
Abstract
Alzheimer’s disease (AD) is the most common form of dementia, which is progressively affecting elderly people. The dyshomeostasis of biometals and accumulation of toxic metals are usually observed in numerous neurodegenerative diseases including AD. In the central nervous system, metal imbalance–caused neurotoxic activities are usually linked with decreased enzymatic activities, increased aggregation of proteins, and oxidative stress, where a series of processes can result in neurodegeneration and cell death. Even though the relations between neurodegenerative diseases and biometal imbalance are still elusive, there is a growing interest in a group of major endogenous proteins that are associated with the transports of metals. Aberrant expression of these endogenous proteins is associated with the biometal imbalance and AD pathogenesis. Indeed, heavy metals are extremely toxic to the nervous system. Various studies have demonstrated that the toxic effects of heavy metals can result in amyloid beta (Aβ) aggregation, neurofibrillary tangles, and even loss of neurons. In this article, we have focused on the molecular processes through which exposure to biometals and toxic metals can play roles in AD pathogenesis.
Similar content being viewed by others
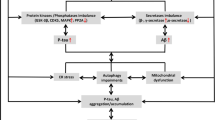

Avoid common mistakes on your manuscript.
Introduction
Alzheimer’s disease (AD) is a chronic and irreversible neurodegenerative disease associated with dementia in elderly people [1, 2]. In AD brain, neuropathological alterations are associated with the amyloid-beta (Aβ) aggregation which generates senile plaques (SPs) and primarily results in various consequences, for example, hyperphosphorylated aggregates of the microtubule-associated tau protein in neurofibrillary tangles (NFTs), impaired neuronal connectivity, and loss of neurons [3,4,5]. Numerous studies have extensively analyzed Aβ’s structure and its harmful effects in inducing oxidative stress (OS), autophagy, and neuroinflammation [6,7,8]. To treat AD, several drug candidates were developed to eliminate or decrease Aβ generation [9,10,11]. Nevertheless, most of them failed in clinical trials [12, 13]. In recent times, it has been revealed that Aβ aggregation is not regarded as the initial event of AD pathogenesis; rather, Aβ aggregation is a subsequent event of the disease [10, 14]. Thus, new research approaches are required to discover effective AD treatments. Several studies have demonstrated that homeostasis of essential biometals (for example calcium, magnesium, manganese, copper, zinc, and iron) is impaired in case of AD. Furthermore, these metals contribute significantly to the metabolism and aggregation of tau and Aβ. Depending on these findings, a metal hypothesis for AD has been proposed [12, 15], which is suggesting that targeting the interactions of metals with Aβ may prove more effective in AD prevention.
Several studies have demonstrated the pathophysiological impacts of metal imbalance in the brain [16, 17]. In a study, Akhtar et al. [18] revealed that chromium picolinate treatment attenuated streptozotocin-induced cognitive impairment. Furthermore, treatment with chromium picolinate reversed pathology of AD, as demonstrated by enhanced memory, decreased oxidative damage, mitochondrial dysfunction, neuroinflammation, and upregulated insulin signaling [18]. Nonetheless, still, there are arguments regarding impaired biometal activity as the causative factor for AD. The presence of the blood-brain barrier (BBB), makes it difficult to treat the brain diseases [19, 20]. As the BBB cannot be passively penetrated by the biometals, thus the mentioned metal imbalance in the AD brain cannot only be associated with the decreased or increased exposure to metals, but rather to a more initial distribution of intracellular ions in an unclear manner. Therefore, brain metal homeostasis is regulated by several metal exporters, importers, and metal sequestering proteins.
It has been demonstrated that heavy metal accumulation in the human body can be harmful for multiple organs. Particularly, heavy metals are well-known to exert toxic effects on the brain. Several studies have particularly focused on the neurological functions of cadmium, mercury, and lead in the brain [21]. In this article, we have focused on the biometals, heavy, and non-essential metals–induced molecular processes of AD.
Biometals and Alzheimer’s Disease
Iron
Iron (Fe, an essential trace metal) is involved with several vital neuronal functions in the brain, including synthesis of myelin, mitochondrial respiration, and transport of oxygen [22]. Furthermore, iron also plays a role as a cofactor for numerous metalloproteins associated with signal transduction and metabolism [23, 24]. Numerous studies also found increased levels of iron in AD brains [25, 26], particularly in putamen and globus pallidus areas [27, 28]. Nonetheless, as compared to healthy individuals, studies observed reduced or unchanged levels of iron in the serum of AD individuals [29, 30]. It has been revealed by a meta-analysis that there is a significantly decreased level of iron in the serum of AD individuals as compared to healthy controls. Nonetheless, iron level in CSF was not influenced by AD; however, further analyses are essential because of the relatively small number of CSF studies carried out till now [29]. Scientists have carefully studied iron content in 12 selective areas of the brain via separated meta-analyses by utilizing cross-referenced statistical methods [29]. They also observed that 8 certain areas of the brain contained an elevated level of iron that correlated with the clinical diagnosis of AD in a statistically validated manner. Indeed, these findings provided rigorous statistical support for the model that iron homeostasis was altered in individuals with AD, along with the finding of lower iron in their serum and evidence for iron overload in various specific areas of the brain [29]. Nevertheless, meta-regression analyses showed in several studies that differences in iron levels in serum might be owing to the different mean ages [30]. Unfortunately, clarification of this aforesaid imbalance is still not known. Studies also showed that in case of AD, excessive levels of iron can stimulate the production of hydroxyl radical by Fenton reaction, which can eventually lead to an elevated level of oxidative stress in AD.
Dynamic relationship between efflux and influx of iron is important to maintain homeostasis of intracellular iron, where various transporter proteins have significant contributions. In AD individuals, impairment of iron exporter ferroportin (FPN), along with iron importers, such as melanotransferrin (MTf), lactoferrin (Lf), divalent metal transporter 1 (DMT1), and transferrin (Tf), can significantly play role in the accumulation of iron in the affected areas of the brain (Fig. 1). Except for oligodendrocytes, DMT1 is expressed on microglia, astrocytes, and neurons. DMT1 is associated with the pathway involved in Fe2+ influx [31]. In the AD brain, DMT1+IRE (iron-responsive element) and DMT1-IRE are the 2 isoforms of DMT1 that were found to colocalize with Aβ in the plaques. In amyloid precursor protein (APP)/presenilin-1 (PS1) transgenic mouse model, these 2 isoforms of DMT1were also found to be elevated in the hippocampus and frontal cortex regions, along with a decreased level of FPN expression [32], which is further indicating that the dysregulated iron metabolism–associated protein DMT1 and FPN have significant contribution in the iron-facilitated AD neuropathogenesis. Hepcidin (a protein, involved in iron homeostasis) is co-located with FPN in astrocytes, and neurons exhibited a decreased level of FPN expression in AD brains. It has been observed that the downregulation of hepcidin can lead to the impaired iron export pathway which can eventually lead to retention of cellular iron [33].

The role of iron in the pathogenesis of Alzheimer’s disease. The ferrous form of iron (Fe2+) can directly enter into the cell via DMT1, whereas transferrin (Tf)-ferric iron (Fe3+) enters via the transferrin receptor (TfR)-facilitated endocytosis. Elevated Fe2+ levels stimulate the Fenton reaction to generate hydroxyl radical (•OH), which eventually results in oxidative stress and neurodegeneration. Furthermore, Fe2+ can elevate phosphorylation of tau via activation of glycogen synthase kinase 3β (GSK3β) and cyclin-dependent kinase 5 (CDK5) that lead to the formation of neurofibrillary tangles (NFTs). The iron chelators reduce the phosphorylation of tau by inhibiting the GSK3β and CDK5. In the cellular environment, Fe2+ binds with the iron-responsive element (IRE) in the 5′ UTR region of amyloid precursor protein (APP) mRNA, which results in the induction of APP translation that leads to the formation of amyloid beta (Aβ). This generated Aβ can interact with Fe2+ as well as increase Aβ aggregation. Fe2+ can only go out of the cell by using ferroportin (FPN) along with the activity of haptoglobin or ceruloplasmin. APP can also interact with FPN to oxidize Fe2+ into Fe3+ for Tf binding. Nevertheless, the binding of hepcidin (HP) with FPN leads to its internalization to avert the export of Fe2+
APP can catalytically oxidize Fe2+ to Fe3+ (Fig. 1) through the activity of ferroxidase and subsequently interact with FPN to mediate the export of iron [34]; nevertheless, this mechanism is suppressed through extracellular zinc (Zn), which originates from Zn-Aβ complexes [35]. In the same study, researchers exhibited that loss of soluble tau can result in retention of iron via weakening APP-induced export of iron [36], and such inhibition can be achieved by the use of lithium [37] or an iron chelator [38]. In addition to this, sirtuin 2 controls homeostasis of cellular iron through deacetylation of nuclear factor erythroid-derived 2-related factor 2, which functions as a transcription factor to regulate the expression of FPN [39]. In a study, it was found that compounds derived from Chinese herbs can decrease the expression of DMT1 and can increase the expression of FPN, which is suggesting a new approach to reduce iron overload-mediated impairment in AD [32]. In BBB’s endothelial cells, Tf-transferrin receptor (TfR) complex was found to play a role in iron uptake. Iron transport across BBB can take place owing to receptor-facilitated endocytosis of Tf-bound iron [40]. A study showed that cerebrospinal fluid (CSF) levels of Tf were markedly different in case of familial AD (FAD) when compared between individuals who carried mutations and related non-carriers [41]. Lf’s structure is similar to the structure of Tf, where both of these iron importers have 2 lobes, in which each lobe contains a binding site for Fe3+ (Fig. 1) [42]. In AD patients, Lf expression is high in macrophages/monocytes and fibrillar-type SPs in the cerebral cortex region of individuals with AD [43]. Furthermore, the formation of SPs mediates the age-related deposition of Lf [44]. Interestingly, lipoprotein receptor–related protein (LRP, a cell surface receptor) is associated with the clearance of Aβ by an endocytic process. Furthermore, Lf can bind with LRP and can significantly increase soluble Aβ clearance instead of Aβ generation [45]. A liposomal system involving surface Lf was developed for the delivery of neuron growth factors through the BBB. Indeed, the aforesaid technique was useful to control AD progression [46, 47].
Copper
Copper (Cu, an essential trace metal) is also involved with various important cellular activities, such as it plays role as a structural component of enzymes that are essential for antioxidant defense and energy metabolism [48]. Association of copper with AD pathophysiology is complex. Increased copper levels have been identified in SPs [49]. In AD brain, a deficiency in the total copper levels in the AD brain has been reported by some studies [50]. Furthermore, another study revealed that even though the combined serum and plasma copper level was higher in AD individuals [51], however, total CSF levels of copper were not different when compared between AD individuals and healthy subjects [52, 53]. The reason for this heterogeneity is that a substantial amount of copper precipitates with SPs in AD-affected areas, which can further lead to a deficiency of copper in other areas. Indeed, copper can interact with both tau and Aβ and can exacerbate their pathological outcomes [54, 55].
Still, the processes associated with copper dislocation in the AD brain are not clear. Copper transporter 1 (CTR1) and the copper transporting P-type ATPases, such as ATPase copper transporting alpha and beta (ATP7A and B), are the main transporters associated with the cellular regulation of monovalent copper [56, 57] (Fig. 2). It was reported that DMT1 might contribute to divalent copper delivery into cells to synthesize copper-containing enzymes [58]. However, in terms of copper overload, dual-roles are played by ATP7A and ATP7B in the export of excess copper out of cells in an adenosine triphosphate (ATP) hydrolysis–dependent manner. Along with the transporters, a group of intracellular proteins, known as molecule chaperones, for example, copper chaperone for superoxide dismutase (SOD), cytochrome oxidase enzyme complex, and antioxidant protein-1, also play roles in copper delivery to certain targets [58, 59]. Interestingly, the genetic knockdown of copper transporter 1C (CTR1C) in a Drosophila model of AD markedly decreased the accumulation of copper in the brain [60]. Similar findings were also seen in flies when DmATP7 (a copper exporter) was increased, or when CTR1B (a copper importer) was suppressed in AD flies. Furthermore, these flies showed elevated Aβ generation but a decrease in Cu-Aβ complex-mediated OS, indicating that Aβ oligomers, or the elevated levels of Aβ aggregates, were less toxic in a decreased influx of copper-induced by CTR1 knockdown [60]. In an AD mouse model, it was observed that ATP7A can be increased in activated microglial cells where the amyloid plaques are gathered, which can further lead to a substantial change of copper trafficking in microglia. This finding suggests a neuromechanism, where inflammation-mediated copper dyshomeostasis in microglia is linked with AD [61]. On the other hand, genetic studies showed that a cohort analysis of a single nucleotide polymorphism in ATP7B is responsible for impairments in circulating non-ceruloplasmin-bound copper which can elevate the AD risk, which further indicates that alterations in copper homeostasis might speed up the neurodegeneration process which can lead to AD [53, 62].

