
Abstract
New and little known Gryllones insects (Insecta: Gryllones) from the Triassic of Eurasia are described. The genus Yontala Aristov, 2005 from the Lower Triassic locality of Yontala (Induan Stage of the Vologda Region, Russia) is re-described and transferred to the family Sylvabestiidae of the order Cnemidolestida. Sharovala triassica gen. et sp. nov. (Cnemidolestida: Sylvabestiidae) from the Middle Triassic Madygen locality (Ladinian Stage of the Osh region of Kyrgyzstan) is described. New representatives of the order Reculida, Chauliodites sakmaris sp. nov. (family Chaulioditidae) from the Lower Triassic locality of Petropavlovka (Olenekian Stage of the Orenburg Region, Russia) and Batkenopterum kirgizicum gen. et sp. nov. (family Sylvaphlebiidae) from Madygen are described. Changes in Cnemidolestida and Reculida faunas at the boundary of the Paleozoic and Mesozoic are discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Changes in the faunas of some orders of Gryllones insects (Cnemidolestida and Reculida) in the time interval close to the boundary of the Permian and Triassic are discussed in this paper. The interval that extends from the Vyatkian Stage of the Upper Permian to the Ladinian Stage of the Triassic (Madygen locality) is considered. The distribution of the families of the orders discussed is inferred from the studies (Storozhenko, 1998; Rasnitsyn et al., 2013; Aristov, 2014, 2018a, b) for the Permian and from the studies (Storozhenko, 1998; Aristov, 2003, 2020; Aristov and et al., 2011; Bashkuev et al., 2011; Aristov, 2014, 2018a, b) for the Triassic, as well as from the author’s original data.
Cnemidolestida and Reculida faunas from a range of localities (Fig. 1) are considered in this study: the Upper Permian localities include the Vyatkian Belmont (Australia, New South Wales), Aristovo (Russia, Vologda Region), Sokovka (Russia, Vladimir Region), Nedubrovo (Russia, Vologda Region), Babii Kamen (Russia, Kemerovo Region), Tunguska (a group of localities in the Krasnoyarsk Region of Russia), Zalazna (Russia, Kirov Region) and Yaman-Us (Mongolia, South Gobi Aymag: Uranbileg, 2019). Yontala (Russia, Vologda Region) represents the Induan Stage of the Lower Triassic. The localities of Tikhvinskoe (Russia, Yaroslavl Region) and Petropavlovka (Russia, Orenburg Region) date back to the Olenekian age, whereas Hammelburg and others (a group of localities in Germany, Lower Franconia, and Thuringia) and Vosges (a group of localities in France, Alsace, and Lorraine) represent the Anisian Stage, and Madygen (Kyrgyzstan, Osh region) is assigned to the Ladinian layer of the Middle Triassic. The age of the Madygen locality is usually reported as the Ladinian or Carnian of the Middle or Upper Triassic (Shcherbakov, 2008). This concept emerged, because I.A. Dobruskina (1995) did not discriminate between Ladinian and Carnian floras. Our data show that only three of the six families found at Madygen are known from younger sediments (Fig. 1), and therefore we can confirm the assumption of the Ladinian age of the Madygen locality (A.G. Ponomarenko, personal communication, 2018).

Distribution of families of the orders Cnemidolestida and Reculida in the Late Permian–Middle Triassic (the sequence of localities given after Rasnitsyn, 2012).
Three cnemidolestid families (Tillyardembiidae, Psoropteridae, and Sylvabestiidae) and five reculid families (Chaulioditidae, Geinitziidae, Liomopteridae, Sylvaphlebiidae, and Skaliciidae) transitioned from the Middle Permian to the Upper Permian (Fig. 1). Last finds for three of these eight families were reported in the Upper Permian: Tillyardembiidae, Liomopteridae, and Skaliciidae. Endemic families (in this case, Tunguskapteridae) are not considered in calculations of this type, because the first find of such a family is also its last. Thus, 37% of cnemidolestid and reculid families became extinct at the Paleozoic–Mesozoic boundary (29% of all other insect families became extinct at the Paleozoic–Mesozoic boundary: Rasnitsyn et al., 2013).
