
Abstract
The tomato leafminer, Tuta absoluta (Lepidoptera: Gelechiidae), is a widespread devastating pest reported to develop on economically important solanaceous plants. The characterization of its effective host range could help to understand and prevent the dispersion behavior of the insect in the environment. In this study, the ability of T. absoluta to locate and develop on wild (Solanum nigrum, Atropa belladonna, Datura stramonium) and cultivated (Solanum tuberosum) solanaceous plant species under laboratory conditions was assessed. Dual-choice behavioral assays performed in flying tunnels (S. tuberosum vs. another plant) revealed that adult distribution and female oviposition did not differ between Solanum species, which were preferred to the other tested plants. The volatile molecules released by each tested plant species provide some explanations in the observed behavioral discrimination: S. nigrum and S. tuberosum volatile profiles were similar and were presenting quantitative and qualitative differences with the other tested solanaceous plants. To determine whether the host plant choice was adaptive or not, we have finally conducted fitness assays, by rearing T. absoluta larvae on each plant species and have shown that Solanum species allowed higher larval survivability and lower development time (from egg to adult emergency) compared to the other plants. We conclude that Solanum species are suitable host plants for T. absoluta, but other solanaceous plant species could be opportunistically colonized with fewer incidences.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The tomato leafminer, Tuta absoluta (Lepidoptera: Gelechiidae), is a widespread invasive species originated from South America and damaging economically important tomato crops (Solanum lycopersicum L., Solanaceae) (Desneux et al. 2010). Larvae feed on the mesophyll of all aerial parts of the plants, as well as on the fruits, resulting in significant yield losses, cosmetic damages, and rot diseases to fresh market tomatoes. During the last decade, the pest has spread worldwide and is now a serious threat to commercial tomato production in South and Central America as well as in Europe (Desneux et al. 2010, 2011; Tropea Garzia et al. 2012). Since its introduction in Europe in 2006, the moth is reported on the EPPO A2 list of species recommended for regulation as quarantine pests (EPPO/OEPP 2014).
The emergence of an alien invasive phytophagous insect within a geographical range is determined by its ability to localize and successfully develop on new hosts. Host range is constrained by the behavioral and physiological traits of the insect (Suckling et al. 2014). Although causing large injuries on tomatoes, T. absoluta is considered as a polyphagous leafminer that can also develop on alternative hosts such as cultivated Solanaceae including eggplant (Solanum melongena L.), sweet pepino (Solanum muricatum L.), potato (Solanum tuberosum L.), and tobacco (Nicotiana tabacum L.) (Desneux et al. 2010, 2011). Wild plant species have also been cited to be potential hosts for the development of the pest, such as deadly nightshade (Atropa belladonna L.), Jimson weed (Datura stramonium L.), and black nightshade (Solanum nigrum L.) (Desneux et al. 2010, 2011; Tropea Garzia et al. 2012), that can be found indifferently in croplands, wastelands, logging, and urban areas (Lambinon et al. 2004). In this context, host plant species in the wild could provide reservoirs or refuges for T. absoluta.
Host range prediction is a key step for risk assessment that determines whether an invasive alien species should be eradicated, managed, or tolerated (Suckling et al. 2014). However, little is known about the effective developmental and reproductive capacity of T. absoluta on most of the agricultural and non-cultivated plants that are reported as potential hosts in the literature. The characterization of its host range could help to understand and prevent the dispersion behavior of the insect in the environment for more efficient integrated management strategies.
In the present study, we investigated whether non-cultivated Solanaceae could act as refuges or reservoirs for T. absoluta. Previous works having demonstrated the ability of T. absoluta to efficiently develop and reproduce on S. tuberosum (Caparros Megido et al. 2013), we have first conducted dual-choice bioassays to evaluate the attractiveness of different wild solanaceous plant species. Plant volatile organic compounds (VOCs) were characterized for each tested wild Solanaceae species to provide explanations regarding host location preferences. Finally, the ability of each plant species to sustain T. absoluta larvae was assessed.
Materials and methods
Plant and insect material
Tomato (Solanum lycopersicum cv. Moneymaker), potato (Solanum tuberosum cv. Charlotte), and wild solanaceous plants (Atropa belladonna, Datura stramonium, Solanum nigrum) were cultivated in a greenhouse (25 ± 5 °C) in plastic pots (20 cm diameter × 20 cm height) filled with loam (VP113BIO; Peltracom, Belgium) and grown with a 16L:8D photoperiod. The plants were watered once every 2 days.
