
Abstract
We investigated the female oviposition preference and larval performance and behavior of Trichoplusia ni (Hubner) on cabbage, tomato, soybean (host plants), and wormseed (nonhost plant) in laboratory experiments. In addition, we investigated the role of trichomes in the oviposition of females. Females oviposited more often on cabbage than on tomato, soybean, or wormseed plants. Tomato was the second most preferred plant, followed by soybean and wormseed. Neonate larvae gained more weight on cabbage and tomato than on soybean or wormseed, while second-instar larvae grew better on cabbage than on tomato, soybean, or wormseed. The least growth of neonate larvae occurred on wormseed plants. The orientation of neonate and second-instar larvae to cabbage, tomato, soybean, and wormseed did not differ significantly. Neonate larvae settled equally on leaf discs of cabbage, tomato, soybean, and wormseed, while most second-instar larvae settled on leaf discs of cabbage in comparison with the other plants after 24 h of release. The foliar area consumed by neonate larvae was quite similar among plants, but second-instar larvae consumed more cabbage than tomato, soybean, or wormseed. Comparing different types of leaves, females oviposited more often on mature than young leaves of tomato, soybean, and wormseed. In contrast, females did not show any preference for ovipositing on young or mature leaves of cabbage. In general, we found that the density or length of nonglandular and glandular trichomes of tomato, soybean, and wormseed plants negatively affected oviposition of T. ni females.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Host location plays an integral role in the fitness of all phytophagous insects; for instance, the oviposition site chosen by females is crucial for the survival of their offspring (Stephens and Krebs 1986; Thompson and Pellmyr 1991). Thus, it would be expected that females would oviposit on suitable hosts for development of their offspring. This idea was formalized as the preference–performance hypothesis, which states that herbivore females will choose to oviposit on hosts on which their offspring will have better performance (Levins and MacArthur 1969; Jaenike 1978). However, in some cases, there is no correlation or the relationship is weak between female preference and offspring performance (Rausher 1979; Berdegué et al. 1998; Refsnider and Janzen 2010), and several life history and ecological factors may account for this discrepancy, including diet breadth, aggregation of offspring, and presence of natural enemies, among others (Thompson 1988; Gripenberg et al. 2010). For example, a meta-analysis showed that the relationship between female preference and larval performance was stronger for oligophagous species than for polyphagous insects (Gripenberg et al. 2010). Laboratory and field experiments showed that Spodoptera littoralis (Boisduval) females prefer to oviposit on alfalfa than cotton plants, but larvae performed better on the latter host. However, eggs oviposited on cotton were more parasitized by Chelonus inanitus (L.) than those laid on alfalfa (Sadek et al. 2010), suggesting that females prefer to oviposit on an inferior larval host plant since it provides higher protection from natural enemy.
Herbivorous insects must overcome a number of hurdles to use host plants, including nutrition, attachment, desiccation, and plant defenses; for example, insects have to overcome the constitutive defense represented by trichomes. Depending on their size and density, trichomes may form structural barriers that affect the movement or ability of insects to oviposit or feed on host plants (Simmons et al. 2005; Simmons and Gurr 2005). Furthermore, glandular trichomes produce sticky exudates, proteins, terpenoids, or toxins that may affect the behavior and physiology of herbivorous insects (Wagner et al. 2004; Lange 2015).
The cabbage looper moth, Trichoplusia ni (Hübner), is a generalist moth, whose larvae feed on more than 150 plant species from 36 families (Sutherland and Greene 1984). However, larvae do not develop or survive equally on all host plants (Soo Hoo et al. 1984). Two previous studies have investigated the relationship between female preference and larval growth in the cabbage looper moth, with contrasting results, because only on some plants was a match found between preference and performance (Cameron et al. 2007; Shikano et al. 2010). Could such results be due to the fact that both studies used cut leaves instead of whole plants? It is well documented that mechanical damage may activate plant defenses, thus influencing the behavior and physiology of insects (Haukioja and Niemelä 1977; Reymond et al. 2000; Savatin et al. 2014); for instance, leaf damage to cotton and cabbage plants positively and negatively affects the attraction and oviposition of T. ni females on these plant species (Landolt 1993).
In this study, we investigated the female oviposition preference and larval performance of T. ni on cabbage, tomato, soybean (host plants), and wormseed (a possible nonhost plant) using whole plants. We also evaluated the choice (orientation and settling) and movement (arrest and dispersal) of neonate and second-instar larvae on the experimental plants. In addition, we investigated the role of trichomes in oviposition behavior of T. ni females.
