
Abstract
In this study, we aimed to clarify the forest structure and species composition of a coastal sand dune and the relationship between topographical factors and woody plant distribution at the Bang Boet coastal sand dune, Southern Thailand. Three permanent transect plots (10 × 100 m) covering both windward and leeward aspects were established 300 m apart. In subplots (10 × 10 m) of each transect, all woody plants with a diameter at breast height larger than 1 cm were tagged, measured, and identified. The topographic factors were recorded for each subplot. Non-multidimensional scaling was employed to analyse the woody plant distribution, which was tested for statistical significance. We identified 36 species from 32 genera and 23 families. Species diversity was moderate based on the Shannon–Wiener index (2.81). The density and basal area of all woody plants was 3110 ± 361 stem·ha−1 and 7.34 ± 0.87 m2·ha−1, respectively. The dominant trees species based on the importance value index were Pandanus odorifer, Syzygium antisepticum and Syzygium grande. The forest structure could be classified into two canopy layers, with an open and closed crown canopy on the windward and leeward aspects, respectively. Overall, the distribution of woody plants was significantly correlated with aspect. Species diversity, tree abundance, and basal area differed significantly between the windward and leeward aspects and quantitative values were low on the windward aspect. Due to strong winds and salt spray on the windward side, the woody plant community and distribution favoured the leeward side.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Overexploitation, habitat modification, and pollution have caused estuarine and coastal zone degradation, biodiversity loss, and a decline in ecological resilience (Lotze et al. 2006). Coastal zones are important because they support a wide range of contrasting ecosystems, such as coral reefs, lagoons, seagrass meadows, kelp forests, mangrove wetlands, sand dunes, and other types of coastal vegetation. Sand dunes consist of ridges or mounds formed by wind deposition of loose sand (Pye and Tsoar 2009) or aeolian systems that develop on sandy beaches (Martínez et al. 2004; Lancaster 2009). Sand dunes bear the full force of coastal winds, which move loose sand, burying plants and exposing their roots. Moreover, the sand surface can reach 60–70 °C under full sun exposure (Louge et al. 2013). The harsh dune environment changes from the windward (front) to the leeward (back) aspect, providing unique habitats for plants and wildlife. Native dune vegetation can reduce wind speed, thereby stabilising and trapping loose sand particles (Levin and Ben-Dor 2004; Tsoar 2005). In addition, this ecosystem protects human developments and is an important habitat for mediating the negative impacts of global climate change, including rising sea levels, large storms, and increased erosion (Cochard et al. 2008). However, it is a fragile ecosystem that is sensitive to human activities, especially tourism-driven land development.
Coastal sand dunes are useful for studying the interactions between plant species and their environment from an initial successional stage. For instance, soil is the most important factor for plant establishment. On coastal sand dunes, the soil is very fine grained (Prachantasen et al. 2008) and extremely porous, has low water-holding capacity (Yair et al. 2008), and may have increased salinity due to salt spray (Sevink 1991). It can cause burial, abrasion, drought, and nutrient stress in plants, especially on the foredune (Hesp and Martinez 2008). Wind is considered the main factor influencing sand movement (McLachlan 1991; Çakan et al. 2011) and is an important factor in controlling the distribution, establishment, and characteristics of plant communities (Lubke 1983; Moreno-Casasola 1986; Maun 2009). In addition, salt spray has an important role in determining plant succession on coastal sand dunes (Hesp 1991; Wilson and Sykes 1999; Griffiths and Orians 2003; Maun 2009), and most plant species are distributed based on their tolerance to salt, with those growing near the coast being more tolerant (Yura 1997). Furthermore, topography can protect plants from wind and salt spray and is related to soil nutrients; therefore, it may be an important determinant of plant distribution. These factors may act alone or in combination, and the impacts of each vary according to local environmental conditions. Thus, plant establishment on coastal sand dunes is mainly initiated by primary succession based on these crucial environmental factors (Robertson and Vitousek 1981; Lichter 2000; Walker and del Moral 2003; Álvarez-Molina et al. 2012), especially the presence of grass and some herbaceous species that function to build sand dunes due to their extensive root systems and ability to grow under constant sand burial (Botkin 1993; Maun 1998). Plant community complexes on coastal sand dunes result from the interaction between plant tolerance and the environment, with climate, rainfall, growth season, and dry periods as the main factors (Maun 2009).
