
Abstract
This study aimed to understand bark thickness variations of Araucaria angustifolia (Bertol.) Kuntze trees growing in natural forest remnants in southern Brazil, and their relationship with quantitative and qualitative attributes. Bark thickness must be accurately estimated in order to determine timber volume stocks. This is an important variable for the sustainable management and conservation of araucaria forests. In spite of its importance and visibility, bark thickness variations have not been evaluated for this key species in southern Brazil. A total of 104 trees were selected, and their qualitative and quantitative attributes such as diameter at breast height (DBH), height (H), crown base height (CBH), crown length (CL), social position (SP), stoniness (ST), position on the relief (PR), vitality (VT) and branch arrangement (BA) were measured. The trees were categorized into two groups: red bark or gray bark. Regression analysis and artificial neural networks (ANN) were used for modelling bark thickness. The results indicate that: (1) bark thickness showed good correlation to DBH, with 0.76 as coefficient of determination (R2), 0.540 as Mean Absolute Error (MAE) and 22.4 root-mean-square error in percentage (RMSE%); (2) the trend changed according to bark colour, with significant differences for the intersection (\( \beta_{0} \) – Pr > F: p = 0.0124) and slope (\( \beta_{1} \) – Pr > F: p = 0.0126) of bark thickness curves between groups; (3) the highest correlation of bark thickness was found with: DBH (ρ = 0.88), H (ρ = 0.58), CBH (ρ = 0.46), SP (ρ = − 0.52), and BA (ρ = − 0.32); (4) modelling with ANN confirmed high adjustment (R2 = 0.99) and accuracy (RMSE% = 3.0) of the estimates. ANN is an efficient and robust technique for the modelling of various qualitative and quantitative attributes commonly used in forest mensuration. The effective use of ANN to estimate araucaria bark in natural forests reinforces its potential, besides the possibility of application for other forest species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Forest inventory is used to obtain information about the quantity, quality and condition of the resource components of a forest, particularly tree species, and other land characteristics (Kershaw et al. 2017). With the advancement of technology by industries that handle forest products and the growing demand for alternative products and energy sources, tree bark volume estimates have become relevant (Kozak and Yang 1981).
To provide accurate and timely information on current and future growing stock, forest managers need to estimate both volume and tree taper. Since inner bark volume and taper—required for accurately estimating bark volume—are based on the diameter measured for bark, bark thickness needs to be determined or predicted. As such, bark thickness is an important variable in forest mensuration (Muhairwe 2000), and in order to convert outside-bark diameters into inside-bark diameters, several equations for bark thickness and volume have been developed for many species worldwide (Li and Weiskittel 2011).
The bark is a vital woody component close to the surface but its characteristics and thickness have been only recently considered in the context of communities and biomes (Pausas 2015). The bark includes all tissues outside the cambium—the inner living phloem and dead outer tissues or rhytidome. The phloem plays an essential role translocating carbohydrates, while the periderm or outer bark tissue reduces water loss and provides protection from both biotic and abiotic injuries (Kozlowski and Pallardy 1996). Trees with a thick bark have vital tissue protection against attack from pathogens, herbivores, frost and drought. Thick bark characteristics are vital in large trees subjected to surface fires (Pausas 2015). However, bark thickness and texture vary significantly from extremely rough to soft according to the species (West 2015) and the environment (Zeibig-Kichas et al. 2016).
Regression models are commonly used to model forest attributes. However, artificial neural networks (ANN) have been applied in various fields of science due to their learning ability from using a dataset, and the generalization capability of this learning with unknown data has very accurate estimates (Haykin 2007). The use of ANN as an estimate method in forest science is promising for acquiring values for height (Binoti et al. 2013; Castaño-Santamaría et al. 2013), volume (Gorgens et al. 2009; Silva et al. 2009; Castro et al. 2013a; Binoti et al. 2014a; Leal et al. 2015), diameter distribution (Binoti et al. 2014b), bark (Diamantopoulou 2005, 2006), and relative diameter, as well as studies on stem shape (Leite et al. 2011; Soares et al. 2011; Souza 2013; Schikowski et al. 2015; Sanquetta et al. 2017), growth and production models at individual tree level (Castro et al. 2013b), and allows for the modelling of various qualitative and quantitative attributes simultaneously.
