
Abstract
In this paper, we study the global properties of a computer virus propagation model. It is, interesting to note that the classical method of Lyapunov functions combined with the Volterra–Lyapunov matrix properties, can lead to the proof of the endemic global stability of the dynamical model characterizing the spread of computer viruses over the Internet. The analysis and results presented in this paper make building blocks towards a comprehensive study and deeper understanding of the fundamental mechanism in computer virus propagation model. A numerical study of the model is also carried out to investigate the analytical results.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
By development of computer technologies and network applications, the Internet has become a powerful mechanism for propagating computer virus. Because of this, computers connected to the Internet become much vulnerable to digital threats. Computer viruses, including the narrowly defined viruses and network worms, are loosely defined as malicious codes that can replicate themselves and spread among computers. In this scenario, the large number of existing computer viruses and their high level of destructivity appear as an important risk factor for corporations and individuals. Developing a mathematical model for the computer viral propagation is of critical importance not only for understanding better the behavior of computer virus but also for stopping the spread of the virus. These models lead to a better understanding and prediction of the scale and speed of computer virus propagation (Murray 1988; Yang and Yang 2015; Piqueira et al. 2008). Due to the high similarity between computer virus and biological virus, some classic epidemic models were established for computer virus propagation, such as the SIRS model (Han and Tan 2010; Ren et al. 2012; Bradley et al. 2008), the SEIR model (Yuan and Chen 2008), the SEIRS model (Mishra and Saini 2007), and the SEIQV model Wang et al. (2010), which share a common assumption that an infected computer in which the virus resides is in latency can not infect other computers (Yang et al. 2013; Li and Knickerbocker 2007).
The study of the endemic global stability is not only mathematically important, but also essential in predicting the evolution of the disease in the long run, so that prevention and intervention strategies can be effectively designed, and public health administrative efforts can be properly scaled. There are some methods, i.e., those based on the monotone dynamical systems (Li et al. 1999), and Lyapunov functions (Chong et al. 2014; Liu et al. 2015), to conduct global stability analysis for epidemic models (Xu and Ma 2010; Bhunu and Mushayabasa 2013; Imran et al. 2014). In addition, the method of Lyapunov functions has been known for many decades. The challenge in the application of this method is that there is no systematic way to construct Lyapunov functions (particularly, the determination of the appropriate coefficients is often a matter of luck), so that its success largely depends on trial and error as well as on specific problems. In this paper, we apply the method of Lyapunov functions combined with the Volterra–Lyapunov matrix properties which lead to the proof of the endemic global stability (Redheffer 1985a, b; Rinaldi 1990; Wang and Liao 2012; Chavez et al. 2002). In fact, we incorporate the Volterra–Lyapunov matrix theory (Rinaldi 1990) into Lyapunov functions, under certain conditions, which can leverage the difficulty of determining specific coefficient values, and, as such wider application of Lyapunov functions to dynamical systems could be promoted. Although the method of Lyapunov functions has been widely applied to various dynamical systems, the main contributions of our analysis is based on the less well-known results of Volterra–Lyapunov stable matrices. We are able to investigate more complex model systems with nonlinear incidence rates, with the aid of the Volterra–Lyapunov matrix properties.
Recently, Yang et al. (2013) proposed a computer virus propagation model, which incorporates the two features mentioned above. One major difficulty in studying the qualitative properties of this model lies in the construction of suitable Lyapunov functions. They used linear combinations of quadratic functions in independent variables as the candidate Lyapunov functions. Equipped with this tool, it is proved that the dynamic behavior of the model is determined by a threshold \(R_0.\) Specifically, the virus-free equilibrium is globally asymptotically stable if \(R_0 \le 1,\) whereas the viral equilibrium is locally asymptotically stable if \(R_0 > 1.\) In the present work, we employ the Volterra–Lyapunov metod, to investigate the endemic global stability for the computer virus propagation model (Yang et al. 2013).
We organize the remainder of this paper as follows. In section “Model formulation”, we analyze the computer virus propagation model and investigate the mathematical analysis of this model. In section “Some notations and preliminaries”, we investigate the global stability of the model. Finally, we close the paper by conclusions and discussion.