In July 2011, 200 third-instar larvae of the tomato leafminer, Tuta absoluta, were collected from a commercial tomato plantation located in Saint-Andiol (France) and were subsequently kept under laboratory conditions at 24 ± 1 °C, 60–70 % RH, and with a 16L:8D photoperiod. T. absoluta colony was maintained on tomato in net cages (46.5 × 46.5 × 46.5 cm). Caterpillars were provided with fresh plants three times a week until pupation.
Behavioral assays
Flying tunnels (232.5 cm × 46.5 cm × 46.5 cm) were used to study the preferences of T. absoluta reared on tomato plants toward different solanaceous plants (Solanum tuberosum cv. Charlotte, versus Atropa belladonna, Datura stramonium or Solanum nigrum; 4 weeks old). The tunnels were divided into three areas (Caparros Megido et al. 2014): a central area for insect release and two areas at opposite sites containing the plants. Twenty newly emerged T. absoluta adults were placed in the neutral area at a distance of 116.5 cm from each plant. After 48 h, the number of males, females, and eggs laid on each plant was counted. Forty-eight hours allow the couples to explore the tunnel and to mate, and also allow the females to lay eggs in sufficient numbers on one of the two plants. Each plant combination was randomly tested five times (for a total of 100 insects tested per dual-choice assay). The experiments were carried out under laboratory conditions (20 ± 1 °C, 65 ± 5 % RH, and a 16L:8D photoperiod under cool white LED lights (77 µmol/sqm/s)). These conditions were monitored using an automatic datalogger (HOBO RH/TEMP 8 K; Onset Computer Corporation, Bourne, MA, USA).
Collection of plant volatiles
Entire 4-week old plants (n = 4 for each species) were separately placed into 40-L glass jars that were closed with a glass lid. Before volatile collection, each glass and Teflon® pipe used to connect the experimental setup was washed with Extran® solution (MA 01; Merck, Darmstadt, Germany). In addition, each plant pot was wrapped in aluminum foil to avoid contamination from the pot and the soil. A pull air pump (Escort Elf® Pump, MSA, Pittsburgh, PA, USA) provided constant airflow from the bottom to the top of the jar at 1 L/min, after being charcoal-filtered with a carbon trap (Carbon Cap 75, Whatman, Maidstone, England). Volatiles were collected from each plant during 24 h, after a 45-min acclimation period. To trap the VOCs released into the plant headspace, a 45 mg HayeSep Q glass trap (80/100 mesh; Hayes Separation Inc., Houston, TX, USA) was connected to the setup with a Teflon® pipe connecting the pump and the glass jar. The VOCs collections were conducted in a laboratory at 20 ± 1 °C, 65 ± 5 % RH, and a 16L:8D photoperiod under cool white light emitting diode (LED) lights (77 µmol/sqm/s). All conditions were monitored using an automatic datalogger (HOBO RH/TEMP 8 K; Onset Computer Corporation, Bourne, MA, USA).
Before use, all traps were washed with 600 µL dichloromethane and 600 µL n-hexane (95 % purity; Sigma-Aldrich, St. Louis, MO, USA). After volatile collection, the traps were immediately eluted with 200 µL n-hexane, and 10 µL of a hexanic solution of n-butyl-benzene (750 µg/mL) was added to each sample as the internal standard. The samples were stored in closed vials wrapped with a Teflon® strip at −80 °C before analysis.
Extracts were analyzed by gas chromatography coupled with mass spectrometry (GC–MS) using an Agilent Technologies (Santa Clara, CA, USA) model 6890n GC System/5973 Mass Selective Detector. Aliquots (1 µL) were injected with a splitless injector maintained at 220 °C. The column (HP-5MS, 30 m × 0.25 mm × 0.25 µm) was maintained at 35 °C for 2 min, before being heated to 100 °C at a constant rate of 5 °C/min. The column was then heated to 260 °C at a constant rate of 20 °C/min. The quantification of compounds was realized by comparing their areas with the internal standard (applying a factor response ratio of 1:1 for each compound), using Chemstation software (Agilent Technologies, Palo Alto, CA, USA). The components were identified by comparing the recorded mass spectra with the National Institute of Standards and Technology (NIST) and Wiley spectral databases. Further identification was carried out by calculating non-isothermal retention indices under identical chromatographic conditions. Identification was finally confirmed by the injection of standards (when commercially available) and comparison of retention times. Volatile chemicals identified at least in two replicates were considered for analytical purposes. Compounds commonly associated with the earth’s atmosphere (e.g., toluene and benzene), as well as compounds associated with the analytical system (e.g., siloxanes or phthalates), were excluded (Jansen et al. 2009).