The role of copper in the pathogenesis of Alzheimer’s disease. In the human body, copper is most commonly shifting between cuprous (Cu+) and cupric (Cu2+) forms. Cu+ enters into the brain cells via copper transporter 1 (CTR1), whereas Cu2+ uses divalent metal transporter 1 (DMT1) to enter into the brain cells. Cu+ accumulation is sequestered into specific locations in the cells via several chaperones of copper such as antioxidant protein-1 (ATOX1). ATOX1 plays a role in the Cu+ transfer to ATPase copper transporting alpha and beta (ATP7A and B), which also aids the Cu+ import into synaptic vesicles for release and/or directly facilitate copper efflux that interacts with amyloid beta (Aβ). Excessive levels of intracellular Cu2+ might cause activation of Fenton reaction to elevated oxidation of biomolecules. In addition, Cu2+ is associated with the activation of GSK3β that eventually leads to hyperphosphorylation of tau. Cu2+ also increases the phosphorylation of amyloid precursor protein (APP) and generate Aβ, which results in the formation of senile plaques
Zinc
Zinc (Zn, an essential trace metal) is an important constituent of about 100 s of proteins and enzymes [63]. This trace metal is also extensively associated with cell signaling as compared to other metals, mainly because it can also play a role as a neurotransmitter [64]. Since a former study reported zinc redistribution into extracellular SPs, therefore functions of zinc have been widely studied in AD pathogenesis (Fig. 3) [65]. In AD individuals, inconsistent findings have been observed with the levels of zinc. However, numerous studies have reported increased zinc levels in the CSF and brains of AD individuals [66]. Other studies observed reduced or even no difference in the levels of zinc in the serum and brain of AD individuals as compared to controls [30, 67, 68]. Still, the exact cause of such diverse findings with the levels of zinc is not known. Henceforth, the development of zinc as an AD treatment is hindered. It is known that zinc binds with the histidine residues in the Aβ’s C-terminus and promotes the formation of aggregates. As compared to copper and iron, zinc has a greater affinity to bind with Aβ in a wide pH range [69].

The role of zinc in the pathogenesis of Alzheimer’s disease. In the plasma membrane, zinc importing protein (ZIP) mainly regulates the zinc (Zn2+) entry, while zinc transporter (ZnT) regulates the efflux of Zn2+. An elevated level of Zn2+ increases the accumulation of Aβ, tau modification as well as increase the formation of reactive oxygen species (ROS). Conversely, lower levels of Zn2+ reduce the bioavailability of Zn2+ and amyloid beta (Aβ) clearance as well as cause synaptic dysfunction
Homeostasis of zinc in neurons is mainly regulated by 3 groups of transporters including metallothioneins (MTs), zinc-regulated transporter-like, and iron-regulated transporter-like proteins (ZIPs), and zinc transporters (ZnTs) (Fig. 3). It has been found that ZnTs mediate the efflux of zinc from cells or facilitate excessive zinc from the cytoplasm into intracellular vesicles and organelles [70]. Functions of ZIPs are nearly opposite to the roles of ZnTs. ZIPs mediate the import of zinc into cells or facilitate zinc movement from intracellular vesicles into the cytoplasm [70, 71]. MTs also play roles in the maintenance of zinc homeostasis and regulate the cellular levels of zinc and other related signaling mechanisms [72]. In the brains of AD individuals, immunofluorescence studies confirmed that various ZnTs (including ZnT1, 3, 4, 5, 6, and 7) are widely present in Aβ plaques of cortex region [73]. Among them, ZnT3 is mainly found on the synaptic vesicles of zinc-containing glutamatergic neurons [74]. ZnT3 levels reduce in elderly people with aging, especially in AD individuals [75].
Furthermore, an age-dependent impairment in cognitive functions has been observed in ZnT3 knockout mouse models [76]. However, in the ZnT3 deficit mouse models (while overexpressing APP), reduced levels of plaque burden and synaptic zinc were observed [77], which further indicating the role of synaptic zinc in amyloid plaque deposition in case of AD. In AD, ZnT3 knockout-mediated elevated levels of intra-neuronal zinc worsened the damages of neurons [49]. However, more studies are required on the mechanisms related to zinc reuptake into the presynapses. ZIPs play vital roles in facilitating the influx of zinc, and other processes may also indirectly contribute in this regard including presenilins (presenilin mutation is known as a causative factor for FAD) [78]. Moreover, MTs are the main zinc-buffering peptides that play roles in the maintenance of cytosolic zinc balance. MT-1 to MT-4 are the 4 major isoforms of MT that are found to be expressed in brains. In AD individuals, level of MT-1 and MT-2 increases, while the MT-3 level decreases [79]. Deficiency in MT-1/2 reduced the amyloid plaque burden in an animal model of AD and therefore recovered the APP-mediated alterations in mortality [80].
Interestingly, MT-3 is linked with the aggregation of Aβ via cysteine oxidation. The deficiency of MT-3 was found to partially rescue the APP-mediated mortality of females and resulted in alterations in APP-mediated behavioral phenotypes of mouse models [81]. Following the polymerization of actin, MT-3 controls the uptake of Aβ in astrocytes via its positive activity [82]. Studies involving spectroscopy and microscopy confirmed that MT3 averts Cu-Aβ-induced neurotoxic activities perhaps through a metal exchange in between aggregated Cu-Aβ (1–40) and Zn-MT-3, which can further result in inhibition of reactive oxygen species (ROS) generation [83]. A different study reported the protective functions of MTs (derived from astrocytes) on primary cortical neurons against Aβ toxicity via suppressing the generation of proinflammatory cytokines, increasing the B-cell lymphoma 2 levels, and decreasing contents of ROS [84].
Zinc has been found to induce tau aggregation under reducing conditions [85]. Interestingly, zinc suppressed the intramolecular disulfide bond formation but mediated intermolecular bonds between important cysteine residues. In addition to this, exposure to zinc elevated the phosphorylation of phosphoinositide 3 kinase and mitogen-activated protein kinase-dependent pathways which are vital for tau modifications [86]. For a better understanding of the AD-linked zinc dyshomeostasis, more studies are required to assess the functions of zinc transporters in AD pathogenesis.
Manganese
Manganese (Mn, an important trace element) exerts various vital physiological functions for intracellular homeostasis and growth [87]. Manganese plays an important role as a cofactor for important enzymes that are associated with normal cell function, for instance, glutamine synthetase and SOD. There is growing evidence that indicates that an overload of manganese is linked with neurodegenerative diseases and a slight increase in the level of manganese can trigger symptoms that are related to manganese poisoning [88]. Processes that are linked with manganese-induced cytotoxicity include reduced cellular antioxidant defense and autophagy, buildup of intracellular toxic metabolites, aberrant energy metabolism, mitochondrial dysfunction, and ROS overgeneration [89, 90].
Significantly increased level of manganese was found in the brain of AD individuals with dementia in comparison with the healthy subjects, while the maximum manganese level was found in the parietal cortex [91, 92]. This finding indicates that an overload of manganese might be associated with the cognitive deficit and AD pathology. Chronic exposure of manganese altered gene expression which dispersed Aβ plaques in non-human primates. Interestingly, p53 mostly targeted the altered genes; amyloid-beta precursor-like protein 1 (APLP1) was one such gene, which was found to be the major upregulated gene in the frontal cortex [93]. Exposure to manganese mainly affects the frontal cortex which can lead to incipient dementia [94]. Treatment with manganese elevated Aβ levels both in vitro and in vivo; the associated process is perhaps linked with the interruption of Aβ degradation [92]. In a study, it was revealed that manganese may weakly bind with the specific Aβ sites [95]. Nonetheless, more studies are required to demonstrate the effects of such manganese binding with Aβ in mediating Aβ aggregation. Indeed, manganese is a constituent of manganese (Mn) SOD (Mn-SOD), which is an antioxidant enzyme that significantly contributes to preserving the vitality of mitochondria. Elevated manganese levels can hinder oxidative respiration, which can increase ROS generation and eventually can result in mitochondrial dysfunction [96]. In a transgenic mouse model of AD, partial Mn-SOD deficiency elevated Aβ plaque deposition and phosphorylation of tau [97, 98]. However, Mn-SOD overexpression exerted various benefits against the pathology of AD via decreasing the burden linked with cortical plaques [99], which further confirmed the associations between the AD pathophysiology and mitochondrial oxidative stress. Moreover, manganese toxicity triggered cognitive impairment in humans, and it has been hypothesized that high level of manganese uptake can result in deficiency of iron in the Golgi apparatus, which is in line with the finding that iron and manganese compete with the same transport processes and binding sites, at least to some degree [100].
Manganese transport is facilitated by several importers, including dopamine transporter, ZIP14, ZIP8, Tf/TfR, DMT1, and also via multiple exporters including FPN, SLC30A10 (solute carrier family 30 member 10), park9/ATP13A2, and the secretory pathway Ca2+-ATPase 1 (SPCA1). Among them, DMT1 mediates iron influx and this transporter is the first mammalian transporter for cellular uptake of manganese. It has been found that DMT1 mediates manganese movement across the BBB, mainly under the conditions of iron deficiency [101]. Through a ligand-receptor endocytosis process, DMT1 transports divalent manganese, whereas Tf transports trivalent manganese into cells via a ligand-receptor endocytosis process [102]. ZIP14 and ZIP8 possess increased binding capacity with zinc; however, multiple studies revealed that these transporters are also associated with the manganese absorption from the lungs and liver [103,104,105]. The contribution of exporter proteins in the maintenance of manganese levels has also been studied. It has been identified by genome analysis that SLC30A10 might transport both manganese and zinc. Manganese accumulation has been detected in the carriers of mutations in SLC30A10 and individuals with Parkinson’s disease (PD) [106]. In the frontal cortex of APP/PS1 transgenic mouse models and AD individuals, the level of SLC30A10 was found to be markedly decreased, which further indicates that its dysregulation can play role in AD pathology [107]. In a pH-dependent manner, FPN (an iron exporter) can play a role as a cellular exporter of manganese to attenuate cytotoxicity and manganese accumulation [108, 109]. Moreover, ATP13A2 (ATPase cation transporting 13A2) plays a role as a cation transporter in the transportation of zinc and manganese. It has been revealed by studies that ATP13A2 overexpression decreases the intracellular level of manganese, which as a result can alleviate manganese-induced lethality; loss-of-function mutations in ATP13A2 are associated with the rise in both Aβ plaques and α-synuclein in Lewy body disease [110]. In addition to this, a SPCA1 homolog in yeast, plasma membrane ATPase-related 1, mediated the transport of manganese and Ca, and ectopic SPCA1 expression in yeast increased sensitivity to manganese toxicity [111]. Therefore, it has been indicated that SPCA1 functions as another regulator for cellular manganese homeostasis. However, further studies are required to investigate the affinity between manganese and SPCA1 and the functions of SPCA1 in the pathogenesis of AD.
Magnesium and Calcium
Magnesium (Mg, a major macro element) plays crucial roles in several enzymatic synthesis and cellular mechanisms such as synaptic plasticity, ion channels, and energy metabolism [112, 113]. Calcium (Ca, a major macro element) plays a role as a ubiquitous second messenger and its role in regulating cellular function has been extensively studied [114]. Levels of intracellular calcium are strictly controlled via several calcium-binding proteins, calcium channels, pumps, and are also controlled by other metal ions including magnesium. It has been found that magnesium can act as a calcium antagonist. Magnesium also plays a role in the maintenance of intracellular calcium concentrations and protecting neurons from excitatory responses mediated via calcium overload, under physiological conditions [115]. Nonetheless, disruptions in calcium and magnesium homeostasis change a series of processes that can result in various diseases including neurodegeneration [116].
Magnesium levels in serum and brain are considerably lower, while levels of calcium were found to be considerably increased in AD individuals as compared to age-matched healthy subjects [117,118,119]. Frequently elevated calcium concentrations can lead to the raised expression of ApoE and APP and can also mediate the generation of Aβ aggregation via a mechanism involving γ-secretase stabilization [120, 121], while Aβ aggregation can lead to altered membrane calcium permeability that can further worsen AD [122]. Some studies have also assessed the contribution of magnesium in AD pathogenesis. It has been revealed by an in vitro study that both magnesium and calcium can induce the mechanism of hyperphosphorylated tau aggregation [123]. Magnesium-l-threonate administration elevated concentrations of magnesium in the brain, which resulted in reduced β-secretase (BACE1) levels thus decreased the β c-terminal fragments and soluble APP levels, therefore alleviated the cognitive impairments and synaptic loss linked with Alzheimer’s symptoms [124]. Moreover, magnesium sulfate treatment decreased hyperphosphorylated tau levels via suppressing the phosphorylation of glycogen synthase kinase 3β (GSK3β) and elevating the activity of phosphatidylinositol 3 kinase (PI3K) and protein kinase B (Akt) [125, 126], therefore indicating that magnesium can play a role as a neuroprotective factor in AD development. The underlying mechanism associated with magnesium blocking the long-term activation of N-methyl- d-aspartate (NMDAR)-induced calcium influx and therefore decreasing calcium-mediated neuroinflammation. Indeed, NMDARs are cationic channels that are activated by glutamate, containing an increased permeability to calcium ions upon the synaptic activity, for instance, memory and learning. Aβ aggregation–mediated NMDAR over-activation might take place during the early AD stages [127]. Intracellular calcium concentrations can be increased by continuous calcium influx, which can further trigger numerous enzymatic processes that can lead to neuronal death, protein destruction, and peroxidation [128]. As an endogenous blocker, magnesium can bind with the NMDAR subtypes including NR1/2A and NR1/2B, which are components of NMDARs present in brain areas affected by AD, under normal conditions [129]. Blocking channels by the addition of magnesium decreased influx of calcium into post-synaptic neurons, to decrease excitotoxic cell death during dementia. In case of neurodegenerative diseases, activation of ATP-gated P2X purinergic receptors (P2XRs) linked with neuroinflammation has also been reported [130]. P2X7R can help in the formation of an oligomer to form membrane pores in microglia to facilitate the influx of calcium [131]. It has been revealed by using tissue culture that magnesium can decrease intracellular calcium levels via P2X7R, and that improved stimulation of purinergic receptor-activated neuroinflammation, which further indicates that elevated magnesium level can play role as an effective calcium entry inhibitor via cell surface channels [132].
