
Abstract
We describe two large predators from the hominoid-bearing Khorat sand pits, Nakhon Ratchasima Province, northeastern Thailand: a new genus of pantherine, Pachypanthera n. gen., represented by partial mandible and maxilla and an indeterminate sabre-toothed cat, represented by a fragment of upper canine. The morphological characters of Pachypanthera n. gen., notably the large and powerful canine, the great robustness of the mandibular body, the very deep fossa for the m. masseter, the zigzag HSB enamel pattern, indicate bone-cracking capacities. The genus is unique among Felidae as it has one of the most powerful and robust mandibles ever found. Moreover, it may be the oldest known pantherine, as other Asian pantherines are dated back to the early Pliocene. The taxa we report here are the only carnivorans known from the late Miocene of Thailand. Although the material is rather scarce, it brings new insights to the evolutionary history of Neogene mammals of Southeast Asia, in a geographic place which is partly “terra incognita.”
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The extant family Felidae comprises about 14 genera (Kitchener et al. 2017) and numerous species adapted to hunting and feeding mainly on the flesh of living animals although they may sometimes crack the bones. Statistical analysis (Werdelin 1983) separate the extant felids in two clusters: large and small cats; the two groups being separated by size but also by their different proportions. When considering the fossil record, Felidae are divided into two groups: the fossil Machairodontinae or sabre-toothed cats and fossil and extant Felinae, most of them having conical upper canines (Werdelin et al. 2010). The fossil record of Felidae dates back to the Oligocene with the small-sized European Proailurus Filhol, 1879. During the Miocene, Felinae are represented by small- and medium-sized species. The large-sized felines only appear during the Pliocene and belong to pantherine (Panthera [lion, tiger, leopard, jaguar, snow leopard] and Neofelis, the clouded leopard), puma-like or cheetah-like forms. Here we describe the first remains of carnivorans from the late Miocene hominoid-bearing Khorat sand pits, in Thailand: a partial hemi-mandible and maxilla of a large-sized feline and an upper canine of a machairondontine. We discuss the ecological role of the new fossils in the past communities of Thailand. These discoveries further document the diversity of large-sized carnivorans in the late Miocene ecosystems of Southeast Asia.
Material and methods
The studied material is a left hemi-mandible (CUF-KR-1), a piece of right maxilla (CUF-KR-2), and a canine fragment (KHT 1) from Khorat sand pits. In the case of CUF-KR-1 and CUF-KR-2, we performed comparisons with fossil or extant felids. Additionally, we used high-resolution X-ray tomography to access and describe the internal structure of the lower jaw (CUF-KR-1). The microtomographic acquisition was carried out using the EasyTom XL Duo (RX-solutions, France) available on the PLATINA platform (IC2MP, Université de Poitiers). A sealed microfocus x-ray source (L12161-07, Hamamatsu Photonics, Japan) was used coupled to a flat panel detector (PaxScan 2520DX, Varian, USA; 1920 × 1536 matrix pixel; pixel pitch of 127 μm; 16 bits of dynamics). The whole specimen was scanned in stacking mode with 2624 projections distributed over 2 turns with a spatial resolution of 75 μm. Parameters of the acquisition are 130 kV (tube voltage), 350 μA (tube current), 12.5 frames per second, averaging of 15 frames per projections, a source-to-detector distance, and a source-to-object distance of 497 mm and 293 mm, respectively. An aluminum filter of 1.2 mm thickness was used to reduce beam hardening effects. An anti-ring procedure with random shifts of the detector was used to avoid ring artifacts. At the end of the acquisition, a set of 32 additional reference projections were recorded to correct for possible beam drift. Reconstruction of the data was done with the XAct software package (RX-solutions) with a filtered back projection algorithm based on the Feldkamp method for cone beam geometry and with a Tukey filter. Beam hardening artifacts were corrected through linearization with a polynomial function. Moreover, we acquired surface data of the upper jaw (CUF-KR-2) with an 3D Artec Space Spider and obtained 1,412,644 triangles for a surface of 234.82 cm2. The 3D models of the mandible and maxilla are figured in the Supplementary Material 1 and are available at morphomuseum.com (Grohé et al. 2023). Observations and image acquisitions of the Hunter-Schreger bands on the p4 of CUF-KR-1 were made with a Leica microscope M205C with a Flexacam C1 camera.
Linear measurements were taken with Mitutoyo calliper to the nearest 1/100 mm with results given to the nearest 1/10 mm.
Abbreviations: “CUF-KR,” Collections of Geological Department, Chulalongkorn University, Bangkok-Khorat Fossils. “KHT,” Collections of Nakhon Ratchasima Rajabhat University-Khorat Fossils. “UPPal CAR-5-001,” Collection of the University of Poitiers, Paleontology -Carnivore.
Geological and faunal context
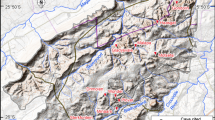
The Khorat sand pits are located along the Mun River in Nakhon Ratchasima Province, northeastern Thailand (Fig. 1). Chaimanee et al. (2006) mentioned two distinct sedimentary units in the stratigraphic sections of these sand pits: the upper unit consisting of yellowish sands and gravels of about 7–8 m thickness, and the lower unit consisting of grayish sands and gravels (more than 50-m thick). The upper unit shows oxidized yellowish sands and gravels, but the lowermost part of the unit consists of gravels, which contain rare reworked tektites, abundant tree trunks, and some clay lenses rich in organic matter, including leaves and seeds. The tektites were dated to about 780,000 years old (Haines et al. 2004) and this upper unit is therefore of Pleistocene age. The lower unit is composed of organic-rich sand that corresponds to fluviatile channel deposits with intense cross-bedding and has produced fossil remains of a few tree trunks, mammals, turtles, and crocodiles. Organic-rich clay lenses about 5–30 cm thick are interbedded within these sands and gravels. The mammalian fossils collected from this lower unit include remains of the hominoids Khoratpithecus piriyai and Khoratpithecus magnus (Chaimanee et al. 2004, 2022); hipparionines sp. indet.; the rhinocerotids Alicornops complanatum, Acerorhinus paleosinensis, Acerorhinus porpani, and Brachypotherium perimense (Chaimanee et al. 2004; Deng et al. 2013; Handa et al. 2021); several species of proboscideans (Saegusa et al. 2005; Thasod et al. 2012; Duangkrayom et al. 2017); the suids Hippopotamodon cf. sivalensis and Propotamochoerus cf. hysudricus; the anthracotheres Merycopotamus medioximus, M. thachangensis, and Microbunodon milaensis (Lihoreau et al. 2007; Hanta et al. 2008); the giraffid Bramatherium sp. and the bovids Selenoportax vexillarius, Selenoportax falconeri, Selenoportax sp., and Pachyportax giganteus (Nishioka et al. 2014, 2020).

