
Abstract
The family Percrocutidae was erected to include some Miocene and Pliocene hyaena-like carnivoran taxa of Eurasia and Africa, which were included to the Hyaenidae for a long time. The percrocutids were dispersed from Spain to China and in the Eastern Mediterranean region are traced in Serbia, Bulgaria, and Turkey. In Greece, Percrocutidae are scarcely known from the middle and late Miocene of northern Greece and Samos Island. It is, however, quite diverse and the three identified certain species (two of them new) belong to the genera Percrocuta, Dinocrocuta, and Belbus. The members of the family are characterized by the absence of the M2/m2 and the enlarged premolars, mainly P3/p3.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Percrocutidae is an extinct family including some Miocene–Pliocene hyaena-like carnivoran taxa of the Old World, called percrocutoids, which were referred to Hyaenidae for a long time. The genera Percrocuta, Dinocrocuta, Belbus, and Allohyaena are now included to their own family, the Percrocutidae. Their taxonomy has been a matter of debate and is mainly based on the presence of some derived characters as the enlarged premolars and the absence of M2/m2. Based on the deciduous dentition, Schmidt-Kittler (1976) and Chen and Schmidt-Kittler (1983) proposed that the percrocutoids are phylogenetically distinct from Hyaenidae and share a common ancestry with Stenoplesictinae, a distinct carnivoran family. The discovery of a cranium of Dinocrocuta gigantea in China indicated that the morphology of the basicranium and auditory bullae is quite different from that of the true hyaenids. Based on these differences, as well as to those of the deciduous dentition, Werdelin and Solounias (1991) erected the new family Percrocutidae (Qiu et al. 1988).
The family had a great distribution in Eurasia, expanding from Spain to China, and in Africa. In Greece, Percrocutidae were also present, but the known material is scarce; they are traced in Axios Valley, Samos Island, and Chalkidiki Peninsula.
2 Historical Overview
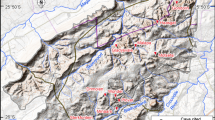
The family Percrocutidae was early recognized in Greece after the discovery of the maxillary fragment of a large-sized hyaenid in Axios Valley (Macedonia, Greece), described as Hyaena salonicae (Andrews 1918). The specimen, after a long taxonomic wandering, was classified as a separate species of the genus Dinocrocuta by Qiu (1987). Quite later, a large-sized hyaenid with percrocutoid features was discovered in the locality Pentalophos 1 of Axios Valley, which was described as Dinocrocuta gigantea (Koufos 1995). Some hyaenid specimens from Samos have also a long taxonomical history, referred under various names. Finally, they are classified in the family Percrocutidae, named Belbus beaumonti by Werdelin and Solounias (1991). More recently, the genus Percrocuta was recognized in the early/middle Miocene locality Antonios, near Thessaloniki (Koufos 2008). In total, three different percrocutoid genera with three different species—two of them are new—have been recognized in the Greek fossil record (Fig. 1 and Appendix).

Map of Greece showing the geographic distribution of the most important localities with percrocutid fossils (the uncertain locality in Samos is not shown). See Appendix for more information. Image exported from Google Earth Pro © 2019, map data from US Dept. of State Geographer, SIO, NOAA, U.S. Navy, NGA, GEBCO, image from Landsat/Copernicus. Scale bar equals 10 km, North faces upward
3 Phylogenetic Relationships
The Percrocutidae as they share some derived features (enlarged premolars, absence of the second molar in both jaws) with the Hyaenidae s.s. were nearly always referred to this family. However, the percrocutids present some derived characters (absence of the metaconid and reduction of the talonid in the m1, reduced protocone in the upper carnassial) distinguishing them from the Hyaenidae s.s. Although the monophyly of the Hyaenidae was early questioned (Thenius 1966; Schmidt-Kittler, 1976) the first clear indications originated from the study of a mandible with deciduous dentition from China (Chen and Schmidt-Kittler 1983). The two authors suggested that: (a) the Percrocutidae are phylogenetically separated from the Hyaenidae s.s., having a parallel but independent evolution, and (b) their feloid dp4 could be an indication that they are phylogenetically far from the Hyaenidae s.s. and share an ancestry with the “stenoplecictines.” The phylogenetic distance from the Hyaenidae s.s. was supported by the different cranial morphology of Dinocrocuta gigantea described by Qiu et al. (1988). Recently, Xiong (2019) scanned and reconstructed the cranium of Dinocrocuta gigantea from China and suggested that the basicranium is like that of the Hyaenidae, questioning the validity of the family Percrocutidae; he suggested that “it could be an early side-branch of the Hyaenidae.” Besides these evidences, the phylogenetic relationships of the Percrocutidae need more fossils and studies to be clarified.
4 Systematic Paleontology
Type Species
Hyaenictis beaumonti Qiu, 1987.
