
Abstract
Urbanization and human-led development have increased more rapidly along shorelines and in coastal watersheds than inland regions over the past century. The result of major land use changes for both urban tracts and agriculture to serve the urban areas, as well as infrastructure development is increased runoff carrying sediments, nutrients, pollutants, pharmaceuticals, and toxins downstream to estuarine systems. The increased runoff levels are only the tip of the iceberg, with human development resulting in increased fecal bacteria from urbanization and excess nutrients from agriculture leading to harmful algal blooms. Estuaries act as a natural filter between land and sea, but have been overloaded by the influx of sediments and pollutants in recent decades. As a result, there have been a variety of impacts to estuarine ecosystems and water quality including increased sediment load, eutrophication, harmful algal blooms, fecal bacteria, as well as shellfish and fisheries declines. In some estuarine systems, the reduction in light penetration to the benthos has led to the loss of seagrasses. In others, seasonal hypoxia is a visible symptom of prolonged eutrophication. There is a need to augment long-term monitoring techniques with new technologies and data processing methods to better understand the current state of estuaries and work towards mitigating human impacts on estuarine ecosystems and water quality.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Urbanization is transforming the world’s coastlines on timescales of years to decades. Human population centers tend to be near rivers and coastlines. In 2010, 123.3 million people, or 39% of the US population, lived in counties directly on the shoreline (Fig. 1). This population is expected to increase by about 10 million people, or 8%, from 2010 to 2020 (https://oceanservice.noaa.gov/facts/population.html). While less than 20% of land in the USA is in a coastal watershed county, these counties are home to more than half of the population (53%). From 1970 to 2010, coastal shoreline counties added 125 people per square mile, over three times the national average of 39 people per square mile (http://stateofthecoast.noaa.gov). As human populations continue to grow and urbanization escalates in coastal and watershed counties, waterways and aquatic ecosystems are often adversely affected. Estuaries, the natural buffer zone between rivers and the ocean, are among the most impacted waterways and aquatic ecosystems. Estuaries are particularly vulnerable because they often have a large watershed, much greater than their own area. Impacts of urbanization on estuaries include increases in sediment, nutrient, and fecal microbial loading and subsequent shifts in water quality, potentially leading to harmful algal blooms; changes in streamflow and salinity; and the subsequent effects of these environmental shifts on plankton, marshes, seagrasses, shellfish, and fish within the estuary (Dauer et al. 2000; Mallin et al. 2000; Alberti 2005; Carle et al. 2005; Campos and Cachola 2007; Paerl et al. 2014; Lemley et al. 2018a).

US population density by county in 2010 (a) and population change by county from 1990 to 2000 (b), courtesy US Census Bureau. Note higher population densities in coastal counties as well as more rapid population growth along coasts and in watersheds in the twenty-first century
Water quality further impacts quality of life for humans living near estuaries as events like harmful algal blooms and high turbidity may limit recreational use of the waters, and could incur an economic cost as well. Despite the value of the ecosystem services provided by estuaries, most urban design planning takes little account for neighboring estuarine waterways. Rapid urbanization in the past century has put immense stress on these systems that has resulted in sometimes dramatic shifts in water quality and fish/shellfish stocks, with some oyster grounds completely extirpated due to microbial pathogens and historical levels a distant memory (Figs. 2 and 3). Those shellfish beds that are still surviving are often closed to fishery collections due to microbial pathogens. It is thus critical to understand the specific effects of urbanization on estuarine waterways and ecosystems to mitigate the potential harm caused by this process. This is more imperative now than ever given many estuaries are now beyond their capacity to assimilate or process these inputs, although that the same pollutants, excess nutrient loads, and toxic materials make their way downstream to the oceans.

Oyster ground extent in US estuaries historically (a) and presently (b). Note significant decline throughout US coastal and estuarine regions regardless of latitude or coast (Zu Ermgassen et al. 2012, reprint of open access material)

Percentage of estuary area containing oyster grounds historically and present by US coast line (Zu Ermgassen et al. 2012, reprint of open access material)
Many estuaries have monitoring systems in place in response to the stress of human impacts and the intrinsic value of these systems. In 1987, the Environmental Protection Agency (EPA) National Estuary Program recognized 28 estuaries of national significance along the US Atlantic, Gulf, and Pacific coasts and in Puerto Rico (https://www.epa.gov/nep). Several years later, the National Oceanographic and Atmospheric Administration (NOAA) National Estuarine Research Reserve System, a network of 29 coastal sites, was designated to protect estuarine systems (https://coast.noaa.gov/nerrs/; Voulgaris and Meyers 2004). Globally, marine protected areas such as these serve to protect estuarine systems as well as monitor flora, fauna, and water quality (http://www.mpatlas.org/; Washburn and Sanger 2011). While monitoring is critical for understanding changes over time, methods utilized today have changed little over the past decades. Likely due to their proximity to human population centers, many estuaries have had the luxury of scientific monitoring for decades including measurements of nutrients and suspended sediment loads. These historic data provide important baseline information for studying changes to estuaries over time. The benefit is that multi-decadal stages are being published examining ecosystems (e.g., Thronson and Quigg 2008; Paerl et al. 2014; Steichen and Quigg 2018) which reveal the prevailing consequences of long-term land use changes on water quality, including its degradation.
Given the high flux volumes and rapid transformations occurring in estuaries worldwide, there is a pressing need to incorporate modern technologies and methods alongside traditional monitoring to improve our understanding of estuaries and human impacts on water quality and estuarine ecosystems. Here, several direct impacts of urbanization on estuarine ecosystems and water quality are discussed. The breadth of topics included is representative of the wide range of human impacts, many of which are not immediately apparent to the individuals living in population centers adjacent to the coast. Effects are potentially only perceivable downstream or after a time delay, making it even more important to understand adverse effects of human impacts and urban development on our natural buffer zones so that preventative action and mitigation strategies can be enacted.
Sediment Dynamics in Tidal Creek Systems
Tidal creeks act as a gateway between uplands and marshes to estuaries and coastal waters. These smaller water bodies are influenced by both freshwater flows running off the landscape and continuous tidally driven changes in salinity (Holland and Sanger 2008). The upland areas of these watersheds are often popular locations for development due to the adjacent coastal landscape. These ecosystems are important habitat for commercial and recreation fishes, including seatrout, crevalle jack, flounder, spadefish, spot, black drum, blue crab, brown shrimp, and white shrimp (Hackney et al. 1976; Shenker and Dean 1979; Kneib 1997; Beck et al. 2001). Tidal creeks act as a gateway of sediments, nutrients, and water to and from adjacent wetlands. The wetlands serve vital roles for ecosystems, act as niche habitats for organisms, act as an initial “filter” of water from the landscape, and provide coastal protection from storms through wave attenuation (Daborn et al. 1993; Goodwin et al. 2001; Granek et al. 2009; Townend et al. 2011; Shi et al. 2012; Möller et al. 2014). The physical and biological dynamics of tidal creek and wetland ecosystems are intricately connected. Changes that occur at that landward (e.g., stormwater runoff, shoreline hardening, land clearing) and seaward (e.g., trawling, sea level change) ends of tidal systems may alter the resilience of these habitats in the future. Rapid economic development and associated land use change in these coastal areas can increase point and nonpoint source pollution loading into these tidal creeks and nearby estuaries, influencing productivity, biodiversity, and ecological functioning of these systems (Vitousek et al. 1997). Previous research on tidal creek systems has focused primarily on freshwater discharge, water quality, macrobenthic communities, and biodiversity (Voulgaris and Meyers 2004; Corbett et al. 2007; Sanger et al. 2008; Corbett et al. 2009; DiDonato et al. 2009; Washburn and Sanger 2011; Gleeson et al. 2013; Webster et al. 2013; Sanger et al. 2015). There has been much less focus on the response of sediment dynamics associated with land use change, particularly development, in these critical coastal ecosystems.
Suspended sediment is cited as the most prolific water pollutant (USEPA 2004, 2006) and therefore studying sediment dynamics within coastal tributaries and estuaries is critical to better management. It is commonly found that sediments introduced to coastal systems are chemically and physically bound with terrestrially based nutrients and other sediment associated pollutants (e.g., heavy and trace-metals, herbicides, pesticides) and ultimate degradation of these water bodies (Schropp et al. 1990; Riggs et al. 1991; Phillips 1997; Paerl et al. 1998; Hyland et al. 2000; Buzzelli et al. 2001; Paerl 2006;). In shallow coastal tributaries and estuaries, there is continuous exchange and deposition of particle-reactive materials between the water column and bed sediment (Giffin and Corbett 2003). Because of this, there is a need to study the sediment dynamics of these urbanized estuarine systems if we are to better understand their function as sediment sources or sinks, as well as their sedimentological characteristics. However, only focusing on the sediments does not provide insight into the drivers that lead to the ultimate accumulation of material. Additional focus on the biogeochemical transformations of the material along its route to deposition/accumulation would better link the sediments to the biological community and productivity. Other studies have shown the impact of development in tidal creek watersheds to detrimental changes in the benthic biological community (Lerberg et al. 2000; Holland et al. 2004). Linking sediment delivery, quality, and dynamics to land use change and biological diversity/productivity would be a next logical step in better managing these systems.