Materials and methods
Biological material
T. ni larvae were reared on artificial diet (Rojas et al. 2003) under controlled conditions of temperature (26 ± 2 °C), relative humidity (70 ± 5 %), and photoperiod (14:10 h, L:D). The colony has been maintained in the laboratory for more than 50 generations, although periodic additions of wild insects have been performed. Naïve neonate larvae were used 2–3 h after hatching. They did not contact plant material or artificial diet, while second-instar larvae (10 days old) were fed artificial diet. To ensure mating, groups of 10 females and 15 males were kept together in cages (30 cm × 30 cm × 30 cm) from the first night of emergence to obtain mated females. Moths were fed with 10 % sucrose solution dispensed on cotton wool in a Petri dish. After the assays, 30 % of females were dissected to determine their mating status by evaluating the presence of spermatophores in the bursa copulatrix. Five-day-old naïve females were used in all experiments.
Host plants used were cabbage, Brassica oleracea L. var. capitata (Cruciferae), unknown cultivar; tomato, Solanum lycopersicum Mill. (Solanaceae), unknown cultivar; and soybean, Glycine max (L.) c.v. cristalina (Fabaceae); while wormseed, Chenopodium ambrosioides (L.) var. blanco (Chenopodiaceae), was used as nonhost plant. Seeds were sown individually in sterile soil in plastic containers (12 cm high × 10 cm diameter), which were placed under natural conditions of light (12 h), temperature of 27 ± 3 °C, and relative humidity of 65 ± 5 %. Containers were placed inside screen cages (4 m × 4 m) to protect plants from insect infestation. Plants were fertilized with N-P-K (17-17-17) fertilizer once or twice before use in experiments. Plants of different ages were used to aim for a total plant height as similar as possible between species. Thus, we used 90-day-old cabbage plants, 60-day-old tomato and wormseed, and 20-day-old soybean.
Female oviposition preference
The female preference for cabbage, tomato, soybean, or wormseed was evaluated in multiple-choice tests. In each assay, one plant of each species were put together in pyramidal screen cages (75 cm × 60 cm × 60 cm), which were placed outdoors under a covered area with circulating air under natural conditions of temperature and relative humidity. The position of each plant was randomly determined in each cage. The cages were separated from each other by 1 m, and the distance between plants was 30 cm. Trials began at 19:00 h, considering the nocturnal activity of T. ni. In each trial, a gravid moth was released on a platform 15 cm high, placed at the entrance of the cage. Plants and insects were used once. Oviposition was recorded 12 h later. A total of 15 replicates (cages) were performed. We took photographs of the plants at the same distance (60 cm) and zoom using a Sony DCR-DVD508 camera (Sony Mexico) to calculate the leaf area of each plant species. In total, we processed 15 images per plant species using Adobe PhotoShop CS5 software to convert pixels to cm2.
Larval performance
In this experiment, we evaluated the growth performance of neonate and second-instar larvae on cabbage, tomato, soybean, or wormseed. Using a paintbrush, an individual larva was placed onto a potted plant. Larvae were weighted before release. Potted plants were covered with a 3-l polyethylene terephthalate (PET) bottle to avoid larvae leaving the plant. Infested plants were maintained at 25–27 °C and relative humidity of 55–65%. Larvae were collected 10 and 5 days after release, respectively, and at weighted to record final weight. Mortality was also calculated. A total of 15 larvae per host for each stage were used.
Larval orientation and settling
Orientation and settlement of neonate and second-instar larvae on cabbage, tomato, soybean, or wormseed were investigated in multiple-choice tests. The arena consisted of a Petri dish (15 cm diameter × 1.5 cm height). Leaf discs of 2 cm diameter of each plant species were placed 2 cm from each other inside the arena with random distribution, 0.5 cm from the Petri dish’s wall. The leaf discs were made from mature leaves. The assays were performed under the same conditions as the female oviposition experiment. An individual larva was released in the center of each arena and its position recorded at 1 and 24 h to determine orientation and settling preference (Mutyambai et al. 2014), respectively. The Petri dish was covered with its lid and placed in a dark room with red light. At the end of the assay, the remaining parts of the leaf discs were recovered and images digitalized using a scanner (Epson) with optical resolution of 1200 DPI. The foliar area consumed by larvae was converted from pixels to cm2 using PhotoShop CS5 software. A total of 20 replicates were performed.