Model formulation
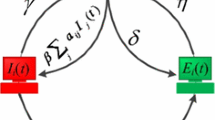
We shall study the mathematical model of computer virus proposed by Yang et al. (2013). A computer is classified as internal and external depending on weather it is connected to internet or not. In this model, assumes that only internal computers are concerned, and all internal computers are categorized into three classes: uninfected computers (i.e., virus-free computers), infected computers that are currently latent (latent computers, for short), and infected computers that are currently breaking out (seizing computers, for short). Due to the fact that in the future, the total amount of computers in the world would tend to saturation, it is reasonable to suppose that this total number is constant.
Let S(t), L(t), and B(t) denote, at time t, the percentages of uninfected, latent, and seizing computers in all internal computers, respectively. Then, \(S(t) + L(t) + B(t) = 1.\) Unless otherwise stated, let S, L, and B stand for S(t), L(t), and B(t), respectively. By carefully considering the features of a computer virus, the following hypotheses are made:
-
(H1) External computers are connected to the Internet at positive constant rate \(\delta\), and internal computers are disconnected from the Internet also at this rate.
-
(H2) All newly connected computers are virus free.
-
(H3) The percentage of internal computers infected at time t increases by \(\beta S(L + B),\) where \(\beta\) is a positive constant. This hypothesis says that both seizing and latent computers have infectivity. In contrast, all the traditional models assumed that only seizing individuals have infectivity, i.e., at time t the force of infection can be described as \(\beta S f(B)\) (Britten 2003).
-
(H4) Latent computers break out at positive constant rate \(\alpha .\)
-
(H5) Latent computers are cured at positive constant rate \(\gamma _1\), while breaking-out computers are cured at positive constant rate \(\gamma _2\). For the graded cure rates, we have \(\gamma _2>\gamma _1>0.\)
Based on the previous assumptions, one can derive the following computer virus propagation model:
Remark 2.1
From the biological point of view, the spread of virus is similar to sexually disease, worm is similar to flu, and logic bomb is similar to HIV. Kephart used the epidemic models to find out the rule in the computer virus (Kephart and White 1991) and he focused his attention on the effect of topological structure of the network on the spread of the virus.
Now, we will discuss the existence of all possible equilibria of the model system (1). We found that system (1) has two possible non-negative equilibria, namely, the virus-free equilibrium \(E_0\) and the viral equilibrium \(E_1.\)
Virus-free equilibrium \(E_0\) and the basic reproduction number \(R_0\)
In this subsection, we shall investigate the existence of equilibria of system (1). The virus-free equilibrium (VFE) is always feasible, as at this equilibrium the infection eradicates from the population. We calculate the basic reproduction number, \(R_0\), using the next generation approach, developed in Van den Driessche and Watmough (2002).
Let consider the non-negative orthant of \(\mathbb {R}^n\) by \(\mathbb {R}_+^n\). We let \(x(t) \in \mathbb {R}_+^n,\) where \(x_i(t)\) denotes the number of individuals in compartment i at time t. For ease of notation, we order the compartments, such that the first m (for \(m\le n\)) compartments correspond to the states with infection. Note that the definition of a state with infection needs to be made from an understanding of the system being modeled and not from the infections themselves (and that this definition may not be unique). For the model
with non-negative initial conditions, \(x(0) \in \mathbb {R}_+^n\), we define \(X_s\) as the set of all disease-free states
We rewrite the model (2) as
where \(\mathcal {F}_i(x)\) is the rate of new infections entering compartment i, and
where \(\mathcal {V}_i^+(x)\) is the rate of transfer into compartment i by any other means, and \(\mathcal {V}_i^-(x)\) is the rate of transfer out of compartment i. We list five reasonable assumptions for these functions below.
-
(A1) If \(x\in \mathbb {R}_+^n\), then \(\mathcal {F}_i(x)\), \(\mathcal {V}_i^+(x)\), \(\mathcal {V}_i^-(x)>0\) for \(1\le i \le n.\) This implies that no rate of movement can be negative.