Remote Sensing of Suspended Sediment Concentrations
Estuaries have long been considered a “filter” for suspended sediments and material bound to those sediments such as pollutants and organic debris (Schubel and Carter 1984). Classically suspended sediment study was limited case studies using water samples and cores. Satellite and aerial imaging of waterways provides a means of rapidly filling spatiotemporal gaps in traditional sampling of suspended sediment concentration; however, it does so at the cost of spatial resolution. Satellite-based instruments image a particular scene on a daily to weekly timescale, with a spatial resolution of 15–250 m. Studies combining field data collection of water samples with remote sensing imagery evaluation have found a high degree of accuracy even in coarse spatial resolution (250 m) MODIS imagery (Miller and McKee 2004). The broad spatial information offered by remote sensing is critical for examining sediment loads in estuarine systems as transport may occur on scales of 1000s of kilometers (e.g., Van Rijn 1993; Miller and McKee 2004). Use of remotely sensed imagery also allowed assessment of a large spatial scale on prolonged time scales (20+ years) from historical records (e.g., Zhou et al. 2017; Wang et al. 2018).
Many studies have explored the relationship between remote sensing reflectance at the sea surface from an atmospherically corrected satellite image and suspended sediment concentration, as early as the 1980s (Stumpf and Pennock 1989). There is an exponential relationship between these quantities, particularly at mid-range sediment concentrations from about 10 to 30 mg/L (Freeman et al. 2017). A wide variety of algorithms have been developed to link these two quantities using in situ data alongside concurrently collected satellite or aerial imagery. It is critical to carefully assess the conditions under which an algorithm was developed prior to applying it to a new imagery dataset. Many field-based algorithms are linear and are most applicable to the conditions under which they were developed, and will not perform well for regions with much higher or lower concentrations of suspended sediment (Freeman et al. 2017) (Fig. 4). That said, there is immense information contained in remotely sensed imagery for studies of suspended sediment loads in estuarine systems, or related material such as organic matter, chlorophyll, and pollutants which bind to sediments. Studies combining in situ samples with remotely sensed imagery provide the highest degree of accuracy and spatial extent; however, enough ground work has been laid to date to extrapolate meaningful information about sediment concentration from satellite imagery for nearly any estuarine system.

Comparison of suspended particulate matter (SPM) measured in situ from water samples in a variety of water types with calculated SPM utilizing at surface reflectance from a handheld radiometer and a suite of algorithms relating reflectance to sediment load. Note that there is increased spread between calculated values at higher sediment concentrations, and that each algorithm has a “niche range” in which it performs well. (Freeman et al. 2017)
While earlier studies focused on validating relationships between remote sensing reflectance and suspended sediment concentration, the abundance and improved resolution of remotely sensed imagery has allowed for more detailed studies. Mixing over distance is now being studied remotely and on a larger scale thanks to Landsat imagery (Umar et al. 2018). Interplay of different physical forcing from tides, waves, and seasonal events such as monsoon are now being tied to sediment load with a high level of fidelity utilizing a combination of geostationary and mobile platforms (Zhou et al. 2017). Furthermore, remote sensing has opened a window to understand variability of different physical influences (such as freshwater inflow and wave events) on regions of clear and turbid water in both shallow and deep water (Ruhl et al. 2001). Established relationships tend to link one band (usually red or infrared) of remote sensing reflectance to suspended sediment concentration. Modest performance increase has been obtained using multi-band algorithms for scenarios where a higher degree of accuracy may be necessary (DeLuca et al. 2018).
Harmful Algal Blooms—Insight from a South African Case Studies
The increased global incidence of harmful algal bloom (HAB) events is closely linked to the urbanization of coastal areas resulting in intensification of anthropogenic nutrient loading (Paerl and Scott 2010; Havens and Paerl 2015). The primary concern arising from such HAB events is founded in the array of possible consequences. Some of these include oxygen depletion related to bloom decay processes (i.e., respiration), direct toxic effects on higher trophic levels (i.e., shellfish poisoning and bioaccumulation), mechanical interference and suffocation of faunal communities (e.g., mucilage production), and habitat destruction through shading of submerged aquatic vegetation. Further complicating the issue of HABs are the variety of mechanisms employed by species—both microalgae and protozoans—which enable their proliferation. Therefore, global issues, such as the accelerated distribution, frequency, magnitude, and variety of HABs, “require the interfacing of phenomena that occur on very different scales of space, time and ecological organization” (Levin 1992).
The dynamics of HABs, and phytoplankton communities in general, are controlled by both local processes and physical transport processes (Qin and Shen 2017). A combination of bottom-up (e.g., nutrient supply, light availability, temperature) and top-down (e.g., grazing pressure and interspecies competition) controls constitute the local processes structuring these communities. Alternatively, external transport processes (e.g., tidal exchange, freshwater discharge and wind-driven circulation) serve as the physical forces driving observed patterns. Therefore, given the plasticity regarding the periodic fluctuations for each of these processes (hourly, daily, seasonally, annually, or stochastic), the relative importance of local and external transport drivers varies with timescales. As such, gaining an understanding of external and internal processes governing spatio-temporal patterns of phytoplankton communities allows for natural progressions to be discerned from anthropogenically induced change (Hall et al. 2015). Detailed site-specific investigations of HABs are thus required to explain the diversity of patterns at multiple temporal scales and to understand the ecological significance thereof (Cloern and Jassby 2010).
Given the significant economic and social development opportunities provided by aquatic ecosystems, the prevention of continued urbanization and expansion of catchment activities is unlikely (Satterthwaite et al. 2010), thus necessitating an adaptive and proactive approach to conservation of these ecosystems. Potential management options geared at restoring a degree of natural phytoplankton functionality at an annual or seasonal scale are primarily catchment based, while those aimed at disrupting short-term processes can be implemented at the catchment scale or in situ. The Zandvlei (Cape Town) and Swartkops (Port Elizabeth) estuaries provide two South African examples of where increased urbanization has culminated in eutrophic conditions and the proliferation of HABs. The temporarily open/closed Zandvlei Estuary is subject to numerous impacts associated with urbanization, including catchment hardening (increased runoff), river canalization, dredging, habitat loss, flow obstruction (weir and causeway construction), and elevated nutrient inputs. The impacted nature of the system, low salinity environments (< 10), and the propensity for closed mouth conditions have facilitated periodic HAB proliferations of Prymnesium parvum (Prymnesiophyceae) since the 1970s (Whitfield et al. 2016). In 2012, an extensive bloom of P. parvum significantly discolored surface waters and resulted in oxygen depletion and mass fish mortalities within the estuary. Subsequently, an effective adaptive management approach geared at preventing such incidents, together with continuous monitoring, has been adopted by the local authorities. Current management interventions include submerged macrophyte harvesting (maintain nutrient uptake capacity), mouth manipulation (increased salinity variability), and dredging (improved connectivity).
Unlike the Zandvlei Estuary, management of the permanently open and eutrophic Swartkops Estuary is more complex given its size and multiplicity of anthropogenic activities concentrated in the lower reaches of its catchment, including industrial, wastewater treatment plants (WWTP), and stormwater inputs (Scharler and Baird 2003; Lemley et al. 2017a). Of these, the WWTP inputs have the most notable impacts, with the effects being twofold, i.e., consistent daily inputs of freshwater and inorganic nutrients. Therefore, because the probability of reducing discharge rates from the WWTPs is unlikely, due to the high demand placed on these facilities, efforts should be focused on reducing the high N and P loading. One way in which this can be achieved is through the adoption of a dual-nutrient reduction approach to wastewater treatment (Conley et al. 2009). Interestingly, the high inorganic P levels in the Swartkops Estuary currently serve to skew inorganic N:P molar ratios which in turn constrain the magnitude of phytoplankton blooms through N-limitation. In saying this, HABs of species belonging to the Peridinium genus (Dinophyceae) have been documented to occur in the stratified, occasionally hypoxic, upper reaches of the estuary (Lemley et al. 2017a). Therefore, exacerbated eutrophication symptoms (e.g., HABs) could be expected in the estuary by only reducing P loads.