Arrest and dispersal of larvae
In this experiment, we evaluated arrest and dispersal of neonate and second-instar larvae in multiple-choice tests. The arena consisted of a Petri dish (15 cm diameter × 1.5 cm height). A leaf disc (2 cm diameter) of a given plant was placed at the center of the arena, and three leaf discs of the other plants were placed 2 cm away from the central leaf disc such that each of them lay at one of the vertices of an imaginary triangle, with random distribution. The leaf discs were made from mature leaves. Using a paintbrush, a larva was deposited on the central leaf disc. The position of larvae was recorded at 1 and 24 h. The assays were conducted under the conditions described in the section on larval orientation and settling. At the end of the assay, the foliar area consumed by each larva was calculated as described above. This experiment was replicated 20 times per category of leaf disc and per kind of larvae.
Role of trichomes in female oviposition
In this experiment, we investigated the role of trichomes in the oviposition of T. ni females. First, the oviposition of females on cabbage, tomato, soybean, or wormseed plants was investigated in nonchoice tests. A plant of a given species was placed at the center of a pyramidal screen cage (75 cm × 60 cm × 60 cm). Then, an individual gravid female was released into the cage. After 12 h, the number of eggs deposited on young and mature leaves was recorded. In addition, the density and type of trichomes in 1 cm2 of one young and one mature leaf of the same plant were recorded under a binocular microscope. Trichome type classification was performed for tomato (Kennedy 2003; Glas et al. 2012; Simmons and Gurr 2005), soybean (Permana et al. 2012; Franceschi and Giaquinta 1983), and wormseed (Bonzani et al. 2003; Sá et al. 2016). In addition, length of trichomes was measured using AxioVision software version 4.7.1 (Imaging Associates Ltd., Bicester, UK). In total, 15 young and 15 mature leaves per plant species were analyzed.
Statistical analysis
Statistical analyses were performed using the R statistical software package (R Development Core Team 2014). Female oviposition preference was analyzed by analysis of covariance (ANCOVA), with plant type as independent variable and foliar area of plants as covariable. Larval performance and the effect of trichome density or length on the number of eggs oviposited were analyzed by one-way analysis of variance (ANOVA). Significant ANOVAs were followed by honest significant difference (HSD) test for multiple comparison of means. Larval preference was investigated using log-likelihood ratio contingency table analysis.
Results
Female oviposition preference
Oviposition depended on plant species (F = 52.61; df = 3, 76; P = 0.001) but not on the foliar area of plants (F = 0.07; df = 1, 75; P = 0.78). Females oviposited more often on cabbage than on tomato, soybean, or wormseed plants. Tomato was the second preferred host plant, followed by soybean and wormseed (Fig. 1).

Mean (+standard error of the mean, SEM) number of eggs laid by Trichoplusia ni females on host (cabbage, tomato, and soybean) and nonhost (wormseed) plants in multiple-choice tests. Different letters on bars indicate significant difference (Tukey test, α = 0.05)
Larval performance
Neonate (F = 32.6; df = 3, 47; P < 0.001) and second-instar larvae (F = 35.7; df = 3, 70; P < 0.001) grew differently on cabbage, tomato, soybean, and wormseed plants. Neonate larvae grew better on cabbage and tomato than on soybean or wormseed, with the least growth of neonate larvae occurring on wormseed. Older larvae grew better on cabbage than on tomato, soybean, or wormseed (Fig. 2). No mortality of neonate larvae was observed on cabbage or tomato, but 20 and 35 % mortality occurred on soybean and wormseed, respectively. With second-instar larvae, no mortality was observed.

Mean (±SEM) weight (mg) of Trichoplusia ni larvae after 10 (neonate) and 5 days (second instar) on cabbage, tomato, soybean, and wormseed plants. Different letters on bars indicate significant difference (Tukey test, α = 0.05)
Larval orientation and settling
The orientation of neonate (χ 2 = 4.4, df = 3, P = 0.22) and second-instar larvae (χ 2 = 2.8, df = 3, P = 0.42) to cabbage, tomato, soybean, and wormseed did not differ significantly between plant species (Fig. 3a). Neonate larvae settled equally on leaf discs of cabbage, tomato, soybean, and wormseed (χ 2 = 6.8, df = 3, P = 0.07), and there was little movement between the first hour (orientation) and 24 h (settling) (Fig. 3a, b). The foliar area consumed by neonate larvae was quite similar among cabbage, soybean, and wormseed, but decreased in tomato (Table 1a). For second-instar larvae, we found that they settled more on leaf discs of cabbage in comparison with tomato, soybean, or wormseed after 24 h of release (χ 2 = 45.2, df = 3, P < 0.001) (Fig. 3b), coinciding with the larger area of cabbage leaves consumed by larvae (64 %) compared with tomato (3 %), soybean (1 %), or wormseed (2 %) (F = 159; df = 3, 16; P < 0.001) (Table 1a).