-
(A2) If \(x_i = 0,\) then \(\mathcal {V}_i^-(x)=0\). If there are no individuals in a compartment, there can be no movement of individuals out of that compartment.
-
(A3) \(\mathcal {F}_i(x) = 0\) for \(i > m.\) There can be no infections entering classes that are defined as noninfectious.
-
(A4) If \(x \in X_s,\) then \(\mathcal {F}_i(x) = 0\) and \(\mathcal {V}_i^+(x)=0\) for \(1\le i \le m.\) If there is no infection in the population, there can be no input into the infectious populations.
-
(A5) if F(x) is set to 0, then all eigenvalues of the corresponding Jacobian of (2) evaluated at a disease-free equilibrium point \(x_{dfe}\), have negative real part. Note that the disease-free equilibrium point is not required to be unique.
For a disease-free equilibrium point, \(x_{\mathrm{dfe}}\), of (2), where \(x_{\mathrm{dfe}}\) and f(x) satisfy assumptions (A1)–(A5), we define \(m\times m\) matrices, F and V
For this model, the matrices F and V, for the new infection and the remaining transfer, are, respectively, given by
and
Then, the basic reproduction number is defined, as the spectral radius of the next generation matrix, \(FV^{-1}\). For this model, the basic reproduction number, which is defined as the number of previously uninfected computers that are infected by a single infected computer during its life cycle, can be derived as
Clearly, system (1) always has a virus-free equilibrium \(E_0=(1,0,0).\)
The endemic equilibrium
A positive equilibrium of (1), if one exists, is called an endemic equilibrium, and denoted by
satisfy the following equilibrium equations:
where,
The authors in Yang et al. (2013) answered the basic questions of the stability of the DFE and the local dynamics of the endemic equilibrium, for the system (1). The global stability of the endemic equilibrium for this model, however, is much more difficult to analyze. Below, we will combine the method of Lyapunov functions and Volterra–Lyapunov stable matrices to address this challenge.
Some notations and preliminaries
Our goal here is to show that the endemic equilibrium is globally asymptotically stable. First, we introduce necessary concepts and notations that will facilitate our global stability analysis.
Notation 3.1
We write a matrix \(A > 0(<0)\) if A is symmetric positive (negative) definite. The following fundamental result on matrix stability was originally proved by Lyapunov:
Lemma 3.2
(Cross 1979). Let A be an \(n\times n\) real matrix. Then, all the eigenvalues of A have negative (positive) real parts if and only if there exists a matrix \(H > 0,\) such that \(HA + A^TH^T < 0(>0).\)
Definition 3.3
We say a nonsingular \(n\times n\) matrix A is Volterra–Lyapunov stable if there exists a positive diagonal \(n\times n\) matrix M, such that \(MA + A^TM^T < 0.\)
The following lemma determines all \(2\times 2\) Volterra–Lyapunov stable matrices.
Lemma 3.4
(Rinaldi 1990; Cross 1979). Let \({ D=}\left[ \begin{array}{cc} {{ d}}_{{ 11}} &{} {{ d}}_{{ 12}} \\ {{ d}}_{{ 21}} &{} {{ d}}_{{ 22}} \end{array}\right]\) be a \(2\times 2\) matrix. Then D is Volterra–Lyapunov stable if and only if \(d_{11}< 0, ~ d_{22} < 0,\) and \(det(D) = d_{11}d_{22}- d_{12}d_{21} > 0.\)
The characterization of Volterra–Lyapunov stable matrices of higher dimensions, however, is much more difficult. We need the following definition.
Definition 3.5
We say a nonsingular \(n\times n\) matrix A is diagonally stable (or positive stable) if there exists a positive diagonal \(n\times n\) matrix M, such that \(MA + A^TM^T > 0.\)
From Definitions 3.3 and 3.5, it is clear that a matrix A is Volterra–Lyapunov stable if and only if its negative matrix, \(-A\), is diagonally stable.
Notation 3.6
For any \(n\times n\) matrix A, let \(\widetilde{ A}\) denote the \((n-1)\times (n-1)\) matrix obtained from A by deleting its last row and last column.