Thorough assessments of HAB dynamics in South African estuaries have been largely understudied, with the Sundays Estuary providing the only example in the literature where this has been addressed. Linked to the urban demand for agricultural products (Satterthwaite et al. 2010), accelerated rates of urbanization are central to the expansion of agricultural activities in the Sundays Estuary catchment. An early study in the late 1980s (Hilmer and Bate 1991) was the first to document the occurrence of two HAB species, namely Heterocapsa rotundata (Dinophyceae) (formerly Katodinium rotundatum) and Heterosigma akashiwo (Raphidophyceae). As such, a recent review of microalgae as indicators in South African estuaries (Lemley et al. 2016) highlighted—among others—the need for (1) “hypothesis-driven,” fine-scale experimental research, and (2) comprehensive autecological studies of HAB species. Geared at addressing these knowledge gaps, an intensive 3-year sampling program was initiated in the Sundays Estuary. Annual (Lemley et al. 2017a) and seasonal (Lemley et al. 2017b) studies verified the persistently eutrophic condition of the estuary and, more specifically, the progressive and recurrent nature of extensive HABs of H. akashiwo and H. rotundata (> 550 μg Chl-a/L) occurring in spring/summer and winter, respectively. The Sundays Estuary provided an ideal ecosystem to delve into the short-term processes (daily and hourly studies) driving phytoplankton dynamics, due to its highly regulated (interbasin water transfer scheme and irrigation canal network) and seasonal (temperature profiles, nutrient inputs and phytoplankton dynamics) nature. These investigations (Lemley et al. 2018a, b) identified seasonal bottom-water hypoxia trends to be a direct consequence of collapsing H. akashiwo spring/summer HABs. Additionally, inorganic nutrient availability (nitrate and phosphate) was identified as the key bottom-up control promoting biomass accumulation, while also limiting the magnitude and duration of HAB events, while a vertically stratified water column concomitant with mesohaline surface waters (~ 10) were identified as conditions promoting the success of observed HAB species (H. akashiwo, H. rotundata, Mesodinium rubrum, and Karenia cf. mikimotoi). The significance of internal biotic processes was revealed during the hourly investigation, with the plasticity of diel vertical migration behavior, reliance of M. rubrum on suitable cryptophyte “prey” resources, and the allelopathic effects of H. akashiwo on co-occurring taxa explaining phytoplankton community dynamics beyond the influence of physicochemical variability. These findings provided insight regarding the ecology of HAB taxa and how they have adapted to thrive in anthropogenically manipulated environments. As such, the broad- to fine-scale information obtained for the Sundays Estuary (Fig. 5) offered the opportunity to suggest management options specifically geared at mitigating against the occurrence of HAB proliferations in similar ecosystems, including urbanized systems. Potential management actions during HAB periods include mechanical harvesting and algal flocculation techniques as a means of biomass removal (Dai et al. 2015). However, many HAB species are well-adapted to persisting during unfavorable conditions, e.g., alternating benthic-pelagic life histories and mixotrophic capabilities. Therefore, the most feasible approach regarding the management of HABs in estuaries is to implement measures at the catchment scale (e.g., beneficial management practices, recycling of wastewater, and scheduled dam releases). These will serve to increase the incidence of episodic disturbances and reduce the extent of press disturbances, with the ultimate objective of (1) maintaining a state of flux in the phytoplankton community, (2) limiting the frequency and magnitude of HAB events, and (3) disrupting the seasonal nature of bottom-water hypoxia (Fig. 6).

Sundays Estuary region discussed in the harmful algal bloom South Africa case study

Summary of the processes facilitating the proliferation and recurrent nature of two commonly occurring harmful algal bloom species in the Sundays Estuary (note: the factors promoting and limiting the growth of these two species are indicated in green and red text, respectively)
The persistent occurrence of harmful phytoplankton taxa is a new feature in South African estuaries, and as such further research is required to assess the distribution, magnitude, variety, and ecology thereof. However, given the logistical constraints associated with many developing regions, including South Africa, there are numerous research gaps which need to be addressed to facilitate effective management of HABs, some of which include:
-
1.
Identification of alternative resource pools (i.e., organic nutrients and bacterial communities) that facilitate the persistence and dominance of HAB species.
-
2.
Microcosm experiments aimed at isolating the effect of specific environmental variables on HAB species to make predictions regarding global change scenarios.
-
3.
Assessment of how primary consumer communities (e.g., copepods and larval fishes) respond during HAB periods.
-
4.
Molecular comparisons with global strains of HAB species to elucidate the potential dispersal mechanisms facilitating their global occurrence.
-
5.
Feasibility studies of remote sensing as a tool with which to monitor estuarine HABs.
-
6.
Installation and implementation of continuous in situ HAB monitoring technologies.
Pathogen Accumulation in Shellfish Due to Urbanization Near Estuarine Watersheds
Human illness associated with bacterial pathogens has risen alongside population density growth in major cities (i.e., New York City) since the late nineteenth century. This time period corresponds with the initial construction of sewage systems in many urban areas, which delivered human wastes to the nearby estuaries (Rippey 1994). Since then, over 400 different outbreaks have been reported, many of these due to bacteria in wastewater (Rippey 1994). The National Shellfish Sanitation Program and sanitation surveys created in the 1920s (to track effluent and point source inputs) helped to reduce the number of human illnesses (DePaola et al. 2010). Coastal urbanization can lead to elevated inputs of enteric bacteria (fecal coliforms) and nutrient rich waters over subtidal/intertidal shellfish beds, including commercially and ecologically important areas of clams, scallops, and oysters (Mallin et al. 2000; Campos and Cachola 2007; Maalouf et al. 2010; Campos et al. 2013). Many of these organisms live in small estuaries and creeks surrounded by impervious surfaces and agricultural land, and shellfish beds can be closed due increasing fecal coliform bacterial concentrations (Mallin et al. 2000). For example, in the New Hanover County watersheds, NC, there are five tidal creeks with 42.9–77.8% developed land, and in the low salinity regions of the creeks, all five had elevated fecal coliform counts over a sampling season.
EPA standards for shellfish sanitation and shell fishing (1986) indicate that regular fecal coliform sampling should occur to adequately monitor conditions in the water column that could impact the oysters on the bottom (Campos et al. 2013). If routine sampling shows a medial fecal coliform concentration above 14MPN/100 mL (or 10% above 43MPN/100 mL), then water is not safe for shellfish harvesting (EPA Water Quality Criteria 1986). The concentration of fecal indicator organisms that are in the water column serve as a good indicator of what is in the shellfish meat; however, multiple environmental and physiological factors influence the accumulation of fecal coliforms within oyster tissue (reviewed in Campos et al. 2013). Historically, most outbreaks of bacterial pathogens occur in the summer months (Rippey 1994). A nationwide survey of marketed oysters in 2006 examined microbial indicators in shellfish (along with other viral pathogens) and found that mean levels of fecal coliforms and E. coli during summer were above the FDA limits for consumption in some regions (DePaola et al. 2010); in areas with closed shellfish beds, illegally harvested oysters could have much higher pathogen loads and cause illness when consumed (DePaola et al. 2010). Maalouf et al. (2010) examined historical data looking at illness outbreaks associated with sewage discharges over shellfish beds, and found many linkages between the two. Outbreaks of gastroenteritis due to fecal coliforms in sewage effluent were linked to contaminated oysters in Australia, New York, and France in the past three decades (Maalouf et al. 2010).
The amount of urbanization in the surrounding watershed can be estimated by the degree of impervious surfaces sounding the waterways. Impervious surfaces mean that during periods of high rainfall, any pollutants on the surfaces wash into the storm drains/creeks. Mallin et al. (2000) found that if creeks were surrounded by greater than 10% impervious surfaces, they were periodically closed to shell fishing activities due to increased fecal coliform bacteria. Comparison of the fecal coliform abundance with the percentage of impervious surface showed a strong correlation (Pearson correlation r = 0.975, p < 0.005). Impervious surfaces mean that during periods of high rainfall, any pollutants on the surfaces would wash into the storm drains/creeks. During a thorough sampling season highlighting various rainfall amounts, Coulliette and Noble (2008) saw a significant relationship between the quantity of rain and the quantity of fecal coliform bacteria (E. coli and Enterococcus) in the water columns of the Newport River Estuary (NC, USA). Not only did rainfall (above the recommended threshold for shellfish sanitation safety) cause E. coli and Enterococcus levels to rise above the regulated MPN/100 mL levels, there was also a significant rise in fecal indicator bacteria during dry periods, possibly due to a reservoir of cells in the sediment that were suspended into the water column due to human activity in the creeks (Fig. 7).

a Fecal coliform (FC) concentrations found in the Newport River Estuary, NC, according to general rainfall categories of < 0.25 cm, 0.25–2.54 cm, > 2.54 cm, and then in regard to the management action plan of < 3.81 cm and > 3.81 cm. The dotted line indicates the FC standard limit of 14 MPN per 100 ml. Asterisks indicate a significant difference as compared to other categories. Error bars indicate ± 1 standard error. b Mean FIB concentrations at Newport River Estuary sampling sites categorized by distance from land (close ≤ 0.25 km to shoreline, distant ≥ 0.25 km to shoreline). Error bars indicate ± 1 standard error. Reproduced from Journal of Water and Health Volume 6, issue number 4, page 478, with permission from copyright holders IWA Publishing (Coulliette and Noble 2008)
Input of pathogenic bacteria into an enclosed waterway (i.e., a tidal creek) can impact not only the water column but the sediments as well. Fecal coliform bacteria can be deposited onto the substrate during sedimentation, adding to the sediment microflora. It is highly likely that they would be resuspended during tidal cycles, or human interactions (wading in the creek, swimming, etc.). Following a raw sewage spill, Mallin et al. (2007) found that sediment bacteria counts remained elevated for weeks, and increased yet again following an intense precipitation event (Mallin et al. 2007). Once resuspended, fecal coliform bacteria that were present in the sediments (i.e., 200 CFU/cm2) showed corresponding quantities in the water column (i.e., 200 CFU/100 mL). Since sediments can accumulate viable bacterial cells rapidly (acting as a reservoir for cells), this could cause a rapid increase in water column bacteria concentrations above the recommended threshold for human contact (Al Aukidy and Verlicchi 2017; Madoux-Humery et al. 2015; Coulliette and Noble 2008; Mallin et al. 2007). Stormwater input can cause increased turbidity as well, due to both the input of particulates in the runoff as well as the resuspension of existing sediments (Coulliette and Noble 2008). Several studies in both subtropical and temperate regions have demonstrated the survival of fecal indicator bacteria in the sediments; Coulliette and Noble (2008) linked periods of increased fecal indicator bacteria even during dry times to this ‘reservoir effect’ (Fig. 7).
