
Abstract
sn-2-Monoacylglycerol (2-MAG) and sn-1(3)-monoacylglycerol [1(3)-MAG] are important but yet little studied intermediates in lipid metabolism. The current study compared the metabolic fate of 2-MAG and 1(3)-MAG in isolated caecal enterocytes and hepatocytes of brown trout (Salmo trutta). 1(3)-Oleoyl [9,10-3H(N)]-glycerol and 2-Oleoyl [9,10-3H(N)]-glycerol were prepared by pancreatic lipase digestion of triolein [9,10-3H(N)]. The 1(3)-MAG and 2-MAG were efficiently absorbed by enterocytes and hepatocytes at similar rates. The 2-MAG was quickly resynthesized into TAG through the monoacylglycerol acyltransferase (EC: 2.3.1.22, MGAT) pathway in both tissues, whereas 1(3)-MAG was processed into TAG and phospholipids at a much slower rate, suggesting 2-MAG was the preferred substrates for MGAT. Further analysis showed that 1(3)-MAG was synthesized into 1,3-DAG, but there were no accumulation of 1,3-DAG in either enterocytes or hepatocytes, which contrasts that of mammalian studies. Some of the 1(3)-MAG may be acylated to 1,2(2,3)-DAG and then utilized for TAG synthesis. Alternatively, 1(3)-MAG can be hydrolyzed to free fatty acid and glycerol, and re-synthesized into TAG through the glycerol-3-phosphate (Gro-3-P) pathway. The overall data suggested that the limiting step of the intracellular 1(3)-MAG metabolism is the conversion of 1(3)-MAG itself.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In mammals, pancreatic lipase (EC: 3.1.1.3) is the major lipase for triacylglycerol (TAG) digestion [1]. Although it has a strong sn-1,3-specificity towards TAG, it will also hydrolyze in 1 and 3 positions of TAG producing small amounts sn-1-monoacylglycerols (1-MAG) and sn-3-monoacylglycerols (3-MAG), respectively, in addition to the main product sn-2-monoacylglycerols (2-MAG) [2]. Mammals also possess bile salt-dependent lipase (BSDL; EC: 3.1.1.1) that has lower activity than pancreatic lipase [3]. BSDL has low positional specificity and will produce a variety of MAG in addition to the complete hydrolysis to glycerol and free fatty acids (FFA) [4, 5]. Overall, these processes will generate 2-MAG, FFA and glycerol as the major products of TAG digestion followed by small but significant amounts of 1-MAG and 3-MAG.
Fish do not seem to have a typical pancreatic lipase [6]. Rather the main digestive lipase appears to be versions of the BSDL [6], as shown in rainbow trout (Oncorhynchus mykiss) [7], red sea bream (Pagrus major) [8] and Atlantic salmon (Salmo salar) [9]. In most cases, fish BSDL appears to share the similarities of the mammalian BSDL in that it shows low positional specificity, attacking all three positions of TAG. Experimental incubations with midgut extract in Atlantic salmon have shown that all varieties of MAG are produced during TAG digestion with nearly half being 1,(3)-MAG [10]. Other studies have indicated some sn-1,3 specificity in Atlantic cod (Gadus morhua) [11, 12].
Following luminal digestion, all types of MAG are absorbed into enterocytes for further metabolism. The 2-MAG are quickly processed into TAG via the sequential acylation by monoacylglycerol acyltransferase (EC: 2.3.1.22, MGAT) and diacylglycerol acyltransferase (EC: 2.3.1.20, DGAT), the so-called MGAT pathway. When TAG is completely hydrolyzed to FFA and glycerol, the resynthesis of TAG needs to undergo the de novo pathway from glycerol-3-phosphate (Gro-3-P) pathway. It involves acylation of Gro-3-P into lyso-phosphatidic acid (lysoPtdOH), and then conversion to phosphatidic acid (PtdOH), which is hydrolyzed to 1,2-DAG and finally acylated to TAG by DGAT. It is an energy consuming pathway and is often observed in liver and adipose tissue rather than the intestine [13]. As in mammals, the MGAT pathway is well functioning in the intestine of Atlantic salmon [14, 15], sea bream (Sparus aurata) [16] and Atlantic cod [17].
However, as opposed to mammals, many fish species are also expected to produce significant amounts of 1- and 3-MAG during digestion that need to be processed efficiently following uptake into enterocytes. The fate of these MAG is, however, not well described in mammals, and not at all in fish. One possible pathway described in mammals is the hydrolysis of 1-MAG by monoacylglycerol lipase (EC: 3.1.1.23, MGL) to produce FFA and glycerol, which are then used as substrates for resynthesis of TAG through the Gro-3-P pathway [18, 19]. Alternatively, 1(3)-MAG can proceed to 1,3-DAG through the MGAT pathway [20, 21]. However, in mammals, DGAT does not seem to utilize this efficiently for TAG synthesis leading to the accumulation of 1,3-DAG [22]. Such DAG oil is becoming popular for humans because its possible anti-obesity effect [23], increasing fat oxidation and decreasing body weight [24]. If 1,3-DAG oil has a similar anti-obesity effect for fish, it would be detrimental in aquaculture when produced in large amounts, causing slower growth of fish.