Proportion of Trichoplusia ni larvae occurring on leaf discs of cabbage, tomato, soybean, or wormseed in multiple-choice tests: a orientation and b settling
Arrest and dispersal
Most of the neonate larvae remained on the plant where they were placed at 1 and 24 h after release (Table 2). Neonate larvae only fed on the plant where they were initially deposited (Table 1b). In contrast, second-instar larvae showed more movement among plant disks (Table 2); for example, 1 h after larvae were deposited on cabbage, only 67 % remained on this plant species, while 20 and 13 % were found on soybean and tomato, respectively. However, all larvae were found on cabbage after 24 h. In the same way, less than 50 % of larvae deposited on tomato, soybean, and wormseed remained on these plants at 1 h after release, although most of these larvae were found on cabbage 24 h after release, suggesting that few larvae were arrested by leaf discs of tomato, soybean, or wormseed (Table 2). Second-instar larvae consumed more cabbage, followed by tomato, soybean, and wormseed (F = 20.36; df = 3, 56; P < 0.001) (Table 1b). In some cases, although larvae were not found on a determined plant species at 1 or 24 h after release, some consumption occurred on that plant species, implying that there was considerable movement of larvae among different plant species within the 24-h period.
Role of trichomes in female oviposition
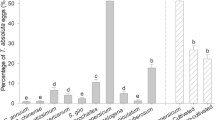
Females preferred to oviposit on the abaxial surface (underside) of young and mature leaves compared with the adaxial (upper) side of young leaves of cabbage. Oviposition on the adaxial surface of mature leaves was intermediate and not significantly different from that occurring on the abaxial surface of young and mature leaves, or the adaxial surface of young leaves (Fig. 4). Cabbage leaves are not pubescent but have abundant waxes on both leaf surfaces. On tomato, females oviposited more often on the abaxial surface of mature leaves than on the abaxial surface of young leaves, or the adaxial surface of mature and young leaves (Fig. 4). Tomato plants have three types of trichomes, corresponding to two glandular types I and VI, and nonglandular type V. There is significant difference between the density of trichomes and surface of mature and young leaves. The abaxial surface of mature leaves, the most preferred surface for oviposition, had less density of type V trichomes (F = 44.79; df = 3, 53; P < 0.001), and the abaxial surface of young leaves had lesser density of type VI trichomes than the adaxial surface of young and mature leaves (F = 6.74; df = 3, 53; P < 0.001) (Table 3). We found the least number of eggs when the density (F = 18.01; df = 2, 57; P < 0.001) or length (F = 14.40; df = 2, 57; P < 0.001) of type VI trichomes was higher. Regarding trichome type I, we found that their density (F = 5.81; df = 2, 57; P = 0.019) but not length (F = 0.66; df = 2, 57; P > 0.05) had a significant effect on oviposition. The density (F = 1.98; df = 2, 57; P > 0.05) or length (F = 0.56; df = 2, 57; P > 0.05) of type V trichomes did not affect the oviposition of T. ni females. On soybean, T. ni females preferred to oviposit on the abaxial surface of mature leaves in comparison with the abaxial surface of young leaves, or adaxial surface of mature and young leaves (Fig. 4). This plant species has one type of trichome, viz. a glandular type, with significant difference between the density and surface of mature and young leaves (F = 695.4; df = 3, 76; P < 0.001; Table 3). We found the least number of eggs when the density (F = 5.57; df = 2, 57; P = 0.022) or length (F = 24.19; df = 2, 57; P < 0.001) of trichomes was highest. On wormseed, females oviposited more often on the adaxial surface of mature leaves than on the adaxial surface of young leaves, or the abaxial surface of mature and young leaves (Fig. 4). This plant species has three types of trichomes, viz. nonglandular (flagelliform), noncapitate glandular, and capitate glandular. The number of noncapitate (F = 7.27; df = 3, 56; P < 0.001) and capitate glandular (F = 187.8; df = 3, 56; P < 0.001) trichomes were reduced on the adaxial surface of mature leaves, while the nonglandular type was absent from the adaxial surface (Table 3). Higher density (F = 42. 06; df = 2, 57; P < 0.001) or larger size (F = 42.14; df = 2, 57; P < 0.001) of nonglandular trichomes negatively affected the oviposition of T. ni females. The length (F = 18.87; df = 2, 57; P < 0.001) of noncapitate glandular trichomes but not their density (F = 1.35; df = 2, 57; P = 0.250) affected the oviposition of females, which oviposited less on leaf surfaces with larger trichomes. Finally, the density (F = 1.64; df = 2, 57; P = 0.204) or length (F = 0.02; df = 2, 57; P = 0.898) of capitate glandular trichomes did not affect the oviposition of T. ni females.