The following generalized result was obtained by Redheffer (1985a, b) which will be frequently used in our global stability analysis. For simplicity, we only state the sufficient condition below.
Lemma 3.7
Redheffer (1985a, b). Let \(D = [d_{ij}]\) be a nonsingular \(n\times n\) matrix \((n\ge 2)\) and \(M = diag(m_1, \ldots ,m_n)\) be a positive diagonal \(n\times n\) matrix. Let \(E = D^{-1}.\) Then, if \(d_{nn} > 0,\) \(\widetilde{{ M}}\widetilde{{ E\ }}{ +}{{ (}\widetilde{{ M}}\widetilde{{ E\ }}{ )}}^{{ T}}{ >0}\), and \(\widetilde{{ M}}\widetilde{{ D}}{ +}{{ (}\widetilde{{ M}}\widetilde{{ D}}{ )}}^{{ T}}{ >0\ }\), it is possible to choose \(m_n > 0\) , such that \(MD + D^TM^T > 0.\)
Lemma 3.8
(Chavez et al. 2002). Consider a disease model system written in the form:
where \(X_1\in \mathbb {R}^m\) denotes (by its components) the uninfected populations and \(X_2\in \mathbb {R}^n\) denotes (by its components) the infectious populations; \(X_0=(X_1^E,0)\) denotes the disease-free equilibrium of the system.
In addition, assume the conditions (C1) and (C2) below:
-
\(C_1\) : For \(\frac{d {X_1}}{ dt} =F(X_1,0),\) \(X_1^E\) is globally asymptotically stable;
-
\(C_2\) : \(G(X_1,X_2)=A X_2-\hat{G}(X_1,X_2),\) with \(\hat{G}(X_1,X_2)\ge 0\) for \((X_1,X_2) \in \Omega\), where the Jacobian matrix \(A=\frac{\partial G}{\partial X_2}(X_1^E,0)\) has all non-negative off-diagonal elements, and X is the region where the model makes biological sense.
Then, the DFE \(X_0 =(X_1^E,0)\) is globally asymptotically stable provided that \(R_0 < 1.\)
Global stability of disease-free equilibrium
We will use the theorem by Castillo-Chavez et al. (2002), to prove the global stability result.
Theorem 3.9
The fixed point \(E_0 = \left( 1,0,0\right)\) is a globally asymptotically stable equilibrium of system (1) provided that \(R_0 < 1\) and the assumptions in Eq. (5) are satisfied.
Proof
Applying Lemma 3.8 to system (1), consider \(X_1=S,\) \(X_2=\left[ \begin{array}{c} L\\ B \\ \end{array} \right] .\)
When \(L =B= 0,\) the uninfected subsystem (i.e., the equation for S) becomes
which has the solution
obviously, \(S(t) \rightarrow 1\) as \(t \rightarrow \infty\) regardless of the initial value S(0). Therefore, it shows that condition \((C_1)\) in Lemma 3.8 holds for our model.
Next, the right-hand side of the infectious subsystem (i.e., the equations for L and B) can be written as
where
and
It is obvious that, \(S\le 1\), hence, it is clear that condition \((C_2)\) holds for our model. We also notice that the matrix A is an M matrix, since all its off-diagonal elements are non-negative. Hence, this proves the global stability of the DFE \((E_0).\) \(\square\)
Global stability of the endemic equilibrium of the model system (1)
Our goal here is to show that the endemic equilibrium of the model system (1) is globally asymptotically stable. It is, however, interesting to note that the classical method of Lyapunov functions combined with the Volterra–Lyapunov matrix properties Redheffer (1985a, b) can lead to the proof of the endemic global stability. The details are provided below.
To prove global stability result, we propose the following Lyapunov function:
where \(w_1,\) \(w_2,\) and \(w_3\) are positive constants. Calculating the time derivative of V along the trajectories of the system (1), we obtain
Then, we add the expression \(\beta S^*L\) and \(\beta S^*B\) into the first and second square bracket. As a result, we obtain
therefore, we have
where \(Y = [S-S^*, L-L^*,B-B^*], W= diag(w_1,w_2,w_3),\) and
To discuss the global asymptotic stability of \(\text{E}_{1} = (\text{S}^{*}, \text{L}^{*}, \text{B})\), we proceed to show that the matrix P defined in Eq. (10) is Volterra–Lyapunov stable or \(-P\) is diagonal stable. For this goal, we prove the following lemmas.
