
Abstract
Salt stress is a critical factor that affects the growth and development of plants. Salicylic acid (SA) is an important signal molecule that mitigates the negative effects of salt stress on plants. To elucidate salt tolerance in large pink Dianthus superbus L. (Caryophyllaceae) and the regulatory mechanism of exogenous SA on D. superbus under different salt stresses, we conducted a pot experiment to evaluate leaf biomass, leaf anatomy, soluble protein and sugar content, and the relative expression of salt-induced genes in D. superbus under 0.3, 0.6, and 0.9% NaCl conditions with and without 0.5 mM SA. The result showed that exposure of D. superbus to salt stress lead to a decrease in leaf growth, soluble protein and sugar content, and mesophyll thickness, together with an increase in the expression of MYB and P5CS genes. Foliar application of SA effectively increased leaf biomass, soluble protein and sugar content, and upregulated the expression of MYB and P5CS in the D. superbus, which facilitated in the acclimation of D. superbus to moderate salt stress. However, when the plants were grown under severe salt stress (0.9% NaCl), no significant difference in plant physiological responses and relevant gene expression between plants with and without SA was observed. The findings of this study suggest that exogenous SA can effectively counteract the adverse effects of moderate salt stress on D. superbus growth and development.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Large pink Dianthus superbus L. (Caryophyllaceae) is a loosely tufted perennial species whose flowers with long calyx tubes produce large amounts of nectar and strong scent. So it’s extensively utilized in ornamental horticulture and housing decoration (Bittrich 1993; Ma et al. 2017). The genus Dianthus L. comprises about 300 species that are distributed across globe but is most abundant in the Mediterranean. D. superbus is widely but patchily distributed in western, central, and eastern Europe, as well as in northern Asia (Bittrich 1993). Also, D. superbus is a rare and strictly protected plant in Poland (Rosenthal 2010; Opdekamp et al. 2012), Latvia, and Lithuania (Hooftman et al. 2003), and has been shown to be of significant medicinal value because of its sedative, analgesic, and urinary effects. Based on these features, an expansion of areas for the propagation of D. superbus is warranted (Ma et al. 2017). However, techniques for the management of D. superbus and their physiological response to the environment remain unclear. Therefore, it is necessary to investigate the effects of environmental factors on the growth of D. superbus.
Salinity stress is one of the most important environmental stressors that limit plant growth and productivity in various parts of the world, particularly arid and semiarid regions (Saleh 2012; Li et al. 2005). One of the most sensitive aspects of plant growth is the growth and development of leaves. In crop plants, a decline in leaf expansive growth is often the first detectable response to salt stress (Bradford and Hsiao 1982). Furthermore, the anatomical basis for inhibited leaf growth under saline conditions has been investigated in various species, as example of cotton, bean (David and Park 1979).
Salt stress, in addition to growth and morphology effects, alters various biochemical and physiological processes in plants, thereby resulting in a metabolic imbalance in plants (Iqbal et al. 2006;Roussos et al. 2007). Changes in protein hydration is one of the responses of plant cells to high-salt stress (Doganlar et al. 2010). Soluble proteins are the most active components of plant cells as these serve as a resource for enzymes and a modulator of metabolic processes. Total soluble protein content increases in Pancratium maritimum when subjected to high salt concentrations (Khedr et al. 2003). Soussi et al. (1998) and Roussos et al. (2013) reported that salt stress significantly affected the carbon metabolism in plants. Salt-stressed maize plants also exhibit a decrease in soluble sugar (SS) content (Khodary 2004). However, extensive evidence indicates that exogenously applied salicylic acid (SA) can ameliorate salt stress in cucumber seedlings (Yildirim et al. 2008), maize (Gunes et al. 2005), and soybean (Hamayun et al. 2010) by influencing various processes in plants, including carbon metabolism, proteometabolism, and the expression of relevant genes (Jayakannan et al. 2013).