Khorat sand pits are located along the Mun River in Nakhon Ratchasima Province, northeastern Thailand
Fossil mammals discovered from these sand pits correspond in part to those of the lower Dhok Pathan mammalian zone of the Siwaliks (Northern India and Pakistan) indicating a late Miocene age, between 9 and 6 Ma (Barry et al. 2002; Chaimanee et al. 2006). This fossiliferous sand unit corresponds to fluvial channel deposits of the paleo-Mun River system as indicated by the abundant crocodile and turtle remains. The paleoenvironment of the area was swampy mixed with closed woodland habitats based on the occurrence of the rhinocerotid Brachypotherium, which is considered a swamp dweller. The pollen assemblage, dominated by thermophilous trees and grassland, indicates a forest-grassland transition environment (Sepulchre et al. 2010). However, hydromorphic plant pollen spectra dominate, suggesting that large areas of the floodplain were covered by grasslands (Chaimanee et al. 2006).
The fossils described in this study were recovered in a Khorat sand pit by a local amateur. They originated from the fossiliferous lower sand unit and are therefore dated between 9 and 6 Ma. They will be stored after study in the Collections of the Geological Department, Chulalongkorn University, Bangkok.
Systematics
Order Carnivora Bowdich, 1821
Sub-order Feliformia Kretzoi, 1945
Family Felidae Batsch, 1788
Sub-family Felinae Batsch, 1788
Pachypanthera n. gen.
Etymology: from the greek “Pachy” = thick.
Type species: Pachypanthera piriyai n. sp.
Diagnosis: Very large felid whose mandible is characterized by a dilated anterior region raised around the huge canine; medium-sized i3 situated in front of the canine while very small i1 and i2 located behind i3; thick and very robust corpus mandibulae with a straight ventral border, and a deep and large masseteric fossa; no p1 and p2, diastema between canine and p3 with a ridge running from the canine alveolus to the anterior alveolus of p3; relatively low and elongate p4 with two well-developed accessory cuspids located mesial and distal to the main cuspid; horizontal wear facets on the cuspids of p4. Large upper canine; large infra-orbital foramen; P2 out of the line of the tooth row; P3, with a concave mesial profile, is posteriorly enlarged with a small distal accessory cusp and a narrow and flat talon; P4 with a small parastyle, a large mesio-lingually projecting protocone with a small conic cusp at the lingual extremity; relatively large M1. Presence of zigzag Hunter-Schreger bands (HSB) on the premolars.
Pachypanthera piriyai n. sp
Origin of the name: in honor of Piriya Vachajitpan, who played a critical part in recovering the fossils.
Holotype (CUF-KR-1): left hemi-mandible with the alveoli for i1–i3, canine alveolus, remains of roots of p3, p4 and partially broken off m1 crown.
Paratype (CUF-KR- 2): large fragment of right maxilla with canine alveolus, P3–P4, M1 alveolus.
Diagnosis: As for the genus.
Locality: Khorat sand pit, Nakhon Ratchasima Province, northeastern Thailand
Age: late Miocene, 9 to 6 Ma.
Description
The jaw of Pachypanthera piriyai n. gen. n. sp. (CUF-KR-1) is striking in the morphology of its anterior part (Figs. 2 and 3). This part is dilated and elevated, the alveolar plane of the canine being distinctly higher than that of the cheek teeth. There is a round alveolus for the third incisor (diameter 6.7 mm) and two small and laterally compressed alveoli for i2 and i1 (Table 1). The incisors were very small compared to the huge canine, and the third incisor was not situated medially relative to the canine but in front of the canine so that the distance between the two lower canines of the mandibles would have been quite small (Fig. 4). The alveolus of the canine is very large (length = 28.8 mm; width = 23 mm) and the long axis of the tooth is little curved (Fig. 2). There is a large diastema between canine and p3; a thick crest runs on the dorsal edge of the diastema from the posterior limit of the canine alveolus to the anterior limit of the p3 alveolus. A shallow depression situated on the buccal side of the diastema could indicate the presence of a huge upper canine being in contact with the mandible when the jaws were closed. There are two mental foramina at mid-height below the diastema, a medium-sized anterior one and a smaller posterior one (Fig. 2, Supplementary Material 1). Except under the canine where there is a small bump, the height of the corpus mandibulae is uniform from p3 to the angular process. This bump corresponds to a high steep symphysis and a long canine alveolus, the depth of the symphysis being correlated with the size of the canine root (Kurten 1957, p. 379). The corpus is also very thick (Table 2) and the lower border is straight below the masseteric fossa up to the fossa of the angular process. The fossa seems to have been deep, but there is a polymorphism in this area in some species of felids (Salles 1992, fig. 5). The ascending process (ramus) is very oblique relative to the corpus (angle between the line of the mesial border and the horizontal is about 155°). The masseteric fossa is very deep. The masseter muscle inserts on a high and robust flange running along the ascending ramus, and it also has an anterior expansion of the masseteric fossa to the level of the middle part of m1. The tooth row is slightly convex buccally. The posterior alveolus of p3 is larger than the anterior one, meaning that the distal part of the crown was probably wider than the mesial part. This tooth is not very developed (length from alveoli 17.2 mm) and is situated slightly obliquely relatively to the tooth row. The p4 is longer than the p3 (Table 1). It is elongate and relatively low, even when taking into account the horizontal wear facet on the top of the main cuspid. If we take into account this wear facet (about 2 mm), the height of p4 divided by its length is 0.7. There are two well-developed accessory cuspids. Large horizontal wear facets are present on the main and both accessory cuspids. Small shear facets are visible on the buccal side of the main and accessory cuspids under the horizontal wear facets. Although the m1 is partially broken off, remains of the base of the crown allow measurements to be taken. It was a stout tooth (Table 1); the base of the m1 paraconid is in contact with the distolingual end of p4 (tiles disposition) and the talonid tapers distally (Fig. 2a). The robustness index of m1 (maximum width divided by the length and multiplied by 100) reaches 52. The distance between canine and m1 is relatively short (64.5 mm) and is evidence for a shortened anterior part of the jaw and, of course, a shortened snout. There is no m2.

Pachypanthera piriyai n. gen. n. sp., left hemi-mandible CUF-KR-1. a Occlusal view (stereopair); b inferior view; c lateral view; d medial view (scale bar = 20 mm)

Pachypanthera piriyai n. gen. n. sp., left hemi-mandible CUF-KR-1 (scale bar = 20 mm). a Virtual rendering of the jaw in buccal view showing the roots of the check teeth. Note the absence of trace of p2. b Parasagittal section showing the huge alveolus for the canine. Abbreviations: ca, canine alveolus; d, diastema; mc, mandibular canal; mf, masseteric fossa

Digital reconstruction of the mandible of Pachypanthera piriyai n. gen. n. sp (a) in comparison with the mandible of Panthera leo (UPPal CAR-5-001) (b) in occlusal view. Scale bar = 20 mm
The microtomographic data allow for observation of the internal structures of the jaw (Fig. 3), showing a part of the large alveolus of the canine, the relatively short roots of p3, the anterior one being the shorter, with both roots being anteriorly directed, the two long anteriorly directed roots and the crown of p4, followed by the very large vertical mesial root and the smaller vertical distal one of m1. There is also a part of the carnassial crown. A longitudinal section along the tooth row (Fig. 3b) shows the large size of the canine alveolus whose direction indicates a root that is less curved and closer to the vertical than in some other carnivores. Under the diastema, the anterior alveolus of p3 and the canine alveolus are separated by a short distance.