In Thailand, the Bang Boet coastal sand dune is one of the largest sand dunes located on the inner gulf of Thailand. It was created by wind-blown sand under dry conditions, probably during and after the mid-Holocene 2220–2960 years ago (Lertnok et al. 2010), and the sediments on the sand dune were deposited by unusual storms about 100 years ago. However, this area has not been affected by typhoons in the past four decades (Prachantasen et al. 2008). Vegetation establishment on this sand dune may be affected by environmental factors similar to those affecting other coastal sand dunes, such as strong wind, salt spray, and soil. In general, coastal woody species on coastal sand dunes in Southern Thailand include Casuarina equisetifolia, Terminalia catappa, Hibiscus tiliaceus, and Premna serratifolia (Hayasaka and Fujiwara 2005). Meanwhile, the dominant species of terrestrial coastal forest along the eastern coast of peninsular Thailand are Syzygium antisepticum and Syzygium grande (Sridith and Laongpol 2003). Coastal grassland includes Ipomoea pes-caprae, Ipomoea imperati, Fimbristylis sericea, and Cyperus stoloniferus, while coastal scrub is composed of woody plants such as Sindora siamensis, Planchonella obovata, and S. grande (Laongpol et al. 2009). However, the interaction between plant species distribution and environmental factors is not well documented, and the destabilisation of sand dunes, especially due to damaged vegetation (e.g. during road construction, sand stabilisation, and invasion of non-native plants) requires study. Therefore, in this study, we aimed to clarify the forest structure and species composition of the Bang Boet coastal sand dune in Thailand and the relationship between topographic factors and woody plant distribution.
Materials and methods
Study site
Study site was revised as “The study was conducted on the Bang Boet natural coastal sand dune (10°55′22″–10°56′6″N, 99°29′25″–99°29′49″E), Pak Klong sub-district, Pathio district, Chumphon Province, Southern Thailand (Fig. 1), 425 km south of Bangkok. The highest elevation is approximately 20 m above mean sea level (Lertnok et al. 2010; Choowong 2011). The Bang Boet coastal sand dune experiences a tropical monsoon climate with three main seasons: rainy, winter, and dry seasons. The wet season lasts 6 months from May to October and is influenced by the southwest monsoon. Winter occurs between November and January and is influenced by the northeast monsoon. The dry season occurs between February and April and is under the influence of the humid and hot southeast monsoon. The mean annual rainfall between 1990 and 2017 was 1730.52 mm, with a minimum of 1148.10 mm in 1994 and a maximum of 2656.50 mm in 1996. The mean monthly temperature over this period was 24.83 °C, with a minimum of 19.12 °C in April and a maximum of 30.54 °C in December. The soil is Bacho seties, sandy (99.55%), and rather acidic (pH 4.20–6.30), and has a low water-holding capacity and a low nutrient content (Marod et al. 2020)”.

Study site at Bang Boet coastal sand dune on the inner gulf of Thailand
Data collection
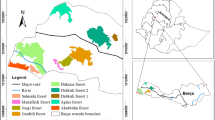
Three permanent transect plots that crossed the windward and leeward aspects were established in 2012 (Fig. 1). Each transect was 10 × 100 m, with 300 m between transects. Subplots of 10 × 10 m were created, yielding 30 subplots, and all woody plants with a diameter at breast height (DBH) larger than 1 cm were tagged, measured, and identified to the species level. Identification of specimens was confirmed by the Forest Herbarium, Department of National Parks, Wildlife and Plants Conservation, and species nomenclature was based on Smitinand (2014). Stratification based on Richards (1952) was used to describe the vertical and horizontal plant community structure. The topographic attribute of elevation was measured using a dumpy level in the four corners of each subplot. Mean elevation, convexity, slope, and aspect were calculated according to Legendre et al. (2009).