Salicylic acid is a plant phenolic that is often considered as a signaling molecule that triggers plant defense responses to biotic and abiotic stress (Khan et al. 2013). Salicylic acid could indirectly act as cell stress protector by altering the gene expression and synthesis of defense compounds such as proline and jasmonic acid (JA) (Wani et al. 2016). Generally, plants resist stress injuries or elicit an effective defense response by activating dormancy-associated genes and synthesize stress resistance-related proteins. Various genes in different species have been reported to respond to salt or drought stress by expressing proteins that show sequence homology with well characterized proteins such as DREB, MYB, P5CS, and BADH (Ingram and Bartels 1996; Bray 1997). The expression patterns of salt-inducible genes are complex. The expression of some genes rapidly respond to salt stress, whereas others are slowly induced after the accumulation of salt-resistance proteins. Durner et al. (1997) showed that SA could also induce the expression of resistance-related genes to protect plants from stressful injuries as a defense mechanism against salt stress.
The mechanisms underlying the physiological and molecular responses of plants to SA remain unclear. Therefore, the present study aimed to investigate the effects of exogenous SA application on the growth, metabolism, and expression of defense genes of D. superbus under 0.3, 0.6 and 0.9% salt conditions by analyzing the leaf biomass, anatomy, soluble protein and sugar content, and expression of the MYB and P5CS genes. A better understanding of the mechanism underlying the response of D. superbus to salt stress is vital to the expansion of plantations in other regions.
Materials and methods
Plant materials and experimental procedures
In late March 2015, a pot experiment was conducted in a controlled environmental room at the Zhejiang Institute of Subtropical Crops in Zhejiang Province, China (N28°23′, E120°72′). One-year-old homogenous and healthy D. superbus seedlings were transferred to plastic pots (13.5 cm inner diameter, 15 cm height, with a hole at the bottom) filled with a mixed substrate of humus:peat:soil (3:5:2, v/v/v, pH 6.11). The substrate was loam with a total nitrogen (N) content of 1.78 g kg−1, total phosphorus (P) content of 1.98 g kg−1, total potassium (K) content of 16.63 g kg−1, and an organic matter content of 22.2 g kg−1. Twelve weeks later, a completely randomized design with five replicates per treatment and five plants (plastic pot) per replicate was adopted.
Plants were divided into eight treatments consisting of four salt concentrations (0, 0.3, 0.6, and 0.9% NaCl) and two SA concentrations (0 and 0.5 mM). The eight treatments were as follows: T1 (distilled water), T2 (distilled water with 0.5 mM SA), T3 (0.3% NaCl), T4 (0.3% NaCl with 0.5 mM SA), T5 (0.6% NaCl), T6 (0.6% NaCl with 0.5 mM SA), T7 (0.9% NaCl), and T8 (0.9% NaCl with 0.5 mM SA). The salt solution was gradually added to the substrate in eight steps to achieve the final concentrations of 0.3, 0.6, and 0.9%, respectively. The 0.5 mM SA solution in ethanol was sprayed onto the adaxial and abaxial surfaces of the leaves of D. superbus seedlings twice daily at 7:30 and 18:00 (5 days prior to salinity treatment). At 45 days after treatment, fresh leaves were collected from the five replicates of the same treatment, which were then used in the determination of soluble protein content, SS content, structural microscopy analysis, and expression analysis of salt stress-related genes (MYB and P5CS).
Leaf growth analysis
At 45 days after treatment, the D. superbus were harvested and divided into leaves and roots for growth analysis. The leaves were incubated at 105 °C for 15 min, then dried in an oven at 80 °C until constant weight. The leaves were then weighed on an electronic scale (Tang et al. 2015).
Leaf anatomy analysis
To observe the anatomical structure of the mesophyll of D. superbus, leaf samples were collected from seedlings subjected to different salt treatments, cut into 3-mm segments, and fixed in glutaraldehyde for at least 24 h as described by Deng et al. (2010). The samples were then dehydrated across a graded ethanol series, cleared in xylene, embedded in paraffin wax, and then cut into 8–10 μm thick sections using a rotary microtome (KD2258, Zhejiang, China). The sections were stained with both safranin and Fast Green and then examined under a microscope (Bx41, Olympus Optical Co. Ltd., Tokyo, Japan). The thickness of the lower and upper epidermis, mesophyll, palisade, and spongy layer of the leaves of seedlings subjected to different treatments were measured using an ocular and a stage micrometer.