Evaluation of Tuta absoluta fitness
For each tested plant species, twenty newly emerged T. absoluta adults (ten males and ten females) were placed in a net cage (46.5 × 46.5 × 46.5 cm) in the presence of a single non-flowering host plant (4 weeks old) until the females death. Fifty eggs were then taken from each host plant species. Eggs were individually placed in a Petri dish (5 cm diameter) lined with a moistened filter paper and containing an excised leaf taken from the host plant species on which they were laid. Each Petri dish was considered to be a replicate (i.e., n = 50 for each tested plant species). The emerging larvae were fed with a single piece of leaf (2 × 3 cm), which was changed every day. Water was also added to humidify the filter paper until pupation. Individuals were sexed according to the methods of Coelho and França (1987), and male and female pupae were grouped in couples. Each couple of pupae was placed in an individual Petri dish containing a moistened filter paper and a single corresponding host plant leaf until adult emergence. Following adult emergence, males and females were left together in the Petri dishes for their entire lives. Following the visual observation of the first mating (occurring rapidly after adult emergence), the number of eggs laid on a single plant leaf was evaluated daily until the female died. Water was supplied on the filter paper covering each Petri dish, every 2 days, to optimize adult survival. During the development of the insects, egg-laying date, hatching date, pupation date, adult emergence date, and sex were recorded for each individual. Pupal weight was not recorded, since the ratio between fecundity and pupal weight may not be reliable and should be treated with caution when attempting to understand oviposition (Leather 1988). Experiments were carried out under laboratory conditions (24 ± 1 °C, 65 ± 5 % RH, and 16L:8D photoperiod) and were monitored using an automatic datalogger (HOBO RH/TEMP 8 K; Onset Computer Corporation, Bourne, Massachusetts, USA).
Statistical analyses
Binomial proportion tests (equal distribution hypothesized) were used to compare the number of males, females, and eggs laid on each studied plant observed during the dual-choice behavioral assays conducted in the flying tunnel.
Principal component analysis was performed on a correlation matrix for the visual comparison of the VOC profiles from the different solanaceous plants. One-way ANOVAs were applied to compare the mean quantities of each VOC released by the tested plants.
One-way ANOVAs were applied to compare the mean development times of each life stage of T. absoluta on the various host plant species. When assumptions of the one-way ANOVA (normality of populations and homoscedasticity) were not met, Kruskall–Wallis tests were applied followed by Mann–Whitney comparisons of specific samples pairs. Chi-square tests of independence were applied to compare survival percentages at each life stage and sex ratios on each plant species studied.
All tests were performed using Minitab® version 16 software (http://www.minitab.com/fr-FR/default.aspx).
Results
Behavioral assays
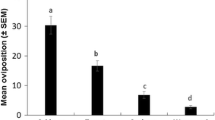
Table 1 lists the total number of males, females, and eggs on each side of the flying tunnels (S. tuberosum versus another plant). Adult males and females attraction did not differ between the two Solanum species (P = 0.701 and P = 0.256), but female oviposition occurred preferentially on S. nigrum (P < 0.001). By contrast, S. tuberosum was preferred to the other tested plants, i.e., Atropa belladonna and Datura stramonium, considering both adult attraction and female oviposition.
Plant volatiles analysis
A total of 34 compounds were found in the headspace of all tested plants (Table 2; Fig. 1). The total amount of VOCs varied among the four tested Solanaceae species, ranging from 5.59 ± 4.93 (S. nigrum) to 50.70 ± 15.07 ng (D. stramonium) per gram of fresh plant (Table 2). Terpenoids (mono- and sesquiterpenes) were the most abundant identified compounds. Monoterpenes represented 32.5, 84.26, 93.14, and 18.19 % of the total VOCs collected, respectively, for S. tuberosum, S. nigrum, A. belladonna, and D. stramonium. Sesquiterpenes represented 66.82, 5.01, 0.00, and 81.81 % of the total emission, respectively, for S. tuberosum, S. nigrum, A. belladonna, and D. stramonium. Related percentages were 0.68, 10.73, 6.86, and 0.00 % for aldehydes.