Araucaria (Araucaria angustifolia), also known as Brazilian pine, is an important tree species in southern Brazil of considerable economic importance due to its exceptional wood quality (Carvalho 1994). This species originally covered 200,000 km2 in the south of the country in early 1900s (Hueck 1972). Due to intensive and often indiscriminate harvesting, its area has been reduced significantly. However, it still has potential to be used sustainably, combined with strategic conservation (IUNC 2006). A pioneer study by Reitz and Klein (1966) revealed the natural occurrence of some Araucaria varieties whose major characteristics are related to the maturation of edible seeds that are palatable to both fauna and humans. However, due to annual climatic variations that determine maturation time, this differentiation is a challenging task. As an alternative, it is easy to categorize bark colour into red or gray while in the field (Fig. 1).

Illustrative bark colour characteristics: (a) gray, (b) red
To identify factors that influence changes associated with bark thickness and might serve as support tools for silvicultural activities, this study was developed with the following hypotheses: H1—there are differences in bark thickness of Brazilian pines according to their colour (Fig. 1); H2—qualitative variables of the crown and site are important to bark thickness; and, H3– ANN can estimate accurately bark thickness of Araucaria.
Given this context, this study aimed to model bark thickness for diameter at breast height (DBH) of araucaria growing in a natural forest in southern Brazil. The specific objectives were: (1) to characterize the in situ measured variables by qualitative and quantitative means; (2) to compare the bark thickness of gray- and red-coloured trees; (3) to develop a regression model of bark thickness in DBH function; (4) to verify the dendrometric, qualitative variables of crown, and local site variables associated with bark thickness; (5) to model the bark thickness using ANN; and, (6) to compare regression and ANN estimators.
Methods and materials
Characteristics of the species
Araucaria angustifolia (Bertol.) Kuntze belongs to the genus Araucaria, family Araucariaceae consisting of 16 species, exclusive to the southern hemisphere with two in South America. Other species are found in Oceania. It is an evergreen species and a heliophyte pioneer, adapted to intensive solar isolation, typical of high altitude regions, with rectilinear or cylindrical, rarely forked stems, containing four to eight verticils or whorls upon insertion of branches which stand out with age. As a dioecious plant—rarely monoeciousonly by trauma or disease— it has reproductive structures in male and female strobiles (Carvalho 2003).
Araucaria can reach heights of 20 to 50 m and up to two m in diameter. The crowns have cylindrical primary branches curved upwards, with the lower ones curved more than the upper ones; both branch types with secondary branches (grimpas), alternating and grouped at the apex (Reitz and Klein 1966). The trunk has thick, gray, rough, and deeply cracked bark, peeling off in rectangular plates and blades at the top of the trunk (Marchiori 2005). The species has a bark thickness between 5–10 cm (Ceccatto 1943), accounting for an average 32.6% of the total tree volume (Silva et al. 1975).
Study area
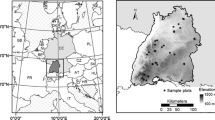
The study was conducted in a natural forest on a rural property in the municipality of Lages (27º48ʹS and 50º19ʹW), in the state of Santa Catarina (SC). The local climate is humid subtropical with no dry season and with a temperate summer (Cfb) (Alvares et al. 2013) according to the Köppen climate classification. The forest is predominantly Araucaria, with over 300 trees of this species per hectare, and with an average basal area of 21.9 m2 ha−1. Thus the local climate (Cfb) is favoured to this species in southern Brazil (Silveira et al. 2018).
Data collection
The attributes measured were based on 104 sample trees within the natural forest as a representation of population characteristics (Table 1). A detailed description of the field measurements can be found in Costa (2011).