Various factors including buffering proteins, exchangers, channels, and numerous transporters are associated with the maintenance of cellular calcium and magnesium homeostasis. Various channels can facilitate the influx of magnesium into cells, for instance, transient receptor potential melastatins 6 and 7 (TRPM6/TRPM7), magnesium transporter 1 (MagT1), and cyclin M (CNNM) transporter. Solute carrier family 41 member 1 (SLC41A1) and sodium-independent magnesium exchanger are essential to favor magnesium extrusion [133, 134]. In addition to this, intracellular calcium balance can be attained via several calcium transporters. NMDAR, store-operated calcium channels, voltage-gated calcium channels, and α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptors (AMPARs) are accountable for the increased level of calcium. On the other hand, calcium-binding buffering proteins including calbindin can mediate calcium storage in the endoplasmic reticulum (ER), while the activities of the calcium-ATPase pump and sodium-calcium exchanger facilitate calcium export out of cells. Indeed, mutant presenilins activate 2 types of calcium receptors and that plasma membrane calcium-permeable channels permit the leakage of calcium ions from the ER into the cytoplasm, therefore triggering a vital effect on ER-Ca dynamics in the AD brain [135, 136]. Furthermore, Aβ oligomers can either induce the formation of calcium-permeable channels or bind with NMDARs, therefore can mediate the entry of calcium through the plasma membrane [137, 138]. Nonetheless, the contribution of magnesium transporters in AD pathology is poorly understood. The physiological function of TRPM7 was found to be coordinated by presenilins, the mutation of which can result in familial AD [139]. TRPM2 removal in APP/PS1 mouse models ameliorated age-dependent memory impairments and decreased ER stress, while in vitro studies revealed that the knockdown of TRPM2 blocked the Aβ-induced rise in the magnitude of whole-cell current, therefore suggesting the significance of TRPM2 effect in Aβ neuronal toxicity [140]. It was also mentioned that TRPM2 alteration can result in calcium imbalance, even though its contribution in the regulation of magnesium linked with AD was overlooked [140]. Moreover, PD dementia and amyotrophic lateral sclerosis were found to be linked with lower levels of magnesium and calcium as compared to healthy subjects, which is further suggesting the dysfunction of TRPM2 and TRPM7 channels [141].
Pathogenic Mechanisms of Heavy Metal–Induced Alzheimer’s Pathology
Various studies have revealed the heavy metal-induced molecular mechanisms that are involved in AD pathogenesis. In this section, we have explained the molecular processes and analyzed the signaling mechanisms based on molecular networks linked with various heavy metals (cadmium, mercury, and lead) mediated AD pathogenesis. Furthermore, we have also discussed the connections of various molecular objects with the signaling pathways (Table 1).
Cadmium
Indeed, Aβ aggregation is an important hallmark of AD pathogenesis [172,173,174]. Interaction between cadmium (Cd) and Aβ is associated with elevated AD risk [172, 175, 176]. Certain metals are also involved with the NFTs formation [172, 177]. Cadmium also can interact with Aβ1–42 [172]. It is known that Aβ1–42 is an important component of SPs that contributes significantly in AD pathogenesis [178,179,180]. Cadmium is also associated with the Aβ aggregation (Fig. 4) [172, 181]. In line with this finding, tyrosine and histidine residues located at the N-terminal part of the peptide and the binding blocked ion channel of Aβ1–42 [172, 181]. AD risk can also be increased via decreased or elevated expression of certain proteins and these alterations are induced by the exposure to cadmium [182]. For instance, M1 receptor can be blocked by cadmium, which can result in AChE (acetylcholinesterase)-R downregulation and AChE-S overexpression [182]. In basal forebrain, cadmium was also found to activate the cell death of cholinergic neuronal cells [182]. This finding is similar to the mechanism involved in AD-related brain degeneration [182]. Interestingly, these symptoms are linked with elevated levels of Aβ, GSK3β, and formation of tau filament (Fig. 4) [182]. Indeed, GSK3β is an important constituent of tau paired helical filaments, which is located in the deposits of NFTs that disturb functions of neurons, and it is used as a marker for AD-related neurodegeneration [183].

The role of cadmium in the pathogenesis of Alzheimer’s disease. Cadmium (Cd) induced the aggregation of amyloid beta (Aβ) oligomers or fibrils by blocking the Aβ1–42 ion channel. Furthermore, Cd blocks M1 receptor that causes overexpression of AChE-S and downregulation of AChE-R and finally increase the expression of GSK3β that leads to hyperphosphorylation of tau
As compared to healthy subjects, potential neurotoxic activity of cadmium has been detected owing to increased levels detected in plasma [184], liver [185], and brain tissues [67] of AD individuals [172]. Elevated cadmium levels were also identified in the hair and blood of AD individuals. Cadmium-exposed workers experienced neurobehavioral problems in memory, psychomotor speed, and attention [186]. Like humans, Li X et al. [187] confirmed that APP/PS1 transgenic mouse models exhibit symptoms of the ethological disorder including memory and learning following exposure to cadmium. Indeed, these observed symptoms are AD characteristics. It is considered that cadmium may play roles in AD and this heavy metal might be involved with the generation of NFTs and Aβ aggregation [172].
Mercury
The tubulin proteins polymerize into long chains or filaments that form microtubules [188, 189]. Mercury (Hg) suppresses the effects of tubulin, which results in neuronal damage and eventually AD [190]. This protein shows a very high affinity towards mercury and following binding this metal-ligand, the structural integrity of tubulin is affected, which leads to a suppression of polymerization of tubulin to micro-tubulin, which further leads to the generation of NFTs and SPs [190]. Indeed, these NFTs and SPs are characteristic features in the brains of AD individuals [190]. Activities of mercury were analyzed in studies involving animal neuronal cell experiments. In these studies, degeneration of axon and formation of NFTs were also reported [191]. A study on stem cells showed that mercury played role in neuronal apoptosis and this metal also inhibited the activities of tubulin [191]. Furthermore, mercury can cause tau hyperphosphorylation; as a result, stabilization of microtubules in the neuron can be affected [192, 193].
Mercury-induced OS can also affect the phosphorylation state of tau by elevating its level [193, 194]. Indeed, tau is closely linked with Aβ that can exacerbate both Aβ pathology and tau-Aβ interactions in AD [195]. Accumulation of Aβ can trigger tau hyperphosphorylation in AD [195], which further suggests that Aβ accumulation can stimulate the signal transduction pathways for the hyperphosphorylation of tau [195,196,197,198,199]. It was suggested that tau dysfunction might lead to AD pathogenesis [200]. Several studies exhibited that APP gene expression can be affected by mercury [200,201,202]. In a study, Song et al. [203] estimated whether mercury influences Aβ accumulation facilitated by an imbalance between Aβ generation and clearance. In that study, mercury and methylmercury elevated the level of APP, which is associated with Aβ generation. Levels of neprilysin were reduced in PC12 cells by the treatment of mercury and methylmercury. Collectively, these findings indicated that mercury stimulated Aβ accumulation via the overgeneration of APP and NEP reduction [203]. Astrogliosis caused by methylmercury was also observed in case of AD neuropathology, which eventually plays a role in APP expression via glial activation [200, 201, 204,205,206,207].
Increased levels of mercury were also identified in the blood of AD individuals and were also detected in brain tissue [208,209,210]. Increased levels of mercury were also found in hair [211]. Furthermore, as compared to the control group, increased mercury levels were identified in the hair of patients with degenerative brain disease [211]. In the nervous system, mercury can trigger dementia, attention deficits, and memory loss [212, 213]. In a study, Haut et al. [214] examined workers who were exposed to the vapor of mercury. Increased mercury levels were found in the blood of these workers and they also suffered from cognitive impairments.
Lead
Associations between AD and exposure to lead (Pb) have been extensively studied at the molecular level by generating oxidative DNA damage [195, 215]. In the brain, oxidative DNA damage has been identified during the aging process and this damage can also contribute to AD pathogenesis [195, 215,216,217,218]. Following exposure to lead, OS might play a role in the elevation of Aβ levels which can eventually result in oxidative damage of the nervous system [215, 219]. OS-mediated apoptosis is found to be involved with the accumulation of Aβ [220,221,222]. Lead toxicity–mediated oxidative DNA damage might be associated with the imbalance between 8-hydroxyguanosin (oxo8dG) accumulation and the effect of Ogg1 mediating oxidative DNA damage [215, 219]. In a study, Bolin et al. [219] stated that oxo8dG was briefly modified at the early stage of life (postnatal day 5); however, it later increased 20 months following cessation of lead exposure, but the activity of Ogg1 was found to be not altered. Besides, an age-dependent loss in the inverse correlation between accumulation of oxo8dG and Ogg1 activity was noticed. In old age, lead activity on oxo8dG levels did not take place if the animals were exposed to lead. Furthermore, these elevations in DNA damage took place in the absence of any lead-mediated alterations in manganese-SOD2, copper/zinc-SOD1, and reduced form glutathione (GSH). Collectively, these findings suggest that neurodegeneration and oxidative damage in the aging brain might be affected by the developmental disturbances [219]. The generation of oxo8dG owing to oxidative DNA damage is commonly known as a marker for oxidative DNA [215, 219]. Indeed, the imbalance may be associated with the processes of AD pathogenesis [215]. In a study, Wu et al. [223] found that AD-associated genes (BACE1, APP) were upregulated due to the lead exposure [223]. It was observed that snippets of APP caused the formation of Aβ aggregates in AD [224].
In a different study, Bolin et al. [219] revealed that lead exposure in an earlier stage of life resulted in gene alterations via hypomethylation of the APP gene, which is a gene responsive to lead [225]. Furthermore, this hypomethylation triggered APP gene overexpression and resulted in APP protein generation [219, 225]. Since the level of APP becomes high due to the lead exposure, thus the effect of Sp1 (a transcription factor) which controls AD-related proteins is increased [219, 224, 225]. Therefore, the aggregation of Aβ was induced and resulted in plaque formation in the brain [219, 224]. In a mouse hippocampal cell line, increased expression of APP was identified under chronic exposure to low-dose lead [226]. Besides, Wu et al. [223] also confirmed that lead might also induce AD pathogenesis in the monkey. Following lead exposure in the brain of monkeys, these researchers also observed intracellular staining of total Aβ and dense-core plaques via immunohistochemical analysis [223]. The obtained findings suggested that there was a buildup of immunoreactive Aβ aggregates inside neuronal cells and this further indicated the possibility of AD [223].
In the battery industry, occupation-related lead exposure is confirmed from the studies on workers [227, 228]. It has been found that workers of this industry are exposed to an increased level of lead in comparison with the average adults. These workers exhibited various symptoms of psychological dysfunctions including headaches, vertigo, forgetfulness, and paresthesia [229]. Moreover, an increased level of lead has also been detected in the blood [227,228,229]. In a study, Sharma et al. [227] confirmed that the workers of the industry suffered from schizophrenia-associated differential structural problems of the brain. Furthermore, workers in that study exhibited functional differences in brain activity in frontal lobes and hippocampus [227]. Indeed, these symptoms are usually seen in AD individuals. The researchers summarized that lead can trigger neurodevelopmental diseases showing neurocognitive impairments for example AD and schizophrenia [227].
Non-Biologically Relevant Metal
Aluminum
Indeed, the physiological roles of aluminum (Al) are not well-known, but this metal is toxic to organisms [230]. In daily life, we inevitably come into contact with aluminum because of its ubiquitous presence in the environment. Luckily, aluminum compounds that are present in our ingested food items are not dissolved at physiological pH; rather, it is then eliminated from the body. Indeed, aluminum toxicity can take place if increased levels of aluminum are inhaled or ingested. Aluminum accumulates primarily in the frontal cortex and hippocampal regions of the brain, therefore correlating to the impairment of other essential biometals. This can result in oxidative stress and can affect numerous signaling cascades, features which can further result in neuronal death and induction of neurodegenerative diseases [231,232,233].