Chromatograms of VOCs emitted by S. tuberosum, S. nigrum, A. belladonna, and D. stramonium. The number on peaks refers to the compound number given in Table 2
The principal component analysis (Fig. 2) showed that the volatile compound profiles differed between the four Solanaceae species. All tested plants released α-pinene (1), δ-3-carene (5), limonene (7) and nonanal (12) in equivalent amounts. S. tuberosum was the only plant to release β-phellandrene (2), α-cubebene (18), α-copaene (19), β-gurjunene (21), (E)-β-farnesene (25), germacrene-D (28), and bicyclogermacrene (31). In comparison, only S. nigrum released β-pinene (3) and myrcene (4). Only A. belladonna released 1,4-cineole (6), 1,8- cineole (8), α-terpinolene (11), terpinene-1-ol (13), α- terpineol (16), β-terpineol (14), borneol (15), and tridecanal (30). Only D. stramonium released (E)-β-ocimene (9), longifolene (20), valencene (29), α-Muurolene (32), and caryophyllene oxide (35).

Spatial distribution of the volatile blends released by S. tuberosum, S. nigrum, A. belladonna, and D. stramonium (n = 4)
Evaluation of Tuta absoluta fitness
Table 3 lists survivability rates and sex ratios, while Table 4 lists the development times and fecundity rates of T. absoluta reared on the different tested solanaceous host plants.
Mean development times (±SE) of T. absoluta eggs were not statistically different according to the host plants on which they were laid (H 3 = 5.25; P = 0.155). Egg viability differed according to the host plant species (χ 2(3) = 8.15; P = 0.043) and was statistically higher on S. tuberosum (80 %) than on D. stramonium (56 %).
Larval development times differed according to the host plant (H 3 = 45.96; P < 0.001). Larvae fed on S. tuberosum developed faster (8.5 ± 0.2 days) than those fed on S. nigrum (11.7 ± 0.3 days), D. stramonium (11.8 ± 1.3 days), and A. belladonna (13.0 ± 0.5 days). Moreover, differences in terms of larvae survival were observed between the four tested host plants (χ 2(3) = 25.81; P < 0.001). Survivability of larvae fed on S. nigrum (79 %) did not differ from S. tuberosum (67 %), but was higher than on A. belladonna (50 %) and D. stramonium (20 %). Survivability of larvae fed on D. stramonium was significantly lower than those fed on any of the other host plants.
Pupae development times were not statistically different according to the host plants on which pupae were reared (H 3 = 5.78; P = 0.123). However, association tests performed on sample pairs indicated a significant difference between S. nigrum (7.4 ± 0.2 days) and D. stramonium (8.8 ± 0.4 days). There were no significant differences in pupae survival percentages (χ 2(3) = 0.37; P = 0.946), which ranged from 83 % on A. belladonna and D. stramonium to 88 % on S. nigrum.
Development times from egg to adult emergence were significantly lower for insects reared on S. tuberosum (20.4 ± 0.4 days) than those reared on S. nigrum (22.9 ± 0.3 days), A. belladonna (24.7 ± 0.6 days), and D. stramonium (24.4 ± 1.4 days) (H 3 = 33.77; P < 0.001). Survival rates from egg to adult emergence were lower with insects reared on D. stramonium (9 %) than those fed on S. tuberosum (45 %), S. nigrum (45 %), and A. belladonna (39 %).
Adult longevity was not affected by the host plants, ranging from 8.6 ± 1.4 days on D. stramonium to 15.0 ± 1.5 days on A. belladonna (F 3,73 = 2.05; P = 0.114). Similarly, the sex ratio (male/female) (χ 2(3) = 7.45; P = 0.059) and female fecundity rates (H 2 = 1.03; P = 0.598) did not differ significantly between S. tuberosum, S. nigrum, and A. belladonna. Only adult males have emerged from pupae reared on D. stramonium.