Data analysis
Data analysis was revised as “Species richness (at the species, genus, and family levels) was assessed for all transects. Species diversity based on the Shannon–Wiener (H’) index and Fisher’s alpha diversity index (Shannon and Weaver 1949; Fisher et al. 1943) was analysed using the diversity and fisher.alpha function, respectively, in the vegan package (Oksanen et al. 2015). The forest horizontal structure was described in terms of woody plant density (stem·ha-1), basal area (m2·ha-1), and DBH class distribution. To detect the dominant species, saplings (1 ≤ DBH < 5 cm) and trees (DBH ≥ 5 cm) were used to analyse the importance value index (IVI). The IVI was calculated as the sum of three indices: the relative dominance in basal area, relative density, and relative frequency (Curtis and McIntosh 1950). To detect the relationship between topographic factors and woody plant distribution, ordination analysis based on non-metric multidimensional scaling (NMDS) was used. The ordination dimension selected was that which provided lower stress. The data were organised into subplot × species matrices with the cells filled with the abundance values of each species and into subplot × topographic matrices with the cells filled with the values of each factor. NMDS was used with the metaMDS function as implemented in the vegan package and Bray–Curtis dissimilarity as the metric distance (Oksanen et al. 2015). Subsequently, the envfit function over the NMDS ordination was used to fit vectors representing the sand dune communities. Finally, the significance of the factors was tested using the permutation test (9999 permutations). Then, one-way analysis of similarity (ANOSIM) was used to test for significant differences in the composition patterns of woody plant species revealed in the NMDS ordination (Clarke 1993). The data were organised into subplot × species matrices with the cells filled with the abundance values of each species and into subplot × side matrices with the cells filled with the code side of each subplot. ANOSIM was performed using the anosim function as implemented in the vegan package (Oksanen et al. 2015). The Wilcoxon statistical test was used to identify significant differences among the community types on the coastal sand dune. All analyses were performed using the R data analysis package ver. 3.2.5 (R Development Core Team 2016).”
Results
Forest structure and species composition
Woody plants consisted of 933 stems representing 36 species from 32 genera and 23 families (Appendix Table 3). The dominant families based on species numbers were Myrtaceae (six species) and Rubiaceae (five species), whereas other families were represented by only one or two species. The genus Syzygium had the highest number of species (four species), whereas other genera were represented by only one or two species. The woody plant density for all woody plant species, trees, and sapling stages varied among the transects (Table 1). The mean (± standard deviation) density and mean basal area for all woody plants were 3110 ± 361 stem·ha−1 and 7.34 ± 0.87 m2·ha−1, respectively. The diameter class distribution was significantly represented by a negative power growth form (Fig. 2). Moderate plant species diversity was found based on the H′ index (2.81) and Fisher’s alpha index (7.45).

Diameter class distribution of all woody plants on the Bang Boet coastal sand dune
The dominant species were mostly saplings and trees. Of the trees, Pandanus odorifer had the highest IVI (53.20%), followed by Syzygium antisepticum (46.97%), Syzygium grande (39.39%), Planchonella obovata (32.34%), Mischocarpus sundaicus (20.92%), Sindora siamensis (16.92%), Syzygium claviflorum (13.67%), Chaetocarpus castanocarpus (12.19%), Suregada multiflora (11.74%), and Atalantia monophylla (10.27%). Of the saplings (including shrubs and small trees), M. sundaicus had the highest IVI (55.39%), followed by Eurycoma longifolia (33.45%), Ochna integerrima (26.02%), S. antisepticum (24.27%), Scaevola taccada (18.19%), S. siamensis (17.79%), P. obovata (16.16%), Aporosa planchoniana (14.95%), Champereia manillana (11.02%), and S. grande (9.06%).
Two canopy layers, a top and a middle layer, were apparent, and there was a large amount of overlap between the layers. Correspondingly, trees higher than 7 m tended to be grouped separately from shorter trees. The top layer (height range: 7–12 m) consisted of predominantly adults of species such as P. obovata, S. grande, S. antisepticum, S. multiflora, P. odorifer, and Casuarina equisetifolia. The middle layer (< 7 m) was dominated by shrubby trees and shrubs such as M. sundaicus, Aporosa planchoniana, O. integerrima, A. monophylla, E. longifolia, and S. taccada. The windward and leeward sides were characterised by an open and a closed canopy, respectively (Fig. 3).

Profile diagram across the altitudinal gradient of the Bang Boet coastal sand dune
Relationship between topographic factors and woody plant distribution
The NMDS analysis revealed lower stress (0.075) for the two-dimensional ordination, which was chosen as the most appropriate NMDS ordination. The first and second axes revealed two distinct groups of subplots in the Bang Boet community (Fig. 4). The windward group included 14 subplots (P1, P2, P3, P8, P11, P12, P13, P14, P15, P16, P21, P23, P24, and P25), the leeward group included 12 subplots, and the remaining four plots contained no woody plants. This pattern was confirmed by the results of the one-way ANOSIM, which indicated that woody plant species composition differed significantly between the windward and leeward sides (r = 0.335, P = 0.001). However, the Wilcoxon tests of species diversity, tree abundance and basal area differed significantly between the windward and leeward aspects (W = 10, W = 12, and W = 21, P < 0.001, respectively), with lower quantitative values on the windward side.