Former studies stated the aluminum hypothesis, which indicated that exposure to aluminum is associated with AD etiology [234, 235]. Even though the validity of the aluminum hypothesis in AD has been argued for years and is progressively being regarded as only a fringe hypothesis as compared to various other theories in AD research, however, exposure to aluminum still remains important and endures to be the center of interest [236]. It has been reported by an in vivo study that chronic administration of aluminum increased Aβ generation in the cortical and hippocampal areas in rats [237]. In transgenic mouse models of AD, aluminum administration increased Aβ plaques accumulation and elevated amyloidogenesis, even though this activity might be removed through antioxidant treatment [238]. Collectively, these findings suggest that the neurotoxic activity of aluminum takes place in an elevated level of oxidative damage. In addition, these findings were observed in cultured neurons, where prolonged aluminum exposure caused aggregation of Aβ and fibrillar deposits on the surface of cells [239, 240]. In the body, increased aluminum levels influenced the activities of 3 crucial genes including presenilin-1, presenilin-2, and APP [241]. Aluminum also decreased the effects of some important enzymes associated with Aβ catabolism via triggering the activation of the amyloidogenic pathway [242, 243], which is suggesting a probable decrease of Aβ degradation. Moreover, aluminum increased hyperphosphorylated tau aggregation via suppressing the activity of protein phosphatase 2A [244]. Even though aluminum loading stimulates neurotoxic activities and causes behavioral alterations that partially model AD, but more studies are required to confirm whether toxic exposure of aluminum plays a role as a causal factor for AD.
In neurons, the molecular processes that are linked with aluminum transport are still unclear. Studies have revealed that uptake and transport of aluminum into cells is rather complicated because of other metal ions including iron, which is suggesting that aluminum competes with iron to bind with iron transporters (Tf/TfR or Lf/LfR), which are also involved with aluminum transport across the BBB [245]. A homolog of human DMT1, SMF-3, was involved with aluminum transport into neurons in Caenorhabditis elegans, which ultimately resulted in increased aluminum levels that decreased the levels of cellular ATP and mitochondrial membrane potential [246]. Besides, aluminum suppressed iron-mediated oxidation and the iron regulatory protein 2 (IRP2) degradation via the ubiquitin-proteasome pathway, which is indicating that aluminum stabilizes IRP2 to affect the balance of intracellular iron level [247]. Indeed, aluminum-mediated neurodegeneration seems to be linked with a different molecular pathway that is independent of tau- or Aβ-related toxicity and is mainly mediated by ROS generation and iron buildup in the brain [232].
Conclusion
The imbalances of intracellular biometal homeostasis and toxic metal exposure are linked to AD pathology. Various biometals have been reported to deposit in the brains of AD individuals, which further increased APP expression, Aβ aggregation, and hyperphosphorylation of tau. Toxic metal exposure might also trigger characteristics of AD pathology through various mechanisms including protein modification, neuroinflammation, and OS. Therefore, more studies are required to identify wider alterations of combined metal ion homeostasis in AD. Moreover, it is possible to identify the probable solutions for AD by recognizing the association between specific genes and particularly heavy metals. Indeed, major responsible genes for AD can also be targeted to treat this disease.
Abbreviations
- AChE:
-
Acetylcholinesterase
- AD:
-
Alzheimer’s disease
- APP:
-
Amyloid precursor protein
- ATOX1:
-
Antioxidant protein-1
- ATP:
-
Adenosine triphosphate
- ATP13A2:
-
ATPase cation transporting 13A2
- ATP7A and B:
-
ATPase copper transporting alpha and beta
- Aβ:
-
Amyloid beta
- BACE1:
-
Beta-secretase 1
- BBB:
-
Blood-brain barrier
- CDK5:
-
Cyclin-dependent kinase 5
- CSF:
-
Cerebrospinal fluid
- CTR1:
-
Copper transporter 1
- CTR1C:
-
Copper transporter 1C
- DMT1:
-
Divalent metal transporter 1
- FAD:
-
Familial AD
- FPN:
-
Ferroportin
- GSK3β:
-
Glycogen synthase kinase 3β
- IRE:
-
Iron-responsive element
- IRP1:
-
Iron regulatory protein 1
- IRP2:
-
Iron regulatory protein 2
- Lf:
-
Lactoferrin
- LRP:
-
Lipoprotein receptor–related protein
- MTf:
-
Melanotransferrin
- MTs:
-
Metallothioneins
- NFTs:
-
Neurofibrillary tangles
- OS:
-
Oxidative stress
- oxo8dG:
-
8-Hydroxyguanosin
- PS1:
-
Presenilin 1
- ROS:
-
Reactive oxygen species
- SLC30A10:
-
Solute carrier family 30 member 10
- SOD:
-
Superoxide dismutase
- SPCA1:
-
Secretory pathway Ca2+-ATPase 1
- SPs:
-
Senile plaques
- Tf:
-
Transferrin
- ZIPs:
-
Zinc-importing proteins
- ZnT:
-
Zinc transporter
References
Al Mamun A, Sahab Uddin M, Fahim Bin Bashar M et al (2020) Molecular insight into the therapeutic promise of targeting APOE4 for Alzheimer’s disease. Oxidative Med Cell Longev 2020:5086250. https://doi.org/10.1155/2020/5086250
Uddin MS, Devesh T, Mamun AAL et al (2020) Circadian and sleep dysfunction in Alzheimer’s disease. Ageing Res Rev 60:101046. https://doi.org/10.1016/J.ARR.2020.101046
Kabir MT, Uddin MS, Mathew B et al (2020) Emerging promise of immunotherapy for Alzheimer’s disease: a new hope for the development of Alzheimer’s vaccine. Curr Top Med Chem 20:1214–1234. https://doi.org/10.2174/1568026620666200422105156
Uddin MS, Kabir MT (2019) Oxidative stress in Alzheimer’s disease: molecular hallmarks of underlying vulnerability. In: Biological, diagnostic and therapeutic advances in Alzheimer’s disease. Springer Singapore, Singapore, pp. 91–115
Kolarova M, García-Sierra F, Bartos A et al (2012) Structure and pathology of tau protein in Alzheimer disease. Int J Alzheimers Dis 2012:731526. https://doi.org/10.1155/2012/731526
Jomova K, Vondrakova D, Lawson M, Valko M (2010) Metals, oxidative stress and neurodegenerative disorders. Mol Cell Biochem 345:91–104
Uddin MS, Kabir MT, Mamun A Al, et al (2020) Pharmacological approaches to mitigate neuroinflammation in Alzheimer’s disease. Int Immunopharmacol 84:106479. https://doi.org/10.1016/j.intimp.2020.106479
Uddin MS, Upaganlawar AB (2019) Oxidative stress and antioxidant defense biomedical value in health and diseases. Nova Science Publishers, New York
Kabir MT, Uddin MS, Begum MM et al (2019) Cholinesterase inhibitors for Alzheimer’s disease: multitargeting strategy based on anti-Alzheimer’s drugs repositioning. Curr Pharm Des 25:3519–3535. https://doi.org/10.2174/1381612825666191008103141
Uddin MS, Kabir MT, Jeandet P et al (2020) Novel anti-Alzheimer’s therapeutic molecules targeting amyloid precursor protein processing. Oxidative Med Cell Longev 2020:7039138. https://doi.org/10.1155/2020/7039138
Kabir MT, Sufian MA, Uddin MS et al (2019) NMDA receptor antagonists: repositioning of memantine as a multitargeting agent for Alzheimer’s therapy. Curr Pharm Des 25:3506–3518. https://doi.org/10.2174/1381612825666191011102444
Ayton S, Lei P, Bush AI (2015) Biometals and their therapeutic implications in Alzheimer’s disease. Neurotherapeutics 12:109–120
Kabir MT, Uddin MS, Mamun A Al, et al (2020) Combination drug therapy for the management of Alzheimer’s disease. Int J Mol Sci 21:3272. https://doi.org/10.3390/ijms21093272
Kepp KP (2017) Ten challenges of the amyloid hypothesis of Alzheimer’s disease. J Alzheimers Dis 55:447–457. https://doi.org/10.3233/JAD-160550
Bush AI, Tanzi RE (2008) Therapeutics for Alzheimer’s disease based on the metal hypothesis. Neurotherapeutics 5:421–432. https://doi.org/10.1016/j.nurt.2008.05.001
Zhang Z, Miah M, Culbreth M, Aschner M (2016) Autophagy in neurodegenerative diseases and metal neurotoxicity. Neurochem Res 41:409–422. https://doi.org/10.1007/s11064-016-1844-x
Chin-Chan M, Navarro-Yepes J, Quintanilla-Vega B (2015) Environmental pollutants as risk factors for neurodegenerative disorders: Alzheimer and Parkinson diseases. Front Cell Neurosci 9:124. https://doi.org/10.3389/fncel.2015.00124
Akhtar A, Dhaliwal J, Saroj P et al (2020) Chromium picolinate attenuates cognitive deficit in ICV-STZ rat paradigm of sporadic Alzheimer’s-like dementia via targeting neuroinflammatory and IRS-1/PI3K/AKT/GSK-3β pathway. Inflammopharmacology 28:385–400. https://doi.org/10.1007/s10787-019-00681-7
Harilal S, Jose J et al (2020) Revisiting the blood-brain barrier: a hard nut to crack in the transportation of drug molecules. Brain Res Bull. https://doi.org/10.1016/j.brainresbull.2020.03.018
Pardridge WM (2012) Drug transport across the blood-brain barrier. J Cereb Blood Flow Metab 32:1959–1972
Lee HJ, Park MK, Seo YR (2018) Pathogenic mechanisms of heavy metal induced-Alzheimer’s disease. Toxicol Environ Health Sci 10:1–10. https://doi.org/10.1007/s13530-018-0340-x
Ward RJ, Zucca FA, Duyn JH et al (2014) The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol 13:1045–1060. https://doi.org/10.1016/S1474-4422(14)70117-6
Castellani RJ, Moreira PI, Perry G, Zhu X (2012) The role of iron as a mediator of oxidative stress in Alzheimer disease. BioFactors 38:133–138
Biasiotto G, Di Lorenzo D, Archetti S, Zanella I (2016) Iron and neurodegeneration: is ferritinophagy the link? Mol Neurobiol 53:5542–5574
Bartzokis G, Sultzer D, Mintz J et al (1994) In vivo evaluation of brain iron in Alzheimer’s disease and normal subjects using MRI. Biol Psychiatry 35:480–487. https://doi.org/10.1016/0006-3223(94)90047-7
Levine SM (1997) Iron deposits in multiple sclerosis and Alzheimer’s disease brains. Brain Res 760:298–303. https://doi.org/10.1016/S0006-8993(97)00470-8
Wang D, Li YY, Luo JH, Li YH (2014) Age-related iron deposition in the basal ganglia of controls and Alzheimer disease patients quantified using susceptibility weighted imaging. Arch Gerontol Geriatr 59:439–449. https://doi.org/10.1016/j.archger.2014.04.002
Moon Y, Han SH, Moon WJ (2016) Patterns of brain iron accumulation in vascular dementia and Alzheimer’s dementia using quantitative susceptibility mapping imaging. J Alzheimers Dis 51:737–745. https://doi.org/10.3233/JAD-151037
Tao Y, Wang Y, Rogers JT, Wang F (2014) Perturbed iron distribution in Alzheimer’s disease serum, cerebrospinal fluid, and selected brain regions: a systematic review and meta-analysis. J Alzheimers Dis 42:679–690. https://doi.org/10.3233/JAD-140396
Wang ZX, Tan L, Wang HF et al (2015) Serum iron, zinc, and copper levels in patients with Alzheimer’s disease: a replication study and meta-analyses. J Alzheimers Dis 47:565–581. https://doi.org/10.3233/JAD-143108
Song N, Jiang H, Wang J, Xie JX (2007) Divalent metal transporter 1 up-regulation is involved in the 6-hydroxydopamine-induced ferrous iron influx. J Neurosci Res 85:3118–3126. https://doi.org/10.1002/jnr.21430
Xian-Hui D, Wei-Juan G, Tie-Mei S et al (2015) Age-related changes of brain iron load changes in the frontal cortex in APPswe/PS1ΔE9 transgenic mouse model of Alzheimer’s disease. J Trace Elem Med Biol 30:118–123. https://doi.org/10.1016/j.jtemb.2014.11.009