Lemma 3.10
For the matrix P defined in Eq. (10), \(-{P},\) is diagonal stable.
Proof
To prove the diagonal stability of \(-P\) and based on Lemma 3.7, we need to show that the following three conditions are satisfied:
Condition 1. We show that the matrix \(D=-\tilde{P}\) is diagonal stable. From Eq. (10), we obtain
For this purpose, it is necessary to show that \(-D\) is Volterra–Lyapunov stable:
Clearly, \(-{D}_{11} < 0.\) Next, we show \(-{D}_{22} < 0\); according to (4), we have
and it is obvious that
hence, \(-{D}_{22} < 0\). Now, we show \(-{D}_{12} < 0\), that is
using (4), (5), we can see that
since \(0<\gamma _1<\gamma _2\), we have
therefore
hence, \(-{D}_{12} < 0\). It is easy to see \(-{D}_{21} > 0\). Therefore, \(-{D}\) is Volterra–Lyapunov stable based on Lemma 3.4.
Condition 2. We show that the matrix \(E=\widetilde{-P^{-1}}\) is diagonal stable. In fact, we show that \(-E\) is Volterra–Lyapunov stable:
where
It is obvious that \(-E_{21}<0\) and \(-E_{22}<0.\) Below, we show \(-E_{11}=0\) and \(-E_{12}>0.\)
The (1, 1) entrie of this \(-E\) is writen as
multiplying the (4) by \(\alpha\), and using (5) we have
Therefore
from where
hence, \(-E_{11} = 0\).
It is easy to see \(det(-E)>0,\) see the Appendix of this paper. Therefore, \(-E\) is Volterra–Lyapunov stable based on Lemma 3.4.
Condition 3. It is obvious that \(-P_{33}>0\).
Hence, Lemma 3.7 guarantees that \(-P\) is diagonal stable. \(\square\)
Theorem 3.11
The matrix P defined in Eq. (10) is Volterra–Lyapunov stable.
Proof
Based on Lemmas 3.7 and 3.10, there exists a positive diagonal matrix W, such that \(W(-P) + (-P)^TW^T > 0.\) Thus \(WP + P^TW^T < 0.\) \(\square\)
Theorem 3.12
The endemic equilibrium, \(E_1 = (S^*,L^* ,B^*),\) of model (1) is globally asymptotically stable.
Proof
Based on Lemmas 3.7 and 3.10 and Theorem 3.12, we obtain \(\frac{\mathrm{d}V}{\mathrm{d}t} < 0\) when \(X \ne X^*\) and X is not on the S-axis (a set of measure zero). It implies that the endemic equilibrium of the model system (1) is globally asymptotically stable. \(\square\)
Numerical results
In this subsection, we carry out numerical simulations and discuss results. Consider system (1) with \(\alpha = 0.6, \beta = 0.3, \delta = 0.1, \gamma _1 = 0.1, \gamma _2 = 0.3.\) We plot the phase plane portrait of L vs. S and B vs. S in Figs. 1, 2 for \(R_0 = 0.9375,\) a typical case of \(R_0 < 1,\) where the DFE is globally asymptotically stable. This is evidenced in these figures by the fact that all the five orbits converge to the DFE at \(S = 1\) and \(L=B=0.\)
In addition, consider system (1) with \(\alpha = 0.3, \beta = 0.4, \delta = 0.1, \gamma _1 = 0.1, \gamma _2 = 0.3.\) Then, \(R_0= 1.4\) in this case, and the unique positive endemic equilibrium is located at \(S^*= 0.71,~ L^*= 0.1\) and \(B^*=0.1.\) We pick five different initial conditions, and plot these five solution curves by the phase plane portrait of L vs. S and B vs. S in Figs. 3, 4. From which one can see all these five orbits converge to the endemic equilibrium, showing the global asymptotic stability of the endemic equilibrium.