Therefore, to gain better understanding of the metabolic fate of different MAG, the present study compares the fate of 2-MAG and 1(3)-MAG in isolated caecal enterocytes and hepatocytes of brown trout (Salmo trutta). To achieve this, radiolabeled 2-MAG and 1(3)-MAG were used as substrates for in vitro incubations. The incorporation of label into different lipid classes was analyzed and the pathways involved were discussed.
Materials and Methods
Experimental Fish
This study was carried out within the Norwegian animal welfare act guidelines, in accordance with the Animal Welfare Act of 20th December 1974, amended 19th June 2009, at a facility with permission to conduct experiments on fish (code 93) provided by the Norwegian Animal Research Authority (FDU, http://www.fdu.no). The experiment was approved by the Norwegian Animal Research Authority (NARA).
Brown trout were purchased from Settefiskanlegget Lundamo AS, and ranged from 400 to 700 g. They were kept at 10 °C in freshwater, with natural daylight regime at the experimental facility of NTNU Sealab in Trondheim, Norway. Fish were fed a standard commercial diet (Skretting AS) and they were starved for 48 h before the experiment.
Isolation of Caecal Enterocytes and Hepatocytes
Fish were killed by an overdose of MS222. The enterocytes and hepatocytes were prepared as described previously [25], with minor modification. In brief, pyloric caeca was dissected, cleaned of adhering tissue, finely chopped with scissors and incubated with 20 ml of Solution A [Hank’s balanced salt solution (HBSS) supplemented with 10 mM HEPES and 1 mM EDTA], containing 1 mg ml−1 collagenase for 45 min in an incubator at 20 °C. The digested tissue were filtered through a 100-μm cell strainer and the cells were collected by centrifugation at 500g for 5 min. The cell pellet was washed with 20 ml of solution A containing 10 mg ml−1 fatty acid free bovine serum albumin (FAF-BSA) and re-centrifuged at 500g for 3 min. The cell pellet was then washed with 30 ml freshly prepared solution B (calcium free minimum essential medium (MEM) supplemented with 100 U Penicillin ml−1, 100 μg Streptomycin ml−1, 0.25 µg Amphotericin B ml−1 and pH adjusted to 7.1–7.4 by sodium bicarbonate) and re-centrifuged at 500g for 3 min. The final cell pellet was resuspended in 10 ml of solution B. The liver was perfused via the hepatic vein and it was then treated the same as pyloric caeca.
For protein analysis of enterocytes and hepatocytes, 100 µl of the cell suspensions were collected and assayed by the method of Lowry [26].
Preparation of sn-1(3)-Monoacylglycerol (1(3)-MAG) and sn-2-Monoacylglycerol (2-MAG)
Both 1(3)-Oleoyl [9,10-3H(N)]-glycerol and 2-Oleoyl [9,10-3H(N)]-glycerol were prepared by pancreatic lipase (porcine pancreas, Sigma-Aldrich Co. MO) digestion of triolein [9,10-3H(N)] (American Radiolabeled Chemicals, Inc. MO) as described in detail by Oxley et al. [15], based on the method of Myher and Kuksis [27]. In brief, 10 mg of cold triolein was mixed with 50 μCi hot triolein, dried under a stream of nitrogen, and suspended in 1 ml of buffer (1 M Tris-pH 8, 100 g l−1 gum Arabic, 25 g l−1 CaCl2). Then 1 ml of buffer containing 10 mg of pancreatic lipase was added to the mixture, vortexed vigorously for 30 s and then incubated at 37 °C for 30 min. The reaction was terminated by 4 ml of ice-cold diethyl ether, and the upper phase was collected after centrifugation. The extraction was repeated four times with 2 ml of diethyl ether. The pooled diethyl ether passed through an anhydrous Na2SO4 column, evaporated under nitrogen and re-suspended in 30 µl of chloroform.
Both 1(3)-MAG and 2-MAG were separated on HPTLC (high performance thin layer chromatography) plates impregnated with boric acid (25 mg ml−1 in ethanol/water 1:1, by vol), in a chloroform/acetone (88:12, by vol) solvent system [28]. Lipid bands were visualized under UV light after spraying with 1 mg ml−1 2,7-dichlorofluorescein in methanol/water (95:5, by vol). Both 1(3)-MAG and 2-MAG were extracted from silica four times with 2 ml of chloroform/acetone (9:1, by vol) followed by rinsing the mixture with 2 ml of ice-cold water twice [29]. The hypophase was collected, passed through an anhydrous column and evaporated under N2. Both 1(3)-MAG and 2-MAG were dried in a vacuum desiccator for 30 min, weighed, resuspended in chloroform, and the specific activity determined. The yield of 2-MAG and 1-MAG was 18.7 and 3.6%, respectively, with purity better than 90% when stored in −80 °C and analyzed by borate-TLC before and after the experiments.
Incubation of Enterocytes and Hepatocytes
Either 1(3)-MAG or 2-MAG (40 µM final concentration, 0.06 μCi for each) were dried under N2, dissolved in ethanol (0.25% volume of the final volume), and subsequently dispersed in 40 µM FAF-BSA solution (50 mg ml−1 in solution B). The mixture was sonicated in ice-cold water for 5 min and used immediately to minimize isomerization.