Crowns were categorized into geometric formats: cone, umbel, hemisphere or champagne glass; these characteristics are caused by the ontogenetic alteration of the crowns, a characteristic of the species as described by Seitz (1986), and by competition with other trees (Costa and Finger 2017).
Statistical analysis
Descriptive statistics characterized the measured variables. A t test verified the existence of differences in mean between bark thickness measured in the north position and in the perpendicular measurement. The same test verified differences between the gray- and red-coloured bark groups. The following logarithmic model (Silva et al. 1975) was used to describe the relationship between bark thicknesses:
where \( B_{T} \) = Bark thickness; \( D_{\text{BH}} \) = Diameter at breast height; \( \beta_{0} \), \( \beta_{1} \) = estimated regression coefficients.
An analysis of covariance (ANCOVA) was used to verify differences between trends [intercept (\( \beta_{0} \)) and slope (\( \beta_{1} \))] of the adjusted regression lines (Milliken and Johnson 2002) for bark thickness for the tree groups between gray- and red-coloured bark.
Spearman’s rank correlation assessed the association between bark thickness and dendrometry, crown qualitative attributes, and site variables in addition to diameter. The variables with the highest correlation were also examined for their contribution to improve estimates of bark thickness. In addition to other advantages of using ANN, it is possible to model with categorical variables (qualitative) aside from numeric variables (quantitative) (Vieira et al. 2018).
The perceptron-type neural network from multiple layers (Haykin 2007) was architecture: n–n-1. Linear normalization of numeric variables was set between 0 and 1. The input variables were: DBH, H, HCB, SP, BA, Gr and BTpred; the continuous output variable was bark thickness. The group (Gr) variable expressed the differentiation between gray- (1) and red-barked trees (2). BTpred was the bark thickness estimate: BT = − 6.3248 + 2.4908∙ln (DBH), considering both bark colour groups (Table 3—All). In the hidden layer and in the output layer, the logistic activation and identity activation functions were applied.
To assess the performance of the adjusted models, the most appropriate being the one that presented the highest value of the coefficient of determination (R2) (Eq. 2), the lower mean absolute error (MAE) (Eq. 3) and root-mean-square error in percentage (RMSE%) (Eq. 4), and a regular and good waste distribution graphic. Statistical analyses used the statistical program R, version 3.5.2 (http://cran.r-project.org).
where R2 is coefficient of determination, MAE the mean absolute error, RMSE% the root-mean-square error in percentage, \( y_{\text{i }} \) the observed values, \(\hat{y}_{i}\) the estimated values; \(\bar{y}\) the average of the observed values, and n is number of observations.
Results
Qualitative and quantitative characteristics
The sample trees had a wide range of DBH, (17.0–117.8 for gray- and 14.7–79.3 for red-barked trees), and heights ranging from 11.8 to 32.9 for gray- and 9.1 to 24.9 for red-barked trees, and covered a wide variability of dendrometric characteristics such as CBH, CL, ST, PR and bark thickness (Table 2).
Trees were located in different positions throughout the site (‘1’ had 30 trees; ‘2’, 41; ‘3’, 17; ‘4’, 16), with different levels of stoniness (‘0’ = 55 trees; ‘1’ = 27; ‘2’ = 17; ‘3’ = 5). A similar trend was observed for these variables within the gray- and red- barked groups. The three social positions analyzed were also representative (SP1 = 79 trees; SP2 = 10; SP3 = 15) and the tree frequency according to the classification pertaining to the branch arrangement was: ‘1’ = 60 trees; ‘2’ = 31; ‘3’ = 13. No red-barked tree was class 3 of branch arrangement. The irregularities in the branch arrangement of dominant Brazilian pines in these quadrants are the result of competition and wind intensity, confirmed by fallen branches. No trees with low vitality were identified but 19 were classified as average vitality. This is especially related to the effect of intra-interspecific competition between individuals and the amount of light that reaches the understory.