Discussion
The insect ability to localize suitable host plants in a given environment determines its propensity to persist and spread to new areas (Suckling et al. 2014). In Europe, the tomato leafminer is abundantly present in the Mediterranean countries, where it mainly infests tomato production sites. But recent studies have suggested that alternative cultivated and non-cultivated host plants could be used by T. absoluta to spread and reproduce in Northern Europe (Desneux et al. 2010, 2011). In the present study, we have shown that T. absoluta females preferentially orientate in dual-choice assays to Solanum species rather than to other wild Solanaceae. However, oviposition has occurred on each of the tested plants, suggesting that all of these plants could be localized and identified as a potential host for the offspring development. During host selection, female insects are exposed to a variety of cues, including plant volatiles, contact chemicals, and visual signals related to plant morphological characteristics, which help to determine the suitability of a host plant (Awmack and Leather 2002; Faria et al. 2008). That T. absoluta females were preferentially attracted by Solanum species (i.e., potato and black nightshade) was expected because these plants are related to the tomato (Lambinon et al. 2004), which is the primary host for the insect development (Desneux et al. 2010, 2011). Male preferences are rather related to the female choice as most of lepidopteran males are primarily attracted by female sex pheromones and rarely exhibit adaptive behavior in response to host plant volatiles (Ramaswamy 1988). All T. absoluta tested individuals were reared on tomato prior to dual-choice assays, and this fact should be underlined because larval experience has been reported to have an impact on the host finding behavior of the subsequent adults (Caparros Megido et al. 2014; Facknath and Wright 2007). The host plant preferences of individuals reared for one or more generations on wild solanaceous host plants have to be described in further studies in order to better understand their dispersion behavior in the environment.
Recent advances in chemical ecology demonstrated that the attraction and oviposition of T. absoluta females are mediated by the volatile signature of their host plant. VOCs released by leaves provide capital information to the females on the suitability of the plant as a larval substrate. It was found that tomato leaf odors mainly include volatile terpenoid compounds which elicit in mated females upwind orientation flight followed by landing as well as egg-laying (Proffit et al. 2011). Tomato volatiles are principally made of monoterpenes including α-pinene, β-pinene, α-phellandrene, β-phellandrene, δ-2-carene, δ-3-carene, limonene, myrcene, and sabinene, as well as the sesquiterpene (E)-β-caryophyllene (Caparros Megido et al. 2014; Proffit et al. 2011). Similar observations were made for potato plants: the volatile emissions of S. tuberosum, principally made of sesquiterpenes as (E)-β-caryophyllene (E)-β-farnesene, and germacrene-D, are implicated in the host selection by T. absoluta females (Caparros Megido et al. 2014). Our results are consistent with these previous headspace analyses regarding the latter species. Despite presenting similarities, each solanaceous plant species released a qualitatively and quantitatively specific volatile blend, as visually demonstrated by the principal component analysis. The fact that the insect has laid eggs on each plant suggests that one or more combinations of volatile compounds are responsible for the attraction and oviposition behavior, some of which being more attractive than others. By example, even both are principally made of monoterpenes, A. belladonna emissions differed from those of S. nigrum which are mainly constituted with tomato-related compounds such as α-pinene, β-pinene, δ-3-carene, limonene and myrcene. The total quantity of terpenoids is likely not affecting host selection, as the total amount of monoterpenes and sesquiterpenes released by the four Solanaceae is not correlated with the attraction results and the number of eggs laid. Rather than the total quantity of VOCs released by a plant, the presence of specific blend of certain chemicals can be involved in the selection of a host and the quantity of eggs deposited. Further electrophysiological and behavioral assays conducted on natural blends as well as on separated identified chemicals are required to confirm the effect of specific chemicals on the choice of the oviposition site in T. absoluta.
Oviposition on a suitable host in response to plant cues is of major importance for an insect to maximize its fitness. A significant variability in egg survivability was observed among the tested plants. This discordance may be due to either morphological characteristics of the plants or defense mechanisms in response to egg presence, impacting embryonic development. By example, low humidity on the leaf surface due to stomatal closure or hypersensitive response (i.e., formation of necrotic tissue at the site of egg deposition) may lead to their desiccation (Hilker and Meiners 2011; Woods 2010). Volatile and contact chemicals from plant tissues could also alter the embryonic development (Hilker and Meiners 2011). Another hypothesis could be that females adjust the nutritional content of their eggs in response to a poor quality host plant, thus allocating fewer resources to embryos, which has an evident impact on the performances of the resulting offspring (Boggs 1992). By example, Lymantria dispar L. (Lepidoptera: Lymantriidae) was reported to lay eggs on Populus tremuloides Michx. (referred as a good-quality host plant) that contained the greatest amount of vitellogenin compared with Quercus rubra L., Q. prinus L., and Pinus rigida Mill. (referred as lower-quality host plant species) (Rossiter et al. 1993).