Soluble protein and SS content analysis
Soluble protein content was measured as described by Bradford (1976) with bovine serum albumin used as standard. For analysis of soluble protein content, each leaf sample (0.3 g) was homogenized with 8 mL of 0.1 M sodium phosphate buffer (pH 7.4). The homogenate was centrifuged at 80,000 rpm for 20 min at 4 °C. The supernatant was collected for measurement of soluble protein, and a mixture of 1 mL of the supernatant and 0.1% Coomassie brilliant blue G250 (5 mL, dissolved in 50% alcohol) was allowed to react for 10 min. The absorbance of the mixture was then measured at a wavelength of 595 nm. SS content was determined using anthrone colorimetry according to the method of Li (2000), with glucose used as the standard. For the measurement of SS content, each leaf sample (0.5 g) was immersed in 15 mL of distilled water and 8 mol L−1 hydrochloric acid (10 mL), which was then heated at 100 °C for 25 min. After rapid cooling, the mixture was centrifuged at 50,000 rpm for 20 min at 4 °C, and then the supernatant was collected. Then, 1 mL of the supernatant was mixed with 4 mL of 2% anthrone reagent (dissolved in sulfuric acid) and boiled for 15 min, and then cooled to room temperature. The absorbance was measured at a wavelength of 620 nm.
Expression analysis of salt-inducible genes
Total RNA from the leaves of D. superbus subjected to different salt treatments was extracted according to Pang et al. (2005), and then reversely transcribed to cDNAs using a cDNA synthesis kit (Toyobo, Osaka, Japan). The expression analysis of salt-inducible genes (MYB and P5CS) with an actin gene as a reference was determined by quantitative real-time PCR (qRT-PCR) with an ABI PRISM 7500 sequence detection system (Applied Biosystems, Foster City, CA, USA) and a SYBR Green PCR MasterMix (Applied Biosystems) and performed in triplicate (Liu et al. 2015) (Table 1 presents primer information used in the PCR amplification of specific genes).
Statistical analysis
Statistical analysis was conducted by one-way ANOVA and Duncan’s multiple range test using the statistical software package SPSS19.0, and the standard error of differences between means was calculated (P ≤ 0.05).
Results
Leaf growth and anatomy
Salt significantly affected the leaf growth of D. superbus (P < 0.05) (Table 2). After 45 days of salt treatment at concentrations of 0.3, 0.6 and 0.9%, the total leaf dry weight per plant decreased by 34.9, 44.1, and 56.7%, respectively, while the leaf fresh weight decreased by 17.3, 35.9, and 38.6% respectively. Salicylic acid treatment increased the leaf dry weight and leaf fresh weight by 13.3 and 14.9% respectively, under 0.3% salt treatment, and by 39.5 and 32.9% under 0.6% salt treatment, whereas no detectable effect was observed with the 0.9% treatment.
Salinity had a marked effect on the anatomy of mesophyll cells of the leaves of D. superbus seedlings (Fig. 1, Table 3). The thickness of mesophyll, palisade tissue, and spongy parenchyma decreased with salinity from 0 to 0.9% concentration, whereas the SA treatment caused a slight increase under the 0.3, 0.6, and 0.9% salt treatments. However, no significant difference in the upper and lower epidermis under different salt and SA treatments was detected.