Bark thickness variations between groups
After assessing the average bark thickness of the north position (2.96 cm) and its perpendicular E or W measurement (2.99 cm), according to the t test, there were no significant differences (t = − 0.14; p < 0.8867). There were also no differences between the gray- (2.94 cm) and red-coloured bark trees (3.02 cm) (t = − 0.31; p < 0.7557). These results reflect only the central value comparison between groups.
Adjustments and comparison of the regression model
The three adjustments of the logarithmic model, according to bark characteristics, demonstrated all regression coefficients to be significant (p < 0.0001; Table 3). The largest value of R2 and smallest MAE and RMSE% were for trees with gray bark.
Covariance analysis (F = 115.02; Pr > F: < 0.0001) showed significant differences for the intersection (\( \beta_{0} \) – Pr > F: p = 0.0124) and slope (\( \beta_{1} \) – Pr > F: p = 0.0126) of bark thickness curves between bark colour groups. Based on these results, it was necessary to use specific equations for each group. Graphic analysis of observed and estimated values showed that red bark trees smaller than 40.0 cm DBH have the thickest bark and, as such, they become the thinnest bark trees (Fig. 2). During data collection, we did not find red bark trees with DBH > 80.0 cm.

Observed and estimated values of bark thickness (BT) with diameter at breast height (DBH)
Variables related to bark thickness
Correlation analysis of bark thickness with qualitative and quantitative variables are shown in Table 4. The more representative variables were DBH (ρ = 0.88; p < 0.0001); H (ρ = 0.58; p < 0.0001); and CBH (ρ = 0.46; p < 0.0001), all with well-distributed points, similar to a straight line, followed by the qualitative variables SP (ρ = − 0.52; p < 0.0001); and BA (ρ = − 0.32; p = 0.0009). PR, CL, ST and VT were not significantly correlated (p > 0.01).
Modelling using artificial neural networks
The architectures of 3500 trained artificial neural networks are shown in Table 5. A total of 500 networks of each type (1–7) were trained, according to the evaluated input variables and differing number of neurons in the hidden layer (2–12), to obtain single bark thickness estimation (BT) in the output layer.
The number of trained networks in accordance with the input variables and neurons of the hidden layer ranged from a minimum of 41 to a maximum of 97 (Fig. 3a). Thus, mean values were calculated for statistics R2 (Fig. 3b), MAE (Fig. 3c) and RMSE% (Fig. 3d) according to each of the trained networks (1–7). With the inclusion of independent variables during network modelling (1–7), they showed, on average, increased adjustment and accuracy compared to the previous network architecture: DBH–H (ΔR2 ≈ 7.7% and ΔRMSE% ≈ − 5.2); DBH–H–CBH (ΔR2 ≈ 3.9% and ΔRMSE% ≈ − 3.4), DBH–H–CBH–SP (ΔR2 ≈ 1.3% and ΔRMSE% ≈ − 1.2), DBH–H–HBC–SP–BA (ΔR2 ≈ 3.2% and ΔRMSE% ≈ − 4.0), DBH–H–CBH–SP–BA–Gr (ΔR2 ≈ 0.8% and ΔRMSE% ≈ − 1.2), DBH–H–CBH–SP–BA–Gr–BTpred (ΔR2 ≈ 1.1% and ΔRMSE ≈ − 2.3), respectively, considering the largest number of neurons in the hidden layer (Fig. 3b–d).

Trained networks: a Amount of trained networks with different input variables and neurons in the hidden layers; Average of R2 values (b); Average MAE (c); and RMSE(%) (d) calculated for the trained networks with different input variables and number of neurons in the hidden layer to estimate the bark thickness (BT) of Brazilian pines. (Network types 1–7 considered the input variables DBH; DBH–H; DBH–H–CBH; DBH–H–CBH–SP; DBH–H–HBC–SP–BA; DBH–H–CBH–SP–BA–Gr; DBH–H–CBH–SP–BA–Gr–BTpred respectively. DBH Diameter at breast height, H Height, CBH Crown base height, SP Social position, BA Branch arrangement, Gr group, BTpred bark-thickness-regression model prediction)
Table 6 shows neural networks selected to describe bark thickness (BT) according to the variation of neurons in the hidden layer. In the input layer, the introduction of other independent variables increased network adjustment and accuracy criteria. Network type 7, which included the input variables: DBH–H–CBH–SP–BA–Gr–BTpred; showed better performance, with an increase in R2 from 0.924 to 0.997, whereas the RMSE% was reduced from 14.9 to 3.0 in accordance with the number of neurons obtained in the hidden layer.