Raha AA, Vaishnav RA, Friedland RP et al (2014) The systemic iron-regulatory proteins hepcidin and ferroportin are reduced in the brain in Alzheimer’s disease. Acta Neuropathol Commun 2:55. https://doi.org/10.1186/2051-5960-1-55
McCarthy RC, Park Y, Kosman DJ (2014) sAPP modulates iron efflux from brain microvascular endothelial cells by stabilizing the ferrous iron exporter ferroportin. EMBO Rep 15:809–815. https://doi.org/10.15252/embr.201338064
Duce JA, Tsatsanis A, Cater MA et al (2010) Iron-export ferroxidase activity of β-amyloid precursor protein is inhibited by zinc in Alzheimer’s disease. Cell 142:857–867. https://doi.org/10.1016/j.cell.2010.08.014
Lei P, Ayton S, Finkelstein DI et al (2012) Tau deficiency induces parkinsonism with dementia by impairing APP-mediated iron export. Nat Med 18:291–295. https://doi.org/10.1038/nm.2613
Lei P, Ayton S, Appukuttan AT et al (2017) Lithium suppression of tau induces brain iron accumulation and neurodegeneration. Mol Psychiatry 22:396–406. https://doi.org/10.1038/mp.2016.96
Lei P, Ayton S, Appukuttan AT et al (2015) Clioquinol rescues parkinsonism and dementia phenotypes of the tau knockout mouse. Neurobiol Dis 81:168–175. https://doi.org/10.1016/j.nbd.2015.03.015
Yang X, Park SH, Chang HC et al (2017) Sirtuin 2 regulates cellular iron homeostasis via deacetylation of transcription factor NRF2. J Clin Invest 127:1505–1516. https://doi.org/10.1172/JCI88574
Moos T, Morgan EH (2000) Transferrin and transferrin receptor function in brain barrier systems. Cell Mol Neurobiol 20:77–95. https://doi.org/10.1023/a:1006948027674
Ringman JM, Schulman H, Becker C et al (2012) Proteomic changes in cerebrospinal fluid of presymptomatic and affected persons carrying familial Alzheimer disease mutations. Arch Neurol 69:96–104. https://doi.org/10.1001/archneurol.2011.642
Baker EN, Anderson BF, Baker HM et al (1994) Three-dimensional structure of lactoferrin in various functional states. In: Advances in experimental medicine and biology, pp. 1–12
An L, Sato H, Konishi Y et al (2009) Expression and localization of lactotransferrin messenger RNA in the cortex of Alzheimer’s disease. Neurosci Lett 452:277–280. https://doi.org/10.1016/j.neulet.2009.01.071
Wang L, Sato H, Zhao S, Tooyama I (2010) Deposition of lactoferrin in fibrillar-type senile plaques in the brains of transgenic mouse models of Alzheimer’s disease. Neurosci Lett 481:164–167. https://doi.org/10.1016/j.neulet.2010.06.079
Qiu Z, Strickland DK, Hyman BT, Rebeck GW (1999) α2-Macroglobulin enhances the clearance of endogenous soluble β-amyloid peptide via low-density lipoprotein receptor-related protein in cortical neurons. J Neurochem 73:1393–1398. https://doi.org/10.1046/j.1471-4159.1999.0731393.x
Kuo YC, Wang CT (2014) Protection of SK-N-MC cells against β-amyloid peptide-induced degeneration using neuron growth factor-loaded liposomes with surface lactoferrin. Biomaterials 35:5954–5964. https://doi.org/10.1016/j.biomaterials.2014.03.082
Meng F, Asghar S, Gao S et al (2015) A novel LDL-mimic nanocarrier for the targeted delivery of curcumin into the brain to treat Alzheimer’s disease. Colloids Surfaces B Biointerfaces 134:88–97. https://doi.org/10.1016/j.colsurfb.2015.06.025
Scheiber IF, Mercer JFB, Dringen R (2014) Metabolism and functions of copper in brain. Prog Neurobiol 116:33–57. https://doi.org/10.1016/j.pneurobio.2014.01.002
Lovell MA, Robertson JD, Teesdale WJ et al (1998) Copper, iron and zinc in Alzheimer’s disease senile plaques. J Neurol Sci 158:47–52. https://doi.org/10.1016/S0022-510X(98)00092-6
Deibel MA, Ehmann WD, Markesbery WR (1996) Copper, iron, and zinc imbalances in severely degenerated brain regions in Alzheimer’s disease: possible relation to oxidative stress. J Neurol Sci 143:137–142. https://doi.org/10.1016/S0022-510X(96)00203-1
Klevay LM (2008) Alzheimer’s disease as copper deficiency. Med Hypotheses 70:802–807. https://doi.org/10.1016/j.mehy.2007.04.051
Vaz FNC, Fermino BL, Haskel MVL et al (2018) The relationship between copper, iron, and selenium levels and Alzheimer disease. Biol Trace Elem Res 181:185–191. https://doi.org/10.1007/s12011-017-1042-y
Bucossi S, Ventriglia M, Panetta V et al (2011) Copper in Alzheimer’s disease: a meta-analysis of serum, plasma, and cerebrospinal fluid studies. J Alzheimers Dis 24:175–185. https://doi.org/10.3233/JAD-2010-101473
Sparks DL, Schreurs BG (2003) Trace amounts of copper in water induce β-amyloid plaques and learning deficits in a rabbit model of Alzheimer’s disease. Proc Natl Acad Sci U S A 100:11065–11069. https://doi.org/10.1073/pnas.1832769100
Kitazawa M, Cheng D, Laferla FM (2009) Chronic copper exposure exacerbates both amyloid and tau pathology and selectively dysregulates cdk5 in a mouse model of AD. J Neurochem 108:1550–1560. https://doi.org/10.1111/j.1471-4159.2009.05901.x
Kuo Y-M, Gybina AA, Pyatskowit JW et al (2006) Copper transport protein (Ctr1) levels in mice are tissue specific and dependent on copper status. J Nutr 136:21–26. https://doi.org/10.1093/jn/136.1.21
Yu CH, Dolgova NV, Dmitriev OY (2017) Dynamics of the metal binding domains and regulation of the human copper transporters ATP7B and ATP7A. IUBMB Life 69:226–235
Zheng W, Monnot AD (2012) Regulation of brain iron and copper homeostasis by brain barrier systems: implication in neurodegenerative diseases. Pharmacol Ther 133:177–188
Harris ED (2009) Copper homeostasis: the role of cellular transporters. Nutr Rev 59:281–285. https://doi.org/10.1111/j.1753-4887.2001.tb07017.x
Lang M, Fan Q, Wang L et al (2013) Inhibition of human high-affinity copper importer Ctr1 orthologous in the nervous system of Drosophila ameliorates Aβ42-induced Alzheimer’s disease-like symptoms. Neurobiol Aging 34:2604–2612. https://doi.org/10.1016/j.neurobiolaging.2013.05.029
Zheng Z, White C, Lee J et al (2010) Altered microglial copper homeostasis in a mouse model of Alzheimer’s disease. J Neurochem 114:1630–1638. https://doi.org/10.1111/j.1471-4159.2010.06888.x
Bucossi S, Polimanti R, Ventriglia M et al (2013) Intronic rs2147363 variant in ATP7B transcription factor-binding site associated with Alzheimer’s disease. J Alzheimers Dis 37:453–459. https://doi.org/10.3233/JAD-130431
Maret W (2013) Zinc biochemistry: from a single zinc enzyme to a key element of life. Adv Nutr 4:82–91. https://doi.org/10.3945/an.112.003038
Barr CA, Burdette SC (2017) The zinc paradigm for metalloneurochemistry. Essays Biochem 61:225–235
Bush AI, Pettingell WH, Multhaup G, et al (1994) Rapid induction of Alzheimer Aβ amyloid formation by zinc. Science (80- ) 265:1464–1467. https://doi.org/10.1126/science.8073293
Religa D, Strozyk D, Cherny RA et al (2006) Elevated cortical zinc in Alzheimer disease. Neurology 67:69–75. https://doi.org/10.1212/01.wnl.0000223644.08653.b5
Panayi AE, Spyrou NM, Iversen BS et al (2002) Determination of cadmium and zinc in Alzheimer’s brain tissue using inductively coupled plasma mass spectrometry. J Neurol Sci 195:1–10. https://doi.org/10.1016/s0022-510x(01)00672-4
Ventriglia M, Brewer GJ, Simonelli I et al (2015) Zinc in Alzheimer’s disease: a meta-analysis of serum, plasma, and cerebrospinal fluid studies. J Alzheimers Dis 46:75–87. https://doi.org/10.3233/JAD-141296
Yoshiike Y, Tanemura K, Murayama O et al (2001) New insights on how metals disrupt amyloid β-aggregation and their effects on amyloid-β cytotoxicity. J Biol Chem 276:32293–32299. https://doi.org/10.1074/jbc.M010706200
Huang L, Tepaamorndech S (2013) The SLC30 family of zinc transporters-a review of current understanding of their biological and pathophysiological roles. Mol Asp Med 34:548–560
Cousins RJ, Liuzzi JP, Lichten LA (2006) Mammalian zinc transport, trafficking, and signals. J Biol Chem 281:24085–24089
Krezel A, Hao Q, Maret W (2007) The zinc/thiolate redox biochemistry of metallothionein and the control of zinc ion fluctuations in cell signaling. Arch Biochem Biophys 463:188–200
Zhang LH, Wang X, Stoltenberg M et al (2008) Abundant expression of zinc transporters in the amyloid plaques of Alzheimer’s disease brain. Brain Res Bull 77:55–60. https://doi.org/10.1016/j.brainresbull.2008.03.014
Palmiter RD, Cole TB, Quaife CJ, Findley SD (1996) ZnT-3, a putative transporter of zinc into synaptic vesicles. Proc Natl Acad Sci U S A 93:14934–14939. https://doi.org/10.1073/pnas.93.25.14934
Whitfield DR, Vallortigara J, Alghamdi A et al (2014) Assessment of ZnT3 and PSD95 protein levels in Lewy body dementias and Alzheimer’s disease: association with cognitive impairment. Neurobiol Aging 35:2836–2844. https://doi.org/10.1016/j.neurobiolaging.2014.06.015
Adlard PA, Parncutt JM, Finkelstein DI, Bush AI (2010) Cognitive loss in zinc transporter-3 knock-out mice: a phenocopy for the synaptic and memory deficits of Alzheimer’s disease? J Neurosci 30:1631–1636. https://doi.org/10.1523/JNEUROSCI.5255-09.2010
Johnson-Wood K, Lee M, Motter R et al (2002) Amyloid precursor protein processing and a 42 deposition in a transgenic mouse model of Alzheimer disease. Proc Natl Acad Sci 94:1550–1555. https://doi.org/10.1073/pnas.94.4.1550
Greenough MA, Volitakis I, Li QX et al (2011) Presenilins promote the cellular uptake of copper and zinc and maintain copper chaperone of SOD1-dependent copper/zinc superoxide dismutase activity. J Biol Chem 286:9776–9786. https://doi.org/10.1074/jbc.M110.163964
Hidalgo J, Penkowa M, Espejo C et al (2006) Expression of metallothionein-I, -II, and -III in Alzheimer disease and animal models of neuroinflammation. Exp Biol Med 231:1450–1458. https://doi.org/10.1177/153537020623100902
Manso Y, Carrasco J, Comes G et al (2012) Characterization of the role of the antioxidant proteins metallothioneins 1 and 2 in an animal model of Alzheimer’s disease. Cell Mol Life Sci 69:3665–3681. https://doi.org/10.1007/s00018-012-1045-y
Manso Y, Carrasco J, Comes G et al (2012) Characterization of the role of etallothionein-3 in an animal model of Alzheimer’s disease. Cell Mol Life Sci 69:3683–3700. https://doi.org/10.1007/s00018-012-1047-9
Lee SJ, Seo BR, Koh JY (2015) Metallothionein-3 modulates the amyloid β endocytosis of astrocytes through its effects on actin polymerization. Mol Brain 8:84. https://doi.org/10.1186/s13041-015-0173-3
Pedersen JT, Hureau C, Hemmingsen L et al (2012) Rapid exchange of metal between Zn 7-metallothionein-3 and amyloid-β peptide promotes amyloid-related structural changes. Biochemistry 51:1697–1706. https://doi.org/10.1021/bi201774z
Kim JH, Nam YP, Jeon SM et al (2012) Amyloid neurotoxicity is attenuated by metallothionein: dual mechanisms at work. J Neurochem 121:751–762. https://doi.org/10.1111/j.1471-4159.2012.07725.x
Mo ZY, Zhu YZ, Zhu HL et al (2009) Low micromolar zinc accelerates the fibrillization of human tau via bridging of Cys-291 and Cys-322. J Biol Chem 284:34648–34657. https://doi.org/10.1074/jbc.M109.058883
An WL, Bjorkdahl C, Liu R et al (2005) Mechanism of zinc-induced phosphorylation of p70 S6 kinase and glycogen synthase kinase 3β in SH-SY5Y neuroblastoma cells. J Neurochem 92:1104–1115. https://doi.org/10.1111/j.1471-4159.2004.02948.x