The cell suspension was distributed in 1.895 ml aliquots and mixed with 0.105 ml of each radiolabeled MAG-BSA solution. After incubation for 15 min and 120 min at 20 °C, the cells were isolated by centrifugation at 500g for 2 min. To determine the exported lipids, 0.2 ml of the supernatant was transferred to a glass tube containing 2.5 ml ice-cold chloroform/methanol (2:1, by vol). The cell pellet was washed with 2 ml FAF-BSA in solution B (10 mg ml−1), re-centrifuged at 500g for 2 min and homogenized in 2.5 ml of ice-cold chloroform/methanol (2:1, by vol). Total lipid from the incubation medium and cell pellet was extracted according to Folch et al. [30]. The hypophase was transferred into a GC vial, evaporated under N2 and re-suspended in 20 µl of chloroform. The same procedure was repeated four times with fresh cells isolated from different fish individuals.
Lipid Class Separation
The major lipid classes were separated by double-development HPTLC [31]. HPTLC silica gel 60 plates without fluorescent indicator (10 × 10 cm, Merck KGaA Darmstadt, Germany) were activated at 110 °C for 1 h and 10 µl of the extracted lipid sample was applied as a 1 cm streak by a glass micro-syringe, along with oleic acid standards. The plates were first developed to a distance of 4.5 cm from the origin using methyl acetate/isopropanol/chloroform/methanol/0.25% KCl (25:25:25:10:9, by vol) solvent system, dried briefly by a hair drier for 10 s and then placed in a desiccator over dry NaOH for 30 min. Subsequently, plates were developed in hexane/diethyl ether/glacial acetic acid (80:20:2, by vol) to 8.8 cm from the origin.
1(3)-MAG, 2-MAG, 1,3-DAG and 1,2(2,3)-DAG were separated by borate-TLC plates (produced as described above), using chloroform/acetone (96:4, by vol) as the solvent system. A mixture of standards of 1-monoolein, 2-monoolein, 1,3-diolein, 1,2-diolein and oleic acid was added together with 10 µl of the extracted lipid sample.
Lipid bands were visualized by exposure to iodine vapors and quickly marked with a pencil. Each lipid band was scraped into respective scintillation vials containing 10 ml of scintillation cocktail and the amount of radioactivity determined by a liquid scintillation counter.
Calculations and Statistical Analysis
The cellular retained lipid was expressed in nmol mg−1 protein and was calculated as the sum of radioactivity recovered from cellular lipid classes, unesterified fatty acids and MAG. The experimental data were tested for statistical significance by using Independent-Samples t test or two-way analysis of variance (ANOVA) with time and MAG treatment as the two factors. Differences were considered significant if P < 0.05. All of the statistical tests were performed using SPSS 23 for Windows. All Tables were made in Excel 2010 and Figures by Sigma Plot 13.0.
Results
Total Retained Cellular Lipids
The total retained lipids in enterocytes and hepatocytes showed no significant differences between 15 and 120 min (Fig. 1). In general, the retained lipids derived from 2-MAG were slightly higher than those from 1(3)-MAG, but the differences were not significant in enterocytes (P > 0.05) and close to significant in hepatocytes (P = 0.052). The average of retained lipids in enterocytes (3.89 ± 1.46 nmol mg protein−1) was significantly higher (P < 0.05, t-test) than in hepatocytes (2.45 ± 0.64 nmol mg protein−1).

Total retained lipids in enterocytes (a) and hepatocytes (b) after 15 and 120 min of incubation with 1(3)-MAG (black) or 2-MAG (grey). Columns represent the means of four replicates ± SD. Main effects of time, treatment and the interaction between the two were determined by two-way ANOVA. NS not significant
Exported PtdCho and TAG
The exported PtdCho and TAG were determined by analyzing the lipid classes in the medium. No significant differences were found between 1(3)-MAG and 2-MAG treatment (Fig. 2), though slightly higher PtdCho and TAG levels were found in the 2-MAG treatment of hepatocytes at 120 min (Fig. 2b, d). In enterocytes, the exported PtdCho did not increase significantly after 120 min of incubation (Fig. 2a), but the exported TAG were significantly increased (Fig. 2c). In hepatocytes, both exported PtdCho and TAG increased significantly after 120 min incubation (Fig. 2b, d).

Exported PtdCho (a, c) and TAG (b, d) from enterocytes and hepatocytes incubated with 1(3)-MAG (black) or 2-MAG (grey). Columns represent the means of four replicates ± SD. Main effects of time, treatment and the interaction between the two were determined by two-way ANOVA. NS not significant
Lipid Classes
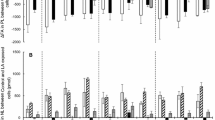
Figure 3 shows the distribution of radiolabeling in lipid classes of enterocytes and hepatocytes at 15–120 min. Similar patterns were found, with TAG and MAG being the predominating lipid classes, followed by PtdCho. The main differences between the 1(3)-MAG and 2-MAG treatments observed were the distribution pattern of these three lipid classes (TAG, MAG and PtdCho). The cellular retained MAG were higher in the 1(3)-MAG treatment (P < 0.05, t-test) compared to that from the 2-MAG treatment. Radiolabeled TAG derived from 2-MAG was significantly higher (P < 0.05, t-test) than that from 1(3)-MAG in both enterocytes and hepatocytes at 15 min. However, at 120 min, these differences became non-significant. In enterocytes, radiolabeled PtdCho was significantly higher in 2-MAG treatment compared to 1(3)-MAG treatment at both 15 and 120 min, but no significant differences were found in hepatocytes. There were no significant differences between the treatments in the other lipid classes (PtdSer&PtdIns, PtdEtn and DAG) and FFA.