Phase plane portraits of L vs. S for system (1). The basic reproduction number is \(R_0 = 0.9.\) The five curves correspond to different initial conditions with \(L(0) = 0.1,0.3,0.5,0.7,0.9,\) respectively

Phase plane portraits of B vs. S for system (1). The basic reproduction number is \(R_0 = 0.9.\) The five curves correspond to different initial conditions with \(B(0) = 0.1,0.3,0.5,0.7,0.9,\) respectively

Phase plane portraits of L vs. S for system (1). The basic reproduction number is \(R_0=1.4.\) The five curves correspond to different initial conditions with \(L(0) = 0.1,0.3,0.5,0.7,0.9,\) respectively

Phase plane portraits of B vs. S for system (1). The basic reproduction number is \(R_0=1.4.\) The five curves correspond to different initial conditions with \(B(0) = 0.1,0.3,0.5,0.7,0.9,\) respectively
Remark 3.13
The method considered in this paper is conceptually simple to implement, and can be applied to different models with nonlinear incidence rates. As can be expected, the implementation of this method will likely be hindered for epidemiological models with more complex incidence rates, or those with even higher dimensions, though such difficulty remains the same for all other existing methods in global stability analysis. As far as the current method is concerned, some symbolic computation software can be possibly used to leverage some of the algebraic difficulty.
Conclusion
We study a dynamical model characterizing the spread of computer viruses over the Internet. It is assumed that all infected computers possess infectivity, and latent computers have a lower cure rate than seizing computers. As we know, a computer user might try to clear viruses spontaneously even if he is not sure that viruses are staying in his computer possibly because:
-
1.
he is accustomed to running antivirus program regularly, or
-
2.
he is informed that viruses are spreading over the Internet.
In this paper, the global stability of a computer virus propagation model, which incorporates the two features mentioned above, is investigated. One major difficulty in studying the qualitative properties of this model lies in the construction of suitable Lyapunov functions, so that its success largely depends on trial and error as well as on specific problems. By combining this classical approach with the Volterra–Lyapunov matrix analysis, we have leveraged the difficulty of determining specific coefficient values, and as such, wider application of Lyapunov functions to dynamical systems could be promoted. The method Volterra–Lyapunov stability in this work is applied for a model of a computer virus propagation model. The analytical expressions of the stability analysis are provided and their numerical implementation is discussed.
Most of us thought that the problem about propagation of computer viruses should be discussed on network; thus, the diffusion can be considered as well (Ma et al. 2016; Song et al. 2016; Yang and Yang 2017; Satorras et al. 2015; Yang et al. 2017). Within our proposed oscillator-like epidemic model, the spatial effect is homogenized and considered using the variables using mean field theory. In the view of network, for example, regular network with nearest-neighbor connection, small-world type, each node will suffer from external forcing and stimuli from other nodes, which can be regarded as external stimuli with diversity. The local kinetics is critical for collective behaviors of the network. Our results in this model could be helpful for further discussion on collective transition of safety and propagation of computer viruses. In addition, we wish this problem can be further discussed on network in the future.