Mean (±SEM) number of eggs laid by T. ni females on young and mature leaves of cabbage, tomato, soybean, and wormseed. YAB, young abaxial surface; YAD, young adaxial surface; MAB, mature abaxial surface; MAD, mature adaxial surface. Bars within each plant with similar letters are not significantly different (Tukey test, α = 0.05)
Discussion
We found a partial match between female oviposition preference and larval performance of T. ni, with females preferring to oviposit on cabbage and offspring performing better on this host plant than on the others. Likewise, females preferred to oviposit on tomato in comparison with soybean or wormseed, and neonate larvae performed better on tomato than on soybean or wormseed. However, second-instar larvae showed similar performance on the three plant species. Females also preferred to oviposit on soybean over wormseed, but second-instar larvae preformed similarly on both plants. Such inconsistency between female oviposition preference and larval performance has been noted in two previous studies (Cameron et al. 2007; Shikano et al. 2010). In general, Cameron et al. (2007) found that female oviposition preference and larval performance of T. ni using broccoli (Brassica oleracea L.) and eight agricultural weeds correlated well. However, they reported that females preferred to oviposit on stinkweed (Thlaspi arvense L.) rather than on broccoli, but only one larva survived for 7 days on stinkweed. Shikano et al. (2010) investigated the relationship between female host plant selection and larval performance using cabbage (B. oleracea L.), pak choi (Brassica rapa L.), bean (Phaseolus vulgaris L.), tomato (S.lycopersicum), anise hyssop [Agastache foeniculum (Pursh) Kuntze)], and geranium hybrid (Perlargonium × hortum). They found a correlation between female oviposition preference and larval performance, except for on anise hyssop. They also reported that females oviposited a higher number of eggs on geranium, despite larva mortality on this plant species. They proposed that the inconsistency in their results could be due to several other factors, including possible confusion of females due to volatiles from the experimental plants being mixed inside the arena, that plants had similar leaf shape, or neuronal constraints of females. In this study, we cannot rule out the first two possibilities, but they seem unlikely because bioassays were performed in ventilated cages and plants were sufficiently spaced to minimize mixing of volatiles from different plants during assays. Although the leaf shapes of the experimental plants are different, in complete darkness these could only be detected as shadows. Thus, the third possibility seems to be the likely explanation for the “mistakes” of females during host plant choice. It has been suggested that generalist herbivorous insects, such as T. ni, have limited capacity for processing sensorial information, making them less efficient in discriminating between host plants (Bernays 2001). Recently, we found that T. ni females were able to discriminate between herbivore-damaged soybean plants and undamaged plants (Coapio et al. 2016). However, it is possible that discriminating plants of different species is more difficult for T. ni females. In fact, in the present study, T. ni females oviposited on wormseed, a nonhost plant. Other factors that may affect the correlation between female oviposition preference and larval performance are limited offspring mobility, presence of competitors and natural enemies, and whether females have the potential to feed as adults (Gripenberg et al. 2010).
The fact that neonate larvae have limited mobility suggests that females have the task of selecting appropriate plants for their offspring. However, if females make a “mistake” and oviposit on a nonhost plant but their offspring are capable of surviving on this plant, eventually this nonhost plant could become a host plant. In our case, T. ni females oviposited on wormseed but no neonate larvae survived after 20 days, although some second-instar larvae survived on this plant, reaching adult stage (Coapio unpublished data). This suggests that wormseed may be considered a suboptimal host for neonates but suitable host for second-instar larvae (Knolhoff and Heckel 2014). Previously, it was reported that T. ni larval experience of nonhost plant extract increased preference for that plant as an oviposition substrate (Chow et al. 2005). However, this is possible only if neonate larvae survive and develop to adult stage. Alternatively, second-instar or older larvae may leave the natal plant and look for food themselves. Our results show that second-instar larvae moved between leaf disks and ultimate chose cabbage for feeding, independent of where they were initially placed.