Percentage radioactivity of each lipid class of total lipid of enterocytes (15 min, a; 120 min, b) and hepatocytes (15 min, c; 120 min, d) incubated with 1(3)-MAG (black) or 2-MAG (grey). Columns represent the means of four replicates ± SD. * Indicates significant difference (P < 0.05) between 1(3)-MAG and 2-MAG treatment
Two way ANOVA analysis using time and treatment as factors (Table 1) was performed to compare the distribution of radiolabeling in the different lipid classes. Significant differences between the treatments were found for PtdCho, MAG and TAG in enterocytes; and for MAG and TAG in hepatocytes. The radiolabeled PtdCho, PtdSer&PtdIns and PtdEtn increased significantly with time in both enterocytes and hepatocytes. Meanwhile, the cellular radiolabeled TAG remained stable. Accordingly, the PtdCho:TAG ratio increased significantly at 120 min compared to 15 min. There was a dramatic decrease in radiolabeled MAG from 15 to 120 min, but the remaining cellular MAG from the 1(3)-MAG treatment was still high compared to other lipid classes at 120 min (Fig. 3b), especially for enterocytes.
The averagess of all the phospholipids (PtdCho, PtdSer&PtdIns and PtdEtn) and PtdCho:TAG ratios in hepatocytes were significantly higher than that in enterocytes (P < 0.05, t-test), whereas the neutral lipids (MAG, DAG and TAG) and FFA were similar (P > 0.05, t-test).
Percentage of MAG and DAG Species
In order to better understand the metabolism pathway of different MAG, the cellular MAG and DAG were further separated into 1(3)-MAG, 2-MAG, 1,2(2,3)-DAG and 1,3-DAG by borate-TLC plates. The cellular 1(3)-MAG derived from added 1(3)-MAG decreased slightly from 15 min (30.5 ± 7.9%) to 120 min (25.6 ± 9.5%) (P > 0.05) in enterocytes, which was significantly higher compared to the retained 1(3)-MAG derived from 2-MAG substrate (Fig. 4a). In hepatocytes, the cellular 1(3)-MAG derived from 1(3)-MAG substrate was significantly higher than that from 2-MAG substrate, but it decreased significantly from 15 to 120 min (Fig. 4b). On the contrary, the cellular 2-MAG showed no significant differences between the two treatments in either enterocytes or hepatocytes (Fig. 4c, d), suggesting a fast utilization of 2-MAG in the cells. There was a slight but significant decrease in the cellular 2-MAG from 15 to 120 min in both enterocytes and hepatocytes.

Percentage radioactivity of 1(3)-MAG (a, b) and 2-MAG (c, d) of total lipids of enterocytes and hepatocytes incubated with 1(3)-MAG (black) or 2-MAG (grey). Columns represent the means of four replicates ± SD. Main effects of time, treatment and the interaction between the two were determined by two-way ANOVA. NS not significant
In enterocytes, the retained 1,2(2,3)-DAG derived from the 2-MAG substrate was significantly higher than that from 1(3)-MAG substrate, and the 1,2(2,3)-DAG level remained stable from 15 to 120 min (Fig. 5a). In hepatocytes, the retained 1,2(2,3)-DAG also remained stable, but no significant difference was found between the treatments (Fig. 5b). Surprisingly, the retained 1,3-DAG was not different between the two treatments in enterocytes (Fig. 5c), and only slightly different (P < 0.05) in hepatocytes (Fig. 5d). The 1,3-DAG level remained low and stable at 15 and 120 min.

Percentage radioactivity of 1,2(2,3)-DAG (a, b) and 1,3-DAG (c, d) of total lipid of enterocytes and hepatocytes incubated with 1(3)-MAG (black) or 2-MAG (grey). Columns represent the means of four replicates ± SD. Main effects of time, treatment and the interaction between the two were determined by two-way ANOVA. NS not significant
Percentage of 1(3)-MAG and 1,3-DAG of Total Cellular MAG and DAG
In order to better understand the fate of 1(3)-MAG and 1,3-DAG, the data in Fig. 6 are presented as the percentages of radiolabeled 1(3)-MAG or 1,3-DAG in total radiolabeled MAG or DAG. When enterocytes and hepatocytes were incubated with 1(3)-MAG, the cellular 1(3)-MAG accounted for more than 84% of the total MAG and it was stable (t-test) with time (Fig. 6a, b). In contrast, when cells were incubated with 2-MAG, the percentage of cellular 1(3)-MAG of total MAG increased significantly from 15 min to 120 min, which was mainly caused by isomerization, but it was still significantly (t-test) lower compared to the levels in 1(3)-MAG treatment.