Urbanization-Driven Land Use Changes: Galveston Bay Case Study
The conversion of land to support growing populations is a major component of human modification of the environment around the world (Alberti et al. 2003; Alberti 2005; Diaz and Rosenberg 2008; Paerl et al. 2014). An examination of land use land cover data is an important element in helping us to understand the environmental and anthropogenic influences affecting waterways and watersheds (Carle et al. 2005; Lee et al. 2009). Urbanization, industrialization, agriculture, deforestation, loss of wetlands, and several other types of land use change have taken place in response to human population growth in watersheds all over the world. Urban-related runoff/stormwater is one of the largest contributors to the impairment of river and stream water quality in most US states (USEPA 2010; Paerl et al. 2014), with high levels of eutrophication reported in 45% of the estuaries surrounding the Gulf of Mexico (Clement et al. 2001).
Recent efforts to understand water quality degradation have focused on the watershed of Galveston Bay (TX), a subtropical estuary in the north-western Gulf of Mexico (Fig. 8). With a billion-dollar commercial and recreational fishery, home to the US energy nexus, several large ports and cities, the Galveston Bay ecosystem is under pressure from a variety of sources (Lester et al. 2013). Yet unlike Chesapeake Bay and San Francisco Bay (see e.g., Dauer et al. 2000; Smith et al. 2003; Kemp et al. 2005; Glibert et al. 2011), clear effects of the environmental pressures associated with eutrophication or invasive species have not yet been observed in Galveston Bay (Gonzalez and Lester 2011; Steichen et al. 2012, 2015). Ongoing efforts are focused on understanding the tipping points in this estuarine system (e.g., Roelke et al. 2013; Dorado et al. 2015; Pinckney et al. 2017).

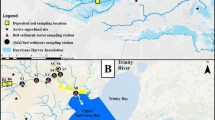
The Galveston Bay watershed extends from Dallas/Fort Worth to Houston to Galveston Island (see inset). Counties surrounding Galveston Bay in the lower watershed and nearby are: Matagorda, Brazoria, Galveston, Chambers, Liberty, Harris, Fort Bend, Wharton, Colorado, Waller, Montgomery, Waller, and Walker. The majority of freshwater inflows (~ 81%) enter from the Trinity and San Jacinto Rivers, and Buffalo Bayou (not shown)
The Galveston Bay watershed (72,000 km2) includes Houston and Dallas, ranked as the fourth and ninth largest cities and fastest growing cities in the USA, respectively (Census 2012, https://www.census.gov). Further, the cities of Austin and Fort Worth rank eleventh and sixteenth, so it is not surprising that the population density in Texas is expected to double by 2050 (Texas Water Development Board TWDB 2007). Galveston Bay (1554 km2), at the southern end of the watershed, is the second largest estuary in the Gulf of Mexico. It has two major rivers flowing into its northwest (San Jacinto) and northeastern (Trinity) reaches. These rivers bring in 16 and 55% of annual freshwater inflows (Guthrie et al. 2012) into the Bay. While the San Jacinto River flows through the highly urbanized and industrialized Houston, the Trinity River is surround primarily by forested and grassland areas immediately north of the Bay (H-GAC 2017).
An examination of land use change maps (National Land Cover Data) between 1996 and 2011 classified by H-GAC using the NOAA Coastal Ocean Program’s Coastal Change Analysis Program (C-CAP) (see H-GAC 2017) indicate an 11% increase in area used for development in Harris County (were Houston is located) since 1996 (Fig. 9). An examination of all the counties surrounding Galveston Bay revealed that forest land cover experienced the greatest loss, primarily due to development (urbanization) during this period (Fig. 9). Forests were also lost to grasslands and more shrubs as well as losses to agricultural (cultivated) lands and wetlands (Fig. 9). In most cases, wetlands were converted into developed lands, to shrubs and grasslands associated with urban community centers, particularly those connected to waterways (Figs. 8 and 9; H-GAC 2017).

The percentage change in land use in counties surrounding Galveston Bay (see Fig. 8). In Harris, Fort Bend, and Montgomery counties, the grow corridor of Houston and surrounding areas, there was a significant increase in developed lands. As a result, there was a significant decrease in forested lands in Harris and Montgomery counties while agricultural lands were lost in Fort Bend county. Development also impacted the land cover in Galveston, Brazoria, Liberty, and Chambers, but to a lesser extent. Loss of forested land was significantly in Liberty; this coincided with increase in grass cover
As a result of the Clean Water Act and other policies, nutrient and chlorophyll a (often used as a proxy for phytoplankton biomass) levels have been declining in Galveston Bay since the 1970s, although less so in more recent years (Gonzalez and Lester 2011). This decline and/or change may also be the result of less agricultural lands surrounding the bay. The losses of wetlands likely preceded the period of record for the land use/land change maps. Nonetheless, we know that consequences of these losses include both habitat quality and dietary changes that impact higher trophic levels. One of the largest increases associated with development in the Galveston Bay watershed has been waterways, either as detention ponds and lakes or canalization associated with residential developments (H-GAC 2017). These are known to be localized zones of degraded water quality, particularly canals that often have a dead end, and experience increased hypoxia and/or harmful algal blooms, predominantly in warmer months (Gonzalez and Lester 2011). Galveston Bay was found to be a hotspot for fish kills over a multi-decadal period (Thronson and Quigg 2008). Despite this, overall fish abundance and diversity has not changed significantly (Steichen and Quigg 2018).
It hypothesized that until now, Galveston Bay has been resilient to the upstream changes in land use and land cover, in terms of eutrophication and subsequent hypoxia and invasive species. The former has not yet been measured, and little information is available to confirm the latter. The lack of obvious signs of eutrophication may in part be facilitated by the relatively high freshwater inflows and short turnover time in the bay (Roelke et al. 2013; Dorado et al. 2015). This may also be due to the fact that the vast majority of developed lands (as urbanization) flushing into the river contributes to only one sixth of the bays freshwater inflows; the other important river (Trinity) contributes more than half the freshwater inflows but has had fewer alterations to its landscape, which is primarily forest and wetlands areas. It may also be that the watershed as a whole has not been overdeveloped, that is, watersheds with ≥ 10% altered land hit the threshold for water quality decreases due to impervious surface effects (Walsh et al. 2005; Zampella et al. 2007). Nonetheless, as regional planning and natural resource management endeavors to determine the appropriate amount of freshwater inflows for Galveston Bay, understanding the balance between land use land cover change and water quality will be key to maintaining ecosystem services and functions for future generations. This is challenging as linear responses to land use change are unlikely given the complex and dynamic nature of estuarine systems.
Hurricane Harvey made landfall from August 25–30 (2017) and set a continental US record for rainfall with more than 100 cm in this short time frame (van Oldenborgh et al. 2017; Emanuel 2017). The massive flooding in urbanized areas is thought to be a result of prolonged changes in land use including development in low lying areas and flood plains along with deferred maintenance projects which results in a weakening of protective barriers around major bayous. Along with extensive flooding, overflowing reservoirs, and overtopping dams, > 100,000 homes were damaged along with numerous schools and businesses (Shultz and Galea 2017). Of the more than 13 million persons directly affected by the storm, 6.5 million were impacted by the rainfall (NOAA NCEI 2017). Highly contaminated Superfund sites in the Houston area were deluged by floodwaters; these along with spills from inundated sewage and wastewater systems released > 31 million gallons of raw sewage (Stuckey 2017). Pollutants released by Texas facilities into Galveston Bay include benzene, 1,3-butadiene, hexane, hydrogen sulfide, toluene, and xylene, at least two of which are cancer-causing chemicals and > 700,000 pounds of sulfur dioxide, which can lead to the formation of dangerous contaminants (FEMA 2017). The impact on the health of the bay is the source of many ongoing studies. While the flood waters effectively “flushed” out the bay, and possibly associated nutrients and pollutants, the concurrent scouring of the sediment bottom, particularly in the industrial areas around Houston and its ship channel may have (re)introduced legacy contaminants such as dioxins, PCBs, lead and others (this data is however not yet published). Ongoing studies will be required to monitor how this massive disruption on urban systems negatively impacts the health of Galveston Bay.
Environmental Limits of Ecosystems: Oyster Case Study
Urbanization has affected coastal species composition and ecosystem function in part by changing biogeochemical processes within the water column (Levin et al. 2014). These alterations can ultimately reduce the survival of species, particularly for those already nearing their physiological tolerance limits. One keystone species found throughout urbanized estuaries along the US East Coast and Gulf of Mexico is the eastern oyster Crassostrea virginica. C. virginica possesses a complex life cycle that includes a benthic adult stage and a pelagic larval phase (Fig. 10). While C. virginica adults are relatively robust to the dynamic physical conditions observed in coastal waters (Bauman et al. 2015), reproduction and survival of early life stages may be disproportionately negatively affected by urbanization. Though factors such as pollution, sedimentation, and competition predation by invasive species have been shown to hurt oyster populations, here the focus is on urbanization-driven changes in physical parameters. Specifically, the effects of eutrophication-driven hypoxia, eutrophication-driven hypercapnia, and lowered salinities from altered streamflow on survival of C. virginica are explored.