This extinction appears quite substantial compared to the second Permian extinction of Cnemidolestida and Reculida in the middle of the Kazanian Age of the Middle Permian, as 20% of the families of the group did not transition into the Upper Kazanian substage (Rasnitsyn et al., 2015). Other Permian extinctions were less pronounced. However, the percentage of extinct Vyatkian families may be overestimated. In fact, the “boundary” between the Vyatkian and Anisian assemblages, rather than the Permian–Triassic boundary, is implicated here. The Lower Triassic insect localities known to date are very poor (one cnemidolestid and one reculid specimen is known from each of Yontala and Tikhvinskoe localities, and seven specimens of reculid insects are known from Petropavlovka). The families Tillyardembiidae, Liomopteridae, and Skaliciidae might have survived into the Induan or Olenekian ages, but they have not been discovered yet. Gryllones from the Anisian localities of Europe are insufficiently characterized. The families listed have not been unambiguously identified in Madygen, where a very large collection of insects was assembled. This allows for the assumption (with some degree of certainty) that these species became extinct before the Ladinian age of the Middle Triassic, but not necessarily at the boundary of the Permian and Triassic.
-
SYSTEMATIC PALEONTOLOGY
-
Order Cnemidolestida
-
Infraorder Cnemidolestina
-
Family Sylvabestiidae Aristov, 2000
-
Genus Yontala Aristov, 2005
-
Yontala camura Aristov, 2005
-
Plate 10, fig. 1
Yontala camura: Aristov, 2005, p. 66, pl. 7, fig. 2; text-fig. 1b; Ponomarenko, 2016, p. 76.
Holotype. PIN no. 4891/2, counterpart of an incomplete forewing; Vologda Region, Kichmenga-Gorodets District, right bank of the Yug River 2 km downstream from the mouth of the Yontala River, Yontala locality; Lower Triassic, Induan Stage, Vetlugian Series, Vokhma Formation, Krasnobakovo Subformation (Ponomarenko, 2016).

Explanation of Plate 10
Fig. 1. Yontala camura Aristov, 2005, holotype PIN, no. 4891/2, forewing; Vologda Region, Yontala; Lower Triassic.
Fig. 2. Sharovala triassica sp. nov., holotype PIN, no. 2785/3021, forewing; Kyrgyzstan, Madygen; Middle Triassic.
Fig. 3. Chauliodites sakmaris sp. nov., holotype PIN, no. 5640/4, forewing; Orenburg Region, Petropavlovka; Lower Triassic.
Fig. 4. Batkenopterum kirgizicum sp. nov., holotype PIN, no. 5343/172, forewing; Kyrgyzstan, Madygen; Middle Triassic.
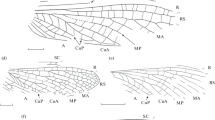
Description (Fig. 2a). Small insects. The anterior margin of the forewing is slightly convex; the costal field at the RS base is twice as wide as the subcostal one. SC is with simple, straight, Y-shaped anterior branches, ends at the border of the distal third of the wing. R is with simple and straight anterior branches. RS begins at the border of the basal third of the wing, fused with MA, RS + MA anastomosis with two or three branches. The interradial field is wide. M5 not close to the first fork of M, which is divided into bifurcated MA and MP at the RS base level, MP starts branching at the base and has four or more endings. CuA is simple, with a slight S-shaped bend. CuA concave anterior to the M5 confluence site and convex posterior to it. The intercubital field is not expanded, CuP is straight. A1 is simple, with an S-shaped bend, A2 is with two or more branches.

Triassic representatives of the orders Cnemidolestida and Reculida, forewings: (a) Yontala camura Aristov, 2005, holotype PIN, no. 4891/2; Vologda Region, Yontala; Induan Stage of the Lower Triassic; (b, c) Sharovala triassica sp. nov., holotype PIN, no. 2785/3021: (b) preserved forewing, (c) reconstruction; Kyrgyzstan, Osh region, Madygen; Ladinian Stage of the Middle Triassic; (d) Chauliodites sakmaris sp. nov., holotype PIN, no. 5640/4, Orenburg Region, Petropavlovka; Olenekian Stage of the Lower Triassic; (e, f) Batkenopterum kirgizicum sp. nov., holotype PIN, no. 5343/172: (e) preserved front wing, (f) reconstruction; Madygen. Scale bar 2 mm in a and d and 3 mm in b, c, e, and f.
Measurements, mm: forewing length approximately 11.