Percentage radioactivity of 1(3)-MAG (a, b) and 1,3-DAG (c, d) of total MAG and DAG of enterocytes and hepatocytes incubated with 1(3)-MAG (black) or 2-MAG (grey). Columns represent the means of four replicates ± SD. Main effects of time, treatment and the interaction between the two were determined by two-way ANOVA. NS not significant
The synthesis of 1,3-DAG in the enterocytes accounted for 54% of the total DAG in the 1(3)-MAG treatment, which was significantly higher compared to the 2-MAG treatment (~25%) at both 15 and 120 min (Fig. 6c). Similar pattern was found for the hepatocytes, where 1,3-DAG synthesis accounted for ~30 versus ~15% in the 1(3)-MAG treatment compared to 2-MAG treatment (Fig. 6d). However, the percentage of 1,3-DAG of total DAG remained stable with time. These data suggested that 1,3-DAG is synthesized by the acylation of 1(3)-MAG in both enterocytes and hepatocytes.
Discussion
In salmonids, results suggest that the bile activated lipase lacks the typical sn-1,3-specificity known to exist in mammalian pancreatic lipase. This could potentially result in the production of all types of MAG. How these MAG, especially 1(3)-MAG, are being processed in fish was not known. Therefore, the main objective of this study was to investigate the comparable metabolic fate of 1(3)-MAG and 2-MAG in enterocytes and hepatocytes of brown trout. The results showed that the fate of 2-MAG in fish was similar to that in mammals (efficiently incorporated into TAG), but there are major differences in the fate of 1(3)-MAG.
MAG Uptake and Lipoprotein (as TAG) Export
At low luminal concentrations (µM), the uptake of sn-2-monoolein was slower compared to the uptake of 18:1n-9 in Caco-2 cells [32]. Previous studies have shown that the uptake of radiolabeled fatty acid by trout caecal enterocytes was close to 1 nmol mg−1 protein over a 2 h incubation period when incubated with 2 µM fatty acid [33]. In the present study, the retained lipids in enterocytes was higher (~4 nmol mg−1 protein), showing that the higher substrate concentrations (40 µM MAG) ensured a maximum uptake rate.
We also found that the retained fatty acids in enterocytes were significantly higher than hepatocytes. This agrees with findings from Diaz-Lopez et al. [34] who used radiolabeled free fatty acid (FFA) in gilthead seabream (Sparus aurata), indicating a general trend across species. The retained cellular lipids did not increase at 120 min compared to 15 min, which could be explained by a significant (P < 0.05) export of lipoproteins (as TAG) at longer incubations as shown in Fig. 2.
Metabolic Fate of 2-MAG and 1(3)-MAG
It is well known that 2-MAG is efficiently reesterified to TAG via the MGAT pathway in enterocytes and hepatocytes in both mammals and fish [13–15, 17, 35]. In this respect, fish and mammals have similar enzyme specificities. The fate of 1(3)-MAG is however little studied in both mammals and fish. The lack of interest is probably due to the high efficiency of mammalian pancreatic lipase producing 2-MAG as the main degradation product [1]. However, in mammals significant amounts of 1(3)-MAG are still produced during lipid digestion [2], and in salmon, data indicate that up to 50% of TAG may be digested to 1(3)-MAG [10].
In mammals, 1(3)-MAG has been suggested to be processed through different pathways. Firstly, 1(3)-MAG could be hydrolyzed to FFA and glycerol by monoacylglycerol lipase (MGL) and then synthesized to TAG following the Gro-3-P pathway. The argument for this pathway can be suggested from Caco-2 cell cultures where 20% of label was found in FFA following incubation with [3H] sn-2-monoolein for 2 h [32]. The typical enzyme for this, a MGL, has been found in many tissues in mammals [36–38]. The enzyme hydrolyzes 1-MAG and 2-MAG with similar efficiencies [39]. Likewise, in brown trout we found that 8% of label in absorbed MAG was recovered in the FFA fraction after 2 h suggesting the possibility of a MAG lipase. The function of MGL in the intestine is unknown, but it could be part of a reorganization of the MAG [37, 38]. A recent study suggested MGL perform the same function in Grass carp (Ctenopharyngodon idella) as in mammals due to conserved gene structure and functional domain [40], but its contribution in the digestion processes remains to be explored.
Secondly, 1-MAG can be processed to 1,3-DAG by MGAT. As 1,3-DAG is a poor substrate for DGAT in mammals, DAG has been reported to accumulate in the intestine with slow processing to TAG. This has been shown to occur in rat intestinal microsomes [41], rat enterocytes [22, 23] and hamster intestinal mucosal homogenates [42]. Although we also observed a significantly slower utilization of 1(3)-MAG compared to 2-MAG in both enterocytes and hepatocytes, the total DAG pool did not increase following incubation with 1(3)-MAG. The acylation to TAG could still have proceeded through 1,3-DAG though as it was still more than half of the DAG pool in the enterocytes incubated with 1(3)-MAG. If this is the case, then 1,3-DAG would have been immediately utilized for synthesis of TAG, which would require the presence of a DGAT with high activity towards 1,3-DAG. This remains to be demonstrated.