Crassostrea virginica reproduction sampling locations. Circles indicate spat collector deployments; squares, reef collections; and stars, continuous water quality sondes
As mentioned in previous sections, one component of urbanization, increased area of impermeable surfaces and subsequent runoff, raises the supply of limiting nutrients to coastal waters, leading to phytoplankton blooms. These blooms are critical to successful fisheries. However, the subsequent bloom die-off and respiration of organic matter produces periods of hypoxia, which may be pronounced in areas of high stratification (Baker and Mann 1992). While adult oysters can withstand prolonged hypoxic periods of days to weeks (Stickle et al. 1989), hypoxia and anoxia reduce both larval set and spat survival in C. virginica (Baker and Mann 1992). In shallow eutrophic tributaries, diel cycling hypoxia is commonly observed during summertime, yet urbanization increases the frequency and severity of the cycling (Keppel et al. 2015). Although normoxic conditions return daily, diel cycling hypoxia both reduces feeding, and increases susceptibility of eastern oysters to the protistan parasite Perkinsus marinus. This potentially leads to estuarine-scale effects of disease acquisition in the species (Breitburg et al. 2015; Keppel et al. 2015). A recent study by Steppe et al. (in prep) demonstrated that severe cycling hypoxia representative of urbanization-impacted estuaries also affects oyster reproduction, both reducing the proportion of reproductive oysters within a sample and lowering gonadosomatic index (the ratio of the germinal tissue to visceral mass). Therefore, eutrophication-driven hypoxia is detrimental to C. virginica throughout its life cycle.
Eutrophication, often a result of urbanization and land use changes, can also lead to the hypercapnic conditions that are frequently found in coastal waters (Cai et al. 2011, Nixon et al. 2015; Bauman et al. 2015). Reduced pH may inhibit C. virginica shell formation (Waldbusser et al. 2010) and dissolve oyster shell base, reducing substrate available for spat set (Waldbusser et al. 2011a, b). Sustained seasonal acidification in line with predictions of severe climate change scenarios has been shown to reduce reproductive success in C. virginica by limiting oogenesis (Boulais et al. 2017). Likewise, constant moderate pH (7.45) has been shown to reduce gametogenesis in C. virginica (Steppe et al. 2019). These values (7.45) coincide with values observed in early summer in Chesapeake Bay tributaries (Steppe and Wallendorf 2017). Cycling pH, on the other hand, does not affect oyster reproduction or disease acquisition (Keppel et al. 2015, though it is associated with increased hemocyte activity, a sign of physiological stress.
Urbanized coastlines have fundamentally changed the physical and chemical parameters of estuaries. Along with reduced oxygen and pH, sharply lowered salinities, such as those caused by coastal storms and dam releases may hurt oyster populations, particularly when these events coincide with gametogenesis and larval development (Butler 1949). Low salinities may reduce the buffering capability of estuarine water. Combined with hypercapnic conditions, this may reduce lipid and glycogen stores, and increase fracturing of juvenile oyster shells (Dickinson et al. 2012). Because mesohaline tributaries are often selected for oyster restoration, due to reduced disease prevalence in these areas (Westby et al. 2015), oysters may already be living close to their salinity tolerance limits. Salinities below 6 may limit gametogenesis (Butler 1949), yet even when gametogenesis is documented, spat recruitment, and subsequent growth is often low in these areas (Bergquist et al. 2006; Steppe and Wallendorf 2017). An example of this has been observed in the Severn River, a mesohaline tributary of Chesapeake Bay, Maryland USA, near the urban centers of Baltimore, Annapolis, and Washington, DC. From 2011 to 2013, high-frequency water quality data (period = 15 min) were collected at multiple sites that coincided with bi-weekly samples of oyster gametogenesis and spat. Gametogenesis was observed each year of the study at three reefs and in oyster grow-out cages, yet of 2760 oyster shell valve “spat collectors” analyzed for recruitment, only 3 spat were recorded. The spat collected were all found in 2012, the only year of the three in which salinities remained above the threshold of 6 throughout the entire spawning period (Steppe et al. 2018).
In addition to spring freshets, salinities may fall due to altered streamflow that cause prolonged periods of low salinity. This includes dam and spillway releases that result from severe storms (Turner 2006; LaPeyre et al. 2016). In 2011, the Conowingo Dam was released in early September, as a result of Hurricane Irene. The release caused the salinity to drop from 9 to 2 at the Severn River study site, likely killing any larvae that may have been in the water column at that time. Similarly, a salinity drop from 8 to 3 in mid-July 2013 may have led to increased larval mortality, and reduced recruitment that year (Steppe et al. 2018).
Further research on the effects of urbanization on ecosystems must be conducted to better understand the role of anthropogenic stresses on water quality and ecosystem function. In particular, high-frequency observations of coastal physical parameters should be colocated with biological sampling of species at each segment of their life history. Additionally, results should be conveyed to managers to better time decisions (such as dam releases) to coincide with period that are less stressful to keystone species.
Closing Remarks
As human populations increase, and technology allows further coastal development, there is a pressing need for improved tools and data collection methods to rapidly obtain meaningful data on broad spatiotemporal scales. Human populations have been experiencing rapid growth over the past century, in addition to a migration towards the coasts. The rapidly changing landscape has altered the nature of estuarine ecosystems and water quality. In general, urbanization land use changes result in increased runoff containing a higher concentration of pollutants, nutrients, and toxic materials, which are introduced to estuarine systems (Carle et al. 2005; DiDonato et al. 2009). Many estuaries are now beyond their capacity to synthesize or process these inputs, meaning that the same pollutants, excess nutrient loads, and toxic materials make their way downstream to the oceans. The dynamics of suspended sediments, nutrients, and pollutants, effects on shellfish, harmful algal blooms, and impacts of land use changes, all have wide reaching effects on estuaries around the world. The reality is that while estuaries provide valuable ecosystem services including seafood, recreation, and a buffer zone from storm energy, urbanization and human development are putting many of these important systems at risk. It is our responsibility to understand the risks and impacts, and make the best use of emerging technologies and data to mitigate and reduce the adverse consequences for estuaries.
References
Al Aukidy, M., and P. Verlicchi. 2017. Contributions of combined sewer overflows and treated effluents to the bacterial load released into a coastal area. Science of the Total Environment. 607: 483–496.
Alberti, M. 2005. The effects of urban patterns on ecosystem function. International Regional Science Review. 28 (2): 168–192.
Alberti, M., J.M. Marzluff, E. Shulenberger, G. Bradley, C. Ryan, and C. Zumbrunnen. 2003. Integrating humans into ecology: Opportunities and challenges for studying urban ecosystems. BioScience. 53 (12): 1169–1179.
Baker, S.M., and R. Mann. 1992. Effects of hypoxia and anoxia on larval settlement, juvenile growth, and juvenile survival of the oyster Crassostrea virginica. Biological Bulletin. 182 (2): 265–269.
Bauman, H., R.B. Wallace, T. Tagliaferri, and C.J. Gobler. 2015. Large natural pH, CO2, and O2 fluctuations in a temperate tidal salt marsh on diel, seasonal, and interannual time scales. Estuaries and Coasts. 38 (1): 220–231.
Beck, M.W., K.L. Heck, K.W. Able, D.L. Childers, D.B. Eggleston, B.M. Gillanders, B. Halpern, C.G. Hays, K. Hoshino, T.J. Minello, R.J. Orth, P.F. Sheridan, and M.P. Weinstein. 2001. The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. BioScience. 51 (8): 633–641.
Bergquist, D.C., J.A. Hale, P. Baker, and S. Baker. 2006. Development of ecosystem indicators for the Suwannee estuary: Oyster reef habitat quality along a salinity gradient. Estuaries and Coasts. 29 (3): 353–360.
Boulais, M., K.J. Chenervet, A.T. Demey, M.R. Robison, J.P. Roberts, and A. Volety. 2017. Oyster reproduction is compromised by acidification experiences seasonally in coastal regions. Scientific Reports 7 (1): 13276.
Breitburg, D.L., D. Hondorp, C. Audemard, R.B. Carnegie, R.B. Burrell, M. Trice, and V. Clark. 2015. Landscape-level variation in disease susceptibility related to shallow-water hypoxia. PLoS One 10 (2): e0116223.
Butler, P.A. 1949. Gametogenesis in the oyster under conditions of depressed salinity. Biological Bulletin. 96 (3): 263–269.
Buzzelli, C.P., R.A. Luettich, H.W. Paerl, J. Fear, J. Fleming, L. Twomey, E. Clesceri, M.J. Alperin, and C.S. Martens. 2001. Neuse River estuary modeling and monitoring project phase 2: monitoring of hydrology and water quality, circulation, phytoplankton physiology and sediment-water coupling, 1–19. WRRI Project.
Cai, W.J., X. Hu, W.J. Huang, M.C. Murrell, J.C. Lehrter, S.E. Lohrenz, W.C. Chou, W. Zhai, J.T. Hollibaugh, Y. Wang, P. Zhao, X. Guo, K. Gundersen, M. Dai, and G.-C. Gong. 2011. Acidification of subsurface coastal waters enhanced by eutrophication. Nature Geoscience Letters. 4 (11): 766–770.
Campos, C.J.A., and R.A. Cachola. 2007. Faecal coliforms in bivalve harvesting areas of the Alvor Lagoon (Southern Portugal): Influence of seasonal variability and urban development. Environmental Monitoring and Assessment. 133 (1-3): 31–41.
Campos, C.J.A., S.R. Kershaw, and R.J. Lee. 2013. Environmental influences on faecal indicator organisms in coastal waters and their accumulation in bivalve shellfish. Estuaries and Coasts. 4: 834–853.
Carle, M.V., P.N. Halpin, and C.A. Stow. 2005. Patterns of watershed urbanization and impacts on water quality. Journal of the American Water Resources Association 41: 693–708.