Under a microscope, we can observe a zigzag pattern of the Hunter-Schreger bands (HSB) of enamel on the top and a large part of the p4 main cuspid and principally acute angle HSB on the distal accessory cuspid (Fig. 5). The HSB are features that increase enamel resistance to mastication stress in mammals. In carnivorans, several HSB patterns have been recognized: the HSB may be undulating with a low amplitude, forming waves with acute angles or may be zigzag (Stefen and Rensberger 1999, fig.11), the latter being characteristic of bone-cracking carnivores such as hyaenids (Stefen 1997a, 1997b). The zigzag HSB may be associated with the other types of bands in a tooth or be the sole type of HSB observed, which is the case for some hyenas (Stefen and Rensberger 1999). When there are several types of HSB in a tooth, the low amplitude ones are situated at the base of the crown and the zigzag HSB at the top of the cuspids. Here the zigzag HSB of the p4 of CUF-KR-1, visible on the top of the main cuspid, and the acute angles on the accessory ones recall those of fossil and modern bone-cracking hyenas (Stefen and Rensberger 1999; Tseng 2011). The zigzag HSB may be present in different carnivorous mammals as Hyaenodonta, Arctocyonidae, and Mesonychidae (Stefen 1997a), North American Hyaenodon (Stefen 1997b) or European Hyaenodon (Bastl et al. 2012). Each time they are associated with very robust enamel and a diet based on tough item and bone-cracking behavior.

Images of the p4 of CUF-KR-1 showing the zigzag HSB pattern in its enamel. a, General buccal view of the p4 main cuspid; b and c, details of the p4 buccal wall. Scale bars = 1 mm
The right maxilla CUF-KR-2 (Fig. 6) fits the mandible well and, considering the scarcity of large carnivores in the locality, with very high probability belongs to the same species. Nevertheless, it is a different individual whose teeth are unworn. The maxilla is preserved from the alveolus of the canine to its posterior part, the top being broken off at the level of a large infra-orbital foramen and it is followed by the beginning of the high (45.8 mm) jugal part of the zygomatic arch. It is anteriorly swollen due to the huge canine alveolus. In lateral view, the maxilla-jugal suture first runs upright from the base and then turns forward to the top up to the infra-orbital foramen; on the medial face, it is oblique from the base to the top. The crown of the single-rooted P2 is broken off (Table 2); it is laterally situated relative to the tooth row. P3 is quite low, elongate, and oblique relative to the tooth row; its occlusal profile is quite trapezoidal but with rounded angles; the crown is encircled by a weak cingulum. There is no mesial accessory cusp; there is a mesio-lingual vertical ridge and the mesial face is slightly concave; there is a moderately developed distal accessory cusp followed by a short talon. The carnassial, P4, is long, with a relatively small parastyle, a large paracone, and an elongate metastyle; a very small cusp is located at the mesio-buccal corner of the tooth; the protocone is very well developed with a conic lingual tubercle; it is as mesial as the parastyle; a mesio-lingual ridge runs from the top of the paracone to the protocone; a weak cingulum runs along the buccal and lingual faces. We observe zigzag HSB on the top of the paracone of P4. There is a single transversally oval and large alveolus for M1.

Right maxilla of Pachypanthera piriyai n. gen. n. sp. (CUF-KR-2). a Occlusal view; b medial view; c lateral view (scale bar = 20 mm)
Comparisons
The lack of p1 and p2, the straight ventral border of the mandible, and the low-crowned P3 and p4 confirm that these specimens belong to the family Felidae. Moreover, the enlarged canines; the strong horizontal wear facets on p4; the acute to zigzag HSB on the lower and upper premolars; and the deep, short, and thick hemi-mandible altogether suggest that this felid probably was a durophagous carnivore. Zigzag HSB are common in fossil and extant bone-cracking mammals (Werdelin 1989; Wang et al. 1999).
Although they are known since the Oligocene with the small-sized Proailurus Filhol, 1879, the Felidae, excepting the Machairodontinae, are represented during the Miocene through small- and medium-sized species. The large-sized felines only appear during the Pliocene and belong either to the genus Panthera, to puma-like or to cheetah-like forms. The infra-orbital foramen of Pachypanthera n. gen. is as large as in pantherines although it is smaller in other felids such as in the cheetah (Sims 2012, fig. 8). Nonetheless, the Thai carnivore differs from other extant large felids in the large size of the canine, the oblique p3/P3, and the robustness of the dentary (Figs. 4 and 7). The obliquity of the third premolars indicates a shortening of the jaws and probably reinforces the pressure during the bite. The protocone is generally smaller in the felids than in Pachypanthera n. gen., but the protocone of lions may also be elongated (photo of Werdelin, in litteris) and the protocone may also be long with a small lingual cusp in a felid (see Neofelis nebulosa in Salles 1992, fig. 11).

Correspondence factor analysis of mandibular measurements of Pachypanthera n. gen. and various extant and fossil carnivorans (Canidae, Felidae, Hyaenidae, and Percrocutidae). Projection on the plane of the two first axes. Variables = log10-transformed deepness (Dy) and thickness (Dx) of the corpus at the gaps p3–p4 (PP) and p4–m1 (PM). Canidae: 1 Vulpes lagopus, 2 Canis latrans, 3 Canis lupus, 4 Canis mesomelas, 5 Cerdocyon thous, 6 Lycalopex culpaeus, 7 Lycaon pictus, 8 Nyctereutes procyonoides, 9 Otocyon megalotis, 10 Urocyon cinereoargenteus, 11 Vulpes velox, 12 Vulpes vulpes. Felidae: 13 Acinonyx jubatus, 14 Prionailurus bengalensis, 15 Felis lybica, 16 Lynx rufus, 17 Leptailurus serval, 18 Leopardus weidii, 19 Herpailurus yagouaroundi, 20 Puma concolor, 21 Panthera leo, 22 Panthera pardus, 23 large-sized Lynx lynx, 24 Lynx lynx, 25 Caracal caracal, 26-29 Panthera spelaea. Hyaenidae: 30 Crocuta crocuta, 31 Hyaena hyaena. Percrocutidae: 32 Dinocrocuta algeriensis, 33 Dinocrocuta senyureki. Convex hulls are represented for Canidae, Felidae, and Hyaenidae + Percrocutidae. Data from Biknevicius and Ruff (1992) completed by Panthera spelaea (pers. comm., G. F. Baryshnikov) and by measurements on original specimens of Lynx lynx and Caracal (collections of the Université de Poitiers). Measurements for percrocutids made from casts
If we consider the fossil record, felids have been described from the upper stage (Pinjor) of the Siwalik hills in the Indian subcontinent. Sivafelis potens Pilgrim 1932 is based on a mandible and a small piece of maxilla (Pilgrim 1932, pl. 3, figs. 2 and 10). This species differs from Pachypanthera n. gen. in the smaller size, the relatively smaller size of the canine, and the shorter diastema between the canine and p3. In the same genus, Pilgrim placed specimens of Felis (Cynailurus) brachygnathus figured by Lydekker (1884, p. 149–151, Pl. 43, figs. 1 and 2) and another specimen from China (Zdansky 1924) that Kretzoi (1938) considered belonging to the genus Sivapanthera. Nevertheless, whatever the generic name, “S.” brachygnathus is larger than S. potens but differs from the Thai felid in the short diastema; the smaller canine and the large size of two mental foramina situated below the short diastema and the gap p2–p3, respectively.