Non-metric multidimensional scaling (NMDS) of woody plant distribution with topographic variables (aspect, elevation, slope, and convexity). P and the corresponding ranking number indicate the subplot number from subplot 1 (P1) to 30 (P30). Appendix Table 3 lists the species codes
Two topographic factors significantly affected woody plant distribution (Table 2), but the correlations between the axes of the topographic factors differed. The distribution of all woody plant species was significantly and positively correlated with aspect (R2 = 0.488, P < 0.001), but not with elevation, slope, and convexity. Therefore, aspect could be used to identify the woody plant distribution at the study site. These results revealed that the species composition and community structure pattern were differentiated along the direction of sand dune advancement, where the leeward aspect had greater plant establishment than the windward aspect.
Discussion
Woody plant community
The woody plant species on the Bang Boet coastal sand dune were mostly similar to the species found in beach forests in Thailand. Except for Casuarina equisetifolia, Pandanus odorifer, and Scaevola taccada found at the study site, the remaining species are common to other tropical to subtropical regions (Appendix Table 3). C. equisetifolia inhabits Australia and other Pacific subtropical areas (Lin et al. 2017). P. odorifer inhabits India, Sri Lanka, and the Maldives in the east to southern China and south through Indochina and the Philippines to Indonesia. The species has also been introduced elsewhere, including eastern Africa (Allen 2011). Finally, S. taccada inhabits tropical and subtropical Indo-Pacific coastal land, from East Africa to Oceania and Japan (Tanaka et al. 2015).
Tree size distribution is related to concurrently measured demographic rates across species (Lima et al. 2016) when describing successional pathways and structural development (Zenner 2005) and predicting future forest stand structure (Feeley et al. 2007). In the present study, as the diameter increased, the number of stems decreased (Fig. 2), indicating that the regeneration process of the plant community was associated with a seral succession stage.
High IVI values indicate that the dominant species are distributed across a wide area (Curtis and McIntosh 1950). In the present study, P. odorifer, Syzygium antisepticum, Syzygium grande, Planchonella obovata, and Mischocarpus sundaicus were indicative of the Bang Boet coastal sand dune. In addition, C. equisetifolia and Hibiscus tiliaceus are generally found on coastal sand dunes in Southern Thailand (Hayasaka and Fujiwara 2005).
Tropical tree species adapted to high-wind environments might be expected to differ systematically in terms of stem allometry and life-history patterns compared to species found in less windy forests (Thomas et al. 2015). Some researchers have reported that strong winds substantially modify tree size and shape along the coast, as evidenced by the development of scrub and branches on only the leeward side (Telewski 1995). In this study, S. grande, S. antisepticum, and Sindora siamensis had characteristics of scrub trees. In contrast, C. equisetifolia and P. odorifer were highly resistant and could easily establish near the sea (Fig. 3). C. equisetifolia is a distinctive species that adapts to coastal stress by altering its tree shape and dry mass density (Lin et al. 2017). Meanwhile, P. odorifer, which had the highest IVI, is adapted to coastal stress. Forests with multiple vertical structures on beaches interspersed with old-growth and regrowth Casuarina stands, dense P. odorifer, and other trees and shrub species may be more resilient to tsunamis (Tanaka et al. 2007).
Woody plant distribution
Topographical habitat is instrumental in determining species composition and community structure (Addo-Fordjour and Rahmad 2015). The present study showed that species diversity, tree abundance, and basal area were differentiated based on aspect. Aspect is usually related to different environmental factors in topographical habitats, especially wind speed and direction (le Roux and McGeoch 2008). According to Qian et al. (2009), wind speed is a strong influence on the windward, rather than the leeward, side. Moreover, the windward aspect of coastal sand dunes is exposed to salt spray, and wind speed is greater than on the leeward aspect (Maun 2009), making wind and salt spray important factors in controlling the distribution, establishment, and characteristics of plants communities (Lubke 1983; Moreno-Casasola 1986; Wilson and Sykes 1999; Maun 2009). In the present study, the leeward side had a higher woody plant distribution and greater establishment by Planchonella obovata (PLANOB), Mischocarpus sundaicus (MISCSU), Syzygium claviflorum (SYZYCL), Chaetocarpus castanocarpus (CHAECA), and Suregada multiflora (SUREMU) compared with the windward side, as shown in Fig. 4. This pattern indicated that strong wind prevents tree establishment on coastal sand dunes, and only a few highly adapted species can succeed, such as Pandanus odorifer (PANDOD), Casuarina equisetifolia (CASUEQ), Hibiscus tiliaceus (HIBITI), and Scaevola taccada (SCAETA).