Interestingly, females and larvae preferred cabbage, and larvae developed better on this plant; however, it appears to be nutritionally inferior compared with tomato, soybean, or wormseed. For example, cabbage has lower protein content and energetic value than the other plants used in this study (Coapio unpublished data). One possibility is that tomato, soybean, and wormseed have defenses to which T. ni is less adapted, and therefore this insect does not prefer them. Although T. ni is a generalist insect, it seems to be better adapted to cope with the defense compounds of cabbage, although females oviposit more often on young than mature leaves. The concentration of glucosinolates is higher in young compared with mature leaves of Brassicaceae plants (Badenes-Perez et al. 2014; Moreira et al. 2016). Glucosinolates act as potent oviposition and feeding stimulants for more than 25 insect species that are specialized on brassicaceous plants and are widely accepted to have a negative impact on generalist insect herbivores, yet detailed studies of their impact on the biology of both generalists and specialists are relatively sparse (Hopkins et al. 2009). Another possibility is that T. ni is less parasitized on cabbage than on the other two host plants, viz. tomato and soybean. To the best of our knowledge, no studies have evaluated whether parasitism of T. ni differs between host plant species.
We found that T. ni females preferred to oviposit on mature compared with young leaves, except in cabbage. Similar results have been reported for other insects. For example, Adoxophyes honmai Yasuda females preferred to oviposit on old rather than young tea leaves (Piyasaengthong et al. 2016). However, most of the neonate larvae fed on young leaves, because after hatching they moved to young leaves to start feeding. In contrast, Plutella xylostella L. oviposited more eggs on young than older leaves of Barbarea plants (Badenes-Perez et al. 2014), but preferred to lay on mature leaves of collar, cauliflower, and cabbage (Moreira et al. 2016). We found that glandular and nonglandular trichomes negatively affected oviposition of T. ni females, suggesting that both physical and chemical cues can inhibit oviposition of T. ni females on young leaves; for example, secondary metabolites may act as oviposition stimulants or deterrents for gravid females (Schoonhoven et al. 2005). Generally, young leaves have higher concentrations of secondary metabolites compared with mature leaves (de Boer 1999; Badenes-Perez et al. 2014; Moreira et al. 2016).
We found that mature leaves of tomato, soybean, and wormseed plants had lower density of trichomes compared with young leaves, and females deposited fewer eggs on young leaves. It is possible that compounds present in glandular trichomes of young leaves act as repellents or oviposition deterrents for T. ni females. However, we also observed that soybean trichomes act as a mechanical defense, because they prevented neonate larvae from feeding on young leaves and negatively affected their movement. Previous study showed that a soybean cultivar with high trichome density was resistant to T. ni larvae, but became susceptible when the trichomes were shaved off (Khan et al. 1986). The trichomes on soybean secrete viscous liquid that hardens as soon as it comes into contact with air, acting like a sticky glue (Metcalf and Luckmann 1982). On the other hand, Woodman and Fernandes (1991) showed that trichomes on Verbascum thapsus L. provide protection against leaf-chewing insects and help to reduce water loss by transpiration from young leaves. Gonzáles et al. (2008) reported that, under water shortage, there is an increment in the density of nonglandular trichomes in Madia sativa Molina. In this study, nonglandular (flagelliform) trichomes were absent from the abaxial surface of wormseed, a result that does not coincide with that reported by Sá et al. (2016), who reported presence of trichomes on both sides of the leaf. Further studies will be performed to investigate in more detail the role of physical or chemical cues as factors to inhibit oviposition of T. ni females on young leaves, and whether larvae perform better on mature rather than young leaves.
In summary, the results of this study confirm that, with some plant species, there is no correlation between female preference and larval performance of T. ni. This lack of correlation is independent of whether cut leaves (Cameron et al. 2007; Shikano et al. 2010) or whole plants (this study) are used. Generally, it seems that there is a correlation between female preference and larval performance of T. ni with plants from Brassicaceae (Cameron et al. 2007; Shikano et al. 2010; this study), which suggests that these plants may be the ancestral hosts of the cabbage looper moth. We also found that there was considerable movement of older larvae among different plants, being particularly notable among tomato, soybean, and wormseed. This suggests that older larvae may be able to search for suitable host plants by themselves. Such larval behavior could remove pressure on females to search for suitable host plants for their offspring. Indeed, restricted offspring movement is a potential critical factor promoting female preference for good-quality host plants (Thompson 1988; Gripenberg et al. 2010). We therefore recommend use of neonate larvae for performance studies.