The middle Miocene Styriofelis turnauensis (Hoernes, 1882), Miopanthera lorteti (Gaillard, 1899), the late Miocene Styriofelis vallesiensis Salesa et al., 2012, and Pristifelis attica (Wagner, 1857) are small cats that are morphologically distant from the large Pachypanthera n. gen.
In Africa, Diamantofelis ferox Morales et al. 1998, is a medium-sized felid which differs from Pachypanthera n. gen. in having a p4 with a very high main cuspid and a convex ventral border of the corpus (Morales et al. 1998, fig. 12).
Other occurrences of quite large non-machairodont cats include taxa phylogenetically closer to the extant genus Panthera. The species from the late Miocene (Vallesian) of Turkey “Felis” pamiri Ozansoy, 1965 was revised and attributed to the genus Miopanthera Kretzoi 1938 (Geraads and Peigné 2017). The authors consider that the specimen figured by Ozansoy (1965) is close to extant Felinae and even to pantherines. Its size (P4 length = 20.4 mm, m1 length = 16.1 mm) would be in the range “from that of a large caracal to that of a small leopard” (p. 423). In their conclusions (p. 424), these authors “favor its inclusion in the pantherin clade, because of its relatively large size, low p3, and large incisors” although Mio. pamiri differs “from modern Panthera species in the absence of canine grooves, longer distance between C and P3, less reduced M1, and long m1 relative to p4”. Mio. pamiri differs from Pachypanthera n. gen. by the smaller size, the relative smaller canines, the relative longer P2, the relative higher P3, the relative larger parastyle of P4, the non-oblique p3, the lower half of the symphysis longer than the upper one, and the slightly convex ventral border of the corpus (Geraads and Peigné 2017, fig. 1d).
Panthera blytheae Tseng et al. 2014 was found in the early Pliocene of Tibet and is considered by the authors as the oldest known representative of the genus Panthera (Tseng et al. 2014). The maxilla of P. blytheae differs from that of the Thai carnivoran by the smaller size (P4 length = 18.4 mm), the relatively smaller canines, the presence of a small supplementary buccal cusp on the posterior cingulum of P3, the presence of converging ridges on the buccal surface of P4, the presence of undulating HSB throughout the cheek teeth, and the absence of zigzag HSB. Some mandibular remains recorded in the type locality and other localities from the same basin are attributed to the same species. They differ from Pachypanthera n. gen. by the smaller size, the steeper symphysis, the premolars on a straight line, p3 not being oblique relative to the tooth row. Note that, for Hemmer (2023), this species P. blytheae is the type of a new genus named Palaeopanthera.
The genus Panthera became more common in Eurasia during the Plio-Pleistocene. Kretzoi (1938) described isolated teeth from a Villafranchian locality as belonging to Leo gombaszögensis (sic) which later was changed to Panthera gombaszoegensis (Kretzoi 1938) and identified from many Pleistocene localities of Eurasia from western Europe to China (Kretzoi 1938; Hemmer 1971, 1972; Sotnikova and Vislobokova 1990; Argant 1991; Hemmer et al. 2001; Langlois 2002; O’Regan 2002; Baryshnikov 2011; Jiangzuo and Liu 2020; and see Marciszak and Lipecki 2022, fig. 1). Some authors, following Hemmer (Hemmer 1971; Hemmer et al. 2010), considered Pa. gombaszoegensis as close to the jaguar but this opinion was discussed recently (Chatar et al. 2022) and this species may be closer to the tiger. Pa. gombaszoegensis has the same oblique p3 and a deep masseter fossa like Pachypanthera n. gen. but differs from the Thai carnivoran in its smaller size, the lack of ridge on the dorsal surface of the diastema, steeper symphysis, two large mental foramina, less robust corpus, relatively smaller canines, larger cingulid around the distal accessory cuspid of p4, no laterally positioned P3, and relatively larger parastyle of P4.
Another large felid was present in the middle and late Pleistocene, Panthera spelaea (Goldfuss, 1810), the cave lion which is recorded from Spain (Alvarez-Lao et al. 2020) to Alaska and Canada (Kurten 1985; Stuart and Lister 2011). Its remains are common in many prehistoric excavations. We include in our comparison Panthera mosbachensis Dietrich, 1962 which is considered by some authors the putative direct ancestor of P. spelaea. Both species are well known for their large size although it is highly variable. The length of m1 varies from 25.5 to 34.2 mm in Eurasian specimens (Baryshnikov and Boeskorov 2001, table 8; Baryshnikov 2011; Sotnikova and Vislobokova 1990) and 25.7 to 34 mm for a width of 14 to 18 mm in another sample (Langlois 2002, fig. 3) against 25 to 31 mm for the extant lion (Turner 1984, fig. 2). These measurements fit those of Pachypanthera n. gen. Nevertheless, in P. spelaea the lower incisors, and especially the two central incisors, are far larger (Argant and Brugal 2017, table 2) and the upper canine is relatively smaller; its length varies from 22.6 to 34.4 mm (Argant and Brugal 2017, table 1), while the estimated length of the alveolus is 48 mm for Pachypanthera n. gen. This difference in canine dimensions between the two species is even greater if considering the relative size of the canine.
The overall size of P. spelaea varies as a function of time and geographic dispersion, allowing the creation of some subspecies (Sotnikova and Nikolaskiy 2006; Marciszak and Stefaniak 2010; Marciszak et al. 2014, 2021; Argant and Brugal 2017) based on size or some morphological details. The crown-root junction of the lower canine is at a higher level than that of the cheek teeth but less so than in Pachypanthera n. gen. Panthera spelaea differs from the latter in its larger incisors, its relatively smaller upper canine, the position of its i3, its P3 in the line with the tooth row and with an accessory mesial cusp, its relative larger parastyle of P4, its narrow symphysis (see Baryshnikov and Petrova 2008, fig. 1), its large mental foramen close to the anterior root of p3, the less robust corpus mandibulae (Fig. 7), and the absence of horizontal wear facets on the cuspids. We observed small horizontal wear facets on the top of premolars on a photograph of a mandible of leopard (Werdelin, in litteris). Nevertheless, this feature is uncommon in the pantherines. When we find a fossil, statistically we have a great chance to get a common specimen of the population, and we may suggest that the presence of a horizontal wear facet on the top of p4 of Pachypanthera n. gen. is probably a character of the population. It seems that this peculiar wear pattern does not exist in the bulk of the P. spelaea populations.