-
★Belbus beaumonti (Qiu, 1987)
Nomenclatural and Taxonomical History
Hyaena sp. in Beaumont 1968 (initial identification); Hyaenictitherium hyaenoides in Howel and Petter 1980 (new combination); Hyaenictis beaumonti in Qiu 1987 (new species); Belbus beaumonti in Werdelin and Solounias 1991 (new genus and combination). The taxonomic history of this material is long and complicated, referred under various generic and specific names (Werdelin and Solounias 1991; Turner et al. 2008).
Type Material
NHMB-SAM-33 (holotype), left mandibular ramus with c–p3, m1, housed in the Naturhistorisches Museum Basel and described and figured by Beaumont (1968:21–26; pl. 1, figs 1, 2) (Fig. 2a).

Selected percrocutids fossils from Greece. (a, b) Belbus beaumonti , right hemimandible, SMNS-SAM-13118, Samos Loc. Unknown, Greece, in (a) lingual and (b) occlusal views, (c) Dinocrocuta gigantea, mandible, LGPUT-PNT-70, Pentalophos 1 (PNT), Axios Valley, Macedonia, Greece, in occlusal view, (d) Dinocrocuta salonicae, right maxillary fragment with P2–P4, NHML-DVT-M.11413 (holotype), Diavata, Axios Valley, Macedonia, Greece, in occlusal view. Scale bar equals 50 mm
Type Locality
Samos Island, Greece, late Miocene, precise locality unknown.
Remarks
This taxon is characterized by the presence of the p1 and m2, the absence of the M2, the large and conical P3 and p3 with strongly curved mesial margin, the weak anterior accessory cusps in the premolars, the large posterior accessory cuspid in the p2 and p3, the large m1 with relatively long talonid and strong metaconid, and the dominance of the hypoconid in the m1 talonid (Beaumont 1968; Werdelin and Solounias 1991).
-
Dinocrocuta Schmidt-Kittler, 1976
Type Species
Hyaena algeriensis Arambourg, 1959.
Remarks
Schmidt-Kittler (1976) erected the subgenus Percrocuta (Dinocrocuta) and defined Hyaena algeriensis Arambourg, 1959 as its type species; however, Qiu et al. (1988) elevated Dinocrocuta to genus level. Besides the various certain species of Dinocrocuta found in Greece (see below), there is a right maxillary fragment with dP2–dP4 (LGPUT-XIR-5) from the late Miocene locality Xirochori 1 of Axios Valley. The deciduous teeth differ from those of Adcrocuta eximia, Percrocuta senyureki, and P. aff. tungurensis. Two large-sized hyaenas are recognized in Axios Valley: D. gigantea and D. salonicae (see below). The deciduous dentition of both species is unknown preventing comparisons and thus the specific determination of the Xirochori specimen is impossible, at the moment; therefore, it is attributed as Dinocrocuta sp. (Koufos 2012).
-
Dinocrocuta gigantea (Schlosser, 1903)
Nomenclatural and Taxonomical History
Hyaena gigantea in Schlosser 1903 (new species); Percrocuta (Dinocrocuta) gigantea in Schmidt-Kittler 1976 (new combination and subgenus); Dinocrocuta gigantea in Qiu 1988 (new combination, upgrade to the genus level). The species has a long taxonomical history and it is referred under different generic or sub-generic names (see Howell and Petter 1985; Werdelin and Solounias 1991).
Type Material
BSPM-1900 XII. 527 (lectotype), right m1, Shansi, China (Schlosser 1903: 35, fig. 6), by the subsequent designation of Kurtén (1957); housed in the Bayerisches Staadtssamlung für Paläontologie und Geologie, München.
Type Locality
Tientsin, Shansi, Tibetfluss (Schlosser 1903), late Miocene.
Distribution
The species is known from the late Miocene of Spain, Bulgaria, Turkey, Georgia, China, Mongolia, and from the Greek locality Pentalophos 1.
Remarks
Although the wide distribution of D. gigantea, the known material is relatively scarce. However, the Chinese material includes some more complete specimens, which allow to see the precise morphology of the taxon. The main morphological characters of the species—modified from Qiu et al. (1988) and Zhang (2005)—are the large-to-very large size, the thick and broad nasal bones, the sharp bending of the cranial roof, the elongated auditory bullae, the short and high cranial proportions, the absence of lingual root in the P3 which is robust and widened mesially, the extremely reduced protocone of the P4, the hypertrophied and strongly oriented lingually p2, the long p4 relative to the p3, and the short m1 relative to the p3. The only known material from Greece is a mandible and two upper canines from Pentalophos 1. The morphology and size of the large-sized hyaenid from the Greek locality Pentalophos 1 (Fig. 2b) allow its attribution to D. gigantea (Koufos 1995).
-
★Dinocrocuta salonicae (Andrews, 1918)
Nomenclatural and Taxonomical History
Hyaena salonicae in Andrews 1918 (new species); ?Crocuta (Percrocuta) salonicae in Kurtén 1957 (new combination); “Hyaena salonicae” in Beaumont 1979 (uncertain attribution); Allohyaena (Dinocrocuta) salonicae in Howell and Petter 1985 (new combination); Dinocrocuta salonicae in Qiu 1987 (new combination).