On coastal sand dunes, shrubs and grasses mainly occupy the windward, rather than the leeward, side (Isermann 2011; Peyrat and Fichtner 2011), particularly S. taccada, which is a coastal shrub with thick fleshy leaves that dominates beach crests and is highly salt and wind tolerant. The presence of a salt-tolerant S. taccada hedge along the coast can result in increased growth of other tree species (Komdeur and Pels 2005). H. tiliaceus is a fast-growing and wind- and salt-resistant coastal shrub or shrubby tree that has adapted to a wide range of environments, and it has been used to stabilise sand dunes and form coastal windbreaks (Tang et al. 2003). In addition, some large tree species (e.g. C. equisetifolia) are dominant tree species due to their pioneering characteristics, including fast growth, adaptability to barren soils, and wind resistance (Liu et al. 2014). C. equisetifolia is important in terms of sand dune stabilisation (Karthikeyan et al. 2013), and many researchers have recommended that it be planted as a shelter belt (Harvey et al. 2004; De Zoysa 2008; Atangana et al. 2014). However, the specimens in the present study were small, and only a few individuals were identified, even though it is recognised as an important species, is widespread along beaches in Southeast Asia (Wong 2005), and it is a common coastal species on coastal sand dunes in Southern Thailand (Hayasaka and Fujiwara 2005). Such facilitator or builder species can stabilise shifting sand and lead to the development of a foredune or windward break, giving way to other herbaceous vegetation, which in turn is taken over by shrubs that may be replaced by trees (Martínez et al. 2001). The results of the present study support the high succession of the woody plant community and distribution on the leeward rather than the windward side on the Bang Boet coastal sand dune.
Conclusions
Low species diversity (36 species from 32 genera and 23 families) was found on the Bang Boet coastal sand dune, Thailand. The DBH class distribution of all woody plant species was negative and approximated a power growth form. The relationship between topographic factors and woody plant distribution was significantly associated with aspect, and most species were distributed on the leeward, rather than the windward, side of the dune. The tree species Pandanus odorifer, which was the dominant species, inhabited both aspects due to its high wind resistance. The other dominant species were mainly shrubby trees and trees such as Syzygium antisepticum and Syzygium grande, which were also well established, especially on the leeward side. These results indicate that strong winds influenced the effects of aspect at the study site, which is an important factor in determining the woody plant community and its distribution on this coastal sand dune.
References
Addo-Fordjour P, Rahmad ZB (2015) Environmental factors associated with liana community assemblages in a tropical forest reserve, Ghana. J Trop Ecol 31(1):69–79 https://doi.org/10.1017/S0266467414000522
Allen DJ (2011) Pandanus odorifer. The IUCN Red List of Threatened Species 2011: https://doi.org/10.2305/IUCN.UK.20112.RLTS.T199828A9128592.en.
