
Abstract
The Plio-Pleistocene site of Kromdraai, covering a chronological range from 1.8 Ma (Kromdraai A Locality) up to older than 2.0 Ma (Kromdraai Member 2), (Cradle of Humankind, Gauteng, South Africa) has been investigating since the first half of the XXth century. These researches have led to the discovery of the type specimen of Paranthropus robustus. Kromdraai is also characterized by an extremely rich bone accumulation (including more than 10000 remains with more than 4800 from the recent field works). Carnivores are highly diverse including Felidae, Hyaenidae, Herpestidae, Viverridae, Mustelidae and Canidae. Based on 27 newly discovered dental and postcranial specimens, a new canid species is described. Canis hewitti sp. nov. is comparable in size to the extant African hunting dog Lycaon pictus but it differs significantly from this species in dental features that are typical of the genus Canis including the m1 with a trigonid of about two-thirds the length of the crown, a metaconid clearly dissociated from the protoconid and a talonid consisting in a hypoconid and an entoconid. Its premolars suggest a certain specialization in meat-cutting (long and thin p4 with a high protoconid backwardly flanked of a well-developed cusp followed by an accessory small denticle). C. hewitti appears as the potential ancestor of the more robust southern African Canis atrox from Kromdraai A (ca. 1.8 Ma).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Quaternary canids include numerous genera and species of small-to-large-sized animals (Tedford and Qiu 1996; Tedford et al. 2009; Wang and Tedford 2010; Amri et al. 2017; Bartolini Lucenti et al. 2020, 2021). However, canids have attracted less attention than other carnivores (e.g., felids or hyenids). Fossil canids are quite scarce in the African fossil record and their identification is limited. Several studies have nevertheless focused on southern African canids (e.g., Hendey 1973, 1974a, b; Ewer, 1956; Ewer and Singer 1956; Hartstone-Rose et al. 2010; Reynolds 2012) with the identification of 7 or 8 genera (Eucyon Tedford and Qiu 1996; Otocyon Müller 1836 or Prototocyon Pohle 1928; Vulpes Frisch 1775; Canis Linnaeus 1758; Lupulella Hilzheimer 1906; Lycaon Brookes 1827; Nyctereutes Temminck 1838) and 14 small-to-large-sized species.
The phylogenic relationships and the evolutionary trends of the southern African canids are poorly known. Some species are represented by only few remains from a single site: Vulpes pattisoni from Taung (Broom 1948); Vulpes pulcher from Kromdraai A and Swartkrans Mb.1 (Broom 1939; Ewer 1956); Canis antiquus from Minnaar’s Cave (Broom 1939; Ewer 1956); Lycaon sekowei from Cooper’s D and Gladysvale (Hartstone-Rose et al. 2010); Canis atrox from Kromdraai A (Ewer 1956). Other taxa are still debated: Lupulella adusta from Makapansgat Member 3 (Ewer 1957); Prototocyon from Swartkrans Member 2 (Hendey 1974b).
The recently discovered and excavated Member 2 from the Plio-Pleistocene site of Kromdraai (Cradle of Humankind) has yet provided more than a hundred specimens of small- to large-sized canids to-date (Fourvel et al. 2016, 2018; Braga et al. 2017). This canid sample includes at least three genera of small-sized species: Vulpes (Vulpes chama), Prototocyon (Prototocyon recki) and Lupulella (Lupulella mesomelas). Among the sample likely related to the canids, 27 specimens are significantly different from both Lupulella mesomelas ssp. and the larger Canis atrox or Lycaon sekowei. Consequently, here, we use these 27 specimens to describe a new species Canis hewitti sp. nov.
The hominin-bearing site of Kromdraai
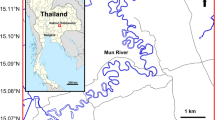
Among the “Cradle of Humankind” (Gauteng, South Africa), Kromdraai appears as a key site for our knowledge of the palaeoenvironmental conditions prevailing during the Pleistocene. It is an unroofed dolomite cave partially shaped by the erosional surface and filled with fossil-bearing deposits, situated approximately 2 km east of Sterkfontein Caves, on the southern side of the Blaauwbank stream (Fig. 1). It yielded the type specimen of Paranthropus robustus, TM 1517 (Broom 1938a, b), and more than 50 other individuals attributed to either Homo or Paranthropus (Braga et al. 2017). Over 4800 faunal remains, which derive from Member 2, have been found during the various field seasons from the new excavations initiated in 2014. The carnivores are composed of 459 specimens (including coprolithes) belonging to 6 families, 22 genera and possibly up to 24 species (Fourvel et al. 2016, 2018; Fourvel 2018). Among the carnivores, the presence of Dinofelis barlowi (which occurs in this region between 3.5 and 2 Ma, Werdelin and Lewis 2001), of Prototocyon recki (which occurs in East Africa at Olduvai (c. 2 Ma) and Laetoli (c. 3.5 Ma), Petter 1964, 1973 but see Werdelin and Dehghani 2011), of Chasmaporthetes silberbergi (which has been identified only in Sterkfontein Members 2 and 4, Werdelin and Peigné 2010) within Kromdraai Member 2 is consistent with an age of or slightly in excess of ca. 2.0 Ma. Among the bovids, the co-occurrence of Connochaetes taurinus prognu and cf. Connochaetes gentryi is noteworthy, because it is reminiscent of the late Pliocene and early Pleistocene of Kenya, as well as the lower levels of Olduvai (Fourvel et al. 2016). Moreover, the presence of a Makapania-like caprine could relate Kromdraai Member 2 to a Plio-Pleistocene transition/early Pleistocene age, since it is abundant at Makapan Limeworks Members 3 and 4 as well as in the basal levels of Olduvai (Gentry and Gentry 1978; Gentry 2010; Fourvel et al. 2016). Accordingly, when compared to other faunal assemblages from the Plio-Pleistocene of Africa, the faunal association from Kromdraai Member 2 suggests an age in excess of ca.2.0 Ma.