The networks with the best performance were those that included 8 to 12 neurons in the hidden layer, represented, respectively, by the networks 138, 501, 1432, 1813, 2286, 2747 and 3157, for each variable inclusion. Figure 4 shows the predicted values versus the measured values for each of these networks.

Relationship between predicted bark thickness (BT) vs. observed values for the best-trained networks for each variable inclusion
Discussion
The purpose of this study was to develop models for predicting bark thickness at arbitrary diameter levels of Araucaria growing in natural forests. The development of a logarithmic regression proved that there is a strong correlation between bark thickness and stem diameter (Table 3). The same relationship has been investigated for other species such as Pinus radiata D. Don (Johnson and Wood 1987), Picea abies (L.) H. Karst. (Laasasenaho et al. 2005), Pinus patula Schiede ex Schltdl. & Cham. (Laar 2007), tropical rain forest species of French Guiana (Paine et al. 2010) and Quercus spp. (Schwilk et al. 2013). Different trends were found in bark thickness estimates between red bark and gray bark trees in our study (Fig. 2).
It is not yet known which factors influence each group that leads to such specific dendrological characteristics. Field observations show that gray- barked trees were older and had, on average, shorter crown lengths compared with red- barked trees (4.2 m and 5.9 m, respectively). It was initially hypothesized that the distinct bark colours were associated with the light intensity inside the forest (Reitz and Klein 1966). However, this hypothesis was rejected during the forest inventory, as Brazilian pines from the two groups were subjected to low, medium and high light incidence.
Dendrological features such as bark colour, size of pine nuts and maturation time have been reported in the literature and used to differentiate Araucaria varieties (Reitz and Klein 1966). In addition, in popular knowledge, the crown architecture differentiates between male and female pines. The dendrometrical and morphological characteristics of the species are reflections of its relationship with its environment, and should be analyzed for a better understanding of the factors that operate throughout its life.
As an attempt to improve bark thickness estimates, the contribution of other quantitative and qualitative variables as inputs beyond diameter are also investigated. In this context, among other advantages of using artificial neural networks, is the possibility of modelling with categorical variables (qualitative) such as sociological position (SP), branch arrangement (BA), and group (Gr), beyond numeric variables (quantitative) (Vieira et al. 2018).
Overall, classifications of qualitative attributes in natural forests should be assessed carefully and with defined standards to allow other researchers to follow the same procedures. Standardization of criteria is one of the greatest challenges in forest surveys when using attributes of this character. Barbosa (2018) described the current state of tree competition in natural forests by crown classification attributes, and proposed new metrics with different classification levels to describe characteristics related to the vertical crown form, the distance between pseudo-whorls, percentage of green leaves and crown size. This permitted a good explanation of Araucaria basal area growth.
Considering the data in Table 6, the Figs. 3a–d and the Fig. 4, it is clear that the use of ANN is a feasible technique to estimate bark thickness of Araucaria. There was more accuracy projecting bark thickness compared to the use of regression models. Moreover, an increase in predictive accuracy was also obtained using variables that consider both qualitative and quantitative information of tree and site. Other researchers have reported the influence of environmental factors on bark thickness such as fire regime (Van Mantgem and Schwartz 2003; Lawes et al. 2011), climate conditions (Pellegrini et al. 2017), and frost events (Molina et al. 2016) which are evidence of the importance of site variables, in addition to those intrinsic to the tree itself.