Prakash A, Dhaliwal GK, Kumar P, Majeed ABA (2017) Brain biometals and Alzheimer’s disease–boon or bane? Int J Neurosci 127:99–108. https://doi.org/10.3109/00207454.2016.1174118
Park RM (2013) Neurobehavioral deficits and parkinsonism in occupations with manganese exposure: a review of methodological issues in the epidemiological literature. Saf Health Work 4:123–135. https://doi.org/10.1016/j.shaw.2013.07.003
Guilarte TR (2013) Manganese neurotoxicity: new perspectives from behavioral, neuroimaging, and neuropathological studies in humans and non-human primates. Front Aging Neurosci 5. https://doi.org/10.3389/fnagi.2013.00023
Martinez-Finley EJ, Gavin CE, Aschner M, Gunter TE (2013) Manganese neurotoxicity and the role of reactive oxygen species. Free Radic Biol Med 62:65–75
Srivastava RAK, Jain JC (2002) Scavenger receptor class B type I expression and elemental analysis in cerebellum and parietal cortex regions of the Alzheimer’s disease brain. J Neurol Sci 196:45–52. https://doi.org/10.1016/S0022-510X(02)00026-6
Tong Y, Yang H, Tian X et al (2014) High manganese, a risk for Alzheimer’s disease: high manganese induces amyloid-β related cognitive impairment. J Alzheimers Dis 42:865–878. https://doi.org/10.3233/JAD-140534
Guilarte TR (2010) APLP1, Alzheimer’s-like pathology and neurodegeneration in the frontal cortex of manganese-exposed non-human primates. Neurotoxicology 31:572–574. https://doi.org/10.1016/j.neuro.2010.02.004
Schneider JS, Williams C, Ault M, Guilarte TR (2013) Chronic manganese exposure impairs visuospatial associative learning in non-human primates. Toxicol Lett 221:146–151. https://doi.org/10.1016/j.toxlet.2013.06.211
Wallin C, Kulkarni YS, Abelein A et al (2016) Characterization of Mn(II) ion binding to the amyloid-β peptide in Alzheimer’s disease. J Trace Elem Med Biol 38:183–193. https://doi.org/10.1016/j.jtemb.2016.03.009
Gunter TE, Gavin CE, Aschner M, Gunter KK (2006) Speciation of manganese in cells and mitochondria: a search for the proximal cause of manganese neurotoxicity. Neurotoxicology 27:765–776. https://doi.org/10.1016/j.neuro.2006.05.002
Li F, Calingasan NY, Yu F et al (2004) Increased plaque burden in brains of APP mutant MnSOD heterozygous knockout mice. J Neurochem 89:1308–1312. https://doi.org/10.1111/j.1471-4159.2004.02455.x
Melov S, Adlard PA, Morten K et al (2007) Mitochondrial oxidative stress causes hyperphosphorylation of tau. PLoS One 2. https://doi.org/10.1371/journal.pone.0000536
Dumont M, Wille E, Stack C et al (2009) Reduction of oxidative stress, amyloid deposition, and memory deficit by manganese superoxide dismutase overexpression in a transgenic mouse model of Alzheimer’s disease. FASEB J 23:2459–2466. https://doi.org/10.1096/fj.09-132928
Carmona A, Devès G, Roudeau S et al (2010) Manganese accumulates within golgi apparatus in dopaminergic cells as revealed by synchrotron X-ray fluorescence nanoimaging. ACS Chem Neurosci 1:194–203. https://doi.org/10.1021/cn900021z
Garrick MD, Singleton ST, Vargas F et al (2006) DMT1: which metals does it transport? Biol Res 39:79–85. https://doi.org/10.4067/S0716-97602006000100009
Subramaniam VN, Summerville L, Wallace DF (2002) Molecular and cellular characterization of transferrin receptor 2. Cell Biochem Biophys 36:235–239. https://doi.org/10.1385/CBB:36:2-3:235
Fujishiro H, Yano Y, Takada Y et al (2012) Roles of ZIP8, ZIP14, and DMT1 in transport of cadmium and manganese in mouse kidney proximal tubule cells. Metallomics 4:700–708. https://doi.org/10.1039/c2mt20024d
Lin W, Vann DR, Doulias PT et al (2017) Hepatic metal ion transporter ZIP8 regulates manganese homeostasis and manganese-dependent enzyme activity. J Clin Invest 127:2407–2417. https://doi.org/10.1172/JCI90896
Aydemir TB, Kim MH, Kim J et al (2017) Metal transporter Zip14 (Slc39a14) deletion in mice increases manganese deposition and produces neurotoxic signatures and diminished motor activity. J Neurosci 37:5996–6006. https://doi.org/10.1523/JNEUROSCI.0285-17.2017
Tuschl K, Clayton PT, Gospe SM et al (2012) Syndrome of hepatic cirrhosis, dystonia, polycythemia, and hypermanganesemia caused by mutations in SLC30A10, a manganese transporter in man. Am J Hum Genet 90:457–466. https://doi.org/10.1016/j.ajhg.2012.01.018
Bosomworth HJ, Adlard PA, Ford D, Valentine RA (2013) Altered expression of ZnT10 in Alzheimer’s disease brain. PLoS One 8:. https://doi.org/10.1371/journal.pone.0065475
Madejczyk MS, Ballatori N (2012) The iron transporter ferroportin can also function as a manganese exporter. Biochim Biophys Acta Biomembr 1818:651–657. https://doi.org/10.1016/j.bbamem.2011.12.002
Seo YA, Wessling-Resnick M (2015) Ferroportin deficiency impairs manganese metabolism in flatiron mice. FASEB J 29:2726–2733. https://doi.org/10.1096/fj.14-262592
Murphy KE, Cottle L, Gysbers AM et al (2014) ATP13A2 (PARK9) protein levels are reduced in brain tissue of cases with Lewy bodies. Acta Neuropathol Commun 2. https://doi.org/10.1186/2051-5960-1-11
Ton VK, Mandal D, Vahadji C, Rao R (2002) Functional expression in yeast of the human secretory pathway Ca(2+), Mn(2+)-ATPase defective in Hailey-Hailey disease. J Biol Chem 277:6422–6427. https://doi.org/10.1074/jbc.M110612200
Misra VK, Draper DE (1998) On the role of magnesium ions in RNA stability. Biopolymers 48:113–35.
Bairoch A (2000) The ENZYME database in 2000. Nucleic Acids Res 28:304–305. https://doi.org/10.1093/nar/28.1.304
Komuro H, Kumada T (2005) Ca2+ transients control CNS neuronal migration. Cell Calcium 37:387–393. https://doi.org/10.1016/j.ceca.2005.01.006
Levitsky DO, Takahashi M (2013) Interplay of Ca2+ and Mg2+ in sodium-calcium exchanger and in other Ca2+-binding proteins: magnesium, watchdog that blocks each turn if able. In: Advances in experimental medicine and biology. Adv Exp Med Biol, pp. 65–78
Volpe SL (2013) Magnesium in disease prevention and overall health. Adv Nutr 4:378S–383S. https://doi.org/10.3945/an.112.003483
Andrási E, Páli N, Molnár Z, Kösel S (2005) Brain aluminum, magnesium and phosphorus contents of control and Alzheimer-diseased patients. J Alzheimers Dis 7:273–284. https://doi.org/10.3233/JAD-2005-7402
Çilliler AE, Öztürk Ş, Özbakir Ş (2008) Serum magnesium level and clinical deterioration in Alzheimer’s disease. Gerontology 53:419–422. https://doi.org/10.1159/000110873
Stutzmann GE (2007) The pathogenesis of Alzheimers disease - is it a lifelong “calciumopathy”? Neuroscientist 13:546–559. https://doi.org/10.1177/1073858407299730
Brzyska M, Elbaum D (2003) Dysregulation of calcium in Alzheimer’s disease. Acta Neurobiol Exp (Wars) 63:171–183
Ho M, Hoke DE, Chua YJ, Li QX, Culvenor JG et al (2011) Effect of metal chelators on γ-secretase indicates that calcium and magnesium ions facilitate cleavage of Alzheimer amyloid precursor substrate. Int J Alzheimers Dis 2011:950932. https://doi.org/10.4061/2011/950932
Kelly BL, Ferreira A (2006) β-Amyloid-induced dynamin 1 degradation is mediated by N-methyl-D-aspartate receptors in hippocampal neurons. J Biol Chem 281:28079–28089. https://doi.org/10.1074/jbc.M605081200
LS Y, H K-R (1999) Ca2+ and Mg2+ selectively induce aggregates of PHF-tau but not normal human tau. J Neurosci Res 55. https://doi.org/10.1002/(SICI)1097-4547(19990101)55:1<36::AID-JNR5>3.0.CO;2-E
Li W, Yu J, Liu Y et al (2014) Elevation of brain magnesium prevents synaptic loss and reverses cognitive deficits in Alzheimer’s disease mouse model. Mol Brain 7. https://doi.org/10.1186/s13041-014-0065-y
Gomez-Ramos A, Dominguez J, Zafra D et al (2006) Inhibition of GSK3 dependent tau phosphorylation by metals. Curr Alzheimer Res 3:123–127. https://doi.org/10.2174/156720506776383059
Xu ZP, Li L, Bao J et al (2014) Magnesium protects cognitive functions and synaptic plasticity in streptozotocin-induced sporadic Alzheimer’s model. PLoS One 9. https://doi.org/10.1371/journal.pone.0108645
Parameshwaran K, Dhanasekaran M, Suppiramaniam V (2008) Amyloid beta peptides and glutamatergic synaptic dysregulation. Exp Neurol 210:7–13
Mota SI, Ferreira IL, Rego AC (2014) Dysfunctional synapse in Alzheimer’s disease - a focus on NMDA receptors. Neuropharmacology 76:16–26
Kotermanski SE, Johnson JW (2009) Mg 2+ imparts NMDA receptor subtype selectivity to the Alzheimer’s drug memantine. J Neurosci 29:2774–2779. https://doi.org/10.1523/JNEUROSCI.3703-08.2009
Witting A, Walter L, Wacker J et al (2004) P2X7 receptors control 2-arachidonoylglycerol production by microglial cells. Proc Natl Acad Sci U S A 101:3214–3219. https://doi.org/10.1073/pnas.0306707101
North RA (2002) Molecular physiology of P2X receptors. Physiol Rev 82:1013–1067. https://doi.org/10.1152/physrev.00015.2002
Lee M, Jantaratnotai N, McGeer E et al (2011) Mg2+ ions reduce microglial and THP-1 cell neurotoxicity by inhibiting Ca2+ entry through purinergic channels. Brain Res 1369:21–35. https://doi.org/10.1016/j.brainres.2010.10.084
Romani AMP (2011) Cellular magnesium homeostasis. Arch Biochem Biophys 512:1–23. https://doi.org/10.1016/j.abb.2011.05.010
de Baaij JHF, Hoenderop JGJ, Bindels RJM (2015) Magnesium in man: implications for health and disease. Physiol Rev 95:1–46. https://doi.org/10.1152/physrev.00012.2014
Tu H, Nelson O, Bezprozvanny A et al (2006) Presenilins form ER Ca2+ leak channels, a function disrupted by familial Alzheimer’s disease-linked mutations. Cell 126:981–993. https://doi.org/10.1016/j.cell.2006.06.059
Cheung KH, Shineman D, Müller M et al (2008) Mechanism of Ca2+ disruption in Alzheimer’s disease by presenilin regulation of InsP3 receptor channel gating. Neuron 58:871–883. https://doi.org/10.1016/j.neuron.2008.04.015
Diaz JC, Simakova O, Jacobson KA et al (2009) Small molecule blockers of the Alzheimer Aβ calcium channel potently protect neurons from Aβ cytotoxicity. Proc Natl Acad Sci U S A 106:3348–3353. https://doi.org/10.1073/pnas.0813355106
Arbel-Ornath M, Hudry E, Boivin JR et al (2017) Soluble oligomeric amyloid-β induces calcium dyshomeostasis that precedes synapse loss in the living mouse brain. Mol Neurodegener 12. https://doi.org/10.1186/s13024-017-0169-9
Oh HG, Chun YS, Kim Y et al (2012) Modulation of transient receptor potential melastatin related 7 channel by presenilins. Dev Neurobiol 72:865–877. https://doi.org/10.1002/dneu.22001
Ostapchenko VG, Chen M, Guzman MS et al (2015) The transient receptor potential melastatin 2 (TRPM2) channel contributes to β-amyloid oligomer-related neurotoxicity and memory impairment. J Neurosci 35:15157–15169. https://doi.org/10.1523/JNEUROSCI.4081-14.2015
Hermosura MC, Garruto RM (2007) TRPM7 and TRPM2-candidate susceptibility genes for Western Pacific ALS and PD? Biochim Biophys Acta - Mol Basis Dis 1772:822–835. https://doi.org/10.1016/j.bbadis.2007.02.008
Lin CF, Chen CL, Chiang CW et al (2007) GSK-3β acts downstream of PP2A and the PI 3-kinase-Akt pathway, and upstream of caspase-2 in ceramide-induced mitochondrial apoptosis. J Cell Sci 120:2935–2943. https://doi.org/10.1242/jcs.03473
Lee HE, Choi ES, Jung JY et al (2014) Inhibition of specificity protein 1 by dibenzylideneacetone, a curcumin analogue, induces apoptosis in mucoepidermoid carcinomas and tumor xenografts through Bim and truncated Bid. Oral Oncol 50:189–195. https://doi.org/10.1016/j.oraloncology.2013.11.006