Effects of SA on the leaf anatomy of D. superbus grown under salt stress. T1 (distilled water), T2 (distilled water with 0.5 mmol SA), T3 (0.3% NaCl), T4 (0.3% NaCl with 0.5 mmol SA), T5 (0.6% NaCl), T6 (0.6% NaCl with 0.5 mmol SA), T7 (0.9% NaCl), and T8 (0.9% NaCl with 0.5 mmol SA)
Soluble protein and SS content
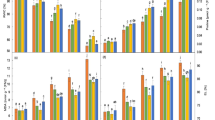
The soluble protein and sugar content in the leaves of D. superbus seedlings significantly (P < 0.05) differed under salt and SA treatments (Fig. 2). We observed a slight decrease in soluble protein content from 0 to 0.9% salt treatment, whereas SA induced a marked increase in soluble protein content by 11.4 and 57% under the 0 and 0.3% salt treatments, respectively. These findings indicate that salt treatment decreases SS content in the leaves of D. superbus and occurs in a dose-dependent pattern. SA treatment increased SS content by 11.5, 28.9, and 8.8% respectively, under 0.3, 0.6 and 0.9% salt treatments.

Effects of SA on soluble protein content (A) and soluble sugar content (B) of D. superbus grown under salt stress (mean ± SD). Different letters indicate significant differences (P < 0.05) based on an LSD test; the same letter indicates no significant differences between the treatments, n = 5
Relative expression of salt-inducible genes
The effect of salinity and SA treatment on the relative gene expression of MYB and P5CS in the leaves of D. superbus was prominent (P < 0.05) and variable (Fig. 3). The variations in the relative expression of MYB and P5CS were similar, such that their relative expression increased with salt treatments of increasing concentrations. The expression of MYB and P5CS under the 0.6% salt condition increased by 22.9 and 70.1%, respectively, compared to T1. Interestingly, the expression of MYB and P5CS under the 0.9% salt treatment increased by 27.2 and 20.4% relative to that of the 0% salt treatment (T1), but showed a minimal decline compared to that of the 0.6% salt treatment. However, SA treatment caused a slight increase in the expressions of MYB and P5CS under the 0.3, 0.6, and 0.9% salt treatments.

Effects of SA on the relative expression of the MYB (A) and P5CS (B) genes in D. superbus grown under salt stress (mean ± SD). Different letters indicate significant differences (P < 0.05) based on an LSD test; the same letter indicates no significant differences between treatments, n = 5
Discussion
The biosynthesis, accumulation, and transport of plant nutrients generally occur in the leaves. Therefore, the leaves are also the most vulnerable plant parts that would immediately respond to biotic and abiotic stresses. A previous study has shown that the leaf biomass of plants grown under water deficit, chilling, salt, and deep shade conditions is suppressed (Larcher 2003). In the present study, the leaf dry weight and leaf fresh weight of D. superbus seedlings decreased with increasing salt concentrations, which was similar to the findings of Li et al. (2014), that showed salt stress causes a water deficit and metabolic disorder in leaves that in turn leads to growth inhibition. Studies have shown that exogenously spraying salicylic acid alleviates salt stress-induced damages by inducing a set of physiological, biochemical, and molecular adaptation responses (Bandurska and Cieślak 2012). In the present study, we found that exogenously spraying SA onto the leaves of D. superbus resulted in a marked increased in leaf biomass under 0.3 and 0.6% salt stress, which then supports the notion that SA plays a positive role in regulating the response of plants to salt stress (Senaratna et al. 2000). However, this study also determined that SA imparted minimal effects on modulating plant defense when the D. superbus were subjected to the severe salt stress (for example, 0.9% salt concentration).
Salt stress causes leaf succulence as well as damages the anatomical features of leaves (Parida et al. 2004). In the present study, a marked decrease in mesophyll thickness was observed in the leaves of D. superbus that were subjected to salt stress from 0% to 0.9%, which was due to the decrease in length of palisade cells and layer of spongy tissue (Fig. 1, Table 3). Our observation was in agreement with the findings of Parida et al. (2004), who reported that changes in leaf anatomy affect CO2 diffusion. Furthermore, salt stress induces a decrease in mesophyll thickness that may result in a decrease in the conductance and photosynthesis in the leaves of bean plants (Brugnoli and Bjorkman 1992). The observed improvement in the growth of D. superbus seedlings under 0.3 and 0.6% salt stress after SA treatment may be a consequence of an increase in the number of palisade and sponge cells that promote CO2 diffusion and thereby enhancing photosynthesis (Table 3).