Holotype
NHML-DVT-M.11413 (holotype), right maxillary fragment with P2–P4, housed in the Natural History Museum of London and described and figured by Andrews (1918: 541) (Fig. 2c, d).
Type Locality
Diavata, Axios Valley, Macedonia, Greece. The locality Diavata could correspond to the fossiliferous level of the locality Pentalophos-1 (PNT) but the absence of information for the exact site of D. salonicae cannot certify it (Andrews 1918; Koufos 1995).
Remarks
Although a certain comparison with the Pentalophos D. gigantea is impossible, as there is different material (maxilla vs. mandible), D. salonicae seems to be smaller than D. gigantea (Koufos 1995). The main characters of the specimen—modified from Pilgrim (1931) and Beaumont (1979)—are the very large size, the large canine, the small P1, the small posterior accessory cusp and the absence of the anterior one in the P2, the absence of anterior and the presence of posterior accessory cusp in the P3, the elongated upper carnassial with extremely strong parastyle, the short protocone, situated behind the mesial margin of the parastyle, and the large M1.
-
Percrocuta Kretzoi, 1938
Type Species
Crocuta carnifex Pilgrim, 1932.
Remarks
An isolated upper carnassial is only known from Greece, found in the early/middle Miocene locality Antonios. This upper carnassial has similarities with P. carnifex and P. abessalomi , but the limited material does not allow a definite identification and thus it is attributed to Percrocuta sp. (Koufos 2008).
References
Andrews CW (1918) Note on some fossil mammals from Salonica and Imbros. Geol Mag 6:540–543
Chen G, Schmidt Kittler N (1983) The deciduous dentition of Percrocuta Kretzoi and the diphyletic origin of the hyaenas (Carnivora, Mammalia). Palaeont Z 57:159–169
de Beaumont G (1968) Une interessante mandibule de Hyaenidae (Carnivora) du Pontien de Samos. Arch Sci Geneve 32:247–250
de Beaumont G (1979) Qu’ est-ce que la “Hyaena salonicae Andrews” (Mammifere, Carnivore). Arch Sci Geneve 32:247–250
Howell FC, Petter G (1985) Comparative observations on some middle and upper Miocene hyaenids genera: Percrocuta Kretzoi, Allohyaena Kretzoi, Adcrocuta Kretzoi (Mammalia, Carnivora, Hyaenidae). Geobios 18:419–476
Koufos GD (1995) The late Miocene percrocutas of Macedonia (Greece). Palaeovertebrata 24:67–84
Koufos GD (2008) Carnivores from the early/middle Miocene locality of Antonios (Chalkidiki, Macedonia, Greece). Geobios 41:365–380
Koufos GD (2012) New material of Carnivora (Mammalia) from the late Miocene of Axios Valley, Macedonia, Greece. C R Palevol 11:49–64
Kurtén B (1957) Percrocuta Kretzoi (Mammalia, Carnivora), a group of Neogene hyenas. Acta Zool Cracoviensia 2(16):375–403
Pilgrim GE (1931) Catalogue of the Pontian Carnivora of Europe in the Department of Geology. The British Museum, Natural History, London
Qiu Z (1987) Die Hyeniden aus dem Ruscinium und Villafranchium Chinas. Münchner Geowiss Abh A 9:1–110
Qiu Z, Xie J, Yan D (1988) Discovery of the skull of Dinocrocuta gigantea. Vertebrata Palasiatica 26:128–138
Schlosser M (1903) Die fossilen Säugetiere Chinas nebst einer Odontographie der recenten Antilopen. Abh Bayer Akad Wiss 22:1–221
Schmidt-Kittler N (1976) Carnivores from the Neogene of Asia Minor. Palaeontographica Abt A 155:1–131
Thenius E (1966) Zur Stammesgeschichte der Hyanen. Z Saugetierkunde 31:293–300
Turner A, Anton M, Werdelin L (2008) Taxonomy and evolutionary patterns in the fossil Hyaenidae of Europe. Geobios 41(5):677–687
Werdelin L, Solounias N (1991) The Hyaenidae: taxonomy, systematics and evolution. Fossils Strata 30:1–106
Xiong W-Y (2019) Basicranial morphology of late Miocene Dinocrocuta gigantea (Carnivora: Hyaenidae) from Fugu, Shaanxi. Vertebrata Palasiatica 57:274–307
Zhang Z (2005) New materials of Dinocrocuta (Percrocutidae, Carnivora) from Lantian, Shaanxi Province, China, and remarks on Chinese Late Miocene biochronology. Geobios 38:685–689
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Koufos, G.D. (2022). The Fossil Record of Percrocutids (Mammalia: Carnivora: Percrocutidae) in Greece. In: Vlachos, E. (eds) Fossil Vertebrates of Greece Vol. 2. Springer, Cham. https://doi.org/10.1007/978-3-030-68442-6_18
Download citation
DOI: https://doi.org/10.1007/978-3-030-68442-6_18
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-68441-9
Online ISBN: 978-3-030-68442-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)