Álvarez-Molina LL, Martínez ML, Pérez-Maqueo O, Gallego-Fernández JB, Flores P (2012) Richness, diversity, and rate of primary succession over 20 years in tropical coastal dunes. Plant Ecol 213(10):1597–1608 https://doi.org/10.1007/s11258-012-0114-5
Atangana A, Khasa D, Chang S, Degrande A (2014) Tropical agroforestry. Springer, Dordrecht, 380 pp
Botkin DB (1993) Forest dynamics: an ecological model. Oxford University Press, Oxford, 309 pp
Çakan H, Yılmaz KT, Alphan H, Ünlükaplan Y (2011) The classification and assessment of vegetation for monitoring coastal sand dune succession: the case of Tuzla in Adana, Turkey. Turk J Bot 35(6):697–711
Choowong M (2011) Quaternary. In: Ridd MF, Barber AJ, Crow MJ (eds) Geology of Thailand. Geological Society of London, London, pp 335–350
Clarke KR (1993) Non-parametric multivariate analysis of changes in community structure. Australian J Ecol 18(1):117–143 https://doi.org/10.1111/j.1442-9993.1993.tb00438.x
Cochard R, Ranamukhaarachchi SL, Shivakoti GP, Shipin OV, Edwards PJ, Seeland KT (2008) The 2004 tsunami in Aceh and southern Thailand: a review of coastal ecosystems, wave hazards and vulnerability. Perspect Plant Ecol 10(1):3–40 https://doi.org/10.1016/j.ppees.2007.11.001
Curtis JT, McIntosh RP (1950) The interrelations of certain analytic and synthetic phytosociological characters. Ecology 31(3):434–455 https://doi.org/10.2307/1931497
De Zoysa M (2008) Casuarina coastal forest shelterbelts in Hambantota City, Sri Lanka: assessment of impacts. Small-Scale For 7(1):17–27 https://doi.org/10.1007/s11842-008-9038-2
Feeley KJ, Davies SJ, Noor MNS, Kenneth AR, Tan S (2007) Do current stem size distributions predict future population changes? An empirical test of intraspecific patterns in tropical trees at two spatial scales. J Trop Ecol 23(2):191–198 https://doi.org/10.1017/S0266467406003919
Fisher RA, Corbet AS, Williams CB (1943) The relation between the number of species and the number of individuals in a random sample of animal population. J Anim Ecol 12:42–58 https://doi.org/10.2307/1411
Griffiths ME, Orians CM (2003) Salt spray differentially affects water status, necrosis, and growth in coastal sand plain heathland species. Am J Bot 90(8):1188–1196 https://doi.org/10.3732/ajb.90.8.1188
Harvey CA, Tucker NIJ, Estrada A (2004) Live fences, isolated trees and windbreaks: tools for conserving biodiversity in fragmented tropical landscapes. In: Schroth G, da Fonseca GAB, Harvey CA, Gascon C, Vasconcelos HL, Izac AMN (eds) Agroforestry and biodiversity conservation in tropical landscapes. Island Press, Washington, DC, pp 261–289
Hayasaka D, Fujiwara K (2005) Species composition and environmental factors, including human impacts, on coastal sand-dunes and maritime strand-forests in southern Thailand. Tropics 14(3):245–254 https://doi.org/10.3759/tropics.14.245
Hesp PA (1991) Ecological processes and plant adaptations on coastal dunes. J Arid Environ 21(2):165–191
Hesp PA, Martinez MLM (2008) Transverse dune trailing ridges and vegetation succession. Geomorphology 99(1–4):205–213 https://doi.org/10.1016/j.geomorph.2007.11.001
Isermann M (2011) Patterns in species diversity during succession of coastal dunes. J Coastal Res 27(4):661–671 https://doi.org/10.2112/JCOASTRES-D-09-00040.1
Karthikeyan A, Chandrasekaran K, Geetha M, Kalaiselvi R (2013) Growth response of Casuarina equisetifolia Forst. Rooted stem cuttings to Frankia in nursery and field conditions. J Biosci 38(4):741–747 https://doi.org/10.1007/s12038-013-9362-3
Komdeur J, Pels DM (2005) Rescue of the Seychelles warbler on Cousin Island, Seychelles: the role of habitat restoration. Biol Conserv 124(1):15–26 https://doi.org/10.1016/j.biocon.2004.12.009
Lancaster N (2009) Dune morphology and dynamics. In: Parsons AJ, Abrahams AD (eds) Geomorphology of desert environments, 2nd edn. Springer, Dordrecht, pp 557–595
Laongpol C, Suzuki K, Katzensteiner K, Sridith K (2009) Plant community structure of the coastal vegetation of peninsular Thailand. Thai Forest Bulletin (Botany) 37(1):106–133
Legendre P, Mi X, Ren H, Ma K, Yu M, Sun I-F, He F (2009) Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 90(3):663–674 https://doi.org/10.1890/07-1880.1