The last variable tested for inclusion in the artificial neural network was the regression model prediction itself (Type 7). The concept is to attribute applicability to previous regression models, for instance, under certain conditions, to allow the readjustment to current variations and to provide greater flexibility to the models. Finally, the possibility of variation in the number of neurons with the artificial neural networks has also resulted in prediction improvements to some extent, where it seems to stabilize (Fig. 3b, 3c and 3d). Approximately eight neurons, regardless of network type, were shown to be sufficient with a tendency to maintain both R2 and RMSE% constant. The continuous addition of neurons may result in overfitting and consequently, lower generalization power (Theodoridis and Koutroumbas 1999; Goodfellow et al. 2016).
Conclusions
There is evidence of statistically significant differences between the regression of gray- and red- barked trees in accordance with Araucaria DBH. This variable can be estimated for a wide range of diameter distributions using adjusted regression equations for both bark colour groups.
Qualitative variables are important in the modelling of several artificial intelligence techniques. Such variables can distinguish characteristics inherent to the data that would be difficult to distinguish with conventional regression modelling.
Artificial neural networks are an efficient and robust technique for various qualitative and quantitative attribute models commonly used in forest mensuration. For this reason, its accuracy, applicability and usability were confirmed in this study for modelling biometric variables in natural forests of high complexity and variability.
References
Alvares CA, Stape JL, Sentelhas PC, Gonçalves JLM, Sparovek G (2013) Köppen’s climate classification map for Brazil. Meteorol Z 2:711–728
Barbosa LO (2018) Efeito da competição no incremento em área transversal de Araucaria angustifolia (Bertol) Kuntze em Floresta Ombrófila Mista no sul do Brasil. Dissertation. Santa Maria: Universidade Federal de Santa Maria, p. 91
Binoti MLMS, Binoti DHB, Leite HG (2013) Aplicação de redes neurais artificiais para estimação da altura de povoamentos equiâneos de eucalipto. Árvore 37(4):639–645
Binoti MLMS, Binoti DHB, Leite HG, Garcia SLR, Ferreira MZ, Rode R, Silva AAL (2014a) Redes neurais artificiais para estimação do volume de árvores. Árvore 38(2):283–288
Binoti MLMS, Binoti DHB, Leite HG, Silva AAL, Pontes C (2014b) Utilização de redes neurais artificiais para a projeção da distribuição diamétrica de povoamento equiâneos. Árvore 38(4):747–754
Carvalho PER (1994) Espécies florestais brasileiras: recomendações silviculturais, potencialidades e usos da madeira. EMBRAPA-CNPF, Colombo, p 640
Carvalho PER (2003) Espécies florestais brasileiras. EMBRAPA-CNPF, Colombo/EMBRAPA-SPI, Brasília, p 1039
Castaño-Santamaría J, Crecente-Campo F, Fernández-Martinez JL, Barrio-Anta M, Obeso JR (2013) Tree height prediction approaches for uneven-aged beech forests in northwestern Spain. For Ecol Manag 307:63–73
Castro RVO, Soares CPB, Martins FB, Leite HG (2013a) Crescimento e produção de plantios comerciais de eucalipto estimados por duas categorias de modelos. Pesq Agropec Bras 48(3):287–295
Castro RVO, Soares CPB, Leite HG, De Souza AL, Nogueira GS, Martins FB (2013b) Individual growth model for Eucalyptus stands in Brazil using Artificial Neural Network. ISRN Forestry 2013:1–12
Ceccatto GNO (1943) Pinho Brasileiro. Ministério da Agricultura, Serviço de Informação Agrícola, Rio de Janeiro, p 39