Location of Kromdraai in the Cradle of Humankind (Gauteng, South Africa). Map realized by B. Chadelle
Among the carnivores, 110 specimens belong to the canids. The canid sample is referred to the cape fox Vulpes chama (NISP = 7), to the extinct bat-eared fox Prototocyon recki (NISP = 4), to the black-backed jackal Lupulella mesomelas (NISP = 60), to some unidentified canid remains (NISP = 12) as well as the 27 dental and postcranial remains belonging to the newly described species Canis hewitti sp. nov.
Sample and methods
The 27 specimens described here are curated at the Evolutionary Studies Institute of the University of Witwatersrand in Johannesburg.
For comparative purposes, we use morphological and metrical data from a large sample of skulls and mandibles representing the black-backed jackal Lupulella mesomelas (Schreber, 1775) (20 individuals), the side-striped jackal Lupulella adusta (Sundevall, 1847) (6 individuals), as well as the African hunting dog Lycaon pictus (Temminck, 1820) (28 individuals) (all specimens curated at the Ditsong National Museum in Pretoria, the Evolutionary Studies Institute of the University of Witwatersrand in Johannesburg as well as the National Museums of Kenya in Nairobi) as well as the African golden wolf Canis lupaster Hemprich and Ehrenberg, 1832 (Geraads pers. com.) and the Ethiopian wolf Canis simensis Rüppell, 1840 (Rook et al. 1996). The comparative fossil sample includes southern African specimens belonging to L. mesomelas from Coopers D (Fourvel, unpublished), L. mesomelas pappos from Kromdraai A (Ewer 1956), and Canis atrox from Kromdraai A (Ewer 1956) that we personally re-examined, as well as northern African specimens of Lupulella paralius from Ahl al Oughlam and Lupulella mohibi from north-western Africa (Geraads 2011).
In order to compare Canis hewitti sp. nov. with the other species of Canis, Lupulella and Lycaon, a principal component analysis was performed using trigonid and talonid lengths and width of the m1. The individual variables were centered and reduced prior to analysis. These three lengths are the ones that could be measured on the most specimens of Canis hewitti sp. nov. The analysis is indeed constrained by the available data: due to the conservation of the material, not all dental lengths could be measured for all 27 specimens.
The measurement protocol follows Von den Driesch (1976) and are expressed in millimeters. The various graphical projections and statistical analyses have been performed with R v.4.1.1 (R Core Team 2021) and associated packages dimensio v.0.2.2 (Frerebeau 2021), ggplot2 v.3.3.5 (Wickham 2016), ggrepel v.0.9.1 (Slowikowski 2021) and patchwork v.1.1.1 (Lin Pedersen 2020).
Abbreviations list
Anatomical abbreviations
Ant. c., anterior cusp; Acc. c., accessory cusp; BPC, breadth of the coronoid process; B, breadth; Bas., basin; Bd, breadth of distal end; Bp, breadth of proximal end; Cing., cingulum; DPA, depth of the Processus anconaeus; Entoc., entoconid (lower tooth); GB, greatest breadth; Hypoc., hypoconid (lower tooth)/hypocone (upper tooth); L, length; Ld, length of distal end; Lp, length of proximal end; Metac., metaconid (lower tooth)/metacone (upper tooth); Parac., paraconid (lower tooth)/paracone (upper tooth); Post. c., posterior cusp; Protoc., protoconid (lower tooth)/protocone (upper tooth); SD, smallest breadth of the diaphysis; Tub. int., tuberculum intermedium.
Systematic paleontology
Order Carnivora Bowdich, 1821.
Family Canidae Fisher, 1817.
Sub-family Caninae Fisher, 1817.
Genus Canis Linnaeus, 1758.
Canis hewitti sp. nov.
Etymology
In honor of Jack Hewitt (1946–2020) who taught one of us (JBF) almost everything he knew about South Africa, for his kindness and his deep friendship throughout the stay of JBF in South Africa.
Diagnosis
Canis hewitti sp. nov. is a large-sized canid with a full dentition composed of 3/3I, 1/1C, 4/4P, 2/3 M. Mandible has deep and robust mandibular corpus. Dental arch is slightly curved. Lower premolars (p2 to p4) are meat-cutting specialized teeth with high and thin crown. The p2 and p3 are high crowned with a mesial accessory cuspid flanking the protoconid. The p4 has a long, thin crown with a high and central protoconid, is flanked posteriorly by a high and well-developed cuspid followed by an accessory small denticle overhanging a thick cingulum. The m1 has a trigonid of about two-thirds the length of the crown. The metaconid is clearly dissociated from the protoconid and inclined posteriorly. The talonid consists in a hypoconid about twice the size of the entoconid, while the latter cuspid is slightly oriented backwards and connected to the metaconid by a low tuberculum intermedium. The hypoconulid is almost absent. The P2 and P3 have a high and central protocone, flanked posteriorly by an accessory cusp. This accessory cusp is reduced to a small tooth ridge on P2, whereas on P3 this cusp is a distinct denticle but lower than the cingulum. The M1 has a paracone almost twice the length of the metacone and significantly higher. The protocone is a long dental ridge forming the lingual margin of the M1.