Comparison. The genus Yontala is most similar to Kazanalicula Aristov et Storozhenko, 2011 from the Middle Permian Soyana locality (Kazanian Stage of the Arkhangelsk Region). A wide costal field and the presence of an RS + MA anastomosis distinguishes Yontala from Kazanalicula. Kazanalicula has a narrow costal field and a free RS (Aristov and Storozhenko, 2011).
Remarks. The genus Yontala was tentatively assigned to the family Chaulioditidae at first description (Aristov, 2005). This genus has a simple CuA, which is not known for Chaulioditidae or any of the Reculida, an order that includes this family (Aristov, 2015a). The discovery of Sylvabestiidae representatives with a similar venation pattern (Aristov and Storozhenko, 2011) enabled the attribution of Yontala to this family.
Y. camura exhibits a characteristic feature, even though it is not unique for Cnemidolestida: the CuA is concave anterior to the site of fusion to M5 and convex posterior to it. This is the primary structure of the forewing CuA for insects (Rasnitsyn, 2007). A similar CuA structure is known, for example, for some Cnemidolestidae from the Chunya site (Carboniferous) in the Krasnoyarsk Region, in which CuA is slightly concave or neutral (Aristov, 2013), although special attention was not paid to this feature. The CuA of Permian cnemidolestids is convex anterior to the point of confluence with M5.
Material. Holotype.
-
Genus Sharovala Aristov, gen. nov.
Etymology. In honor of A.G. Sharov and from the Latin ala (wing); feminine.
Type species – Sh. triassica sp. nov.
Diagnosis. Medium-sized insects. Сostal field at base of RS as wide as subcostal. RS starts anterior to middle of wing, starts branching early, and has six endings. MA with smooth inflexion beyond base. CuA starts branching beyond its basal quarter, not divided into CuA1 and CuA2, and ends at the beginning of distal third of wing.
Species composition. Type species.
Comparison. The new genus is most similar to Mezenalicula Aristov et Storozhenko, 2011 from the Middle Permian Soyana locality (Arkhangelsk Region), from which it is distinguished by the narrow costal field and late emergence of RS. The costal field at the base of RS is wider than the subcostal one in Mezenalicula, and RS begins in the basal third of the wing (Aristov and Storozhenko, 2011).
-
Sharovala triassica Aristov, sp. nov.
-
Plate 10, fig. 2
Etymology. From the Triassic.
Holotype. PIN, no. 2785/3021, counterpart of a forewing; Kyrgyzstan, Osh Region, Batken District, Madygen Stow, Madygen locality; Middle Triassic, Ladinian Stage, Madygen Formation.
Description (Figs. 2b and 2c). The anterior margin of the forewing is slightly convex in the basal half, then straight; the wing apex is rounded. SC ends beyond the middle of the wing, curved parallel to its front edge, with simple anterior branches. The interradial field is wide. RS starts branching in its basal third, with backward-facing serration and an additional branching point in one of the branches. M starts branching at the level of the first branching point in CuA, MA simple, MP with three branches. CuA is with a slight S-bend and irregular backward-facing serration, with five branches. CuP is straight. Transverse veins are simple and form two to three rows of cells.
Measurements, mm: anterior wing length approximately 22.
Material. Holotype.
-
Order Reculida
-
Family Chaulioditidae Handlirsch, 1906
-
Genus Chauliodites Heer, 1864
-
Chauliodites sakmaris Aristov, sp. nov.
-
Plate 10, fig. 3
Etymology. From the Sakmara District.
Holotype. PIN, no. 5640/4, part and counterpart of a forewing; Orenburg Region, Sakmara District, Petropavlovka locality; Lower Triassic, Olenekian Stage, Upper Olenekian Substage, Petropavlovka Formation (Shcherbakov et al., 2019).
Description (Fig. 2d). Medium-sized insects. The anterior margin of the forewing is convex. SC ends immediately beyond the middle of the wing, has an S-shaped bend and simple anterior branches. R has an S-shaped bend anterior to the RS base, with simple anterior branches. RS starts immediately beyond the basal quarter of the wing, with three branches, RS branches are curved towards the apex of the wing. M starts branching beyond the RS base anterior to the middle of the wing, MA and MP are bifurcated, CuA1 and CuA2 are simple, CuA1 has a slight S‑shaped bend. The intercubital field is narrow, sharply narrowed posterior to the CuA fork. The wing membrane is colorless.
Measurements, mm: forewing length 11.