Soluble protein content is an important indicator of the physiological status of plants (Doganlar et al. 2010). In the present study, soluble protein content decreased in response to salt treatment in a dose-dependent pattern. However, compared to the non-SA-treated seedlings, the SA-treated seedlings under salt stress exhibited a slower increase in soluble protein content. This observation indicated that salt stress results in a metabolic disorder in the leaves of D. superbus, and SA treatment may contribute to a more efficient metabolic response in plants (Popova et al. 2009). Similar results were reported in salt-tolerant sunflower and finger millet plants (Parvaiz and Satyavati. 2008). SS, which is an important carbon source for plant growth, reflects the nutritional status of plants (Sasaki et al. 1998; Dong et al. 2011). Salt stress induced a marked decrease in SS content in the leaves of D. superbus, as well as in other plants (Miller et al. 1989; Sasaki et al. 1998), which indicates that salt stress decreases the rate of carbohydrate metabolism in plants. However, compared to the non-SA-treated plants, those treated with SA showed an increase in SS content. This finding thereby confirms the hypothesis that SA application in plants alleviates salt stress-induced decrease in carbohydrate metabolism, thus improving their nutrient content.
In general, plants respond to stresses with transient changes in the expression of related genes, which encode proteins that impart protective effects on plants (Estruch 2000). The MYB gene is involved in osmotic and salt stress in moso bamboo and participates in the biosynthesis pathway of ABA, which protects plants from stress damage (Fujita et al. 2011; Peng et al. 2013). The P5CS gene has been positively correlated with the biosynthesis and accumulation of proline in Arabidopsis and wheat (Hayashi et al. 2000). In the present study, we detected the MYB and P5CS genes in the leaves of D. superbus and observed that salt stress induces the upregulation of the MYB and P5CS genes compared to that in non-salt-treated plants, whereas plants treated with SA showed the highest levels of MYB and P5CS expression. These findings were similar to that observed in Arabidopsis (Qin et al. 2012; Zhu et al. 1998), indicating that the expression of the MYB and P5CS genes are closely related to salt tolerance, and thus play an important role in plant defense against salt stress. Moreover, salicylic acid alleviates the damages that were incurred in D. superbus due to salt stress by activating the expression of salt stress-related genes, thereby enhancing stress resistance.
In conclusion, D. superbus subjected to salt stress conditions resulted in a decrease in leaf thickness, plant growth, and primary metabolite contents together with an increase in the expression of salt-induced genes. Salicylic acid could efficiently prevent the adverse effects of salt stress on the growth and physiology of D. superbus by increasing the thickness of the mesophyll layer and inducing the expression of salt-induced genes to resist higher salinity levels and to improve the metabolic level of the plants. This information facilitates the better understanding how SA regulates the responses of plants to salt stress, particularly in horticulture plants.