Discussion
As we saw above, Pachypanthera n. gen. does not fit exactly any species of Felidae. The robustness of the corpus mandibulae seems to be a major character that separates Pachypanthera n. gen. from the other Felidae. A correspondence analysis is carried on the measurements, depth (DyPP and DyPM), and width (DxPP and DxPM) of the corpus mandibulae at the level of p3–p4 and p4–m1 gaps of some Felidae, Hyaenidae, Percrocutidae, and Canidae (Fig. 7, Table 3). The data come from Biknevicius and Ruff (1992), Baryshnikov (in litteris), and original. We did not work on the raw data but on the logarithmic values and we choose the correspondence analysis to avoid directly the “size effect” in the results (Lebart et al. 1984). In the first axis the variables DxPM and DxPP lie on the positive part and DyPM with DyPP on the negative side. In the first group we find the hyenas (Hyaenidae and Percrocutidae), the large Felidae, and two Canidae (gray wolf and lycaon). On the negative side are the bulk of the Canidae and Felidae. The separation is based on differences in proportion, the positive side being linked to greater robustness. Into this group, there is a vertical gradient from forms in which the larger width is anterior (DxPP) to forms in which the larger width is posterior (DxPM). We note that Pachypanthera n. gen. is isolated in an extreme position with a very robust posterior part of the mandible.
The zigzag pattern of the HSB of the tooth enamel, well developed in the Hyaenidae and in Pachypanthera n. gen., is also present in other carnivorans and could be linked to bone-cracking activity as “the flexible patterns of HSB specialization allow carnivorans to adapt rapidly to changing dietary environments” (Tseng 2011, p. 1165). We may estimate that Pachypanthera n. gen. was probably engaged, at less partially, in breaking tough food items.
Sub-family Machairodontinae Gill, 1872
Genus and species indet.
Description
This left upper canine fragment (KHT-1) was discovered in the Khorat sand pits and stored at the Nakhon Ratchasima Rajabhat University collection, Nakhon Ratchasima Province in northeastern Thailand. It is a small piece (height = 65 mm, mesio-distal diameter = 27 mm) of a flattened upper canine with crenulations (about 20 crenulations by centimetre) on the distal edge (Fig. 8). It certainly belongs to a quite large sabre-toothed felid of the tribe Homotherini in the sense of Kurten (1963), but we cannot give a more precise identification because of the fragmentary nature of the material.

Left upper canine of a machairodontine felid (KHT-1) in lateral and medial views (a, b). cr, crenulations on the distal edge. Scale bar = 1 cm
General discussion
The two large-sized predators of the Khorat basin (Pachypanthera piriyai n. gen., n. sp., a pantherine, and an indeterminate sabre-toothed felid) are the only carnivorans known in the late Miocene of Thailand. Small- and medium-sized carnivorans common in the late Miocene localities of the Old World, such as mustelids and viverrids, were not recovered in the Khorat sand pits, probably due to the grain size of the sand of the deposits. We note that, until the late Miocene, large felids of the size of the extant Pantherini are only present in Machairodontinae which had certainly a different mode of life and an ecological niche different from that of the Felinae. Despite individual teeth appear “less useful for size estimations of fossil material” (Turner and O’Reagan 2002) than other elements, the body mass of Pachypanthera n. gen. is estimated at 142 kg from the size of its m1 following the regression of Legendre and Roth (1988, fig. 1, ln y = k ln X + ln b) for Felidae. Nevertheless, the body mass estimation of one specimen, whatever the method employed, is an approximation that does not give access to the variation into the species, which is known to be important among the extant felids. For example, the range of body mass for the lion is 122–240 kg, and those of the tiger and the leopard are 100–250 and 30–70 kg, respectively (Macdonald 2001; see also Faurby et al. 2021). The body mass of the sabre-toothed cat from Khorat cannot be estimated but should be between that of Lokotunjailurus and Machairodus.
The origin of the pantherine also may be discussed. Because of the presence in the middle Pliocene of Laetoli (Tanzania) of quite large felids (Barry 1987; Werdelin and Dehghani 2011; Hemmer 2022), some authors considered that the origin of Panthera was in Africa (“The origin of the genus Panthera is probably in Africa at 3.5 Ma at Laetoli in Tanzania,” authors’ French translation from Argant and Argant 2018, p. 279). Nonetheless, the true question is where these fossils were coming from. The holotype of an Asiatic species from China, Pa. palaeosinensis (Zdansky 1924), bears the characters of Panthera (Zdansky 1924 pl. 32) and is considered close to the tiger; the locality where it is coming from can be dated to the late Pliocene. Another Asian record is Pa. zdanskyi Mazak et al. 2011, of which holotype comes from Longdan (northwestern China) and is dated to 2.55–2.16 Ma. Another answer was the record of Panthera blytheae, the oldest Panthera coming from the early Pliocene of Asia (Tseng et al. 2014) in the southwestern Tibetan Plateau. The dating is moved further back by Pachypanthera n. gen., which seems to be the earliest recorded large Felinae (Pantherini) in the “Hipparion fauna” of the late Miocene of Southeast Asia.
The premolars P3 and p4 are conical and robust in most of the Hyaenidae and Percrocutidae which are considered being bone crackers. Nevertheless, other carnivorans are considered bone breakers or bone crushers. Several genera of the Borophaginae, a sub-family of Canidae in the Cenozoic of North America, are considered bone crushers. We have tested the index (width p4/length p4) × 100 on the data coming from 23 genera furnished in a large study of Borophaginae (Wang et al. 1999). The index varies from 40 to 74 in this sub-family, the larger indices being those of the larger species. The same index reaches 58 in Pachypanthera n. gen., and it is close to the larger indices of the Borophaginae. Moreover, the presence of zigzag HSB on p4 and P4 indicates a capacity to durophagy in the Thai carnivoran. In a large terrestrial carnivoran this capacity is probably linked to the breaking of bones. We also must remember the large and close themselves canines which could be a useful tool to break. The beginning of a bone-breaking or a bone-crushing process could be a valid hypothesis concerning the way of life of Pachypanthera, although this adaptation to bone cracking seems “weird” in felids.
Pachypanthera n. gen. was a large carnivoran with an extremely robust mandible and very robust canines sustained by a strong swollen anterior mandible and propelled by powerful muscles probably used to perforate and to break large bones. We do not have the limb bones of Pachypanthera n. gen., and it cannot be known whether it was a good runner and an active hunter or specialized in feeding on carcasses of large-sized animals such as proboscideans, rhinos, suids, anthracotheres, giraffids, bovids, the hipparionin equids, or the primate Khoratpithecus, found in the Khorat sand pits. Although the material from these sand pits is rather scarce, it brings new insights into the evolutionary history of Neogene mammals of Southeast Asia, in a geographic place which is partly “terra incognita”.