Feng J, Meng C, Xing D (2015) Aβ induces PUMA activation: a new mechanism for Aβ-mediated neuronal apoptosis. Neurobiol Aging 36:789–800. https://doi.org/10.1016/j.neurobiolaging.2014.10.007
Ekström L, Lyrenäs L, Jakobsson PJ et al (2003) Basal expression of the human MAPEG members microsomal glutathione transferase 1 and prostaglandin E synthase genes is mediated by Sp1 and Sp3. Biochim Biophys Acta - Gene Struct Expr 1627:79–84. https://doi.org/10.1016/S0167-4781(03)00077-0
Satoh K, Nagano Y, Shimomura C et al (2000) Expression of prostaglandin E synthase mRNA is induced in beta-amyloid treated rat astrocytes. Neurosci Lett 283:221–223. https://doi.org/10.1016/S0304-3940(00)00926-5
de Oliveira AC et al (2012) Pharmacological inhibition of Akt and downstream pathways modulates the expression of COX-2 and mPGES-1 in activated microglia. J Neuroinflammation 9. https://doi.org/10.1186/1742-2094-9-2
Li C, Ge Y, Dworkin L et al (2016) The β isoform of GSK3 mediates podocyte autonomous injury in proteinuric glomerulopathy. J Pathol 239:23–35. https://doi.org/10.1002/path.4692
Stephen TL, Niemeyer M, Tzianabos AO et al (2005) Effect of B7-2 and CD40 signals from activated antigen-presenting cells on the ability of zwitterionic polysaccharides to induce T-cell stimulation. Infect Immun 73:2184–2189. https://doi.org/10.1128/IAI.73.4.2184-2189.2005
McQuillan K, Lynch MA, Mills KHG (2010) Activation of mixed glia by Aβ-specific Th1 and Th17 cells and its regulation by Th2 cells. Brain Behav Immun 24:598–607. https://doi.org/10.1016/j.bbi.2010.01.003
Liu H, Mi S, Li Z et al (2013) SB216763, a selective small molecule inhibitor of glycogen synthase kinase-3, improves bleomycin-induced pulmonary fibrosis via activating autophagy. Acta Pharm Sin B 3:226–233. https://doi.org/10.1016/j.apsb.2013.05.004
Ling YH, Tornos C, Perez-Soler R (1998) Phosphorylation of Bcl-2 is a marker of M phase events and not a determinant of apoptosis. J Biol Chem 273:18984–18991. https://doi.org/10.1074/jbc.273.30.18984
Ju TC, Der Chen S, Liu CC, Yang DI (2005) Protective effects of S-nitrosoglutathione against amyloid β-peptide neurotoxicity. Free Radic Biol Med 38:938–949. https://doi.org/10.1016/j.freeradbiomed.2004.12.019
Gupta C, Kaur J, Tikoo K (2014) Regulation of MDAMB-231 cell proliferation by GSK-3β involves epigenetic modifications under high glucose conditions. Exp Cell Res 324. https://doi.org/10.1016/j.yexcr.2014.03.019
Ghosh F, Arnér K, Engelsberg K (2009) Isolation of photoreceptors in the cultured full-thickness fetal rat retina. Invest Ophthalmol Vis Sci:50. https://doi.org/10.1167/iovs.08-2389
Levin EC (2009) Neuronal expression of vimentin in the Alzheimer’s disease brain may be part of a generalized dendritic damage-response mechanism. Brain Res:1298. https://doi.org/10.1016/j.brainres.2009.08.072
Wu K, Liu M, Li A et al (2007) Cell fate determination factor DACH1 inhibits c-Jun-induced contact-independent growth. Mol Biol Cell 18:755–767. https://doi.org/10.1091/mbc.E06-09-0793
Saini MK, Sanyal SN (2012) PTEN regulates apoptotic cell death through PI3-K/Akt/GSK3β signaling pathway in DMH induced early colon carcinogenesis in rat. Exp Mol Pathol 93:135–146. https://doi.org/10.1016/j.yexmp.2012.04.019
Kam TI, Song S, Gwon Y et al (2013) FcγRIIb mediates amyloid-β neurotoxicity and memory impairment in Alzheimer’s disease. J Clin Invest 123:2791–2802. https://doi.org/10.1172/JCI66827
Corsini NS (2009) The death receptor CD95 activates adult neural stem cells for working memory formation and brain repair. Cell Stem Cell 5. https://doi.org/10.1016/j.stem.2009.05.004
Chen Y, Dong C (2009) Abeta40 promotes neuronal cell fate in neural progenitor cells. Cell Death Differ 16. https://doi.org/10.1038/cdd.2008.94
Edelman MJ, Shvartsbeyn M (2012) Epothilones in development for non-small-cell lung cancer: novel anti-tubulin agents with the potential to overcome taxane resistance. Clin Lung Cancer 13. https://doi.org/10.1016/j.cllc.2011.02.005
Stengel C (2014) In vivo and in vitro properties of STX2484: a novel non-steroidal anti-cancer compound active in taxane-resistant breast cancer. Br J Cancer 111. https://doi.org/10.1038/bjc.2014.188
Jans DA, Thomas RJ, Gillespie MT (2003) Parathyroid hormone-related protein (PTHrP): a nucleocytoplasmic shuttling protein with distinct paracrine and intracrine roles. Vitam Horm 66:. https://doi.org/10.1016/S0083-6729(03)01010-0
Kaminsky YG, Marlatt MW, Smith MA, Kosenko EA (2010) Subcellular and metabolic examination of amyloid-β peptides in Alzheimer disease pathogenesis: evidence for Aβ25-35. Exp Neurol 221. https://doi.org/10.1016/j.expneurol.2009.09.005
Fiala M (2007) Innate immunity and transcription of MGAT-III and Toll-like receptors in Alzheimer’s disease patients are improved by bisdemethoxycurcumin. Proc Natl Acad Sci USA:104. https://doi.org/10.1073/pnas.0701267104
Wasiluk KR, McCulloch KA, Banton KL, Dunn DL (2006) Sp1 elements regulate transcriptional activity within the murine Toll-like receptor 4 promoter. Surg Infect 7. https://doi.org/10.1089/sur.2006.7.489
Bai XT, Baydoun HH, Nicot C (2010) HTLV-I p30: a versatile protein modulating virus replication and pathogenesis. Mol Asp Med 31. https://doi.org/10.1016/j.mam.2010.05.004
Linde CI, Baryshnikov SG, Mazzocco-Spezzia A, Golovina VA (2011) Dysregulation of Ca2+ signaling in astrocytes from mice lacking amyloid precursor protein. Am J Physiol Cell Physiol 300. https://doi.org/10.1152/ajpcell.00379.2010
Wang W, Gu L, Verkhratsky A, Peng L (2016) Ammonium increases TRPC1 expression via Cav-1/PTEN/AKT/GSK3β pathway. Neurochem Res 42. https://doi.org/10.1007/s11064-016-2004-z
Souza LB, Ambudkar IS (2014) Trafficking mechanisms and regulation of TRPC channels. Cell Calcium 56. https://doi.org/10.1016/j.ceca.2014.05.001
Notarachille G, Arnesano F, Calò V, Meleleo D (2014) Heavy metals toxicity: effect of cadmium ions on amyloid beta protein 1-42. Possible implications for Alzheimer’s disease. BioMetals 27:371–388. https://doi.org/10.1007/s10534-014-9719-6
Uddin MS, Kabir MT, Tewari D et al (2020) Revisiting the role of brain and peripheral Aβ in the pathogenesis of Alzheimer’s disease. J Neurol Sci 416:116974. https://doi.org/10.1016/j.jns.2020.116974
Uddin MS, Al Mamun A, Rahman MA et al (2020) Emerging proof of protein misfolding and interactions in multifactorial Alzheimer’s disease. Curr Top Med Chem:20. https://doi.org/10.2174/1568026620666200601161703
Yano K, Hirosawa N, Sakamoto Y et al (2003) 500 aggregations of amyloid beta-proteins in the presence of metal ions. Toxicol Lett 144:s134. https://doi.org/10.1016/s0378-4274(03)90499-1
Syme CD, Viles JH (2006, 1764) Solution 1H NMR investigation of Zn2+ and Cd2+ binding to amyloid-beta peptide (Abeta) of Alzheimer’s disease. Biochim Biophys Acta. https://doi.org/10.1016/j.bbapap.2005.09.012
Jiang LF, Yao TM, Zhu ZL et al (2007) Impacts of Cd(II) on the conformation and self-aggregation of Alzheimer’s tau fragment corresponding to the third repeat of microtubule-binding domain. Biochim Biophys Acta - Proteins Proteomics 1774:1414–1421. https://doi.org/10.1016/j.bbapap.2007.08.014
Butterfield DA (2002) Amyloid β-peptide (1-42)-induced oxidative stress and neurotoxicity: implications for neurodegeneration in Alzheimer’s disease brain. A review. Free Radic Res 36:1307–1313
Drake J, Link CD, Butterfield DA Oxidative stress precedes fibrillar deposition of Alzheimer’s disease amyloid beta-peptide (1–42) in a transgenic Caenorhabditis elegans model. Neurobiol Aging 24:415–420. https://doi.org/10.1016/s0197-4580(02)00225-7
Selkoe DJ (2001) Clearing the brain’s amyloid cobwebs. Neuron 32:177–180. https://doi.org/10.1016/S0896-6273(01)00475-5
Syme CD, Viles JH (2006) Solution 1H NMR investigation of Zn2+ and cd 2+ binding to amyloid-beta peptide (Aβ) of Alzheimer’s disease. Biochim Biophys Acta - Proteins Proteomics 1764:246–256. https://doi.org/10.1016/j.bbapap.2005.09.012
del Pino J, Zeballos G, Anadón MJ et al (2016) Cadmium-induced cell death of basal forebrain cholinergic neurons mediated by muscarinic M1 receptor blockade, increase in GSK-3β enzyme, β-amyloid and tau protein levels. Arch Toxicol 90:1081–1092. https://doi.org/10.1007/s00204-015-1540-7
Takashima A (2006) GSK-3 is essential in the pathogenesis of Alzheimer’s disease. J Alzheimers Dis 9:309–317
Basun H, Forssell L, Wetterberg L, Winblad B (1991) Metals and trace elements in plasma and cerebrospinal fluid in normal aging and Alzheimer’s disease. Undefined
Lui E, Wong C, Diaz F (1990) Metals and the liver in Alzheimer’s disease. An investigation of hepatic zinc, copper, cadmium, and metallothionein. J Am Geriatr Soc 38:633–639. https://doi.org/10.1111/j.1532-5415.1990.tb01421.x
Hart RP, Rose CS, Hamer RM (1989) Neuropsychological effects of occupational exposure to cadmium. J Clin Exp Neuropsychol 11:933–943. https://doi.org/10.1080/01688638908400946
Li X, Lv Y, Yu S et al (2012) The effect of cadmium on Aβ levels in APP/PS1 transgenic mice. Exp Ther Med 4:125–130. https://doi.org/10.3892/etm.2012.562
Mamun A, Uddin M, Mathew B, Ashraf G (2020) Toxic tau: structural origins of tau aggregation in Alzheimer’s disease. Neural Regen Res 15:1417. https://doi.org/10.4103/1673-5374.274329
Uddin MS, Kabir MT, Niaz K et al (2020) Molecular insight into the therapeutic promise of flavonoids against Alzheimer’s disease. Mol 25:1267. https://doi.org/10.3390/MOLECULES25061267
Mutter J, Naumann J, Sadaghiani C et al (2004) Alzheimer disease: mercury as pathogenetic factor and apolipoprotein E as a moderator. Neuroendocrinol Lett 25:331–339
Cedrola S, Guzzi GP, Ferrari D et al (2003) Inorganic mercury changes the fate of murine CNS stem cells. FASEB J 17:869–871. https://doi.org/10.1096/fj.02-0491fje
Kosik KS, Joachim CL, Selkoe DJ (1986) Microtubule-associated protein tau (τ) is a major antigenic component of paired helical filaments in Alzheimer disease. Proc Natl Acad Sci U S A 83:4044–4048. https://doi.org/10.1073/pnas.83.11.4044
Olivieri G, Brack C, Müller-Spahn F et al (2000) Mercury induces cell cytotoxicity and oxidative stress and increases β- amyloid secretion and tau phosphorylation in SHSY5Y neuroblastoma cells. J Neurochem 74:231–236. https://doi.org/10.1046/j.1471-4159.2000.0740231.x
Busciglio J, Lorenzo A, Yeh J, Yankner BA (1995) β-Amyloid fibrils induce tau phosphorylation and loss of microtubule binding. Neuron 14:879–888. https://doi.org/10.1016/0896-6273(95)90232-5
Mao P, Reddy PH (2011) Aging and amyloid beta-induced oxidative DNA damage and mitochondrial dysfunction in Alzheimer’s disease: implications for early intervention and therapeutics. Biochim Biophys Acta - Mol Basis Dis 1812:1359–1370
Goedert M, Spillantini MG, Jakes R et al (1989) Multiple isoforms of human microtubule-associated protein tau: sequences and localization in neurofibrillary tangles of Alzheimer’s disease. Neuron 3:519–526. https://doi.org/10.1016/0896-6273(89)90210-9
Zheng WH, Bastianetto S, Mennicken F et al (2002) Amyloid β peptide induces tau phosphorylation and loss of cholinergic neurons in rat primary septal cultures. Neuroscience 115:201–211. https://doi.org/10.1016/S0306-4522(02)00404-9