References
Bandurska H, Cieślak M (2012) The interactive effect of water deficit and UV-B radiation on salicylic acid accumulation in barley roots and leaves. Environ Exp Bot 94:9–18. https://doi.org/10.1016/j.envexpbot.2012.03.001
Bittrich V (1993) Caryophyllaceae. In: Kubitzki K, Rohwer JG, Bittrich V (eds) Families and genera of flowering plants, vol 2 flowering plants, dicotyledons. Springer, Berlin, pp 206–236
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1–2):248–254. https://doi.org/10.1016/0003-2697(76)90527-3
Bradford KJ, Hsiao TC (1982) Physiological responses to moderate water stress. In: Lange OL, Nobel PS, Osmond CB, Ziegler H (eds) Physiological plant ecology II. Water relations and carbon assimilation. Encyl. plant physiol. new series, vol 12. Springer, Berlin, pp 263–324. https://doi.org/10.1007/978-3-642-68150-9_10
Bray EA (1997) Plant responses to water deficit. Trends Plant Sci 2:4–54. https://doi.org/10.1371/journal.pone.0082634
Brugnoli E, Bjorkman O (1992) Growth of cotton under continuous salinity stress: influence on allocation pattern, stomatal and non-stomatal components of photosynthesis and dissipation of excess light energy. Planta 187:335–347. https://doi.org/10.1007/BF00195657
David JIN, Park SN (1979) Salinity effects on leaf anatomy. Plant Physiol 63:700–703
Deng YM, Chen SM, Teng NJ, Chen FD, Li FT, Song AP, Guan ZY (2010) Flower morphologic anatomy and embryological characteristics in Chrysanthemum multicaule (Asteraceae). Sci Hortic 124:500–505. https://doi.org/10.1016/j.scienta.2010.02.009
Doganlar ZB, Demir K, Basak H et al (2010) Effects of salt stress on pigment and total soluble protein contents of three different tomato cultivars. Afr J Agric Res 5(15):2056–2065. https://doi.org/10.5897/AJAR10.258
Dong CJ, Wang XL, Shang QM (2011) Salicylic acid regulates sugar metabolism that confers tolerance to salinity stress in cucumber seedlings. Sci Hortic 129(4):629–636. https://doi.org/10.1016/j.scienta.2011.05.005
Durner J, Shah J, Klessig DF (1997) Salicylic acid and disease resistance in plants. Trends Plant Sci 2(7):266–274. https://doi.org/10.1080/07352689991309397
Estruch F (2000) Stress-controlled transcription factors, stress-induced genes and stress tolerance in budding yeast. FEMS Microbiol Rev 24:469–486. https://doi.org/10.1111/j.1574-6976.2000.tb00551.x
Fujita Y, Fujita M, Shinozaki K et al (2011) ABA-mediated transcriptional regulation in response to osmotic stress in plants. J Plant Res 124(4):509–525. https://doi.org/10.1007/s10265-011-0412-3
Gunes A, Inal A, Alpaslan M et al (2005) Effects of exogenously applied salicylic acid on the induction of multiple stress tolerance and mineral nutrition in maize (Zea mays L.) (Einfluss einer Salicylsäure-Applikation auf die Induktion von Stresstoleranz sowie Nährstoffaufnahme von Mais [Zea mays L.]). Arch Agron Soil Sci 51(6):687–695. https://doi.org/10.1080/03650340500336075
Hamayun M, Khan SA, Khan AL et al (2010) Exogenous gibberellic acid reprograms soybean to higher growth and salt stress tolerance. J Agric Food Chem 58(12):7226–7232. https://doi.org/10.1021/jf101221t
Hayashi F, Ichino T, Osanai M et al (2000) Oscillation and regulation of proline content by P5CS and ProDH gene expressions in the light/dark cycles in Arabidopsis thaliana L. Plant Cell Physiol 41(10):1096–1101. https://doi.org/10.1093/pcp/pcd036
Hooftman DAP, van Kleunen M, Diemer M (2003) Effects of habitat fragmentation on the fitness of two common wetland species, Carex davalliana and Succisa pratensis. Oecologia 134(3):350–359. https://doi.org/10.1007/s00442-002-1096-0
Ingram J, Bartels D (1996) The molecular basis of dehydration tolerance in plants. Annu Rev Plant Physiol Plant Mol Biol 47:377–403