References
Alvarez-Lao DJ, Alvarez-Vena A, Ballesteros D, García N, Laplana C (2020) A cave lion (Panthera spelaea) skeleton from Torca del Leon (NW Iberia): Micromammals indicate a temperate and forest environment corresponding to GI-11 (MIS 3). Quat Sci Rev 229:106123. https://doi.org/10.1016/j.quascirev.2019.106123
Argant A (1991) Carnivores quaternaires de Bourgogne. Doc Lab Géol Lyon 115:1–301
Argant A, Argant J (2018) La brèche à carnivores du Pléistocène moyen de Château (Saône-et-Loire, France). Quaternaire 29:271–285. https://doi.org/10.4000/quaternaire.10390
Argant A, Brugal JP (2017) The cave lion Panthera (Leo) spelaea and its evolution: Panthera spelaea intermedia nov. subspecies. Acta Zool Cracov 60:59–104. https://doi.org/10.3409/azc.60_2.59
Barry JC (1987) Large carnivores (Canidae, Hyaenidae, Felidae) from Laetoli. In: Leakey MD, Harris JM (eds) Laetoli: a Pliocene site in Northern Tanzania. Clarendon Press, Oxford, pp 235–258
Barry JC, Morgan ME, Flynn LJ, Pilbeam D, Behrensmeyer AK, Raza SM, Khan IA, Badgley C, Hicks J, Kelley J (2002) Faunal and environmental change in the late Miocene Siwaliks of northern Pakistan. Paleobiology 28:1–71. https://doi.org/10.1666/0094-8373(2002)28[1:FAECIT]2.0.CO;2
Baryshnikov GF (2011) Pleistocene Felidae (Mammalia, Carnivora) from the Kudaro paleolithic cave sites in the Caucasus. Proc Zool Inst Russ Acad Sci 315:197–226. https://doi.org/10.31610/trudyzin/2011.315.3.197
Baryshnikov GF, Boeskorov G (2001) The Pleistocene cave lion, Panthera spelaea (Carnivora, Felidae) from Yakutia, Russia. Cranium 15:7–24
Baryshnikov GF, Petrova EA (2008) Cave lion (Panthera spelaea) from the Pleistocene of Chuvashyia, European Russia. Russ J Theriol 7:33–40
Batsch AJGK (1788) Versuch einer Anleitung zur Kenntniss und Geschichte der Thiere und Mineralien, für akademische Vorlesungen entworfen, und mit den nöthigsten Abbildungen versehen. Vol. 1. Akademische Buchhandlung, Jena
Bastl K, Semprebon G, Nagel D (2012) Low-magnification microwear in Carnivora and dietary diversity in Hyaenodon (Mammalia: Hyaenodontidae) with additional information on its enamel microstructure. Palaeogeogr Palaeoclim Palaeoecol 348-349:13–20. https://doi.org/10.1016/j.palaeo.2012.05.026
Biknevicius AR, Ruff CB (1992) The structure of the mandibular corpus and its relationship in feeding behaviour in extant carnivorous. J Zool 228:479–707. https://doi.org/10.1111/j.1469-7998.1992.tb04450.x
Bowdich TE (1821) An analysis of the natural classifications of Mammalia: for the use of students and travellers. Smith J, Paris
Chaimanee Y, Suteethorn V, Jintasakul P, Vidthayanon C, Marandat B, Jaeger J-J (2004) A new orang-utan relative from the Late Miocene of Thailand. Nature 427:439–441. https://doi.org/10.1038/nature02245
Chaimanee Y, Yamee C, Tian P, Khaowiset K, Marandat B, Tafforeau P, Nemoz C, Jaeger J-J (2006) Khoratpithecus piriyai, a Late Miocene hominoid of Thailand. Am J Phys Anthropol 131:311–323. https://doi.org/10.1002/ajpa.20437
Chaimanee Y, Lazzari V, Yamee C, Suraprasit K, Rugbumrung M, Chaivanich K, Jaeger J-J (2022) New materials of Khoratpithecus, a late Miocene hominoid from Nakhon Ratchasima Province, Northeastern Thailand, confirm its pongine affinities. Palaeontogr Abt A 323:147–186. https://doi.org/10.1127/pala/2022/0129
Chatar N, Michaud M, Fischer V (2022) Not a jaguar after all? Phylogenetic affinities and morphology of the Pleistocene felid Panthera gombaszoegensis. Pap Palaeontol 8. https://doi.org/10.1002/spp2.1464
Deng T, Hanta R, Jintasakul P (2013) A new species of Aceratherium (Rhinocerotidae, Perissodactyla) from the late Miocene of Nakhon Ratchasima, northeastern Thailand. J Vert Paleontol 33:977–985. https://doi.org/10.1080/02724634.2013.748058
Dietrich WO (1962) Paläontologische Systematik und Nomenklatur. Mitt Geol Staatsinst Hamburg 31:382–384
Duangkrayom J, Wang S, Deng T, Jintasakul P (2017) The first Neogene record of Zygolophodon (Mammalia, Proboscidea) in Thailand: implications for the mammutid evolution and dispersal in Southeast Asia. J Paleontol 91:179–193. https://doi.org/10.1017/jpa.2016.143
Egi N, Ogino S, Takai M (2018) Neogene Irrawaddy fauna of Central Myanmar: Carnivora. Fossils 104:21–33
Faurby S, Morlo M, Werdelin L (2021) CarniFOSS: a database of the body mass of fossil carnivores. Glob Ecol Biogeogr 30:1958–1964. https://doi.org/10.1111/geb.13369
Filhol H (1879) Étude des mammifères fossiles de Saint-Gérand-le-Puy (Allier). Ann Sci Géol Paris 10:1–253
Gaillard C (1899) Mammifères miocènes nouveaux ou peu connus de La Grive-Saint-Alban (Isère). Arch Mus Hist nat Lyon 7: 1–79
Geraads D, Peigné S (2017) Re-Appraisal of ‘Felis’ pamiri Ozansoy, 1959 (Carnivora, Felidae) from the Upper Miocene of Turkey: the Earliest Pantherin Cat? J Mammal Evol 24:415–425. https://doi.org/10.1007/s10914-016-9349-6
Gill T (1872) Arrangement of the families of mammals and synoptical tables of characters of the subdivisions of mammals. Smithsonian Miscel Coll 230:1–98
Goldfuss GA (1810) Die Umgebungen von Muggendorf. Palm JJ, Erlangen
Grohé C, Mazurier A, Blasi-Toccacceli A, Bonis L de, Chaimanee Y, Chavasseau O, Suraprasit K, Rugbumrung M, Jaeger J-J (2023) 3D models related to the publication: A new large pantherine and a sabre-toothed cat (Mammalia, Carnivora, Felidae) from the late Miocene hominoid-bearing Khorat sand pits, Nakhon Ratchasima Province, northeastern Thailand. MorphoMuseuM
Haines PW, Howard KT, Ali JR, Burrett CF, Bunopas S (2004) Flood deposits penecontemporaneous with ~0.8 Ma tektite fall in NE Thailand: impact-induced environmental effects? Earth Planet Sci Lett 225:19–28. https://doi.org/10.1016/j.epsl.2004.05.008
Handa N, Nishioka Y, Duangkrayom J, Jintasakul P (2021) Brachypotherium perimense (Perissodactyla, Rhinocerotidae) from the Miocene of Nakhon Ratchasima, Northeastern Thailand, with comments on fossil records of Brachypotherium. Hist Biol 33:1642–1660. https://doi.org/10.1080/08912963.2020.1723578