Differential diagnosis
Canis hewitti is significantly larger than Vulpes spp. and Nyctereutes spp., smaller than the extinct large-sized Canis atrox and Canis africanus and similar in size to the extant Lycaon pictus. M1s in C. hewitti differ from C. atrox and C. africanus in the reduced metaconule and absence of protoconule in the two latter. While the tuberculum intermedium, observed in C. hewitti, is absent in C. atrox. C. hewitti is also larger than both the modern L. adusta and L. mesomelas as well as the fossil relatives L. paralius and L. mohibi from North Africa. C. hewitti differs from the large jackal L. mesomelas pappos in its larger overall size, as well as in the morphology of the p4 and the presence on the m1 of L. mesomelas pappos of a developed ectostylid that strongly accentuates the angle formed by the cristid obliqua. C. hewitti can be distinguished from these small canids by a much longer jugal series and the prominence of the cutting parts (premolar and trigonid) suggesting some specialisation in meat cutting. C. hewitti shares with the genus Lycaon the anterior and posterior denticles bordering the protoconid of the lower premolars and the p4 in particular. However, C. hewitti differs from Lycaon in the morphology of the lower carnassial talonid, which is composed a central hypoconid in the latter genus while there are an entoconid and an hypoconid in C. hewitti.
Holotype
Portion of left mandible including p1, p2, p3, p4, m1, m2, m3 (KW 9571a); fragment of left maxilla including M1 and M2 (KW 9571b); fragment of right maxilla including M1 and M2 (KW 9571c); left P3 (KW 9571d); right P3 (KW 9571e); left P2 (KW 9571f); right P2 (KW 9571g); right I3 (KW 9571h).
Referred material
Left proximal ulna (KW 6411); radius diaphysis (KW 6895/6889); portion of right mandible including p3 and p4 (KW 6970); left navicular (KW 7415); right humerus cylinder (KW 7433); radius diaphysis (KW 7520); distal metapodial (KW 7609); right M1 (KW 7940); proximal fifth metatarsal (KW 8276); portion of left mandible including p1 and m1 (KW 8743); portion of left mandible including m1 (KW 9071a); left I3 (KW 9071b); right premaxilla with C (KW 9071c); left proximal fifth metatarsal (KW 9150); right upper canine (KW 9570); right m1 (KW 10109); right m3 (KW 10172); right upper canine (KW 10189c); left m1 (KW 10728).
Type Locality
Kromdraai Member 2, Gauteng, South Africa (Plio-Pleistocene Transition-Early Pleistocene).
Description
The sample includes 27 specimens (19 cranial and dental remains and 8 postcranial elements) belonging to 6 adult individuals. KW 9571, KW 6970 and KW 9071 belong to older animals according to their heavy wear stage. Some of the specimens described below have been recently referred to cf. Lycaon (Fourvel et al. 2016) because of the lack of comparative datasets and dental material but they are now referred to Canis hewitti. All the metrics obtained from the Canis hewitti sample are summarized in Tables 1 and 2.
Lower dentition
The lower jaw and teeth are represented by 7 specimens.
KW 9571a is designated as the holotype of C. hewitti because of its completeness (Fig. 2). KW 9571a is an almost complete left mandibular corpus (from the canine alveolus, which is partially preserved, up to the condyle). The corpus is deep and robust with a convex ventral profile reaching its lowest point below the m2. The masseteric fossa is deep and starts just behind the m3. There are two mental foramina both at mid-branch height. The first one is large and just below the p1, while the second foramen is half the size of the first and located below the anterior root of p3. In occlusal view, the dental arch is slightly curved. The p2 is a long, thin tooth (about 2.5 times longer than wide), with a constant width throughout the crown. The main cuspid is located almost above the anterior root. At the rear, centred above the posterior root, there is a small denticle clearly separated from the protoconid. The morphology of p3 is comparable to that of p2, but its imperfect preservation on KW9571a limits observations. Nevertheless, an accessory cuspid is present. It rises on the posterior third of the crown and overhangs the cingulum. The p4 is a high- and sharp-crowned tooth. The protoconid, which is almost a third of the total length of the crown, is centrally positioned on the crown and inclined posteriorly. It is anteriorly flanked by a small denticle almost reduced to a simple tooth ridge but clearly dissociated from the protoconid. Another high and well-developed accessory cuspid rises just behind the protoconid. This cuspid, half as high as the protoconid, occupies a quarter of the length of the crown and displays a rounded profile in both lingual and buccal views. The well-marked posterior cingulum is overhung by a smaller accessory cuspid. This accessory cuspid, as high as the anterior cuspid, is distinct from the cingulum and the posterior cuspid. Even though the m1 on KW 9571a is worn, the typical Canis features remain visible. The trigonid is almost two-thirds of the total length of the m1 crown. The paraconid and the protoconid are similar in length. There is a small metaconid flanking posteriorly the protoconid. Even though it is heavily worn, this metaconid looks independent from the protoconid, although it is partially fused at its basis with the later. The talonid bears two cuspids with a large hypoconid and a smaller entoconid. The heavy wear stage limits the talonid description in KW 9571a. The presence of an m2 and m3 and their morphological features are also typical of the genus Canis. The m2 is triangular with two main anterior cuspids (the protoconid and the metaconid) and two smaller (the hypoconid and the entoconid) posteriorly located. The protoconid is almost twice the size of the metaconid. The talonid is narrower than the trigonid. The wear stage is too advanced to provide any more details of the talonid features. However, these features are similar to the m2 in Canis. The m3 on the KW 9571a is partially broken but looks rounded.

Canis hewitti sp. nov. type specimen KW 9571a. Photographies and 3D model captures. From up to down: lingual, buccal and occlusal views. p4 and m1 + m2 detailed views and cusps location
Two other mandibular portions include the lower carnassials m1 (KW8743 and KW9071a) (Fig. 3). The corpus of KW8743 is partially preserved and shows similar features as observed for the type specimen KW9571a. It is deep, particularly behind the m2 and m3. The masseteric fossa is deep and starts behind m3. The same curvature of the dental arch can be seen in occlusal view as in KW9571a. Some parts of the paraconid and the protoconid are missing in KW9071a while the m1 in KW8743 is entirely preserved. Their m1s are similar to those of the holotype. The trigonid in both KW8743 and KW9071a is about two-thirds of the total length of the m1. The protoconid is slightly longer and higher than the paraconid. On the m1 of both specimens, a well-developed metaconid flanks lingually the posterior margin of the protoconid. The talonid is worn on both specimens, but the wear stage degree is less advanced than in KW9571a allowing a more detailed description. It is composed of a large hypoconid and a smaller, posteriorly positioned, entoconid in both KW8743 and KW9071a. Even though the entoconid and the hypoconid are worn on both specimens we notice that they are clearly individualized but connected by a small dental ridge. Moreover, there is a small well-individualized tuberculum intermedium located between the metaconid and the entoconid. The hypoconulid is almost absent in both specimens.