Considering herbivorous insects, short development times, high survival rates, and high fecundity rates (number of eggs per female) are related to a suitable food quality (Awmack and Leather 2002; Greenberg et al. 2002; Pereyra and Sánchez 2006). In the present study, the host plant affected the development times of T. absoluta from egg to adult emergence: individuals reared on S. tuberosum grew faster than the others. Corresponding survival rates were only found to be statistically lower on D. stramonium, although those observed on S. tuberosum and S. nigrum appeared to be numerically equal to each other and higher than on A. belladonna. In the absence of abiotic and biotic stresses, survival rates are mainly influenced by the feeding stage (larval instar), suggesting differences in nutritive quality and/or production of plant metabolites that have impaired larval development. By example, A. belladonna and D. stramonium tissues contain tropane alkaloids (Krenzelok 2010; Pomilio et al. 2008) involved in insect herbivory resistance (Arab et al. 2012; Shonle and Bergelson 2000). Adult longevity and fecundity rates were similar between the four plants, while no female adults were obtained on D. stramonium. In terms of T. absoluta fitness, it can be assumed that solanaceous plants are not of equal quality for the development of the insect pest. S. tuberosum appears to be the most suitable host among the tested plants considering the biological parameters described above, but S. nigrum, A. belladonna, and D. stramonium can also be exploited as food source with a decreased success in their respective order of citation.
Surprisingly, longer development times (28 days) for similar survivability rates (47 %) were previously observed using an identical protocol for T. absoluta reared on its primary host plant, Solanum lycopersicum (Caparros Megido et al. 2013). However, adult fecundity rate reported on S. lycopersicum (50 eggs per female) was twofold higher than on S. tuberosum and S. nigrum (which allowed the largest offspring in our study). Therefore, it can be assumed that this decrease in oviposition could be directly related to a lower reproductive capacity of the adults resulting from our bioassays, making tomato more beneficial in terms of fitness. Aside this decrease in performances, it is not excluded that a female strategic choice also has occurred in response to a host plant with a lower quality, strengthening the observed egg lay depression. According to this hypothesis, females could have delayed their oviposition, or resorbed eggs and reallocated the nutriments in order to extend their own longevity, waiting to find a more suitable host plant where the offspring performances will be optimized (Awmack and Leather 2002). High adult longevity, which is classically related to a high fecundity (Leather 1988) and low fecundity rates observed with insects reared on A. belladonna, complies with this expectation.
In our experimental design, insect performances were assessed during a single generation, starting with egg-laying adults taken from their primary host plant. The potential effect of food reserves allocated to the eggs on the biological traits of the immature and mature instars should be considered. We might expect that generation time, survivability, and female fecundity will be impacted after several generations on an alternative host plant. Compared to previous results (Caparros Megido et al. 2013), egg to adult development time on S. tuberosum cv. Charlotte increases from 20 to 24 days after five generations, being less beneficial in terms of fitness. Additional experiments are needed to assess the impact on insect performances of a long-term infestation on wild solanaceous host plants as reservoirs.
Finally, genetic variability in plants and insects may have a great impact, respectively, on VOCs emitted or perceived (Harrewijn et al. 1995), and insecticidal compounds secretion or plant defense avoidance (Moore et al. 2014). Climatic variations (Agosta 2006), phenological and physiological state of the plants (Ayabe et al. 2014), local selective pressures on plant traits (Castillo et al. 2014), or the presence of plant endophytic symbionts and insect pathogens (Crawford et al. 2010; Gurulingappa et al. 2010) may also have an influence on host plant availability for the insects. Such variability in plant resource may have a significant impact on the spatiotemporal patterns of plant selection for female oviposition.