References
Badenes-Perez FR, Gershenzon J, Heckel DG (2014) Insect attraction versus plant defense: young leaves high in glucosinolates stimulate oviposition by a specialist herbivore despite poor larval survival due to high saponin content. PLoS One 9:39–42
Berdegué M, Reitz SR, Trumble JT (1998) Host plant selection and development in Spodoptera exigua: do mother and offspring know best? Entomol Exp Appl 89:57–64
Bernays EA (2001) Neural limitation in phytophagous insect: implications for diet breadth. Am Nat 151:451–464
Bonzani NE, Barboza GE, Bugatti MA, Espinar LA (2003) Morpho-histological studies in the aromatic species of Chenopodium from Argentina. Fitoterapia 74:207–225
Cameron JH, Isman MB, Upadhyaya MK (2007) Trichoplusia ni growth and preference on broccoli and eight common agricultural weeds. Can J Plant Sci 87:413–421
Chow JK, Akhtar Y, Isman MB (2005) The effects of larval experience with a complex plant latex on subsequent feeding and oviposition by the cabbage looper moth: Trichoplusia ni (Lepidoptera: Noctuidae). Chemoecology 15:129–133
Coapio GG, Cruz-López L, Guerenstein P, Malo EA, Rojas JC (2016) Herbivore damage and prior egg deposition on host plants influence the oviposition of the generalist moth Trichoplusia ni (Lepidoptera: Noctuidae). J Econ Entomol 109:2364–2372
de Boer NJ (1999) Pyrrolizidine alkaloid distribution in Senecio jacobaea rosettes minimises losses to generalist feeding. Entomol Exp Appl 91:169–173
Franceschi VR, Giaquinta RT (1983) Glandular trichomes of soybean leaves: cytological differentiation from initiation through senescence. Bot Gaz 144:175–184
Glas JJ, Schimmel BCJ, Alba JM, Escobar-Bravo R, Schuurink RC, Kant MR (2012) Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int J Mol Sci 13:17077–17103
Gonzáles WL, Negritto MA, Suárez LH, Gianoli E (2008) Induction of glandular and non-glandular trichomes by damage in leaves of Madia sativa under contrasting water regimes. Acta Oecol 33:128–132
Gripenberg S, Mayhew PJ, Parnell M, Roslin T (2010) A meta-analysis of preference-performance relationships in phytophagous insects. Ecol Lett 13:383–393
Haukioja E, Niemelä P (1977) Retarded growth of a geometrid larva after mechanical damage to leaves of its host tree. Ann Zool Fenn 14:48–72
Hopkins RJ, Van Dam NM, Van Loon JJA (2009) Role of glucosinolates in insect-plant relationships and multitrophic interactions. Annu Rev Entomol 54:57–83
Jaenike J (1978) On optimal oviposition behaviour in phytophagous insects. Theor Popul Biol 14:350–356
Kennedy GG (2003) Tomato, pests, parasitoids, and predators: tritrophic interaction involving the genus Lycopersicon. Annu Rev Entomol 48:51–72
Khan ZR, Ward JT, Norris DM (1986) Role of trichomes in soybean resistance to cabbage looper, Trichoplusia ni. Entomol Exp Appl 42:109–117
Knolhoff LM, Heckel DG (2014) Behavioral assays for studies of host plant choice and adaptation in herbivorous insects. Annu Rev Entomol 59:263–278
Landolt P (1993) Effects of host plant damage on cabbage looper moth attraction and oviposition. Entomol Exp Appl 67:79–85
Lange BM (2015) The evolution of plant secretory structures and the emergence of terpenoid chemical diversity. Annu Rev Plant Biol 66:139–159
Levins R, MacArthur R (1969) An hypothesis to explain the incidence of monophagy. Ecology 50:910–911
Metcalf RL, Luckmann WH (1982) Introduction to Insect Pest Management. Wiley, New York (USA)
Moreira LF, Teixeira NC, Santos NA, Valim JOS, Maurício RM, Guedes RNC, Oliveira MGA, Campos WG (2016) Diamondback moth performance and preference for leaves of Brassica oleracea of different ages and strata. J Appl Entomol 140:627–635
Mutyambai DM, Midega CAO, Bruce TJA, van den Berg J, Pickett JA, Khan ZR (2014) Behaviour and biology of Chilo partellus on maize landraces. Entomol Exp Appl 153:170–181
Permana AD, Johari A, Putra RE, Sastrodihardjo S, Ahmad I (2012) The influence of trichome characters of soybean (Glycine max Merrill) on oviposition preference of soybean pod borer Etiella zinckenella Treitschke (Lepidoptera: Pyralidae) in Indonesia. J Entomol Nematol 4:15–21