Comparison. The new species is most similar to Ch. anisicus Aristov, Grauvogel-Stamm et Marchal-Papier, 2011 from the Anisian Vosges locality (Middle Triassic of France), from which it is distinguished by the curvature of the RS branches towards the apex of the wing. The RS branches in Ch. anisicus are curved towards the front edge of the wing (Aristov et al., 2011).
Material. Holotype.
-
Family Sylvaphlebiidae Martynov, 1940
-
Genus Batkenopterum Aristov, gen. nov.
Etymology. From the Batken District and Greek pteron (wing); neutral.
Type species – B. kirgizicum sp. nov.
Diagnosis. Medium-sized insects. Costal field at the RS base as wide as subcostal. SC reaches almost to the apex of the wing, R with two anterior branches. RS starts anterior to the middle of the wing, with backward-facing serrations. CuA starts branching in its basal quarter, divided into CuA1 and CuA2, ends at the beginning of the distal third of the wing. CuA1 starts branching at its base, with the branches forming a regular posterior-facing serrated pattern at the middle. Anal veins form rows of cells.
Species composition. Type species.
Comparison. The new genus is most similar to Sylviodes from the Permian localities of Chekarda and Soyana (Kungurian Stage of Perm Region and Kazanian Stage of Arkhangelsk Region, respectively), from which it is distinguished by a straight anterior wing margin, narrow costal margin, and very early branching of CuA1. The anterior edge of the wing is convex in Sylviodes, the costal field is wide, and CuA1 starts branching at a certain distance from its base (Aristov, 2004, 2015b).
-
Batkenopterum kirgizicum Aristov, sp. nov.
-
Plate 10, fig. 4
Etymology. From Kyrgyzstan.
Holotype. PIN no. 5343/172, part and counterpart of a forewing; Kyrgyzstan, Madygen locality; Middle Triassic, Ladinian Stage, Madygen Formation.
Description (Figs. 2e and 2f). The anterior margin of the forewing is straight, wing apex pointed. SC with simple anterior branches. The interradial field is broad, RS with four or more branches. M is divided into branches posterior to the first fork in CuA, MA has two branches, MR simple. CuA1 has five branches, CuA2 is with an S-shaped bend. Transverse veins are simple and form a double row of cells in the intercubital field.
Measurements, mm: forewing length approximately 17.
REFERENCES
Aristov, D.S., Revision of the family Tomiidae (Insecta: Grylloblattida), Paleontol. J., 2003, vol. 37, no. 1, pp. 31–38.
Aristov, D.S., The fauna of grylloblattid insects (Grylloblattida) of the Lower Permian locality of Tshekarda, Paleontol. J., 2004, vol. 38, suppl. 2, pp. 80–145.
Aristov, D.S., New grylloblattids (Insecta; Grylloblattida) from the Triassic of Eastern Europe, Eastern Kazakhstan, and Mongolia, Paleontol. J., 2005, vol. 39, no. 2, pp. 173–177.
Aristov, D.S., New and little-known Eoblattida (Insecta) from the Paleozoic of Russia, Paleontol. J., 2013, vol. 47, pp. 272–282.
Aristov, D.S., Classification of the order Cnemidolestida (Insecta; Perlidea) with description of new taxa, Far East. Entomol., 2014, no. 277, pp. 1–46.
Aristov, D.S., Classification of order Eoblattida (Insecta; Blattidea) with description of new taxa, Far East. Entomol., 2015a, no. 301, pp. 1–56.
Aristov, D.S., New gryllones insects (Insecta: Gryllones) from the Permian of Russia, Paleontol. J., 2015b, vol. 49, no. 12, pp. 1310–1333.
Aristov, D.S., New and little known cnemidolestid insects (Insecta: Cnemidolestida) from the Middle Permian–Middle Triassic of Eurasia, Paleontol. J., 2018a, vol. 52, pp. 1381–1390.
Aristov, D.S., Revision of the family Necrophasmatidae (Insecta: Cnemidolestida, Far East. Entomol., 2018b, no. 359, pp. 7–11.
Aristov, D.S., New gryllones insects (Insecta: Gryllones) from the Babii Kamen’ locality (Upper Permian of Russia). 2. Order Reculida and Gryllones ordinis incertis, Paleontol. J., 2020, vol. 54, pp. 132–142.
Aristov, D.S., Grauvogel-Stamm, L., and Marchal-Papier, F., New grylloblattid insects (Insecta: Grylloblattida) from the Grès à Voltzia of the Vosges (Middle Triassic of France), Paleontol. J., 2011, vol. 45, no. 2, pp. 159–166.