References
Adams RP (2007) Identification of essential oil components by gas chromatography/mass spectrometry. Allured Publishing Corporation, Illinois
Agosta SJ (2006) On ecological fitting, plant-insect associations, herbivore host shifts, and host plant selection. Oikos 114:556–565
Arab A, Alves MN, Sartoratto A, Ogasawara DC, Trigo JR (2012) Methyl jasmonate increases the tropane alkaloid scopolamine and reduces natural herbivory in Brugmansia suaveolens: is scopolamine responsible for plant resistance? Neotrop Entomol. doi:10.1007/s13744-011-0001-0
Awmack CS, Leather SR (2002) Host plant quality and fecundity in herbivorous insects. Annu Rev Entomol 47:817–844
Ayabe Y, Minoura T, Hijii N (2014) Plasticity in resource use by the leafminer moth Phyllocnistis sp. in response to variations in host plant resources over space and time. J For Res. doi:10.1007/s10310-014-0467-9
Boggs CL (1992) Resource allocation: exploring connections between foraging and life history. Funct Ecol 6:508–518
Caparros Megido R, Brostaux Y, Haubruge E, Verheggen FJ (2013) Propensity of the Tomato Leafminer, Tuta absoluta (Lepidoptera: Gelechiidae), to develop on four potato plant varieties. Am J Potato Res 90:255–260
Caparros Megido R, De Backer L, Ettaïb R, Brostaux Y, Fauconnier M-L, Delaplace P, Lognay G, Belkadhi MS, Haubruge E, Francis F, Verheggen FJ (2014) Role of larval host plant experience and solanaceous plant volatile emissions in Tuta absoluta (Lepidoptera: Gelechiidae) host finding behavior. Arthropod Plant Interact 8:293–304
Castillo G, Cruz LL, Tapia-Lopez R, Olmedo-Vicente E, Carmona D, Anaya-Lang AL, Fornoni J, Andraca-Gomez G, Valverde PL, Nunez-Farfan J (2014) Selection mosaic exerted by specialist and generalist herbivores on chemical and physical defense of Datura stramonium. PLoS One 9:e102478. doi:10.1371/journal.pone.0102478
ChemSpider (2014) http://www.chemspider.com. Accessed 1 Dec 2014
Coelho MCF, França FH (1987) Biologia e quetotaxia da larva e descrição da pupa e adulto da traça do tomateiro. Pesqui Agropecu Bras 22:129–135
Crawford KM, Land JM, Rudgers JA (2010) Fungal endophytes of native grasses decrease insect herbivore preference and performance. Oecologia 164:431–444
Desneux N, Wajnberg E, Wyckhuys KAG, Burgio G, Arpaia S, Narvaez-Vasquez CA, Gonzalez-Cabrera J, Catalan Ruescas DC, Tabone E, Frandon J, Pizzol J, Poncet C, Cabello T, Urbaneja A (2010) Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J Pest Sci 83:197–215
Desneux N, Luna MG, Guillemaud T, Urbaneja A (2011) The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. J Pest Sci 84:403–408
EPPO/OEPP (2014) EPPO A1 and A2 list of pests recommended for regulation as quarantine pests. http://www.eppo.int/QUARANTINE/listA2.htm. Accessed 1 Dec 2014
Facknath S, Wright DJ (2007) Is host selection in leafminer adults influenced by pre-imaginal or early adult experience? J Appl Entomol 131:505–512
Faria CA, Torres JB, Fernandes AMV, Farias AMI (2008) Parasitism of Tuta absoluta in tomato plants by Trichogramma pretiosum Riley in response to host density and plant structures. Cienc Rural 38:1504–1509
Ferraz EO, Bertolucci SKV, Pinto JEBP, Braga AF, Costa AG (2014) Organic systems in the growth and essential-oil production of the yarrow. Rev Ciênc Agron 45:111–119
Flavornet (2014) http://www.flavornet.org/flavornet.html. Accessed 1 Dec 2014
Gbolade AA, Dzamic AM, Ristic MS, Marin PD (2009) Erratum: essential oil composition of centratherum punctatum from Nigeria (Chem Nat Compd (2009) 45:118–119). Chem Nat Compd 45:296
Greenberg SM, Sappington TW, Sétamou M, Liu TX (2002) Beet armyworm (Lepidoptera: Noctuidae) host plant preferences for oviposition. Environ Entomol 31:142–148
Gurulingappa P, Sword GA, Murdoch G, McGee PA (2010) Colonization of crop plants by fungal entomopathogens and their effects on two insect pests when in planta. Biol Control 55:34–41