Iqbal M, Ashraf M, Jamil A, Rehman S (2006) Does seed priming induce changes in the levels of some endogenous plant hormones in hexaploid wheat plants under salt stress? J Integr Plant Biol 48:181–189. https://doi.org/10.1111/j.1744-7909.2006.00181.x
Ivanitskaya EF (1962) Specific characteristics of the anatomical structure of plants under various soil salinity conditions. Fiziologiya Rastenii 9:199–209
Jayakannan M, Bose J, Babourina O et al (2013) Salicylic acid improves salinity tolerance in Arabidopsis by restoring membrane potential and preventing salt-induced K+ loss via a GORK channel. J Exp Bot. https://doi.org/10.1093/jxb/ert085
Khan MIR, Iqbal N, Masood A et al (2013) Salicylic acid alleviates adverse effects of heat stress on photosynthesis through changes in proline production and ethylene formation. Plant Signal & Behav 8(11):e26374. https://doi.org/10.4161/psb.26374
Khedr AHA, Abbas MA, Wahid AAA, Quick WP, Abogadallah GM (2003) Proline induces the expression of salt stress responsive proteins and may improve the adaptation of Pancratum maritimum L., to salt stress. J Exp Bot 54:2553–2562. https://doi.org/10.1093/jxb/erg277
Khodary SEA (2004) Effect of salicylic acid on the growth, photosynthesis and carbohydrate metabolism in salt-stressed maize plants. Int J Agric Biol 6(1):5–8
Larcher W (2003) Physiological plant ecology: ecophysiology and stress physiology of functional groups. Springer Science & Business Media, Berlin
Li HS (2000) Principles and techniques of plant physiological biochemical experiment. Higher Education Press, Beijing, pp 194–197
Li W, Liu X, Khan MA et al (2005) The effect of plant growth regulators, nitric oxide, nitrate, nitrite and light on the germination of dimorphic seeds of Suaeda salsa under saline conditions. J Plant Res 118(3):207–214. https://doi.org/10.1007/s10265-005-0212-8
Li T, Hu Y, Du X et al (2014) Salicylic acid alleviates the adverse effects of salt stress in Torreya grandis cv. Merrillii seedlings by activating photosynthesis and enhancing antioxidant systems. PLoS ONE 9(10):e109492. https://doi.org/10.1371/journal.pone.0109492
Liu Y, Song L, Yu W et al (2015) Light quality modifies camptothecin production and gene expression of biosynthesis in Camptotheca acuminata Decne seedlings. Ind Crops Prod 66:137–143. https://doi.org/10.1016/j.indcrop.2014.12.046
Ma X, Zheng J, Zhang X, Hu Q, Qian R (2017) Salicylic acid alleviates the adverse effects of salt stress on dianthus superbus (Caryophyllaceae) by activating photosynthesis, protecting morphological structure, and enhancing the antioxidant system. Front Plant Sci 8:600. https://doi.org/10.3389/fpls.2017.00600
Miller JE, Patterson RP, Pursley WA, Heagle AS, Heck WW (1989) Response of soluble sugars and starch in field-grown cotton to ozone, water stress, and their combination. Environ Exp Bot 29:477–486. https://doi.org/10.1016/0098-8472(89)90026-9
Opdekamp W, Beauchard O, Backx H, Fran-ken F, Cox TJS, van Diggelen R et al (2012) Effects of mowing cessation and hydrology on plant trait distribution in natural fen Meadows. Acta Oecol 2012(39):117–127. https://doi.org/10.1016/j.actao.2012.01.011
Pang YZ, Shen GA, Wu WS, Liu XF, Lin J, Tan F, Sun XF, Tang KX (2005) Characterization and expression of chalcone synthase gene from Ginkgo biloba. Plant Sci 168:1525–1531. https://doi.org/10.1016/j.plantsci.2005.02.003
Parida AK, Das AB, Mittra B (2004) Effects of salt on growth, ion accumulation, photosynthesis and leaf anatomy of the mangrove, Bruguiera parviflora. Trees 18(2):167–174. https://doi.org/10.1007/s00468-003-0293-8
Parvaiz A, Satyavati S (2008) Salt stress and phyto-biochemical responses of plants—a review. Plant Soil Environ 54:89–99