Canis hewitti sp. nov. mandibular remains including carnassial. Left mandibular portion KW 8743, from up to down: lingual, buccal and occlusal views and m1 detailed views and cups location. Left mandibular portion KW 9071a, from left to right lingual, buccal and occlusal views and m1 detailed views and cusps location
The isolated right and left m1s (KW10109 and KW10728) are unworn teeth (Fig. 4). The trigonid is composed of a paraconid about a fourth of the crown length, and of a protoconid longer (about a third of the length of the crown) and significantly higher (about a third higher) than the paraconid. The metaconid is almost as high as the paraconid and flanks the posterior margin of the protoconid. The talonid is composed of a well-developed hypoconid located along the buccal surface and of an entoconid significantly smaller than the hypoconid and posteriorly located on the lingual surface.

Canis hewitti sp. nov. lower m1s KW 10,109 (left) and KW 10,728 (right). From up to down: lingual, buccal and occlusal views and cusps location
KW6970 is a small portion of horizontal corpus of a right mandible preserving the p3 and the p4 and the alveolus of the p2, including both anterior and posterior roots (Fig. 5). It belongs to an old individual and is partially covered by breccia. It was firstly assigned to cf. Lycaon (Fourvel et al. 2016), but is similar to KW9571a. The p3 and the p4 are long and narrow and are composed of a prominent main cuspid covering about half of the length of the crown. The main cuspid of p3 is located towards the front of the tooth overhanging the anterior root. There is a posterior cusp distinct from the protoconid and overhanging a developed cingulum as observed on p3 of KW9571a. The protoconid occupies almost one third of the length of the crown and a small anterior denticle is visible. The protoconid is flanked by a large and rounded-profile posterior cuspid of about a quarter of the length of the tooth. At the back of the latter is the small accessory denticle almost as low as the cingulum.

Canis hewitti sp. nov. mandibular remain KW 6970. Right mandibular portion including p3 and p4. From up to down: lingual, buccal and occlusal views
KW 10172 is an isolated, unworn right m3. The tooth is small and round-shaped (similar to the m3 in KW 9571a). The crown base is entirely surrounded by a well-marked cingulum. The tooth bears two main cuspids that are partially fused (presence of a dental ridge which links the cuspids).
Upper dentition
The upper dentition is represented by 12 specimens. Table 1 summarizes the metrics of the upper dentition.
KW 9071b is a left I3 with an elongated and slightly curved root. KW 9071b is morphologically similar to the jackal L. mesomelas but significantly larger. The same comments apply to the right I3 KW 9571 h which belongs to the type specimen. There is no difference with the I3 of the hunting dog in both size and morphology.
KW 9570, KW9071c and KW 10189c are three right isolated upper canines. KW 9071c is still included in the premaxilla. In KW 10189c the root is fully-developed and the crown remains unworn indicating a fully-growth individual (young adult). The crown is as long as the root (about 23 mm each) and its base has an oval-shaped section (transversally compressed). The root is oval-shaped and slightly inflated at the middle of its length.
KW 9571f and KW 9571 g are respectively the left and right P2s of the type specimen (Fig. 6). The P2s are characterized by their anteriorly located, high protocone. There is a small accessory cusp located just behind the protocone, reduced to a vestigial crest and not clearly individualized.

Canis hewitti sp. nov. upper P2s (left) and P3s (right) from KW 9571 type specimen. From left to right: lingual and buccal views and cusps location
The left and right P3s (KW 9571d and KW 9571e) have long (about 2.5 times longer than wide) and thin crowns. The protocone is located anteriorly. At its posterior base, it is flanked by a distinct, low accessory cusp. The posterior cingulum is strong.
Two fragments of maxilla (the left one KW 9571b and the right one KW 9571c) preserve their M1 and M2 (Fig. 7) while KW 7940 is an incomplete right M1 preserving a partial metacone as well as all the lingual part (including the protoconule, the protocone, the metaconule and the hypocone). These M1s are Canis-like molars and differ significantly from L. pictus in which the M1 is characterized by paracone and metacone similar in size, a strong protocone, an almost absent metaconule and the hypocone reduced to a dental ridge. The paracone of the M1s is almost twice the length of the metacone, and significantly higher. The protocone is a long dental ridge forming the lingual margin of the M1. The M2s in C. hewitti are morphologically similar to the M2s in L. mesomelas but larger than the latter.

Canis hewitti sp. nov. maxilla remains from KW 9571 type specimen. Occlusal views of M1 and M2s and cusps location
Postcranial Elements
Eight postcranial remains are tentatively referred to C. hewitti. All the measurements are provided in Table 2. Some of the postcranial elements (ulna KW 6411, radius KW 6895 + 6889, radius KW 7520, humerus KW 7433, navicular KW 7415, distal metapodial KW 7609) have been previously described and assigned to cf. Lycaon (Fourvel et al. 2016). In the light of the newly discovered specimens (dental remains), we now consider these remains as part of the new species C. hewitti. They are all larger than Lupulella spp. but similar in size to the modern L. pictus. KW 8276 and KW 9150 are two proximal ends of fifth metatarsal. These metatarsals have no particular features differing from the other canid species. However, they are larger than the black-backed jackal and almost identical to the extant hunting dog.