Huang HC, Jiang ZF (2009) Accumulated amyloid-β peptide and hyperphosphorylated tau protein: relationship and links in Alzheimer’s disease. J Alzheimers Dis 16:15–27
Ittner LM, Ke YD, Delerue F et al (2010) Dendritic function of tau mediates amyloid-β toxicity in Alzheimer’s disease mouse models. Cell 142:387–397. https://doi.org/10.1016/j.cell.2010.06.036
Monnet-Tschudi F, Zurich M-G, Boschat C et al Involvement of environmental mercury and lead in the etiology of neurodegenerative diseases. Rev Environ Health 21:105–117. https://doi.org/10.1515/reveh.2006.21.2.105
Charleston JS, Body RL, Bolender RP et al (1996) Changes in the number of astrocytes and microglia in the thalamus of the monkey Macaca fascicularis following long-term subclinical methylmercury exposure. Neurotoxicology 17:127–138
Smedman M, Potempska A, Rubenstein R et al (1997) Effects of cadmium, copper, and zinc and beta APP processing and turnover in COS-7 and PC12 cells. Relationship to Alzheimer disease pathology. Mol Chem Neuropathol 31:13–28. https://doi.org/10.1007/bf02815157
Song JW, Choi BS (2013) Mercury induced the accumulation of amyloid beta (Aβ) in PC12 cells: the role of production and degradation of Aβ. Toxicol Res 29:235–240. https://doi.org/10.5487/TR.2013.29.4.235
Eskes C, Honegger P, Juillerat-Jeanneret L, Monnet-Tschudi F (2002) Microglial reaction induced by noncytotoxic methylmercury treatment leads to neuroprotection via interactions with astrocytes and IL-6 release. Glia 37:43–52. https://doi.org/10.1002/glia.10019
Götz J, Chen F, Van Dorpe J, Nitsch RM (2001) Formation of neurofibrillary tangles in P301L tau transgenic mice induced by Aβ42 fibrils. Science (80- ) 293:1491–1495. https://doi.org/10.1126/science.1062097
Hardy J, Selkoe DJ (2002) The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science (80- ) 297:353–356. https://doi.org/10.1126/science.1072994
Oddo S, Caccamo A, Kitazawa M et al (2003) Amyloid deposition precedes tangle formation in a triple transgenic model of Alzheimer’s disease. In: Neurobiology of aging. Elsevier Inc., pp. 1063–1070
Hock C, Drasch G, Golombowski S et al (1998) Increased blood mercury levels in patients with Alzheimer’s disease. J Neural Transm 105:59–68. https://doi.org/10.1007/s007020050038
Ehmann WD, Markesbery WR, Alauddin M et al (1986) Brain trace elements in Alzheimer’s disease - PubMed. Neurotoxicology 7:195–206
Thompson CM, Markesbery WR, Ehmann WD et al (1988) Regional brain trace-element studies in Alzheimer’s disease - PubMed. Neurotoxicology 9:1–7
Mano Y, Takayanagi T, Ishitani A, Hirota T Mercury in hair of patients with ALS. Rinsho Shinkeigaku 29:844–848
Zahir F, Rizwi SJ, Haq SK, Khan RH (2005) Low dose mercury toxicity and human health. Environ Toxicol Pharmacol 20:351–360. https://doi.org/10.1016/j.etap.2005.03.007
Mutter J, Curth A, Naumann J et al (2010) Does inorganic mercury play a role in Alzheimer’s disease? A systematic review and an integrated molecular mechanism. J Alzheimers Dis 22:357–374. https://doi.org/10.3233/JAD-2010-100705
Haut MW, Morrow LA, Pool D et al (1999) Neurobehavioral effects of acute exposure to inorganic mercury vapor. Appl Neuropsychol 6:193–200. https://doi.org/10.1207/s15324826an0604_1
Chen C, Qu L, Li B et al (2005) Increased oxidative DNA damage, as assessed by urinary 8-hydroxy-2′- deoxyguanosine concentrations, and serum redox status in persons exposed to mercury. Clin Chem 51:759–767. https://doi.org/10.1373/clinchem.2004.042093
Hamilton ML, Guo Z, Fuller CD et al (2001) A reliable assessment of 8-oxo-2-deoxyguanosine levels in nuclear and mitochondrial DNA using the sodium iodide method to isolate DNA. Nucleic Acids Res 29:2117–2126. https://doi.org/10.1093/nar/29.10.2117
Sedelnikova OA, Horikawa I, Zimonjic DB et al (2004) Senescing human cells and ageing mice accumulate DNA lesions with unrepairable double-strand breaks. Nat Cell Biol 6:168–170. https://doi.org/10.1038/ncb1095
Von Figura G, Hartmann D, Song Z, Rudolph KL (2009) Role of telomere dysfunction in aging and its detection by biomarkers. J Mol Med 87:1165–1171
Bolin CM, Basha R, Cox D et al (2006) Exposure to lead and the developmental origin of oxidative DNA damage in the aging brain. FASEB J 20:788–790. https://doi.org/10.1096/fj.05-5091fje
Olivieri G, Novakovic M, Savaskan E et al (2002) The effects of beta-estradiol on SHSY5Y neuroblastoma cells during heavy metal induced oxidative stress, neurotoxicity and beta-amyloid secretion. Neuroscience 113:849–855. https://doi.org/10.1016/s0306-4522(02)00211-7
Bains JS, Shaw CA (1997) Neurodegenerative disorders in humans: the role of glutathione in oxidative stress-mediated neuronal death. Brain Res Rev 25:335–358
Selkoe DJ (1994) Cell biology of the amyloid β-protein precursor and the mechanism of Alzheimer’s disease. Annu Rev Cell Biol 10:373–403
Wu J, Basha MR, Brock B et al (2008) Alzheimer’s disease (AD)-like pathology in aged monkeys after infantile exposure to environmental metal lead (Pb): evidence for a developmental origin and environmental link for AD. J Neurosci 28:3–9. https://doi.org/10.1523/JNEUROSCI.4405-07.2008
Basha MR, Wei W, Bakheet SA et al (2005) The fetal basis of amyloidogenesis: exposure to lead and latent overexpression of amyloid precursor protein and β-amyloid in the aging brain. J Neurosci 25:823–829. https://doi.org/10.1523/JNEUROSCI.4335-04.2005
Bihaqi SW, Huang H, Wu J, Zawia NH (2011) Infant exposure to lead (Pb) and epigenetic modifications in the aging primate brain: Implications for Alzheimer’s disease. J Alzheimers Dis 27:819–833. https://doi.org/10.3233/JAD-2011-111013
Davey FD, Breen KC (1998) The interaction between chronic low-level lead and the amyloid Β precursor protein. Amyloid 5:90–98. https://doi.org/10.3109/13506129808995286
Sharma SV , Atam V , Verma A , Kumar P , Murthy RC et al (2015) Lead (Pb) toxicity trigger schizophrenia in battery workers of North Region of India. J Neurol Neurol Disord 2:1–6.
Matte TD, Figueroa JP, Ostrowski S et al (1989) Lead poisoning among household members exposed to lead-acid battery repair shops in Kingston, Jamaica. Int J Epidemiol 18:874–881. https://doi.org/10.1093/ije/18.4.874
Pourabdian S, Eizadi-Mood N, Golshiri P, Amini F (2011) The relationship between blood lead level and neuro-psychological and hematological findings in lead-exposed workers of battery industry. Iran J Toxicol 5:521–526
Jaishankar M, Tseten T, Anbalagan N et al (2014) Toxicity, mechanism and health effects of some heavy metals. Interdiscip Toxicol 7:60–72. https://doi.org/10.2478/intox-2014-0009
Kawahara M, Kato-Negishi M (2011) Link between aluminum and the pathogenesis of Alzheimer’s disease: the integration of the aluminum and amyloid cascade hypotheses. Int J Alzheimers Dis. https://doi.org/10.4061/2011/276393
Wu Z, Du Y, Xue H et al (2012) Aluminum induces neurodegeneration and its toxicity arises from increased iron accumulation and reactive oxygen species (ROS) production. Neurobiol Aging 33:199.e1-199.e12. https://doi.org/10.1016/j.neurobiolaging.2010.06.018
Nampoothiri M, John J, Kumar N et al (2015) Modulatory role of simvastatin against aluminium chloride-induced behavioural and biochemical changes in rats. Behav Neurol 2015:210169. https://doi.org/10.1155/2015/210169
Klatzo I, Wisniewski H, Streicher E (1965) Experimental production of neurofibrillary degeneration: I Light microscopic observations. J Neuropathol Exp Neurol 24:187–199. https://doi.org/10.1097/00005072-196504000-00002
Crapper DR, Krishnan SS, Dalton AJ (1973) Brain aluminum distribution in Alzheimer’s disease and experimental neurofibrillary degeneration. Science (80- ) 180:511–513. https://doi.org/10.1126/science.180.4085.511
Lidsky TI (2014) Is the aluminum hypothesis dead? J Occup Environ Med 56. https://doi.org/10.1097/JOM.0000000000000063
Fattoretti P, Bertoni-Freddari C, Balietti M et al (2004) Chronic aluminum administration to old rats results in increased levels of brain metal ions and enlarged hippocampal mossy fibers. In: Annals of the New York Academy of Sciences. New York Academy of Sciences, pp. 44–47
Praticò D, Uryu K, Sung S et al (2002) Aluminum modulates brain amyloidosis through oxidative stress in APP transgenic mice. FASEB J 16:1138–1140. https://doi.org/10.1096/fj.02-0012fje
Exley C, Price NC, Kelly SM, Birchall JD (1993) An interaction of β-amyloid with aluminium in vitro. FEBS Lett 324:293–295. https://doi.org/10.1016/0014-5793(93)80137-J
Kawahara M, Kato M, Kuroda Y (2001) Effects of aluminum on the neurotoxicity of primary cultured neurons and on the aggregation of β-amyloid protein. Brain Res Bull 55:211–217. https://doi.org/10.1016/S0361-9230(01)00475-0
Sato N, Hori O, Yamaguchi A et al (1999) A novel presenilin-2 splice variant in human Alzheimer’s disease brain tissue. J Neurochem 72:2498–2505. https://doi.org/10.1046/j.1471-4159.1999.0722498.x
Sakamoto T, Saito H, Ishii K et al (2006) Aluminum inhibits proteolytic degradation of amyloid β peptide by cathepsin D: a potential link between aluminum accumulation and neuritic plaque deposition. FEBS Lett 580:6543–6549. https://doi.org/10.1016/j.febslet.2006.10.075
Liang RF, Li WQ, Wang H et al (2013) Impact of sub-chronic aluminium-maltolate exposure on catabolism of amyloid precursor protein in rats. Biomed Environ Sci 26:445–452. https://doi.org/10.3967/0895-3988.2013.06.005
Yamamoto H, Saitoh Y, Yasugawa S, Miyamoto E (1990) Dephosphorylation of r factor by protein phosphatase 2A in synaptosomal cytosol fractions, and inhibition by aluminum. J Neurochem 55:683–690. https://doi.org/10.1111/j.1471-4159.1990.tb04187.x
Yokel RA (2006) Blood-brain barrier flux of aluminum, manganese, iron and other metals suspected to contribute to metal-induced neurodegeneration. J Alzheimers Dis 10:223–253. https://doi.org/10.3233/JAD-2006-102-309
Vanduyn N, Settivari R, Levora J et al (2013) The metal transporter SMF-3/DMT-1 mediates aluminum-induced dopamine neuron degeneration. J Neurochem 124:147–157. https://doi.org/10.1111/jnc.12072
Yamanaka K, Minato N, Iwai K (1999) Stabilization of iron regulatory protein 2, IRP2, by aluminum. FEBS Lett 462:216–220. https://doi.org/10.1016/S0014-5793(99)01533-1
Acknowledgements
This work was funded by Deanship of Scientific Research at Princess Nourah bint Abdulrahman University through the Fast-Track Research Funding Program. This work was supported by King Saud University, Deanship of Scientific Research, College of Science Research Center.
Author information
Authors and Affiliations
Contributions
MSU conceived the original idea and designed the outlines of the study. MTK and MSU wrote the draft of the manuscript and prepared the figures for the manuscript. SZ, YB, GMA, MNB-J, SGB, SAM, and MMA-D revised and improved the manuscript. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Kabir, M.T., Uddin, M.S., Zaman, S. et al. Molecular Mechanisms of Metal Toxicity in the Pathogenesis of Alzheimer’s Disease. Mol Neurobiol 58, 1–20 (2021). https://doi.org/10.1007/s12035-020-02096-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-020-02096-w