le Roux PC, McGeoch MA (2008) Spatial variation in plant interactions across a severity gradient in the sub-Antarctic. Oecologia 155(4):831–844 https://doi.org/10.1007/s00442-007-0954-1
Lertnok W, Choowong M, Thitimakorn T (2010) Geomorphology and ground penetrating radar profiles of Holocene coastal dune, western coastal plain of the gulf of Thailand. Bulletin of Earth Sciences of Thailand 3:17–27
Levin N, Ben-Dor E (2004) Monitoring the dune stabilization process along the coastal dunes of Ashdod-Nizanim, Israel, 1945–1999. J Arid Environ 58(3):335–355 https://doi.org/10.1016/j.jaridenv.2003.08.007
Lichter J (2000) Colonization constraints during primary succession on coastal Lake Michigan sand dunes. J Ecol 88(5):825–839 https://doi.org/10.1046/j.1365-2745.2000.00503.x
Lima RAF, Muller-Landau HC, Prado PI, Condit R (2016) How do size distributions relate to concurrently measured demographic rates? Evidence from over 150 tree species in Panama. J Trop Ecol 32(3):179–192 https://doi.org/10.1017/S0266467416000146
Lin Y, Li J, Bakker JD, Lin H, Chen C, Hong W, Fisher L, Wu C, Hong T, Deng H, Zhang G, Du K (2017) Wind and salt spray alter tree shape and dry mass density in Casuarina equisetifolia L. Trees 31(1):15–26 https://doi.org/10.1007/s00468-016-1450-1
Liu X, Lu Y, Xue Y, Zhang X (2014) Testing the importance of native plants in facilitation the restoration of coastal plant communities dominated by exotics. Forest Ecol Manag 322:19–26 https://doi.org/10.1016/j.foreco.2014.03.020
Lotze HK, Lenihan HS, Bourque BJ, Bradbury RH, Cooke RG, Kay MC, Kidwell SM, Kirby MX, Peterson CH, Jackson JBC (2006) Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312(5781):1806–1809 https://doi.org/10.1126/science.1128035
Louge MY, Valance A, Ould el-Moctoar A, Xu J, Hay AG, Richer R (2013) Temperature and humidity within a mobile barchan sand dune, implications for microbial survival. J Geophys Res-Earth 118(4):2392–2405 https://doi.org/10.1002/2013JF002839
Lubke RA (1983) A survey of the coastal vegetation near Port Alfred, eastern cape. Bothalia 14(3/4):725–738 https://doi.org/10.4102/abc.v14i3/4.1234
Marod D, Sungkaew S, Mizunaga H, Thongsawi J (2020) Association of community-level traits with soil properties in a tropical coastal sand dune. Environ Nat Resour J 18(1):101–109 https://doi.org/10.32526/ennrj.18.1.2020.10
Martínez ML, Psuty NP, Lubke RA (2004) A perspective on coastal dunes. In: Martínez ML, Psuty NP (eds) Coastal dunes: ecology and conservation. Springer, Berlin/Heidelberg, pp 3–10
Martínez ML, Vazquez G, Sanchez-Colon S (2001) Spatial and temporal variability during primary succession on tropical coastal sand dunes. J Veg Sci 12(3):361–372 https://doi.org/10.2307/3236850
Maun MA (1998) Adaptations of plants to burial in coastal sand dunes. Can J Botany 76(5):713–738 https://doi.org/10.1139/b98-058
Maun MA (2009) The biology of coastal sand dunes. Oxford University Press, Oxford, 265 pp
McLachlan A (1991) Ecology of coastal dunes fauna. J Arid Environ 21:229–243
Moreno-Casasola P (1986) Sand movement as a factor in the distribution of plant communities in a coastal dune system. Vegetatio 65(2):67–76 https://doi.org/10.1007/BF00044876
Oksanen J, Blanchet G, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2015) Vegan: Community Ecology Package. R Package Version 2.2-0. http://CRAN.Rproject.org/package=vegan
Peyrat J, Fichtner A (2011) Plant species diversity in dry coastal dunes of the southern Baltic coast. Community Ecol 12(2):220–226 https://doi.org/10.1556/ComEc.12.2011.2.10
Prachantasen N, Choowong M, Pailoplee S, Phantuwongraj S (2008) Sedimentary characteristics of sand dune from bang Berd, Chumphon Province, southern Thailand. Bulletin of Earth Sciences of Thailand 1(1–2):28–34
Pye K, Tsoar H (2009) Aeolian sand and sand dunes. Springer, Berlin/Heidelberg, 458 pp
Qian G, Dong Z, Luo W, Wang H (2009) Variations of horizontal and vertical velocities over two-dimensional transverse dunes: a wind tunnel simulation of the effect of windward slope. J Arid Environ 73(12):1109–1116 https://doi.org/10.1016/j.jaridenv.2009.06.006