Peng Z, Zhang C, Zhang Y et al (2013) Transcriptome sequencing and analysis of the fast growing shoots of moso bamboo (Phyllostachys edulis). PLoS ONE 8(11):e78944. https://doi.org/10.1371/journal.pone.0078944
Popova LP, Maslenkova LT, Yordanova RY et al (2009) Exogenous treatment with salicylic acid attenuates cadmium toxicity in pea seedlings. Plant Physiol Biochem 47(3):224–231. https://doi.org/10.1016/j.plaphy.2008.11.007
Qin Y, Wang M, Tian Y et al (2012) Over-expression of TaMYB33 encoding a novel wheat MYB transcription factor increases salt and drought tolerance in Arabidopsis. Mol Biol Rep 39(6):7183–7192. https://doi.org/10.1007/s11033-012-1550-y
Rosenthal G (2010) Secondary succession in a fallow central European wet grassland. Flora. 205(3):153–160. https://doi.org/10.1016/j.flora.2009.02.003
Roussos PA, Gasparatos D, Tsantili E, Pontikis CA (2007) Mineral nutrition of jojoba explants in vitro under sodium chloride salinity. Sci Hortic 114:59–66. https://doi.org/10.1016/j.scienta.2007.05.001
Roussos P, Gasparatos D, Kyriakou C, Tsichli K, Tsantili E, Haidouti C (2013) Growth, nutrient status and biochemical changes in sour orange (Citrus aurantium L.) plants subjected to sodium chloride stress. Cοmmun Soil Sci Plant Anal 44:805–816. https://doi.org/10.1080/00103624.2013.749438
Saleh B (2012) Salt stress alters physiological indicators in cotton (Gossypium hirsutum L.). Soil Environ 31(2):113–118
Sasaki H, Ichimura K, Okada K, Oda M (1998) Freezing tolerance and soluble sugar contents affected by water stress during cold-acclimation and de-acclimation in cabbage seedlings. Sci Hortic 76:161–169. https://doi.org/10.1016/S0304-4238(98)00143-5
Senaratna T, Touchell D, Bunn E, Dixon K (2000) Acetyl salicylic acid (aspirin) and salicylic acid induce multiple stress tolerance in bean and tomato plants. Plant Growth Regul 30:157–161. https://doi.org/10.1023/A:1006386800974
Soussi M, Ocana A, Lluch C (1998) Effects of salt stress on growth, photosynthesis and nitrogen fixation in chick-pea (Cicer arietinum L.). J Exp Bot 49(325):1329–1337
Tang H, Hu YY, Yu WW, Song LL, Wu JS (2015) Growth, photosynthetic and physiological responses of Torreya grandis seedlings to varied light environments. Trees 29(4):1011–1022. https://doi.org/10.1007/s00468-015-1180-9
Wani AB, Chadar H, Wani AH et al (2016) Salicylic acid to decrease plant stress. Environ Chem Lett. https://doi.org/10.1007/s10311-016-0584-0
Yildirim E, Turan M, Guvenc I (2008) Effect of foliar salicylic acid applications on growth, chlorophyll, and mineral content of cucumber grown under salt stress. J Plant Nutr 31(3):593–612. https://doi.org/10.1080/01904160801895118
Zhu JK, Liu J, Xiong L (1998) Genetic analysis of salt tolerance in Arabidopsis: evidence for a critical role of potassium nutrition. Plant Cell 10(7):1181–1191
Acknowledgements
The Zhejiang Province Wenzhou Science and Technology Plan Projects (N20140021) and The Zhejiang ProvinceWenzhou Demonstration Generalization Projects of Four New Science and Technology of Forestry (WZHX2016-11-236) supported this study. The authors also thank Accdon for its linguistic assistance during the preparation of this manuscript.
Funding
Funding was provided by The Zhejiang Province Wenzhou Science and Technology Plan Projects (N20140021) and The Zhejiang Province Wenzhou demonstration generalization projects of four new science and technology of forestry (WZHX2016-11-236).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Zheng, J., Ma, X., Zhang, X. et al. Salicylic acid promotes plant growth and salt-related gene expression in Dianthus superbus L. (Caryophyllaceae) grown under different salt stress conditions. Physiol Mol Biol Plants 24, 231–238 (2018). https://doi.org/10.1007/s12298-017-0496-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12298-017-0496-x