Hanta R, Ratanasthien B, Kunimatsu Y, Saegusa H, Nakaya H, Nagaoka S, Jintasakul P (2008) A new species of Bothriodontinae, Merycopotamus thachangensis (Cetartiodactyla, Anthracotheriidae) from the Late Miocene of Nakhon Ratchasima, northeastern Thailand. J Vert Paleontol 28:1182–1188. https://doi.org/10.1671/0272-4634-28.4.1182
Hemmer H (1971) Zur Charakterisierung und stratographischen Bedeutung von Panthera gombaszoegensis (Kretzoi, 1938). N Jb Geol Paläontol Mh 12:701–711
Hemmer H (1972) Zur systematischen Stellung von ‘Jansofelis vaufreyi’ Bonifay, 1971, und Felis lunelensis Bonifay, 1971, aus dem Pleistozän Südfrankreichs (Carnivora, Felidae). N Jb Geol Paläont Mh 4:215–223
Hemmer H (2022) The identity of the “lion”, Panthera principialis sp. nov., from the Pliocene Tanzanian site of Laetoli and its significance for molecular dating the pantherine phylogeny, with remarks on Panthera shawi (Broom, 1948), and a revision of Puma incurva (Ewer, 1956), the Early Pleistocene Swartkrans “leopard” (Carnivora, Felidae). Palaeobio Palaeoenv https://doi.org/10.1007/s12549-022-00542-2
Hemmer H (2023) The evolution of the palaeopantherine cats, Palaeopanthera gen. nov. blytheae (Tseng et al. 2014) and Palaeopanthera pamiri (Ozansoy, 1959) comb. nov. (Mammalia, Carnivora, Felidae). Palaeobio Palaeoenv https://doi.org/10.1007/s12549-023-00571-5
Hemmer H, Kahlke R-D, Vekua AK (2001) The Jaguar - Panthera onca gombaszoegensis (Kretzoi, 1938) (Carnivora: Felidae) in the late Lower Pleistocene of Akhalkalaki (South Georgia; Transcaucasia) and its evolutionary and ecological significance. Geobios 34:475–486. https://doi.org/10.1016/S0016-6995(01)80011-5
Hemmer H, Kalhke R-D, Vekua AK (2010) Panthera onca georgica ssp. nov. from the Early Pleistocene of Dmanisi (Republic of Georgia) and the phylogeography of jaguars (Mammalia, Carnivora Felidae). N Jb Geol Paläontol Abh 257:115–127. https://doi.org/10.1127/0077-7749/2010/0067
Hoernes R (1882) Säugethier-Reste aus der Braunkohle von Göriach bei Turnau in Steiermark. Jb K-K geol Reichsanst 32: 153–164
Jiangzuo Q, Liu J (2020) First record of the Eurasian jaguar in southern Asia and a review of dental differences between pantherine cats. J Quat Sci 35:817–830. https://doi.org/10.1002/jqs.3222
Kitchener AC, Breitenmoser-Würsten C, Eizirik E, Gentry A, Werdelin L, Wilting A, Yamaguchi N, Abramov AV, Christiansen P, Driscoll C, Duckworth JW, Johnson W, Luo S-J, Meijaard E, O’Donoghue P, Sanderson J, Seymour K, Bruford M, Groves C et al (2017) A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/ SSC Cat Specialist Group. Cat News Special Issue 11:80
Kretzoi M (1938) Die Raubtiere von Gombaszög nebst einer Übersicht der Gesamtfauna. Ann Mus Natl Hungarici 31:88–157
Kretzoi M (1945) Bemerkungen über das Raubtiersystem. Ann Mus Natl Hungarici 38:59–83
Kurten B (1957) Percrocuta Kretzoi (Mammalia, Carnivora), a group of Neogene hyenas. Act Zool Cracov 2:375–403
Kurten B (1963) Note on some Pleistocene mammal migrations from the Palearctic to the Nearctic. Eiszeitalter und Gegenwart 14:96–103
Kurten B (1985) The Pleistocene lion of Beringia. Ann Zool Fennici 22:117–121
Langlois A (2002) Présence de Panthera gombaszoegensis Kretzoi, 1938 à la grotte XIV (Cénac-et-Saint-Julien). Paleo 14:213–220. https://doi.org/10.4000/paleo.1449
Lebart L, Morineau, A, Warwick, KM (1984) Multivariate descriptive statistical analysis. Correspondence analysis and related techniques for large matrices. Wiley, New York
Legendre S, Roth C (1988) Correlation of carnassial tooth size and body weight in recent carnivores (Mammalia). Hist Biol 1:85–98
Lihoreau F, Barry J, Blondel C, Chaimanee Y, Jaeger J-J, Brunet M (2007) Anatomical revision of the genus Merycopotamus (Artiodactyla; Anthracotheriidae): its significance for Late Miocene mammal dispersal in Asia. Palaeontology 50:503–524. https://doi.org/10.1111/j.1475-4983.2006.00643.x
Lydekker R (1884) Indian tertiary and post-tertiary vertebrata. Part VI. Siwalik and Narbada Carnivora. Mem Geol Surv India, Palaeontol Indica 2:178–351
Macdonald DW (2001) Family of cats. In: The new encyclopedia of mammals. Oxford University Press, Oxford, pp 9–39
Marciszak A, Lipecki G (2022) Panthera gombaszoegensis (Kretzoi, 1938) from Poland in the scope of the species evolution. Quat Int 633:36–51. https://doi.org/10.1016/j.quaint.2021.07.002
Marciszak A, Lipecki G, Pawłowska K, Jakubowski G, Ratajczak-Skrzatek U, Zarzecka-Szubińska K, Nadachowski A (2021) The Pleistocene lion Panthera spelaea (Goldfuss, 1810) from Poland – A review. Quat Int 605–606:213–240. https://doi.org/10.1016/j.quaint.2020.12.018
Marciszak A, Schouwenburg C, Darga R (2014) Decreasing size process in the cave (Pleistocene) lion Panthera spelaea (Goldfuss, 1810) evolution: a review. Quat Int 339–340:245–257. https://doi.org/10.1016/j.quaint.2013.10.008
Marciszak A, Stefaniak K (2010) Two forms of cave lion: Middle Pleistocene Panthera spelaea fossilis Reichenau, 1906 and Upper Pleistocene Panthera spelaea spelaea Goldfuss, 1810 from the Bisnik Cave, Poland. N Jb Geol Paläont Abh 258:339–351. https://doi.org/10.1127/0077-7749/2010/0117
Mazak JH, Christiansen P, Andrew C, Kitchener AC (2011) Oldest known pantherine skull and evolution of the tiger. PLoS ONE 6:e25483. https://doi.org/10.1371/journal.pone.0025483
Morales J, Pickford M, Soria D, Fraile S (1998) New camivores from the basal Middle Miocene of Arrisdrift, Namibia. Eclo Geol Helv 91:27–40
Nishioka Y, Hanta R, Jintasakul P (2014) Note on giraffe remains from the Miocene of continental Southeast Asia. J Sci Tech Mahasarakham Univ 33:365–377
Nishioka Y, Vidthayanon C, Hanta R, Duangkrayom J, Jintasakul P (2020) Neogene Bovidae from Tha Chang Sandpits, Nakhon Ratchasima, Northeastern Thailand. Thailand Nat Hist Mus J 14:59–72
O’Regan HJ (2002) A phylogenetic and palaeoecological review of the Pleistocene felid Panthera gombaszoegensis. PhD dissertation. Liverpool John Moores University. https://doi.org/10.24377/LJMU.t.00004925
Ozansoy F (1965) Etude des gisements continentaux et des Mammifères du Cénozoïque de Turquie. Mém Soc géol France 44:1–92