Alternatively, variants of the MGAT may possess sn-2-acylation activity producing 1,2(2,3)-DAG from 1(3)-MAG. Two such acyltransferases (MGAT1 and MGAT2) have been cloned from mouse showing some acylating activity towards both 1-MAG and 3-MAG in the sn-2 position [20, 21]. In cell homogenates prepared from mouse MGAT2 transfected COS-7 cells, nearly 20% of the total DAG was 1,2(2,3)-DAG when the substrate was rac-1-oleoylglycerol [21]. It is possible that some trout MGAT have similar activities. Our data showed that more than 40 and 60% of the total DAG was 1,2 (2,3)-DAG, produced by enterocytes and hepatocytes, respectively, following incubation with 1(3)-MAG. This could be a natural consequence of salmonids not having a 1,3-specific digestive lipase. Therefore, this pathway could contribute to the utilization of 1(3)-MAG for TAG synthesis.
Finally, monoacylglycerol kinase (EC: 2.7.1.94) catalyzing the biosynthesis of lysophosphatidic acid (lysoPtdOH) from 1-MAG has been demonstrated in brain of mammals [43]. The lysoPtdOH could be further acylated to phosphatidic acid (PtdOH) and finally to TAG or phospholipids following the Gro-3-P pathway [13]. Theoretically, it is a more efficient pathway compared to the hydrolysis of MAG and phosphorylation of glycerol. However, it did not seem to be an important pathway in brown trout, because the label in phospholipids derived from 1(3)-MAG was not more than that derived from 2-MAG.
Hepatocytes Versus Enterocytes
In mammals, the MGAT pathway is the predominant pathway for TAG synthesis in the intestine, whereas TAG is synthesized mainly via the Gro-3-P pathway in liver [13]. The Gro-3-P pathway will produce exclusively 1,2-DAG, while the MGAT pathway would produce all variants including 1,2-DAG and 2,3-DAG. While 1,2-DAG and 2,3-DAG are both good substrates for DGAT for the production of TAG, only 1,2-DAG can be used for the synthesis of PtdCho. This leads to a higher PtdCho:TAG ratio from the Gro-3-P pathway compared to the MGAT pathway [15]. In line with this, significantly higher PtdCho:TAG ratios were found in hepatocytes in the present study compared to enterocytes thus confirming the higher Gro-3-P pathway activity in hepatocytes in fish as well. In the 1(3)-MAG treatment, the cellular 1(3)-MAG level in hepatocytes decreased significantly at 120 min compared to 15 min, but it only decreased slightly in enterocytes. Therefore, compared to enterocytes, hepatocytes could better utilize 1(3)-MAG, possibly for the biosynthesis of phospholipids via the Gro-3-P pathway. However, more studies are needed to clarify the contribution of each pathway. The positional distribution of fatty acid in TAG may be helpful to better understand the pathway. However, the remodeling of TAG via hydrolysis and re-esterification must be considered, as up to 70% of the liver TAG derived from Gro-3-P pathway was found to be remodeled before secretion as very low density lipoproteins [44].
In summary, both 1(3)-MAG and 2-MAG were absorbed efficiently by enterocytes and hepatocytes and esterified into different lipid classes. 2-MAG was quickly resynthesized into TAG through the MGAT pathway in both enterocytes and hepatocytes, whereas 1(3)-MAG was processed to TAG and phospholipids at a much slower rate. The possible pathway of 1(3)-MAG metabolism was discussed based on the results (Figs. 3, 6). It could be hydrolyzed by MGL and utilized for TAG synthesis via the Gro-3-P pathway; alternatively, it could be acylated at the sn-3 or sn-2 position to form 1,3-DAG or 1,2(2,3)-DAG, respectively. However, further studies are needed to demonstrate the contribution of each pathway. The small 1,3-DAG pool in the enterocytes and hepatocytes incubated with 1(3)-MAG may suggest a high DGAT activity towards 1,3-DAG in trout. It seems that hepatocytes utilize 1(3)-MAG better, possibly via the Gro-3-P pathway. The overall data suggested that the limiting step of the intracellular 1(3)-MAG metabolism was the conversion of 1(3)-MAG itself.