R Development Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Richards PW (1952) The tropical rain forest: an ecological study. Cambridge University Press, Cambridge, 450 pp
Robertson GP, Vitousek PM (1981) Nitrification potentials in primary and secondary succession. Ecology 62(2):376–386 https://doi.org/10.2307/1936712
Sevink J (1991) Soil development in the coastal dunes and its relation to climate. Landsc Ecol 6(1–2):49–56 https://doi.org/10.1007/BF00157744
Shannon CE, Weaver W (1949) The mathematical theory of communication. University of Illinois Press, Urbana, 125 pp
Smitinand T (2014) Thai plant names dictionary: revised edition 2014. Department of National Parks, Wildlife and Plant Conservation, Bangkok, 806 pp
Sridith K, Laongpol C (2003) The preliminary study on some natural plant communities of the sandbars along eastern coast of peninsular Thailand. Songklanakarin J Sci Technol 25:103–113
Tanaka KD, Denda T, Ueda K, Emura N (2015) Fruit colour conceals endocarp dimorphism from avian seed dispersers in a tropical beach plant, Scaevola taccada (Goodeniaceae), found in Okinawa. J Trop Ecol 31(4):335–344 https://doi.org/10.1017/S0266467415000218
Tanaka N, Sasaki Y, Mowjood MIM, Jinadasa KBSN, Homchuen S (2007) Coastal vegetation structures and their functions in tsunami protection: experience of the recent Indian Ocean tsunami. Landsc Ecol Eng 3(1):33–45 https://doi.org/10.1007/s11355-006-0013-9
Tang T, Zhong Y, Jian S, Shi S (2003) Genetic diversity of Hibiscus tiliaceus (Malvaceae) in China assessed using AFLP markers. Ann Bot-London 92(3):409–414 https://doi.org/10.1093/aob/mcg156
Telewski FW (1995) Wind-induced physiological and developmental responses in trees. In: Coutts MP, Grace J (eds) Wind and trees. Cambridge University Press, Cambridge, pp 237–263
Thomas SC, Martin AR, Mycroft EE (2015) Tropical trees in a wind-exposed island ecosystem: height–diameter allometry and size at onset of maturity. J Ecol 103(3):594–605 https://doi.org/10.1111/1365-2745.12378
Tsoar H (2005) Sand dunes mobility and stability in relation to climate. Physica A 357(1):50–56 https://doi.org/10.1016/j.physa.2005.05.067
Walker LR, del Moral R (2003) Primary succession and ecosystem rehabilitation. Cambridge University Press, Cambridge, 442 pp
Wilson JB, Sykes MT (1999) Is zonation on coastal sand dunes determined primarily by sand burial or by salt spray? A test in New Zealand dunes. Ecol Lett 2(4):233–236 https://doi.org/10.1046/j.1461-0248.1999.00084.x
Wong PP (2005) Coastal environment of Southeast Asia. In: Gupta A (ed) The physical geography of Southeast Asia. Oxford University Press, Oxford, pp 177–192
Yair A, Veste M, Almog R, Breckle SW (2008) Sensitivity of a sandy area to climate change along a rainfall gradient at a desert fringe. In: Breckle SW, Yair A, Veste M (eds) Arid dune ecosystems: the Nizzana sands in the Negev desert. Springer, Berlin/Heidelberg, pp 425–440
Yura H (1997) Comparative ecophysiology of Chrysanthemum pacificum and Solidago altissima L. 1. Why S. altissima cannot be established on the seashore. Ecol Res 12(3):313–323 https://doi.org/10.1007/BF02529461
Zenner EK (2005) Development of tree size distributions in Douglas-fir forests under differing disturbance regimes. Ecol Appl 15(2):701–714 https://doi.org/10.1890/04-0150
Acknowledgments
This research was supported by the Center for Advanced Studies in Tropical Natural Resources (CASTNaR), Kasetsart University, Bangkok, Thailand, and the Kasetsart University Research and Development Institute (KURDI). We would like to thank the Royal Development Projects of His Majesty King Bhumibol Adulyadej for supplying essential facilities during the fieldwork. In addition, the team at the Thai Forest Ecological Research Network (T–FERN) supported field data collection and analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
Rights and permissions
About this article

Cite this article
Marod, D., Sungkaew, S., Mizunaga, H. et al. Woody plant community and distribution in a tropical coastal sand dune in southern Thailand. J Coast Conserv 24, 44 (2020). https://doi.org/10.1007/s11852-020-00761-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11852-020-00761-9