References
Bhunu CP, Mushayabasa S (2013) Assessing the effects of drug misuse on HIV/AIDS prevalence. Theory Biosci 133–2:83–92
Bradley JT, Gilmore ST, Hillston J (2008) Analysing distributed Internet worm attacks using continuous state-space approximation of process algebra models. J Comput Syst Sci 74(6):1013–1032
Britten NF (2003) Essential mathematical biology. Springer-Verlag, Berlin
Chavez CC, Feng Z, Huang W (2002) On the computation of \(R_0\) and its role on global stability. Math Approaches Emerg Reemerg Infect Dis Intro IMA 125:229–250
Chong NS, Tchuenche JM, Smith RJ (2014) A mathematical model of avian influenza with half-saturated incidence. Theory Biosci 133–1:23–38
Cross GW (1978) Three types of matrix stability. Linear Algebra Appl 20:253–263
Driessche VD, Watmough J (2002) Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math Biosci 180(1):29–48
Han X, Tan QL (2010) Dynamical behavior of computer virus on internet. Appl Math Comput 217:2520–2526
Imran M, Rafique H, Khan A, Malik T (2014) A model of bi-mode transmission dynamics of hepatitis C with optimal control. Theory Biosci 133:91–109
Kephart J, White S (1991) Directed-graph epidemiological models of computer viruses, in. IEEE Symp Secur Pri 1991:343–359
Li MY, Graef JR, Wang L, Karsai J (1999) Global dynamics of a SEIR model with varying total population size. Math Biosci 160:191–213
Li J, Knickerbocker P (2007) Functional similarities between computer worms and biological pathogens. Comput Secur 26(4):338–347
Liu S, Ruan S, Zhang X (2015) On avian influenza epidemic models with time delay. Theory Biosci 134–3:75–82
Ma J, Ying X, Guodong R, Wang C (2016) Prediction for breakup of spiral wave in a regular neuronal network. Nonlinear Dyn 84:497–509
Mishra BK, Saini DK (2007) SEIRS epidemic model with delay for transmission of malicious objects in computer network. Appl Math Comput 188:1476–1482
Murray WH (1988) The application of epidemiology to computer viruses. Comput Secur 7(2):130–150
Piqueira JRC, Devasconcelos AA, Gabriel CECJ, Araujo VO (2008) Dynamic models for computer viruses. Comput Secur 27(7–8):355–359
Redheffer R (1985) Volterra multipliers I. SIAM J Algebraic Discrete Methods 6:592–611
Redheffer R (1985) Volterra multipliers II. SIAM J Algebraic Discrete Methods 6:612–623
Ren JG, Yang XF, Zhu QY, Yang LX, Zhang CM (2012) A novel computer virus model and its dynamics. Nonlinear Anal Real World Appl 13:376–384
Rinaldi F (1990) Global stability results for epidemic models with latent period. IMA J Math Appl Med Biol 7:69–75
Satorras RP, Castellano C, Mieghem PV, Vespignani A (2015) Epidemic processes in complex networks. Rev Mod Phys 87(3):925. doi:10.1103/RevModPhys.87.925
Song X, Wang C, Ma J, Ren G (2016) Collapse of ordered spatial pattern in neuronal network. Phys A 451:95–112
Wang J, Liao S (2012) A generalized cholera model and epidemic-endemic analysis. J Biol Dynam 6:568–589
Wang FW, Zhang YK, Wang CG, Ma JF, Moon SJ (2010) Stability analysis of a SEIQV epidemic model for rapid spreading worms. Comput Secur 29:410–418
Xu R, Ma Z (2010) Global stability of a delayed SEIRS epidemic model with saturation incidence rate. Nonlinear Dyn 61(1):229–239
Yang LX, Yang X (2015) The impact of nonlinear infection rate on the spread of computer virus. Nonlinear Dyn 82(1):85–95
Yang LX, Yang X (2017) The effect of network topology on the spread of computer viruses: a modelling study. Int J Comput Mathe 94(8):1591–1608
Yang LX, Yang X, Zhu Q, Wen L (2013) A computer virus model with graded cure rates. Nonlinear Anal Real 14(1):414–422
Yang LX, Yang X, Wu Y (2017) The impact of patch forwarding on the prevalence of computer virus: a theoretical assessment approach. Appl Math Modell 43:110–125
Yuan H, Chen GQ (2008) Network virus-epidemic model with the point-to-group information propagation. Appl Math Comput 206:357–367
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interests regarding the publication of this article.
Appendix
Appendix
Proof of \(det(-P) > 0\):
Since \(\widetilde{P^{-1}}_{11}=0,\) and \(\widetilde{P^{-1}}_{12}<0\), we have \(det(-P) > 0\).
Hence, it is clear to see \(det(-E) > 0\). The proof is then complete. \(\square\)
Rights and permissions
About this article

Cite this article
Parsaei, M.R., Javidan, R., Shayegh Kargar, N. et al. On the global stability of an epidemic model of computer viruses. Theory Biosci. 136, 169–178 (2017). https://doi.org/10.1007/s12064-017-0253-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12064-017-0253-2