Costa EA.(2011). Influência de variáveis dendrométricas e morfométricas da copa no incremento periódico de Araucaria angustifolia (Bertol.) Kuntze, Lages, SC. Dissertation. Santa Maria: Universidade Federal de Santa Maria, p 140
Costa EA, Finger CAG (2017) Efeito da Competição nas Relações Dimensionais de Araucária. Floresta e Ambiente 24:e20150145
Diamantopoulou MJ (2005) Artificial neural networks as an alternative tool in pine bark volume estimation. Comput Electron Agric 48:235–244. https://doi.org/10.1016/j.compag.2005.04.002
Diamantopoulou MJ (2006) Tree-bole volume estimation on standing pine trees using cascade correlation artificial neural network models. CIGR J 8:1–14
Goodfellow I, Bengio Y, Courville A (2016) Deep Learning. The MIT Press, Cambridge, p 800
Gorgens EB, Leite HG, Gleriani JM, Soares CPB, Ceolin A (2009) Estimação do volume de árvores utilizando redes neurais artificiais. Árvore 33(6):1141–1147
Haykin S (2007) Redes neurais: princípios e prática, 2nd edn. Bookman, Porto Alegre, p 900
Hueck K (1972) As florestas da América do Sul. Ed. da UnB, Brasília/Polígono, São Paulo, p. 466
IUCN (2006) IUCN red list of threatened species. http://www.iucnredlist.org. Accessed 5 Dec 2006
Johnson TS, Wood GB (1987) Simple linear model reliably predicts bark thickness of Radiata pine in the Australian Capital Territory. For Ecol Manag 22:173–183
Kershaw JA, Ducey MJ, Beers TW, Husch B (2017) Forest Mensuration, 5th edn. Wiley, Hoboken, p 630
Kozak A, Yang RC (1981) Equations for estimating bark volume and thickness of commercial trees in British Columbia. Forest Chron 57(3):112–115
Kozlowski TT, Pallardy SG (1996) Physiology of woody plants, 2nd edn. Academic Press, San Diego, p 411
Laar A (2007) Bark thickness and bark volume of Pinus patula in South Africa. South Hemisphere Forest J 69(3):165–168
Laasasenaho J, Melkas T, Aldén S (2005) Modelling bark thickness of Picea abies with taper curves. For Ecol Manag 206:35–47
Lawes MJ, Richards A, Dathe J, Midgley J (2011) Bark thickness determines fire resistance of selected tree species from fire-prone tropical savanna in north Australia. Plant Ecol 212(12):2057–2069
Leal FA, Miguel EP, Matricardi EAT, Pereira RS (2015) Redes neurais artificiais na estimativa de volume em um plantio de eucalipto em função de fotografias hemisféricas e número de árvores. Rev Bras Biometr 33(2):233–249
Leite HG, Silva MLM, Binoti DHB, Fardin L, Takiza WFH (2011) Estimation of inside-bark diameter and heartwood diameter for Tectona grandis Linn. trees using artificial neural networks. Eur J Forest Res 130(2):263–269
Li R, Weiskittel AR (2011) Estimating and predicting bark thickness for seven conifer species in the Acadian Region of North America using a mixed effects modeling approach: comparison of model forms and subsampling strategies. Eur J Forest Res 130(2):219–233
Marchiori JNC (2005) Dendrologia das gimnospermas, 2nd edn. UFSM, Santa Maria, p 161
Milliken GA, Johnson FE (2002) Analysis of messy data volume III: analysis of covariance. Chapman e Hall, Boca Raton
Molina JGA, Hadad MA, Domínguez DP, Roig FA (2016) Tree age and bark thickness as traits linked to frost ring probability on Araucaria araucana trees in northern Patagonia. Dendrochronologia 37:116–125
Muhairwe CK (2000) Bark thickness equations for five commercial tree species in regrowth forests of Northern New South Wales. Aust Forest 63(1):34–43
Paine CET, Stahl C, Courtois EA, Patiño S, Sarmiento C, Baraloto C (2010) Functional explanations for variation in bark thickness in tropical rain forest trees. Funct Ecol 24:1202–1210
Pausas JG (2015) Bark thickness and fire regime. Funct Ecol 29:315–327
Pellegrini AFA, Anderegg WRL, Paine T, Hoffmann WA, Kartzinel T, Rabin SS, Sheil D, Franco AC, Pacala SW (2017) Convergence of bark investment according to fire and climate structures ecosystem vulnerability to future change. Ecol Lett 20:307–316