Clement, C.S., B. Bricker, and D.E. Pirhalla. 2001. Eutrophic conditions in estuarine waters. In NOAA’s state of the coast report. Silver Spring: National Oceanic and Atmospheric Administration http://state-oFcoast.noaa.gov/bulletins/html/eut_18/eut.html. Accessed December 2018.
Cloern, J.E., and A.D. Jassby. 2010. Patterns and scales of phytoplankton variability in estuarine-coastal ecosystems. Estuaries and Coasts. 33 (2): 230–241.
Conley, D.J., H.W. Paerl, R.W. Howarth, D.F. Boesch, S.P. Seitzinger, K.E. Havens, C. Lancelot, and G.E. Likens. 2009. ECOLOGY: Controlling eutrophication: nitrogen and phosphorus. Science 323 (5917): 1014–1015.
Corbett, D.R., D. Vance, E. Letrick, D. Mallinson, and S. Culver. 2007. Decadal-scale sediment dynamics and environmental change in the Albemarle estuarine system, North Carolina. Estuarine, Coastal and Shelf Science. 71 (3-4): 717–729.
Corbett, D.R., J.P. Walsh, and K. Marciniak. 2009. Temporal and spatial variability in the trace metals of two adjacent tributaries of the Neuse River estuary, NC. Marine Pollution Bulletin. 5: 1739–1765.
Coulliette, A.D., and R.T. Noble. 2008. Impacts of rainfall on the water quality of the Newport River estuary (eastern North Carolina, USA). Journal of Water Health. 6 (4): 473–482.
Daborn, G.R., C.L. Amos, M. Brylinksy, H. Christian, G. Drapeau, R.W. Faas, J. Grant, B. Long, D.M. Paterson, G.M.E. Perillo, and M.C. Piccolo. 1993. An ecological cascade effect: Migratory birds affect stability of intertidal sediments. Limnology and Oceanography. 38: 225–231.
Dai, G., J. Zhong, L. Song, C. Guo, N. Gan, and Z. Wu. 2015. Harmful algal bloom removal and eutrophic water remediation by commercial nontoxic polyamine-co- polymeric ferric sulfate-modified soils. Environmental Science and Pollution Research. 22 (14): 10636–10646.
Dauer, D.M., J.A. Ranasinghe, and S.B. Weisberg. 2000. Relationships between benthic community condition, water quality, sediment quality, nutrient loads, and land use patterns in Chesapeake Bay. Estuaries. 23 (1): 80–96.
DeLuca, N., B. Zaitchik, and F. Curriero. 2018. Can multispectral information improve remotely sensed estimates of total suspended solids? A statistical study in Chesapeake Bay. Remote Sensing 10 (9): 1393.
DePaola, A., J.L. Jones, J. Woods, W. Burkhardt III, K.R. Calci, J.A. Krantz, J.C. Bowers, K. Kasturi, R.H. Byars, E. Jacobs, D. Williams-Hill, and K. Nabe. 2010. Bacterial and viral pathogens in live oysters: U.S. markets survey 2007. Applied Environmental Microbiology. 76 (9): 2754–2768.
Diaz, R.J., and R. Rosenberg. 2008. Spreading dead zones and consequences for marine ecosystems. Science. 321 (5891): 926–929.
Dickinson, G.H., A.V. Ivanina, O.B. Matoo, H.O. Pörtner, G. Lannig, C. Bock, E. Beniash, and I.M. Sokolova. 2012. Interactive effects of salinity and elevated CO2 levels on juvenile eastern oysters Crassostrea virginica. Journal of Experimental Biology. 215 (1): 29–43.
DiDonato, G.T., J.R. Stewart, D.M. Sanger, B.J. Robinson, B.C. Thompson, A.F. Holland, and R.F. Van Dolah. 2009. Effects of changing land use on the microbial water quality of tidal creeks. Marine Pollution Bulletin. 58 (1): 97–106.
Dorado, S., T. Booe, J. Steichen, A.S. McInnes, R. Windham, A.K. Shepard, A. Lucchese, H. Preischel, J.L. Pinckney, S.E. Davis, D.L. Roelke, and A. Quigg. 2015. Towards an understanding of the interactions between freshwater inflows and phytoplankton communities in subtropical estuaries. PLoS One 10 (7): e0130931. https://doi.org/10.1371/journal.pone.0130931.
Emanuel, K. 2017. Assessing the present and future probability of Hurricane Harvey’s rainfall. Proceedings of the National Academy of Sciences 114 (48): 12681–12684.
FEMA. 2017. Texas Hurrican Harvey (DR-4332). https://www.fema.gov/disaster/4332. Accessed January 2019.
Freeman, L.A., S.G. Ackleson, and W.J. Rhea. 2017. Comparison of remote sensing algorithms for retrieval of suspended particulate matter concentration from reflectance in coastal waters. Journal of Applied Remote Sensing. 11 (046028): 1.
Giffin, D., and D.R. Corbett. 2003. Evaluation of sediment dynamics in coastal systems via short-lived radioisotopes. Journal of Marine Sciences. 42: 83–96.
Gleeson, J., I.R. Santos, D.T. Maher, and L. Golsby-Smith. 2013. Groundwater–surface water exchange in a mangrove tidal creek: Evidence from natural geochemical tracers and implications for nutrient budgets. Marine Chemistry. 156: 27–37.
Glibert, P.M., D. Fullerton, J.M. Burkholder, J.C. Cornwell, and T.M. Kana. 2011. Ecological stoichiometry, biogeochemical cycling, invasive species, and aquatic food webs: San Francisco Estuary and comparative systems. Reviews in Fisheries Science. 19 (4): 358–417.
Gonzalez, L.A., and L.J. Lester, eds. 2011. State of the bay: A characterization of the Galveston Bay ecosystem, third edition. Houston: Texas Commission on Environmental Quality, Galveston Bay Estuary Program 356 pp.
Goodwin, P., A.J. Mehta, and J.B. Zedler. 2001. Tidal wetland restoration: an introduction. Journal of Coastal Research. 27: 1–6.
Granek, E.F., S. Polasky, C.V. Kappel, D.J. Reed, D.M. Stoms, E.W. Koch, C.J. Kennedy, L.A. Cramer, S.D. Hacker, E.B. Barbier, S. Aswani, M. Ruckelshaus, G.M.E. Perillo, B.R. Silliman, N. Muthiga, D. Bael, and E. Wolanski. 2009. Ecosystem services as a common language for coastal ecosystem-based management. Conservation Biology. 24: 207–216.
Guthrie, C.G., J. Matsumoto, and R. Solis. 2012. Analysis of the influence of water plan strategies on inflows and salinity in Galveston Bay. Austin: Texas Water Development Board.
Hackney, C.T., W.D. Burbanck, and O.P. Hackney. 1976. Biological and physical dynamics of a Georgia tidal creek. Chesapeake Science 17 (4): 271–280.
Hall, N.S., A.C. Whipple, and H.W. Paerl. 2015. Vertical spatio-temporal patterns of phytoplankton due to migration behaviors in two shallow, microtidal estuaries: Influence on phytoplankton function and structure. Estuarine, Coastal and Shelf Science. 162: 7–21.
Havens, K.E., and H.W. Paerl. 2015. Climate change at a crossroad for control of harmful algal blooms. Environmental Science and Technology. 49 (21): 12605–12606.
Hilmer, T., and G.C. Bate. 1991. Vertical migration of a flagellate-dominated bloom in a shallow south African estuary. Botanica Marina. 34: 113–121.
Holland, F., & D. Sanger. 2008. Tidal creek habitats: Sentinels of coastal health. Booklet published by SC Sea Grant Consortium and NOAA for coastal decision makers.
Holland, A.F., D.M. Sanger, C.P. Gawle, S.B. Lerberg, M.S. Santiago, G.H.M. Riekerk, L.E. Zimmerman, and G.I. Scott. 2004. Linkages between tidal creek ecosystems and the landscape and demographic attributes of their watersheds. Journal of Experimental Marine Biology and Ecology. 298 (2): 151–178.
Houston-Galveston Area Council (H-GAC 2017) http://www.h-gac.com/community/socioeconomic/land-use-data/default.aspx (Last accessed December 2017).
Hyland, J.L., W.L. Balthis, C.T. Hackney, and M. Posey. 2000. Sediment quality of North Carolina estuaries: An integrative assessment of sediment contamination, toxicity, and condition of benthic fauna. Journal of Aquatic Ecosystem Stress and Recovery. 8 (2): 107–124.
Kemp, W.M., W.R. Boynton, J.E. Adolf, D.F. Boesch, W.C. Boicourt, G. Brush, J.C. Cornwell, T.R. Fisher, P.M. Glibert, and J.D. Hagy. 2005. Eutrophication of Chesapeake Bay: Historical trends and ecological interactions. Marine Ecology Progress Series. 303: 1–29.
Keppel, A.G., D.L. Breitburg, G.H. Wikfors, R.B. Burrell, and V.M. Clark. 2015. Effects of co-varying diel-cycling and pH on disease susceptibility in the eastern oyster Crassostrea virginica. Marine Ecology Progress Series. 538: 169–183.
Kneib, R.T. 1997. The role of tidal marshes in the ecology of estuarine nekton. Oceanographic Marine Biology Annual Review. 35: 163–220.
LaPeyre, M.K., J. Geaghan, G. Decossas, and J.F. La Peyre. 2016. Analysis of environmental factors influencing salinity patterns, oyster growth, and mortality in lower Breton Sound Estuary, Louisiana, using 20 years of data. Journal of Coastal Restoration. 32: 519–530.