Harrewijn P, Minks AK, Mollema C (1995) Evolution of plant volatile production in insect–plant relationships. Chemoecology 5(6):55–73
Hilker M, Meiners T (2011) Plants and insect eggs: how do they affect each other? Phytochemistry 72:1612–1623
Jansen RMC, Hofstee JW, Wildt J, Verstappen FWA, Bouwmeester HJ, Posthumus MA, van Henten EJ (2009) Health monitoring of plants by their emitted volatiles: trichome damage and cell membrane damage are detectable at greenhouse scale. Ann Appl Biol 154:441–452
Krenzelok EP (2010) Aspects of Datura poisoning and treatment. Clin Toxicol 48:104–110
Lambinon J, Delvosalle L, Duvigneaud J (2004) Nouvelle flore de la Belgique, du G.-D. de Luxembourg, du nord de la France et des régions voisines. Edition du Jardin botanique national de Belgique, Meise
Leather SR (1988) Size, reproductive potential and fecundity in insects: things aren’t as simple as they seem. Oikos 51:386–389
Maggio A, Rosselli S, Bruno M, Spadaro V, Raimondo FM, Senatore F (2012) Chemical composition of essential oil from Italian populations of Artemisia alba Turra (Asteraceae). Molecules 17:10232–10241. doi:10.3390/molecules170910232
Moore BD, Andrew RL, Külheim C, Foley WJ (2014) Explaining intraspecific diversity in plant secondary metabolites in an ecological context. N Phytol 201:733–750
Pereyra PC, Sánchez NE (2006) Effect of two solanaceous plants on developmental and population parameters of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotrop Entomol 35:671–676
Pherobase (2014) Database of pheromones and semiochemicals. http://pherobase.com. Accessed 1 Dec 2014
Pomilio AB, Falzoni EM, Vitale AA (2008) Toxic chemical compounds of the Solanaceae. Nat Prod Commun 3:593–628
Proffit M, Birgersson G, Bengtsson M, Reis R Jr, Witzgall P, Lima E (2011) Attraction and oviposition of Tuta absoluta females in response to tomato leaf volatiles. J Chem Ecol 37:565–574
Ramaswamy SB (1988) Host finding by moths: sensory modalities and behaviours. J Insect Physiol 34:235–249
Rossiter MC, Cox-Foster DL, Briggs MA (1993) Initiation of maternal effects in Lymantria dispar: genetic and ecological components of egg provisioning. J Evol Biol 6:577–590
Shonle I, Bergelson J (2000) Evolutionary ecology of the tropane alkaloids of Datura stramonium L. (Solanaceae). Evolution 54:778–788
Suckling DM, Charles JG, Kay MK, Kean JM, Burnip GM, Chhagan A, Noble A, Barrington AM (2014) Host range testing for risk assessment of a sexually dimorphic polyphagous invader, painted apple moth. Agric For Entomol 16:1–13
Tropea Garzia G, Siscaro G, Biondi A, Zappalà L (2012) Tuta absoluta, an exotic invasive pest from South America now in the EPPO region: biology, distribution and damage. EPPO Bull 42:205–210
Woods HA (2010) Water loss and gas exchange by eggs of Manduca sexta: trading off costs and benefits. J Insect Physiol 56:480–487
Acknowledgments
This research was funded by the Service Public de Wallonie (SPW–DGO3, D31-1298). The authors thank Alain Benko (Biologie végétale, Université de Liège) for plant choice and identification, and Danny Trisman (Chimie générale et organique, Université de Liège) for technical support with GC–MS analyses. Emilie Joie and Maud Fagan (Entomologie fonctionnelle et évolutive, Université de Liège) are acknowledged for their technical support in plant culture and insect rearing.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Stanislav Gorb.
Rights and permissions
About this article

Cite this article
Bawin, T., Dujeu, D., De Backer, L. et al. Could alternative solanaceous hosts act as refuges for the tomato leafminer, Tuta absoluta?. Arthropod-Plant Interactions 9, 425–435 (2015). https://doi.org/10.1007/s11829-015-9383-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-015-9383-y