Comparisons and discussion
Comparisons with Vulpes, Nyctereutes, Lycaon-Xenocyon and Eucyon species
Canis hewitti differs from Vulpes by its significantly larger size and distinct morphological features. The allocation to Nyctereutes spp. is also excluded because of the absence of sub-angular lobe on the mandible. Similarly, the allocation to Eucyon spp. is excluded. The second posterior cusplet in the p4, as observed in Kromdraai specimen, is considered as a typical feature of Eucyon (Tedford and Qiu 1996; Spassov and Rook 2006; Werdelin et al. 2014), however the p4 from Kromdraai differs from Eucyon in its larger size as well as the presence of an anterior cusplet which is absent in the latter genus. Even though C. hewitti is similar in size to L. pictus (Fig. 8), any close relationship with the latter can be excluded. The canines are similar in size to those of Lycaon. However, they are morphologically more Canis-like than Lycaon-like. The premolars in Lycaon species (including L. sekowei, L. magnus and the extant L. pictus) are characterized by their high crown and sharpness. The p4 from Kromdraai is almost similar to those of Lycaon and Xenocyon where the p4s bear both anterior and posterior accessory cuspids (see Hartstone-Rose et al. 2010), but differs from them in the protoconid which is not as high and sharp. Moreover, the lower molars (m1, m2) and the upper molars (M1, M2) differ significantly from those of Lycaon. The lower carnassial and upper molars of Lycaon are characterized by their simplification (talonid in m1 reduced to a central hypoconid; absence of entoconule and reduced hypocone in M1). In contrast, the developed talonid of the m1s as well as the M1s and M2s of Kromdraai, sharing similarities with the genus Canis. Similarly, C. hewitti differs from the large-sized L. sekowei from Cooper’s D and Gladysvale which is significantly larger (Fig. 8a). The P2 and the P3 of the type specimen of L. sekowei from Cooper’s D (CD 8280/8281/8285 figured in Hartstone-Rose et al. 2010:300, Fig. 1) are similar to L. pictus. This accessory denticle of the P2 in C. hewitti flanking the protocone, reduced and integrated into the dental ridge, is well-developed and higher in the P2 of both L. sekowei and L. pictus. Even though the M1 in L. sekowei is much more Canis-like, the protocone is more reduced than in the genus Canis (see Hartstone-Rose et al. 2010:302, Fig. 3). The m3 in C. hewitti differs from the m3 in the hunting dog whose m3 bears a single central cuspid.

Stripcharts of C. hewitti and other African canids tooth lengths. a Length of the p4s. b Length of the m1s. c Cumulative length of the premolars (p1 to p4) and trigonid. d Cumulative length of jugal teeth (premolars and molars). The black dot and line represent the mean and standard deviation for each group
Comparisons with Canis and Lupulella species
The various species of the genus Lupulella are generally smaller than C. hewitti (Figs. 8, 9). Even though the p4 of C. hewitti is metrically close to the p4 of C. lupaster and L. mesomelas pappos (Fig. 8a), the jugal series of C. hewitti is significantly longer than in the small species of the genera Canis and Lupulella (Fig. 8d). In addition, the sectorial parts of the dentition (premolars and m1 trigonid) are more developed in C. hewitti, bringing C. hewitti metrically closer to C. simensis and L. pictus (Fig. 8c). Furthermore, C. hewitti differs morphologically from the small Canis-Lupulella. In L. adusta, the lower premolars are spaced by important diastemata that are absent in C. hewitti. While the lower premolars of C. hewitti bear accessory cuspids anteriorly and posteriorly (two posterior cuspids in the case of p4) to the protoconid, the p3 of L. adusta has a single protoconid and the p4 has a triangular protoconid flanked posteriorly by a single accessory cusp. Similarly, the P2 and P3 of L. adusta are composed of a single protocone, whereas this is flanked by an accessory cusp in C. hewitti. The same observations about the upper (P2 and P3) and lower premolars (p3 and p4) can be made on the fossil forms from Southern Africa (L. mesomelas pappos) and North Africa (L. mohibi and L. paralius). Even though L. mesomelas pappos described in Kromdraai A (Ewer 1956) is considered as a large black-backed jackal, this sub-species is smaller than the specimens that we refer to C. hewitti (Fig. 8). For example, the length of the sectorial parts (premolars and trigonid) reaches 58 mm in C. hewitti against 40 mm in L. mesomelas pappos and the length of the jugal series (premolars and molars) reaches 79 mm in C. hewitti against 61 mm in L. mesomelas pappos. The m1 of L. mesomelas pappos differs from that of C. hewitti by the presence of a developed ectostylid attached to the hypoconid reducing the importance of the cristid obliqua. The p4 of L. mesomelas pappos does not have an anterior denticle as seen in C. hewitti. L. mohibi from the Middle Pleistocene of North Africa is, like L. paralius from Ahl al Oughlam, a small form distinct from C. hewitti (Fig. 8). The most significant morphological difference is the upper molars of L. mohibi, which are trapezoidal in shape, as long as they are wide, and have an extremely developed hypocone (for more details see Geraads 2011). C. hewitti differs from the various extant and extinct Canis as well as Lupulella and Lycaon based on its overall morphology and size (Fig. 9). Among the modern species, C. hewitti is compared to C. lupaster, the African golden wolf, and C. simensis, the Ethiopian wolf. C. lupaster and C. hewitti are morphologically close, although the teeth of C. hewitti are greater overall. Both species have accessory cuspids posterior to the protoconid on the p3 and p4. The diastema separating the p2 and p3 remains larger in C. lupaster than in C. hewitti. The m1s are comparable morphologically, however, the carnassial of C. hewitti is longer (mean L = 22.6 mm) and thicker (mean B = 8.6 mm) than that of C. lupaster (mean L = 19.3 mm; mean B = 7.4 mm). M1 and M2 of C. lupaster differ from that of C. hewitti in the presence of a more prominent cingulum, particularly on the distal border of M2. C. hewitti differs from C. simensis in both general morphology and proportions. The mandible of C. simensis is long and slender, the premolars being separated from each other by long diastemas. The length of the premolars (p1 to p4) is almost 50 mm (compared to 44 mm in C. hewitti). Conversely, tooth comparison shows that they are shorter in C. simensis than in C. hewitti. C. hewitti remains larger than C. brevirostris (a species similar in size of the extant L. mesomelas with shorter premolars and rectangular upper molars, for more details see Ewer 1956) from Sterkfontein in South Africa or C. lupaster (syn. Canis aureus sensu Geraads 2011) from Pleistocene North Africa (Fig. 8). C. hewitti differs from the large-sized C. africanus from the early Pleistocene site of Aïn Hanech, Algeria (Geraads 2011). The latter is significantly larger (M1 breadth = 18.6 mm; M1 length = 15.7 mm) than C. hewitti (M1 breadth = 17.0 mm, 17.1 mm; M1 length = 13.6 mm, 13.4 mm). The metaconule is reduced in C. africanus whereas it is developed in C. hewitti.