Lee, S.W., S.J. Hwang, S.B. Lee, H.S. Hwang, and H.C. Sung. 2009. Landscape ecological approach to the relationships of land use patterns in watersheds to water quality characteristics. Landscape and Urban Planning. 92 (2): 80–89.
Lemley, D.A., J.B. Adams, and G.C. Bate. 2016. A review of microalgae as indicators in south African estuaries. South African Journal of Botany. 107: 12–20.
Lemley, D.A., J.B. Adams, and N.A. Strydom. 2017a. Testing the efficacy of an estuarine eutrophic condition index: Does it account for shifts in flow conditions? Ecological Indicators 74: 357–370.
Lemley, D.A., J.B. Adams, and S. Taljaard. 2017b. Comparative assessment of two agriculturally-influenced estuaries: Similar pressure, different response. Marine Pollution Bulletin. 117: 135–146.
Lemley, D.A., J.B. Adams, and G.M. Rishworth. 2018a. Unwinding a tangled web: A fine-scale approach towards understanding the drivers of harmful algal bloom species in a eutrophic South African estuary. Estuaries and Coasts. 41 (5): 1356–1369.
Lemley, D.A., J.B. Adams, and N.A. Strydom. 2018b. Triggers of phytoplankton bloom dynamics in permanently eutrophic waters of a south African estuary. African Journal o Aquatic Science. 43 (3): 229–240.
Lerberg, S.B., A.F. Holland, and D.M. Sanger. 2000. Responses of tidal creek macrobenthic communities to the effects of watershed development. Estuaries. 23: 83–853.
Lester, L.J., L.A. Gonzalez, and P. Weeks. 2013. Ecosystem-based management of Galveston Bay, Texas. In Gulf of Mexico origin, waters, and biota, volume 4: Ecosystem-based management, ed. J.W. Day and A. Yáñez-Arancibia. College Station: Texas A&M University Press 460 pp.
Levin, S.A. 1992. The problem of pattern and scale in ecology. Ecology. 73: 1943–1967.
Levin, L.A., K.-K. Liu, K.-C. Emeis, D.L. Breitburg, J. Cloers, C. Deutsch, M. Giani, A. Goffart, E. Hofmann, Z. Lachkar, K. Limburg, S.-M. Liu, E. Montes, W. Naqvi, O. Ragueneau, C. Rabouille, S. Kumar Sarkar, D.P. Swaney, P. Wassman, and K.F. Wishner. 2014. Comparative biogeochemistry-ecosystem-human interactions on dynamic continental margins. Journal of Marine Systems 141: 3–17.
Maalouf, H., M. Pommepuy, and F.S. Le Guyader. 2010. Environmental conditions leading to shellfish contamination and related outbreaks. Food and Environmental virology. 2 (3): 136–145.
Madoux-Humery, A.S., S. Dorner, S. Sauvé, K. Aboulfadl, M. Galarneau, P. Servais, and M. Prevost. 2015. Temporal analysis of microbiological and wastewater micropollutant loads from combined sewer overflows: implications for management. Environmental Science: Processes Impacts 17: 965–974.
Mallin, M.A., K.E. Williams, E.C. Esham, and R.P. Lowe. 2000. Effect of human development on bacteriological water quality in coastal watersheds. Ecological Applications 10 (4): 1047–1056.
Mallin, M.A., L.B. Cahoon, B.R. Toothman, D.C. Parsons, M.R. McIver, M.L. Ortwine, and R.N. Harington. 2007. Impacts of a raw sewage spill on water and sediment quality in an urbanized estuary. Marine Pollution Bulletin. 54 (1): 81–88.
Miller, R.L., and B.A. McKee. 2004. Using MODIS Terra 250 m imagery to map concentrations of total suspended matter in coastal waters. Remote Sensing of Environment 93 (1–2): 259–266.
Möller, I., M. Kudella, F. Rupprecht, T. Spencer, M. Paul, B.K. van Wesenbeeck, G. Wolters, K. Jensen, T.J. Bouma, M. Miranda-Lange, and S. Schimmels. 2014. Wave attenuation over coastal salt marshes under storm surge conditions. Nature Geoscience 7 (10): 727–731.
Nixon, S.W., A.J. Oczkowski, M.E.G. Pilson, L. Fields, C.A. Oviatt, and C.W. Hunt. 2015. On the response of pH to inorganic nutrient enrichment in well-mixed coastal marine waters. Estuaries and Coasts. 38 (1): 232–241.
NOAA. 2017. Assessing the U.S. Climate in 2017. https://www.ncei.noaa.gov/news/national-climate-201712. Accessed January 2019.
Paerl, H.W. 2006. Assessing and managing nutrient-enhanced eutrophication in estuarine and coastal waters: Interactive effects of human climatic perturbations. Ecological Engineering. 26 (1): 40–54.
Paerl, H.W., and J.T. Scott. 2010. Throwing fuel on the fire: Synergistic effects of excessive nitrogen inputs and global warming on harmful algal blooms. Environmental Science & Technology. 44 (20): 7756–7758.
Paerl, H.W., J.L. Pinckney, J.M. Fear, and B.L. Peierls. 1998. Ecosystems responses to internal and watershed organic matter loading: consequence for hypoxia in the eutrophying Neuse River estuary, North Carolina, USA. Marine Ecology Progress Series. 166: 17–25.
Paerl, H.W., N.S. Hall, B.L. Peierls, and K.L. Rossignol. 2014. Evolving paradigms and challenges in estuarine and coastal eutrophication dynamics in a culturally and climatically stressed world. Estuaries and Coasts. 37 (2): 243–258.
Phillips, J.D. 1997. A short history of a flat place: Three centuries of geomorphic change in the Croatan National Forest. Annals of the Association of American Geographers. 87 (2): 197–216.
Pinckney, J.L., A. Quigg, and D.L. Roelke. 2017. Interannual and seasonal patterns of estuarine phytoplankton diversity in Galveston Bay, Texas, USA. Estuaries and Coasts. 40 (1): 310–316.
Qin, Q., and J. Shen. 2017. The contribution of local and transport processes to phytoplankton biomass variability over different timescales in the upper James River, Virginia. Estuarine, Coastal and Shelf Science. 196: 123–133.
Riggs, S.R., J.T. Bray, E.R. Powers, J.C. Hamilton, D.V. Ames, K.L. Owens, D.D. Yeates, S.L. Luca, J.R. Watson, and H.M. Williamson. 1991. Heavy metals in organic rich muds of the Neuse River estuarine system. Albemarle-Pamlico Estuarine Study, Report 90–07, 1–169.
Rippey, S.R. 1994. Infectious diseases associated with molluscan shellfish consumption. Clinical Microbiology Reviews. 7 (4): 419–425.
Roelke, D.L., H.-P. Li, N.J. Hayden, C.J. Miller, S.E. Davis, A. Quigg, and Y. Buyukates. 2013. Co-occurring and opposing freshwater inflow effects on phytoplankton biomass, productivity and community composition of Galveston Bay, USA. Marine Ecology Progress Series. 477: 61–76.
Ruhl, C.A., D.H. Schoellhamer, R.P. Stumpf, and C.L. Lindsay. 2001. Combined use of remote sensing and continuous monitoring to analyse the variability of suspended- sediment concentrations in San Francisco Bay, California. Estuarine, Coastal and Shelf Science 53 (6): 801–812.
Sanger, D., Blair A., DiDonato G., Washburn T., Jones S., Chapman R., & White D. (2008). Support for integrated ecosystem assessments of NOAA’s National Estuarine Research Reserves System (NERRS), Volume I: The impacts of coastal development on the ecology and human well-being of tidal creek ecosystems of the US Southeast.
Sanger, D., A. Blair, G. DiDonato, T. Washburn, S. Jones, G. Riekerk, and A.F. Holland. 2015. Impacts of coastal development on the ecology of tidal creek ecosystems of the US southeast including consequences to humans. Estuaries and Coasts. 38 (S1): 49–66.
Satterthwaite, D., G. McGranahan, and C. Tacoli. 2010. Urbanization and its implications for food and farming. Philosophical transactions of the Royal Society B: Biological Sciences 365 (1554): 2809–2820.
Scharler, U.M., and D. Baird. 2003. The influence of catchment management on salinity, nutrient stochiometry and phytoplankton biomass of Eastern Cape estuaries, South Africa. Estuarine, Coastal and Shelf Science 56 (3-4): 735–748.
Schropp, S.J., F.G. Lewis, H.L. Windom, J.D. Ryan, F.D. Calder, and L.C. Burney. 1990. Interpretation of metal concentrations in estuarine sediment of Florida using aluminum as a reference element. Estuaries. 13 (3): 227–235.
Schubel, J.R., and H.H. Carter. 1984. The estuary as a filter for fine-grained suspended sediment. In The estuary as a filter, 81–105. Academic Press.
Shenker, J.M., and J.M. Dean. 1979. The utilization of an intertidal salt marsh creek by larval and juvenile fishes: abundance, diversity and temporal variation. Estuaries. 2 (3): 154–163.
Shi, B.W., S.L. Yang, Y.P. Wang, T.J. Bouma, and Q. Zhu. 2012. Relating accretion and erosion at an exposed tidal wetland to the bottom shear stress of combined current wave action. Geomorphology. 138 (1): 380–389.