Pilgrim GE (1932) The fossil Carnivora of India. Mem Geol Survey India, Palaeontol Indica 18:1–232
Saegusa H, Thasod Y, Ratanasthien B (2005) Notes on Asian stegodontids. Quat Int 126–128:31–48. https://doi.org/10.1016/j.quaint.2004.04.013
Salesa MJ, Antón M, Morales J, Peigné S (2012) Systematics and phylogeny of the small felines (Carnivora, Felidae) from the late Miocene of Europe: a new species of Felinae from the Vallesian of Batallones (MN 10, Madrid, Spain). J Syst Palaeontol 10:87–102. https://doi.org/10.1080/14772019.2011.566584
Salles LO (1992) Felid phylogenetics: extant taxa and skull morphology (Felidae, Aeluroidea). Am Mus Novitates 3047:1–67
Sepulchre P, Jolly D, Ducrocq S, Chaimanee Y, Jaeger J-J, Raillard A (2010) Mid-Tertiary paleoenvironments in Thailand: pollen evidences. Clim Past 6:461–473. https://doi.org/10.5194/cp-6-461-2010
Sicuro FL, Oliveira LFB (2011) Skull morphology and functionality of extant Felidae (Mammalia: Carnivora): a phylogenetic and evolutionary perspective. Zool J Linn Soc 161:414–462. https://doi.org/10.1111/j.1096-3642.2010.00636.x
Sims ME (2012) Cranial morphology of five felids: Acinonyx jubatus, Panthera unca, Panthera pardus, Puma concolor, Uncia uncia. Russ J Theriol 11:157–170
Sotnikova MV, Nikolaskiy P (2006) Systematic position of the cave lion Panthera spelaea (Goldfuss) based on cranial and dental characters. Quat Int 142–143:218–228. https://doi.org/10.1016/j.quaint.2005.03.019
Sotnikova MV, Vislobokova IA (1990) Pleistocene mammals from Lakhuti, southern Tajikistan, U.S.S.R. Quartärpaläontol 8:237–344
Stefen C (1997a) The enamel of Creodonta, Arctocyonidae, and Mesonychidae (Mammalia), with special reference to the appearance of Hunter–Schreger bands. Paläontol Z 71:291–303. https://doi.org/10.1007/BF02988497
Stefen C (1997b) Differentiations in Hunter-Schreger bands of carnivores. In: Von Koenigswald W, Sander PM (eds) Tooth enamel microstructure. Balkema, Rotterdam, pp 123–136
Stefen C, Rensberger JM (1999) The specialized structure of hyaenid enamel: description and development within the lineage including percrocutids. Scanning Microsc 13:363–380
Stuart AJ, Lister AM (2011) Extinction chronology of the cave lion Panthera spelaea. Quat Sci Rev 30:2329-2340. https://doi.org/10.1016/j.quascirev.2010.04.023
Thasod Y, Jintasakul P, Ratanasthien B (2012) Proboscidean fossil from the Tha Chang Sand pits, Nakhon Ratchasima Province, Thailand. J Sci Tech Mahasarakham Univ 31:33–44
Tseng ZJ (2011) Variation and implications of intra-dentition Hunter-Schreger band pattern in fossil hyaenids and canids (Carnivora, Mammalia). J Vert Paleontol 31:1163–1167. https://doi.org/10.1080/02724634.2011.602161
Tseng ZJ, Wang X, Slater GJ, Takeuchi GT, Li Q, Liu J, Xie G (2014) Himalayan fossils of the oldest known pantherine establish ancient origin of big cats. Proc R Soc B 281:20132686. https://doi.org/10.1098/rspb.2013.2686
Turner A (1984) Dental sex dimorphism in European Lions (Panthera leo L.) of the Upper Pleistocene:palaeoecological and palaeoethological implications. Ann Zool Fennici 21:1–8
Turner A, O’Reagan HJ (2002) The assessment of size in fossil Felidae. Estud Geol 58:45–54
Wagner JA (1857) Neue Beiträge zur Kenntniss der fossilen Säugthier-Überreste von Pikermi. Abh Bayer Akad Wiss 8:109–158
Wang XM, Tedford RH, Taylor BE (1999) Phylogenetic systematics of the Borophaginae (Carnivora, Canidae). Bull Am Mus Nat Hist 243:1–391
Werdelin L (1983) Morphological patterns in the skulls of cats. Biol J Linn Soc 19:375–391. https://doi.org/10.1111/j.1095-8312.1983.tb00793.x
Werdelin L (1989) Constraint and adaptation in the bone-cracking canid Osteoborus (Mammalia, Canidae). Paleobiology 15:387–401. https://doi.org/10.1017/S009483730000957X
Werdelin L, Yamaguchi N, Johnson WE, O’Brien SJ (2010) Phylogeny and evolution of cats (Felidae). In: Macdonald D, Loveridge A (eds) Biology and conservation of wild felids. Oxford University Press, Oxford, pp 59–82
Werdelin L, Dehghani R (2011) Carnivora. In: Harrison T (ed) Paleontology and geology of Laetoli: Human evolution in context. Vol 2: Fossil Hominins and the Associated Fauna. Springer, New York, pp 189–232
Zdansky O (1924) Jungtertiäre Carnivoren Chinas. Palaeontol Sinica 2:1–149
Acknowledgements
This work was supported by the PALEVOPRIM laboratory (UMR CNRS 7262 Université de Poitiers). We thank Pratueng Jintasakul for giving permit to study a tooth of Machairodontinae. We are grateful to Mana Rugbumrung (DMR Bangkok) who made a cast of the maxilla of Pachypanthera and Alicia Blasi-Toccacceli (PALEVOPRIM) for scanning this maxilla for us. We thank Sabine Riffaut (PALEVOPRIM) for help in the figures. This work was partially supported by the grant ANR-18-CE92-0029 (to O. Chavasseau). We thank Manuel Salesa and Narimane Chatar for their careful reviews of the manuscript and their relevant suggestions, Lars Werdelin for his relevant comments and the revision of a previous version of the text, G.F. Baryshnikov who sent us measurements of Panthera spelaea and X. Wang for sending additional photographs of P. blytheae. But, all possible mistakes would be at the charge of the authors.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Aurora Grandal d’Anglade
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary material
ESM 1
Composite digital reconstruction of the face of Pachypanthera piriyai n. gen. n. sp. in a, left lateral view. b, anterior view. c, left antero-lateral view. d, posterior view. Based on 2 individuals of similar size. Yellow = fossil hemi-mandible and maxilla. Grey = mirrored parts. (PDF 610 kb)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
de Bonis, L., Chaimanee, Y., Grohé, C. et al. A new large pantherine and a sabre-toothed cat (Mammalia, Carnivora, Felidae) from the late Miocene hominoid-bearing Khorat sand pits, Nakhon Ratchasima Province, northeastern Thailand. Sci Nat 110, 42 (2023). https://doi.org/10.1007/s00114-023-01867-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00114-023-01867-4