Principal component analysis of C. hewitti and other African canids and licaons tooth lengths. a Individuals factor map and associated convex hull. b Variables factor map
Among the canids, C. atrox is the unique large-sized Canis found so far in the “Cradle of Humankind”, at Kromdraai A (Ewer 1956; Turner 1986) and is significantly larger than C. hewitti. The m1 (KA 1288) of C. atrox is similar to the m1s of C. hewitti. The trigonid occupies nearly 62% of the length of the m1 in both C. atrox and C. hewitti. The talonid consists of a large hypoconid and an entoconid almost half as large. The hypoconulid is reduced to a simple ridge in C. atrox. C. atrox differs from C. hewitti by the absence of the tuberculum intermedium. The M1 (KA 1556) of C. atrox differs from the M1 of C. hewitti. The paracone is larger than the metacone in C. atrox. The metaconule is reduced and the protoconule is absent in C. atrox while the hypocone is as developed and high as the protocone. Even though some features (such as the M1) differs between C. hewitti and C. atrox, the Kromdraai species shares much more affinities with C. atrox than with any other canid species from the Plio-Pleistocene of South Africa. Accordingly, we assume that C. hewitti is the most likely ancestor of C. atrox.
Palaeoecological Implications
The various morphological features and metrics presented here are useful data to discuss the palaeoecology of C. hewitti. The lower carnassials (KW 8743, KW 9071a, KW 9571a, KW 10109 and KW 10728) are similar in size to the extant hunting dog (Fig. 8b). The body mass estimation [Log mean body mass (kg) = 3.42 Log m1 − 3.33; for more details see Thackeray and Keiser 1992] is close to that of the extant Lycaon. C. hewitti is a large-sized canid with a weight ranging from 17.7 kg to 22.7 kg (mean 19.8 kg ± 1.8 kg). In the extant hunting dog L. pictus, the males and the females have almost the same weight ranging from 18 kg up to 36 kg. While in the black-backed jackal, the weight ranges from 5.9 to 8.0 kg (mean 7.7 kg ± 0.8 kg) in the females and from 6.4 to 11.4 kg (mean 8.4kg ± 0.8 kg) in the males (Walton and Joly 2003). The extinct C. atrox from Kromdraai A is heavier with a weight estimated to 41.7 kg.
The dentition in C. hewitti has mixed features between the specialized canids (e.g., Lycaon) and the ubiquitous forms (e.g., Canis). In C. hewitti, the premolars are high and sharp with well-developed accessory cusps. This morphology of the premolar is typical of meat-cutting teeth like in wild dogs. In addition, the sectorial part of the cheek-teeth (premolars and trigonid) of C. hewitti is significantly longer than observed in the various fossil forms of Canis-Lupulella and comparable in size to Lycaon (Fig. 8c). This observation reinforces the idea of a certain specialization in meat consumption. Accordingly, C. hewitti should be a hunter and a primary predator differing from the smaller and less-specialized jackal-sized species. However, both upper and lower molars in C. hewitti are well-developed and Canis-like which suggest that C. hewitti had also high crushing capacities.
Conclusion
The phylogeny of the large-sized Canis and Lycaon is still debated. Canis hewitti sp. nov. is a canid almost similar in size to the extant L. pictus but differing morphologically from these highly specialized canids. Canis hewitti sp. nov. is significantly larger than the jackal-sized species and smaller than the large Canis atrox. The large-sized canid (C. atrox and L. sekowei) are relatively confined in time and do not appear until 2 Ma. L. mesomelas has been also identified at Kromdraai Member 2 and, apart from Canis hewitti sp. nov., no large-sized canids have been identified either at Kromdraai Member 2 or in deposits of comparable age. In many ways, Canis hewitti sp. nov. appears reminiscent of Canis atrox of a smaller size. Even though a more detailed study has to be undertaken, it is nevertheless true that Canis hewitti sp. nov. appears to be the most probable ancestor of Canis atrox, thus excluding any synonymy between the latter and Lycaon, as is regularly evoked.
This study opens up numerous prospects for palaeoecological researches to be developed on canids. The identification of a new Plio-Pleistocene canid species shows the underestimated richness and diversity of southern African canid guild. In the future, the aim will be to produce systematic studies of the canid guilds to know their full diversity and to specify their palaeoecological involvement in the dynamics of Plio-Pleistocene ecosystems.