Shultz, J.M., and S.o Galea. 2017. Mitigating the mental and physical health consequences of Hurricane Harvey. JAMA 318 (15): 1437.
Smith, S.V., D.P. Swaney, L. Talaue-Mcmanus, J.D. Bartley, P.T. Sandhei, C.J. McLaughlin, V.C. Dupra, C.J. Crossland, R.W. Buddemeier, and B.A. Maxwell. 2003. Humans, hydrology, and the distribution of inorganic nutrient loading to the ocean. BioScience. 53 (3): 235–245.
Steichen, J.L., and A. Quigg. 2018. Fish species as indicators of freshwater inflow within a subtropical estuary in the Gulf of Mexico. Ecological Indicators 85: 180–189.
Steichen, J. L., R. Windham, R. Brinkmeyer, and A. Quigg. 2012. Ballast water impacts on Galveston Bay, Texas. Bulletin of Marine Pollution. 64: 779–789.
Steichen, J.L., A. Denby, R. Windham, R. Brinkmeyer, and A. Quigg. 2015. A tale of two ports: Dinoflagellate and diatom communities found in the high ship traffic region of Galveston Bay, Texas (USA). Journal of Coastal Research. 31: 407–416.
Steppe, C. N. and L. Wallendorf. 2017. Reproduction potential of Crassostrea virginica in the Severn River, Maryland, USA. A Final report to the Army Corps of Engineers, Baltimore District. 20 p.
Steppe, C.N., A.G. Keppel, R.B. Burrell, and D.L. Breitburg. 2019. Effects of co-varying diel cycling hypoxia and pH on gametogenesis in eastern oysters. In Prep.
Stickle, W.B., M.A. Kaper, L.-L. Liu, E. Gnaiger, and S.Y. Wang. 1989. Metabolic adaptations of several species of crustaceans and molluscs to hypoxia: tolerance and microcalorimetrec studies. Biological Bulletin. 177 (2): 303–312.
Stuckey, A. 2017. Port Arthur plant had largest wastewater spill in Texas after Harvey. Houston Chronicle, online edition. 21 November 2017.
Stumpf, R.P., and J.R. Pennock. 1989. Calibration of a general optical equation for remote sensing of suspended sediments in a moderately turbid estuary. Journal of Geophysical Research: Oceans 94 (C10): 14363–14371.
Texas Water Development Board (TWDB). 2007. Highlights of the 2007 state water plan. Texas Water Development Board. Document number GP-8-1.
Thronson, A., and A. Quigg. 2008. Fifty-five years of fish kills in coastal Texas. Estuaries and Coasts. 31 (4): 802–813.
Townend, I., C. Fletcher, M. Knappen, and K. Rossington. 2011. A review of salt marsh dynamics. Water and Environment Journal. 25 (4): 477–488.
Turner, R.E. 2006. Will lowering estuarine salinity increase Gulf of Mexico oyster landings? Estuaries and Coasts. 29 (3): 345–352.
U.S. EPA. 1986. Quality criteria for water. Vol. 397. Washington, DC: U.S. Environmental Protection Agency.
U.S. EPA. (2004). The incidence and severity of sediment contamination in surface waters of the United States. National Sediment Quality Survey: 2nd edition. EPA-823-R04-007. 280 p.
U.S. EPA. (2006). Contaminated sediment in water. 1 May 2006. http://epa.gov/waterscience/cs/ Accessed Jan 2018.
U.S. EPA. 2010. Texas water quality report. Washington, DC: U.S. Environmental Protection Agency.
van Oldenborgh, G.J., K. van derWiel, A. Sebastian, R. Singh, J. Arrighi, F. Otto, K. Haustein, S. Li, G. Vecchi, and H. Cullen. 2017. Attribution of extreme rainfall from Hurricane Harvey, August 2017. Environmental Research Letters 12 (12): 124009.
Umar, M., B.L. Rhoads, and J.A. Greenberg. 2018. Use of multispectral satellite remote sensing to assess mixing of suspended sediment downstream of large river confluences. Journal of Hydrology 556: 325–338.
Van Rijn, L.C. 1993. Principles of sediment transport in rivers, estuaries and coastal seas. In Aqua publications, vol. 1006. Amsterdam.
Vitousek, P.M., H.A. Mooney, J. Lubchenco, and J.M. Melillo. 1997. Human domination of earth’s ecosystems. Science. 25: 494–499.
Voulgaris, G., and S.T. Meyers. 2004. Temporal variability of hydrodynamics, sediment concentration and sediment settling velocity in a tidal creek. Continental Shelf Research. 24 (15): 1659–1683.
Waldbusser, G.G., H. Bergschneider, and M.A. Green. 2010. Size-dependent pH effect on calcification in post-larval hard clam Mercenaria spp. Marine Ecology Progress Series 417: 171–182.
Waldbusser, G.G., R.A. Steenson, and M.A. Green. 2011a. Oyster shell dissolution rates in estuarine waters: Effects of pH and shell legacy. Journal of Shellfish Research 30 (3): 659–669.
Waldbussuer, G.G., E.P. Voigt, H. Bergschneider, M.A. Green, and R.I.E. Newell. 2011b. Biocalcification in the eastern oyster (Crassostrea virginica) in relation to long-term trends in Chesapeake Bay pH. Estuaries and Coasts 34 (2): 221–231.
Walsh, C.J., A.H. Roy, J.W. Feminella, P.D. Cottingham, P.M. Groffman, and R.P. Morgan. 2005. The urban stream syndrome: current knowledge and the search for a cure. Journal of the North American Benthological Society. 24 (3): 706–723.
Wang, C., W. Li, S. Chen, D. Li, D. Wang, and J. Liu. 2018. The spatial and temporal variation of total suspended solid concentration in Pearl River estuary during 1987–2015 based on remote sensing. Science of the Total Environment 618: 1125–1138.
Washburn, T., and D. Sanger. 2011. Land use effects on macrobenthic communities in southeastern United States tidal creeks. Environmental Monitoring and Assessment. 180 (1-4): 177–188.
Webster, L.F., D.A. Graves, D.A. Eargle, D.E. Chestnut, J.A. Gooch, and M.H. Fulton. 2013. Assessment of animal impacts on bacterial water quality in a South Carolina, USA tidal creek system. Environmental Monitoring and Assessment. 185 (9): 7749–7756.
Westby, S. R., E. Weissburger, K. Gross, C. O’Neill, A. Sowers, S. Allen, K. Muzia, and Ward S.. 2015. Little Choptank River oyster restoration tributary plan: a blueprint for sanctuary restoration. Maryland Interagency Oyster Restoration Workgroup of the Sustainable Fisheries Goal Implementation Team, February 2015, NOAA.
Whitfield, A.K., N.C. James, S.J. Lamberth, J.B. Adams, R. Perissinotto, A. Rajkaran, and T.G. Bornman. 2016. The role of pioneers as indicators of biogeographic range expansion caused by global change in southern African coastal waters. Estuarine, Coastal and Shelf Science 172: 138–153.
Zampella, R.A., N.A. Procopio, R.G. Lathrop, and C.L. Dow. 2007. Relationship of land-use/land-cover patterns and surface-water quality in the Mullica River basin. Journal of the American Water Resources Association 43 (3): 594–604.
Zhou, Z., C. Bian, C. Wang, W. Jiang, and R. Bi. 2017. Quantitative assessment on multiple timescale features and dynamics of sea surface suspended sediment concentration using remote sensing data. Journal of Geophysical Research: Oceans 122 (11): 8739–8752.
Zu Ermgassen, P.S., M.D. Spalding, B. Blake, L.D. Coen, B. Dumbauld, S. Geiger, J.H. Grabowski, R. Grizzle, M. Luckenbach, K. McGraw, and W. Rodney. 2012. Historical ecology with real numbers: past and present extent and biomass of an imperiled estuarine habitat. Proceedings of the Royal Society of London B 27: 3393–3400.
Acknowledgments
The authors are grateful to the participants of CERF 2017 Session 1299: Ruth Carmichael, Nelle D’Aversa, Bethany DeCourten, Naomi Detenbeck, Brooke Frohloff, Alexandria Hounshell, Michael T. Lee, Haley Nicholson, and Guangming Zheng, as well as the organizers of the CERF 2017 Biennial Conference in Providence, RI. LAF appreciates feedback and support from Art Miller & Simon Freeman. AMF is grateful for released-time support from NJCU-College of Arts & Sciences. AQ: the Houston-Galveston Area Council graciously provided the land use land cover data (http://www.h-gac.com/; December 2016). Without this data and support from the Galveston Bay Estuary Program, this work would not have been possible. CNS thanks USACE, Baltimore District, D. Brietburg, R. Burrell, (Smithsonian Environmental Research Center) A. Keppel, and L. Wallendorf (United States Naval Academy). An anonymous reviewer provided constructive suggestions that greatly improved the quality of this manuscript.
Funding
DRC is supported by the NC Sea Grant (R/MG-1522). DAL is supported by the South African Network for Coastal and Oceanic Research (SANCOR) and the National Research Foundation (NRF) of South Africa through a postdoctoral fellowship (grant number 112650)
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Kenneth L. Heck
Rights and permissions
About this article

Cite this article
Freeman, L.A., Corbett, D.R., Fitzgerald, A.M. et al. Impacts of Urbanization and Development on Estuarine Ecosystems and Water Quality. Estuaries and Coasts 42, 1821–1838 (2019). https://doi.org/10.1007/s12237-019-00597-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-019-00597-z