Piyasaengthong N, Sato Y, Kinoshita N, Kainoh Y (2016) Oviposition preference for leaf age in the smaller tea tortrix Adoxophyes honmai (Lepidoptera: Tortricidae) as related to performance of neonates. Appl Entomol Zool 51:363–371
R Development Core Team (2014) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Austria [WWW document]. URL http://www.R-project.org/ [accessed on 11 February 2017]
Rausher MD (1979) Egg recognition: its advantage to a butterfly. Anim Behav 27:1034–1040
Refsnider JM, Janzen JF (2010) Putting eggs in one basket: ecological and evolutionary hypotheses for variation in oviposition-site choice. Annu Rev Ecol Evol Syst 41:39–57
Reymond P, Weber H, Damond M, Farmer EE (2000) Differential gene expression in response to mechanical wooding and insect feeding in Arabidopsis. Plant Cell 12:707–720
Rojas JC, Virgen A, Cruz-López L (2003) Chemical and tactile cues influencing oviposition of a generalist moth, Spodoptera frugiperda (Lepidoptera: Noctuidae). Environ Entomol 32:1386–1392
Sá RD, Santana ASCO, Silva FCL, Alberto L, Soares L, Randau KP (2016) Anatomical and histochemical analysis of Dysphania ambrosioides supported by light and electron microscopy. Rev Bras Farmacogn 26:533–543
Sadek MM, Hansson BS, Anderson P (2010) Does risk of egg parasitism affect choice of oviposition sites by a moth? A field and laboratory study. Basic Appl Ecol 11:135–143
Savatin DV, Gramegna G, Modesti V, Cervone F (2014) Wounding in the plant tissue: the defense of a dangerous passage. Front Plant Sci 5:470
Schoonhoven LM, Van Loon JJA, Dicke M (2005) Insect-Plant Biology. Oxford University Press, New York, p 419
Shikano I, Akhtar Y, Isman MB (2010) Relationship between adult and larval host plant selection and larval performance in the generalist moth, Trichoplusia ni. Arthropod Plant Interact 4:197–205
Simmons AT, Gurr GM (2005) Trichomes of Lycopersicon species and their hybrids: effects on pests and natural enemies. Agric For Entomol 7:265–276
Simmons AT, McGrath D, Gurr GM (2005) Trichome characteristics of F1 Lycopersicon esculentum x L. cheesmanii f. minor and L. esculentum x L. pennellii hybrids and effects on Myzus persicae. Euphytica 144:313–320
Soo Hoo CR, Coudriet DL, Vail PV (1984) Trichoplusia ni (Lepidoptera: Noctuidae) larval development on wild and cultivated plants. Environ Entomol 13:843–846
Stephens DW, Krebs (1986) Foraging Theory. Princeton University Press, New Jersey
Sutherland DWS, Greene GL (1984). Cultivated and wild host plants, In: Lingren PD, Greene GL (eds). Suppression and management of cabbage looper populations. USDA Tech Bull 1684 pp. 1–13.
Thompson JN (1988) Evolutionary ecology of the relationship between oviposition preference and performance of off spring in phytophagous insects. Entomol Exp Appl 47:3–14
Thompson J, Pellmyr O (1991) Evolution of oviposition behavior and host preference in Lepidoptera. Annu Rev Entomol 36:65–89
Wagner GJ, Wang E, Shepherd RW (2004) New approaches for studying and exploiting an old protuberance, the plant trichome. Ann Bot 93:3–11
Woodman RL, Fernandes GW (1991) Differential mechanical defense: herbivory, evapotranspiration, and leaf hairs. Oikos 60:11–19
Acknowledgements
We thank Armando Virgen for assistance in rearing the insects and growing the plants used in this study, Guadalupe Nieto for microscopic assistance during description of trichomes, and Javier Valle Mora for his statistical advice. This study was supported by CONACYT through grant CB 2011-165450 to J.C.R. and a graduate scholarship to G.G.C.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Heikki Hokkanen.
Rights and permissions
About this article

Cite this article
Coapio, G.G., Cruz-López, L., Guerenstein, P. et al. Oviposition preference and larval performance and behavior of Trichoplusia ni (Lepidoptera: Noctuidae) on host and nonhost plants. Arthropod-Plant Interactions 12, 267–276 (2018). https://doi.org/10.1007/s11829-017-9566-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-017-9566-9