References
Amri, L., S. Bartolini Lucenti, M. Saïd Mtimet, N. Karoui-Yaakoub, S. Ros-Montoya, M.P. Esprigares, M.N. BoughdiriBel Haj Ali, and B. Martinez-Navarro. 2017. Canis othmanii sp. nov. (Carnivora, Canidae) from the early Middle Pleistocene site of Wadi Sarrat (Tunisia). Comptes Rendus Palevol 16: 774–783.
Bartolini Lucenti, S., M. Bukhsianidze, B. Martinez-Navarro, and D. Lordkipanidze. 2020. The wolf from Dmanisi and Augmented Reality: review, implications and opportunities. Frontiers in Earth Science (paleontology) 8: 128–140 (131).
Bartolini Lucenti, S., J. Madurell Malapeira, B. Martinez-Navarro, D. Lordkipanidze, and L. Rook. 2021. The early hunting dog from Dmanisi with comments on the social behaviour in Canidae and hominins. Scientific Reports 11: 13501.
Braga, J., J.F. Thackeray, L. Bruxelles, J. Dumoncel, and J.-B. Fourvel. 2017. Stretching the time span of hominin evolution at Kromdraai (Gauteng, South Africa): recent discoveries. Comptes Rendus Palevol 16: 58–70. https://doi.org/10.1016/j.crpv.2016.03.003.
Brookes, J., 1827. In: Griffith, E., Smith, C., Pidgeon, E. (Eds.), The animal kingdom arranged in conformity with its organization by the baron cuvier, with additional descriptions of all the species hitherto named, and of many not before noticed. G. B. Whittaker, London, p. 392.
Broom, R. 1938a. a. The Pleistocene anthropoid apes of South Africa. Nature 142: 377–379.
Broom, R. 1938b. b. Further evidence on the structure of the South African Pleistocene anthropoids. Nature 142: 897–899.
Broom, R. 1939. A preliminary account of the Pleistocene carnivores of the Transvaal caves. Annals of the Transvaal Museum 19: 331–338.
Broom, R. 1948. Some South African Pliocene and Pleistocene mammals. Annals of the Transvaal Museum 21: 1–38.
Ewer, R.F. 1956. The fossil carnivores of the Transvaal caves: Canidae. Proceedings of the Zoological Society London 126: 97–119.
Ewer, R.F. 1957. Some fossil carnivores from the Makapansgat Valley. Palaeontologia Africana 4: 57–68.
Ewer, R.F., and R. Singer. 1956. Fossil Carnivora from Hopefield. Annals of the South African Museum 42: 335–347.
Frisch, J. L. 1775. Das Natur-System der vierfüssigen Thiere. C.F. Günther, Glogau, p 30.
Fourvel, J.-B. 2018. Civettictis braini nov. sp. (Mammalia: Carnivora), a new viverrid from the hominin-bearing site of Kromdraai (Gauteng, South Africa). Comptes Rendus Palevol 17 (6): 366–377. https://doi.org/10.1016/j.crpv.2017.11.005.
Fourvel, J.-B., J. Brink, H. O’Regan, A. Beaudet, and M. Pavia. 2016. Chapter 5, Some preliminary interpretations of the oldest faunal assemblage from Kromdraai. In Kromdraai, a Birthplace of Paranthropus in the Cradle of Humankind, ed. J. Braga and J.F. Thackeray, 71–106. Johannesburg: Sun Media Metro.
Fourvel, J.-B., J.F. Thackeray, J.S. Brink, H. O’Regan, and J. Braga. 2018. Taphonomic interpretations of a new Plio-Pleistocene hominin-bearing assemblage at Kromdraai (Gauteng, South Africa). Quaternary Science Reviews 190: 81–97. https://doi.org/10.1016/j.quascirev.2018.04.018.
Frerebeau, N. 2021. Dimensio: multivariate data Analysis. R package version 0.2.2. Pessac, Université Bordeaux Montaigne. https://doi.org/10.5281/zenodo.4478530.
Gentry, A.W. 2010. Bovidae. In Cenozoic mammals of Africa, ed. L. Werdelin and W.J. Saunders, 741–796. Berkeley: University of California Press.
Gentry, A.W., and A. Gentry. 1978. Fossil Bovidae (Mammalia) of Olduvai Gorge, Tanzania. Part I. Bulletin of the British Museum (natural History) 29: 290–446.
Geraads, D. 2011. A revision of the fossil Canidae (Mammalia) of north-western Africa. Palaeontology 54 (2): 429–446. https://doi.org/10.1111/j.1475-4983.2011.01039.x.
Hartstone-Rose, A., L. Werdelin, D.J. De Ruiter, L.R. Berger, and S.E. Churchill. 2010. The Plio-Pleistocene ancestor of wild dogs, Lycaon sekowei n. sp. Journal of Paleontology 84: 299–308. https://doi.org/10.1666/09-124.1.
Hendey, Q.B. 1973. Carnivore remains from the Kromdraai australopithecine site (Mammalia: Carnivora). Annals of the Transvaal Museum 28 (8): 99–112.
Hendey, Q.B. 1974a. The late Cenozoic Carnivora of the South-Western Cape Province. Annals of the South African Museum 63: 1–369.
Hendey, Q.B. 1974b. New fossil carnivores from the Swartkrans australopithecine site (Mammalia: Carnivora). Annals of the Transvaal Museum 29: 27–47.
Hilzheimer, M. 1906. Die geographische Verbreitung der Afrikanischen Grauschakale. Zoologischer Beobachter 47: 363–373.
Linnaeus, C., 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis. In: Synonymis, Locis, Editio Decima. Tomus I. Laurentius Salvius, Stockholm, reformata.
Lin Pedersen, T. 2020. patchwork: the composer of plots. R package version 1.1.1. https://CRAN.R-project.org/package=patchwork
Müller, J. 1836. Jahresbericht über die Fortschritte der anatomischphysiologischen Wissenschaften im Jahre 1834. Archiv für Anatomie und Physiologie 1835 (1–151): 225–243.