Abbreviations
- 1(3)-MAG:
-
sn-1(3)-Monoacylglycerol
- 1,2(2,3)-DAG:
-
sn-1,2(2,3)-Diacylglycerol
- 1,3-DAG:
-
sn-1,3-Diacylglycerol
- 1-MAG:
-
sn-1-Monoacylglycerol
- 2-MAG:
-
sn-2-Monoacylglycerol
- BSDL:
-
Bile salt-dependent lipase
- DAG:
-
Diacylglycerol(s)
- DGAT:
-
Diacylglycerol acyltransferase
- FAF-BSA:
-
Fatty acid free bovine serum albumin
- FFA:
-
Unesterified fatty acids
- Gro-3-P:
-
Glycerol-3-phosphate
- HBSS:
-
Hank’s balanced salt solution
- HPTLC:
-
High performance thin layer chromatography
- lysoPtdOH:
-
Lysophosphatidic acid
- MAG:
-
Monoacylglycerol(s)
- MEM:
-
Minimum essential medium
- MGAT:
-
Monoacylglycerol acyltransferase
- MGL:
-
Monoacylglycerol lipase
- MS222:
-
Tricaine methanesulfonate
- PtdCho:
-
Phosphatidylcholine
- PtdEtn:
-
Phosphatidylethanolamine
- PtdIns:
-
Phosphatidylinositol
- PtdOH:
-
Phosphatidic acid
- PtdSer:
-
Phosphatidylserine
- TAG:
-
Triacylglycerol(s)
References
Decaro A, Figarella C, Amic J, Michel R, Guy O (1977) Human pancreatic lipase—glycoprotein. Biochim Biophys Acta 490:411–419
Mattson FH, Volpenhein RA (1964) The digestion and absorption of triglycerides. J Biol Chem 239:2772–2777
Mattson FH, Volpenhein RA (1968) Hydrolysis of primary and secondary esters of glycerol by pancreatic juice. J Lipid Res 9:79–84
Wang CS, Kuksis A, Manganaro F, Myher JJ, Downs D, Bass HB (1983) Studies on the substrate specificity of purified human milk bile salt-activated lipase. J Biol Chem 258:9197–9202
Lombardo D (2001) Bile salt-dependent lipase: its pathophysiological implications. BBA-Mol Cell Biol Lipids 1533:1–28
Olsen RE, Ringø E (1997) Lipid digestibility in fish: a review. Recent Res Dev Lipid Res 1:199–265
Tocher DR, Sargent JR (1984) Studies on triacylglycerol, wax ester and sterol ester hydrolases in intestinal ceca of rainbow trout (Salmo gairdneri) fed diets rich in triacylglycerols and wax esters. Comp Biochem Physio 77:561–571
Iijima N, Tanaka S, Ota Y (1998) Purification and characterization of bile salt-activated lipase from the hepatopancreas of red sea bream, Pagrus major. Fish Physiol Biochem 18:59–69
Bogevik AS, Tocher DR, Waagbo R, Olsen RE (2008) Triacylglycerol-, wax ester- and sterol ester-hydrolases in midgut of Atlantic salmon (Salmo salar). Aquac Nutr 14:93–98
Bogevik AS, Oxley A, Olsen RE (2008) Hydrolysis of acyl-homogeneous and fish oil triacylglycerols using desalted midgut extract from Atlantic salmon, Salmo salar. Lipids 43:655–662
Gjellesvik DR (1991) Fatty acid specificity of bile salt-dependent lipase enzyme recognition and super-substrate effects. Biochim Biophys Acta 1086:167–172
Gjellesvik DR, Lombardo D, Walther BT (1992) Pancreatic bile salt dependent lipase from cod (Gadus morhua) purification and properties. Biochim Biophys Acta 1124:123–134
Lehner R, Kuksis A (1996) Biosynthesis of triacylglycerols. Prog Lipid Res 35:169–201
Oxley A, Torstensen BE, Rustan AC, Olsen RE (2005) Enzyme activities of intestinal triacylglycerol and phosphatidylcholine biosynthesis in Atlantic salmon (Salmo salar L.). Comp Biochem Physiol B-Biochem Mol Biol 141:77–87
Oxley A, Jutfelt F, Sundell K, Olsen RE (2007) Sn-2-monoacylglycerol, not glycerol, is preferentially utilised for triacylglycerol and phosphatidylcholine biosynthesis in Atlantic salmon (Salmo salar L.) intestine. Comp Biochem Physiol B-Biochem Mol Biol 146:115–123
Caballero MJ, Gallardo G, Robaina L, Montero D, Fernandez A, Izquierdo M (2006) Vegetable lipid sources affect in vitro biosynthesis of triacylglycerols and phospholipids in the intestine of sea bream (Sparus aurata). Br J Nutr 95:448–454
Li KS, Olsen RE, Ostensen MA, Altin D, Kjorsvik E, Olsen Y (2016) Atlantic cod (Gadus morhua) larvae can biosynthesis phospholipid de novo from 2-oleoyl-glycerol and glycerol precursors. Fish Physiol Biochem 42:137–147
Senior JR, Isselbacher KJ (1963) Demonstration of an intestinal monoglyceride lipase: an enzyme with a possible role in the intracellular completion of fat digestion. J Clin Invest 42:187–195
De Jong B, Hülsmann W (1978) Monoacylglycerol hydrolase activity of isolated rat small intestinal epithelial cells. BBA-Lipid Lipid Metab 528:36–46
Yen CLE, Farese RV (2003) MGAT2, a monoacylglycerol acyltransferase expressed in the small intestine. J Biol Chem 278:18532–18537
Cao JS, Lockwood J, Burn P, Shi YG (2003) Cloning and functional characterization of a mouse intestinal Acyl-CoA: monoacylglycerol acyltransferase, MGAT2. J Biol Chem 278:13860–13866
Kondo H, Hase T, Murase T, Tokimitsu I (2003) Digestion and assimilation features of dietary DAG in the rat small intestine. Lipids 38:25–30