Reitz R, Klein RM (1966) Flora ilustrada catarinense Araucariáceas. Herbário Barbosa Rodrigues, Itaja, p 63
Sanquetta CR, Piva LRO, Wojciechowski J, Corte APD, Schikowski AB (2017) Volume estimation of Cryptomeria japonica logs in southern Brazil using artificial intelligence models. South For 80:29–36
Schikowski AB, Corte APD, Sanqueta CR (2015) Estudo da forma do fuste utilizando redes neurais artificiais e funções de afilamento. Pesqui Florest Bras 35(82):119–127
Schwilk DW, Gaetani MS, Poulos HM (2013) Oak bark allometry and fire survival strategies in the Chihuahuan Desert Sky Islands, Texas, USA. PLoS ONE 8(11):e79285
Seitz RA (1986) Crown development of Araucaria angustifolia in its natural environment during sixty years. In: Fujimori T, Whitehead D (eds) Crown and canopy structure in relation to productivity. Ibaraki, Forestry and Forest Products Research Institute, pp 129–146
Silva JA, Estefanel V, Andrae F (1975) Avaliação da dupla espessura de casca em árvores individuais de pinheiro brasileiro, Araucaria angustifolia (Bert.) O.Ktze, referente ao nível do DAP. Ciência Rural 5(1):17–34
Silva MLM, Binoti DHB, Gleriani JM, Leite HG (2009) Ajuste do modelo de Schumacher e Hall e aplicação de redes neurais artificiais para estimar volume de árvores de eucalipto. Árvore 33(6):1133–1139
Silveira AC, Hess AF, Schorr LPB, Krefta SM, Santos DV, Filho MDHV, Atanazio KA, Costa EA, Stepka TF, Borsoi GA (2018) Management of Brazilian pine (Araucaria angustifolia (Bertol) Kuntze) based on the Liocourt model in a mixed Ombrophilous forest in Southern Brazil. Aust J Crop Sci 12(02):311–317
Soares FA, Flôres EL, Cabacinha CD, Carrijo GA, Veiga ACP (2011) Recursive diameter prediction and volume calculation of Eucalyptus trees using multilayer perceptron networks. Comput Electron Agric 78(1):19–27
Souza RR (2013) Estudo da forma do fuste de árvores de eucalipto em diferentes espaçamentos. Dissertation. Diamantina: Universidade Federal dos Vales do Jequitinhonha e Mucuri, p. 86
Theodoridis S, Koutroumbas K (1999) Pattern Recognition, 1st edn. Academic Press, San Diego, p 625
Van Mantgem P, Schwartz M (2003) Bark heat resistance of small trees in Californian mixed conifer forests: testing some model assumptions. For Ecol Manag 178(3):341–352
Vieira GC, Mendonça AR, Silva GF, Zanetti SS, Silva MM, Santos AR (2018) Prognoses of diameter and height of trees of eucalyptus using artificial intelligence. Sci Total Environ 619–620:1473–1481
West PW (2015) Tree and forest measurement, 3rd edn. Springer, Cham, p 214
Zeibig-Kichas NE, Ardis CW, Berrill JP, King JP (2016) Bark thickness equations for mixed-conifer forest type in Klamath and Sierra Nevada mountains of California. Int J Forest Res 2016:1–10
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Corresponding editor: Yu Lei.
Project funding
This study is supported by the Graduate Program in Forest Engineering of the Santa Catarina State University (UDESC), the Santa Catarina Research Foundation (FAPESC; 2017TR1762, 2017TR639, 2019TR816), the Brazilian National Council for Scientific and Technological Development (CNPq; 313887/2018-7), and the Coordination for the Improvement of Higher Education Personnel (CAPES).
The online version is available at http://www.springerlink.com.
Rights and permissions
About this article

Cite this article
Costa, E.A., Liesenberg, V., Finger, C.A.G. et al. Understanding bark thickness variations for Araucaria angustifolia in southern Brazil. J. For. Res. 32, 1077–1087 (2021). https://doi.org/10.1007/s11676-020-01163-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-020-01163-1