Petter, G. 1964. Origine du genre Otocyon (Canidae africain de la sous-famille des Otocyoninae). Mammalia 28 (2): 330–344.
Petter, G. 1973. Carnivores pléistocènes du ravin d’Olduvai (Tanzanie). In Fossil vertebrates of Africa, vol. 3, ed. L.S.B. Leakey, R.J.G. Savage, and S.C. Coryndon, 44–100. London: Academic press.
Pohle, H. 1828. Die Raubtiere von Oldoway. Wissenschaftliche ergebnisse der Oldowayexpedition herausgegeben von Prof. Dr. Reck 3: 45–54.
R Core Team 2021. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at https://www.R-project.org/
Reynolds, S.C. 2012. Nyctereutes terblanchei: The raccoon dog that never was. South African Journal of Science 108 (1/2): 1–10.
Rook, L., M. Luisa, A. Puccetti, and A. Azzaroli. 1996. Remarks on the skull morphology of the endangered Ethiopian jackal, Canis simensis Rüppel 1838. Rendiconti Lincei 7 (4): 277–302.
Slowikowski, K. 2021. ggrepel: automatically position non-overlapping text labels with ‘ggplot2’. R package version 0.9.1. https://CRAN.R-project.org/package=ggrepel
Spassov, N., and L. Rook. 2006. Eucyon marinae sp. nov. (Mammalia, Carnivora), a new canid species from the Pliocene of Mongolia, with a review of forms referable to the genus. Rivista Italiana Di Paleontologia e Stratigrafia 112 (1): 123–133.
Tedford, R.H., and Z. Qiu. 1996. A new canid genus from the Pliocene of Yushe, Shanxi Province. Vertebrata PalAsiatica 34: 27–40.
Tedford, R.H., X. Wang, and B.E. Taylor. 2009. Phylogenetic systematics of the North American fossil Caninae (Carnivora: Canidae). Bulletin of the American Museum of Natural History 325: 1–218.
Temminck, C.J. 1838. Over de kenniss en de Verbreiding der Zoogdieren van Japan. Tijdschrift voor Natuurlijke Geshiedenis en Physiologie 5: 273–293.
Thackeray, J.F., and J.A. Keiser. 1992. Body mass and Carnassial length in modern and fossil Carnivores. Annals of the Transvaal Museum 35 (24): 337–341.
Turner, A. 1986. Miscellaneous carnivore remains from plio-pleistocene deposits in the Sterkfontein Valley (Mammalia: Carnivora). Annals of the Transvaal Museum 34: 203–226.
Von Den Driesch, A. 1976. A guide to the measurement of animal bones from archaeological sites. Peabody Museum Bulletins 1: 1–137.
Walton, L.R., and D.O. Joly. 2003. Canis mesomelas. Mammalian Species 715: 1–9. https://doi.org/10.1644/0.715.1.
Wang, X., and R.H. Tedford. 2010. Dogs, their fossil relatives and evolutionary history, 232. Columbia: Columbia University Press.
Werdelin, L., and R. Dehghani. 2011. Carnivora. In Paleontology and geology of Laetoli: human evolution in context. Fossil hominins and the associated fauna, vol. 2, ed. T. Harrison, 189–232. New York: Springer Science + Business Media.
Werdelin, L., and M.E. Lewis. 2001. A revision of the genus Dinofelis (Mammalia, Felidae). Zoological Journal of the Linnean Society 132: 147–258.
Werdelin, L., and S. Peigné. 2010. Chapter 32: Carnivora. In Cenozoic mammals of Africa, ed. L. Werdelin and W.J. Saunders, 603–657. Berkeley: University of California Press.
Werdelin, L., M.E. Lewis, and Y. Haile-Selassie. 2014. Mid-Pliocene Carnivora from the Woranso-Mille area, Afar region, Ethiopia. Journal of Mammalian Evolution 21: 331–347.
Wickham, H. 2016. ggplot2: elegant graphics for data analysis. New York: Springer-Verlag.
Acknowledgements
We thank the Cradle Management Authority, the South African Heritage Ressources Agency and R. Lotz for their continuous support. JBF also thanks S. Potze, L. Kgasi, S. Badenhorst, W. Van Zyl (Ditsong Museum, Pretoria) and B. Zipfel (Evolutionary Studies Institute, University of the Witwatersrand, Johannesburg) who gave us access to the modern and fossil samples under their care. JBF also thanks R. Hautefort and B. Lans for their help with the photographs presented in this study as well as B. Chadelle (UMR5608 TRACES Toulouse) for his help in preparing maps. JBF is grateful to J.-P. Brugal (CNRS-UMR7269 LAMPEA) and O. Mwebi (National Museums Kenya, Nairobi) for their help providing data of the extant wild dogs, and D. Geraads (CNRS-UMR7207 CR2P) for the comments and data he provided to us to improve our paper. This research is supported by the French Ministry of Foreign Affairs, the Institut des Déserts et des Steppes (Paris) and the Institut Picot de Lapeyrouse (Toulouse). JBF was supported by the Erasmus Mundus AESOP+ program (Application ES15PD0043). Finally, we would like to thank the reviewers for their insightful comments that improved this manuscript.
Author information
Authors and Affiliations
Contributions
JBF has conceived and designed the analysis, performed the analysis and wrote the paper. NF has performed the analysis and wrote the paper.
Corresponding author
Additional information
Handling Editor: Irina Ruf.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Fourvel, JB., Frerebeau, N. A new canid species (Carnivora: Canidae) from the Plio-Pleistocene hominin-bearing site of Kromdraai (Cradle of Humankind, Gauteng, South Africa). PalZ 97, 163–177 (2023). https://doi.org/10.1007/s12542-022-00628-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-022-00628-4