Aristov, D.S. and Storozhenko, S.Yu., Review of the Permian family Permulidae nomen novum pro Aliculidae Storozhenko, 1997 (Grylloblattida), ZooKeys, 2011, vol. 130, pp. 111–130.
Bashkuev, A., Sell, J., Aristov, D., et al., Insects from the buntsandstein of Lower Franconia and Thuringia, Paläontol. Z., 2012, vol. 85, no. 3, pp. 175–185.
Dobruskina, I.A., Keuper (Triassic) flora from Middle Asia (Madygen, Southern Fergana), New Mexico Mus. Nat. Hist. Sci. Bull., 1995, vol. 5, pp. 1–49.
Ponomarenko, A.G., New beetles (Insecta, Coleoptera) from the lower Triassic of European Russia, Paleontol. J., 2016, vol. 50, pp. 286–292.
Rasnitsyn, A.P., On the discussion of the wing venation of (Archae)Orthoptera (Insecta), Paleontol. J., 2007, vol. 41, pp. 341–344.
Rasnitsyn, A.P., Aristov, D.S., and Rasnitsyn, D.A., Insects of the Permian and Early Triassic (Urzhumian–Olenekian ages) and the problem of the Permian–Triassic biodiversity crisis, Paleontol. J., 2013, vol. 47, no. 7, pp. 793–823.
Rasnitsyn, A.P., Aristov, D.S., and Rasnitsyn, D.A., Dynamics of insect diversity during the Early and Middle Permian, Paleontol. J., 2015, vol. 49, no. 12, pp. 1282–1309.
Shcherbakov, D.E., Madygen, Triassic Lagerstätte number one, before and after Sharov, Alavesia, 2008, no 2, pp. 113–124.
Shcherbakov, D.E., Bashkuev, A.S., Vasilenko, D.V., et al., Petropavlovka, a new Lower Triassic insect locality, Paleostrat-2019. Godichn. sobranie (nauch. konf.) sektsii paleontol. MOIP i Mosk. otdeleniya Paleontol. ob-va pri RAN. Moskva, 28–30 yanvarya 2019 g. (Paleostrat-2019. Ann. Meeting (Sci. Conf.) Moscow Naturalist Soc. Paleontol. Section and Moscow Section of Paleontol. Soc., Russ. Acad. Sci., Moscow, January 28–30, 2019), Moscow: Paleontol. Inst. Ross. Akad. Nauk, 2019, pp. 68–69.
Storozhenko, S.Yu., Systematika, filogeniya i evolyutsiya grilloblattidovykh nasekomykh (Insecta: Grylloblattida) (Systematics. Phylogeny, and Evolution of Grylloblattid Insects: Insecta: Grylloblattida), Vladivostok: Dal’nauka, 1998.
Uranbileg, L., Permian and Jurassic coal deposits of Southern Mongolia and their paleobotanical characteristics, Problemy paleontologii i stratigrafii Mongolii i sopredel’nykh territorii. Mezhdunar. konf., posvyashchennaya 50-letiyu Sovmestnoi Rossiisko-Mongol’skoi paleontologicheskoi ekspeditsii (SRMPE) 1969–2019 (Problems of Paleontology and Stratigraphy of Mongolia and Adjacent Territories. Int. Conf. Dedicated to the 50th Anniversary of the Joint Russian-Mongolian Paleontological Expedition (JRMPE) 1969–2019), Moscow: Paleontol. Inst. Ross. Akad. Nauk, 2019, pp. 47–50.
ACKNOWLEDGMENTS
The author is grateful to A.P. Rasnitsyn and A.G. Ponomarenko (Paleontological Institute of the Russian Academy of Sciences) for comments on the manuscript.
Funding
This work was supported by Program 15, “Problems of the Origin of Life and the Formation of the Biosphere”, of the Presidium of the Russian Academy of Sciences and grant no. 18-04-00322 from the Russian Foundation for Basic Research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by S. Semenova
Rights and permissions
About this article

Cite this article
Aristov, D.S. New and Little Known Gryllones Insects (Insecta: Gryllones: Cnemidolestida, Reculida) from the Triassic of Eurasia. Paleontol. J. 54, 524–530 (2020). https://doi.org/10.1134/S0031030120050020
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030120050020