Murase T, Aoki M, Wakisaka T, Hase T, Tokimitsu I (2002) Anti-obesity effect of dietary diacylglycerol in C57BL/6 J mice: dietary diacylglycerol stimulates intestinal lipid metabolism. J Lipid Res 43:1312–1319
Hibi M, Takase H, Meguro S, Tokimitsu I (2009) The effects of diacylglycerol oil on fat oxidation and energy expenditure in humans and animals. BioFactors 35:175–177
Tocher DR, Fonseca-Madrigal J, Bell JG, Dick JR, Henderson RJ, Sargent JR (2002) Effects of diets containing linseed oil on fatty acid desaturation and oxidation in hepatocytes and intestinal enterocytes in Atlantic salmon (Salmo salar). Fish Physiol Biochem 26:157–170
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Myher J, Kuksis A (1979) Stereospecific analysis of triacylglycerols via racemic phosphatidylcholines and phospholipase C. Can J Biochem 57:117–124
Thomas A III, Scharoun J, Ralston H (1965) Quantitative estimation of isomeric monoglycerides by thin-layer chromatography. J Am Oil Chem Soc 42:789–792
Christie W (2003) Lipid analysis, isolation, separation and structural analysis of lipids. J Barnes and Associates, Dundee
Folch J, Lees M, Stanley GHS (1957) A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem 226:497–509
Olsen RE, Henderson RJ (1989) The rapid analysis of neutral and polar marine lipids using double-development HPTLC and scanning densitometry. J Exp Mar Biol Ecol 129:189–197
Ho SY, Delgado L, Storch J (2002) Monoacylglycerol metabolism in human intestinal Caco-2 cells—evidence for metabolic compartmentation and hydrolysis. J Biol Chem 277:1816–1823
Oxley A, Tocher DR, Torstensen BE, Olsen RE (2005) Fatty acid utilisation and metabolism in caecal enterocytes of rainbow trout (Oncorhynchus mykiss) fed dietary fish or copepod oil. BBA-Mol Cell Biol Lipids 1737:119–129
Diaz-Lopez M, Perez MJ, Acosta NG, Jerez S, Dorta-Guerra R, Tocher DR, Lorenzo A, Rodriguez C (2010) Effects of dietary fish oil substitution by Echium oil on enterocyte and hepatocyte lipid metabolism of gilthead seabream (Sparus aurata L.). Comp Biochem Physiol B-Biochem Mol Biol 155:371–379
Hiramine Y, Emoto H, Takasuga S, Hiramatsu R (2010) Novel acyl-coenzyme A: monoacylglycerol acyltransferase plays an important role in hepatic triacylglycerol secretion. J Lipid Res 51:1424–1431
Young SG, Zechner R (2013) Biochemistry and pathophysiology of intravascular and intracellular lipolysis. Genes Dev 27:459–484
Chon SH, Zhou YX, Dixon JL, Storch J (2007) Intestinal monoacylglycerol metabolism: developmental and nutritional regulation of monoacylglycerol lipase and monoacylglycerol acyltransferase. J Biol Chem 282:33346–33357
Duncan M, Thomas AD, Cluny NL, Patel A, Patel KD, Lutz B, Piomelli D, Alexander SP, Sharkey KA (2008) Distribution and function of monoacylglycerol lipase in the gastrointestinal tract. Am J Physiol Gastrointest Liver Physiol 295:G1255–G1265
Tornqvist H, Belfrage P (1976) Purification and some properties of a monoacylglycerol-hydrolyzing enzyme of rat adipose tissue. J Biol Chem 251:813–819
Sun J, Ji H, Li XX, Shi XC, Du ZY, Chen LQ (2016) Lipolytic enzymes involving lipolysis in Teleost: synteny, structure, tissue distribution, and expression in grass carp (Ctenopharyngodon idella). Comp Biochem Physiol B Biochem Mol Biol 198:110–118
Lehner R, Kuksis A, Itabashi Y (1993) Stereospecificity of monoacylglycerol and diacylglycerol acyltransferases from rat intestine as determined by chiral phase high-performance liquid chromatography. Lipids 28:29–34
Brown JL, Johnston JM (1964) The mechanism of intestinal utilization of monoglycerides. BBA-Lipid Lipid Metab 84:264–274
Pieringer RA, Hokin LE (1962) Biosynthesis of lysophosphatidic acid from monoglyceride and adenosine triphosphate. J Biol Chem 237:653–658
Wiggins D, Gibbons GF (1992) The lipolysis/esterification cycle of hepatic triacylglycerol. Its role in the secretion of very-low-density lipoprotein and its response to hormones and sulphonylureas. Biochem J 284(Pt 2):457–462
Acknowledgements
This work was supported by the Norwegian Research Council (Grant No. 245327). The authors wish to thank the staff at NTNU Sealab for excellent technical assistance.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There are no conflicts of interest to report.
About this article

Cite this article
Li, K., Olsen, R.E. Metabolism of sn-1(3)-Monoacylglycerol and sn-2-Monoacylglycerol in Caecal Enterocytes and Hepatocytes of Brown Trout (Salmo trutta). Lipids 52, 61–71 (2017). https://doi.org/10.1007/s11745-016-4215-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11745-016-4215-0