
Abstract
The predominant feature of type 2 diabetes is insulin resistance. Identifying a drug able to reduce insulin resistance is an urgent requirement. ent-3α-Formylabieta-8(14),13(15)-dien-16,12β-olide had been identified as a new diterpene derivative which showed anticancer activity. This study explores the hypoglycemic effect of ent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide and studied its mechanism. The insulin response of HepG2 cells following ent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide treatment, as a model for liver cancer cells, was assessed. The results demonstrated that hyperglycemia resulted in a significant increase in the levels of insulin receptor substrate-1 (IRS-1) serine phosphorylation and decrease in Akt phosphorylation. High glucose also inhibited the phosphorylation of insulin-dependent GSK3β. ent-3α-Formylabieta-8(14),13(15)-dien-16,12β-olide treatment improved the effect of insulin on the phosphorylation of IRS-1 Ser307. In addition, this study demonstrated that the effect of ent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide was dependent on the activation of AMP-activated protein kinase. Collectively, experimental data indicated an association between insulin resistance and hyperglycemia in HepG2 cells, and that ent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide reduces IRS-1 Ser307 phosphorylation via activating AMPK, thereby decreasing the insulin signaling blockade.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Circulating blood glucose is a serious problem for patients with type 2 diabetes [1, 2]. Chronic hyperglycemia can induce insulin resistance and impair insulin secretion. Also, persistent high blood glucose is toxic to macrovascular and microvascular systems and this effect is known as glucose toxicity [3,4,5]. Among a number of pathological factors, one reason for hyperglycemia is the impairment of glucose homeostasis and hepatic control [6]. Hepatic insulin resistance causes a reduced capacity to trigger downstream signaling cascades. Furthermore, hepatic insulin resistance affects glucose levels and causes lipid synthesis disorders, which may worsen systemic insulin resistance and fatty liver disease [7, 8]. Previous studies have identified that insulin receptor substrate (IRS), a docking protein activated by the insulin receptor (IR), may control hepatic insulin resistance [9,10,11,12]. This defect is likely to be due to the IRS-1 serine phosphorylation stimulated by insulin resulting in decreased phosphoinositide 3-kinase (PI3K) activity [13, 14]. IRS-2 and IRS-1 possess complementary functions in hepatic metabolism, whereas IRS-1 is particularly relevant to glucose homeostasis [15, 16].
When extracellular insulin binds the IR, several intracellular protein substrates including IRS-1 and IRS-2 are stimulated, and downstream insulin signaling cascades are initiated [17,18,19]. This activates the PI3K pathway and suppresses glycogen synthase kinase-3 (GSK-3) [20, 21]. In addition, the forkhead box O1 transcription factor can be phosphorylated by Akt to reduce the expression of glucose 6-phosphatase (G6Pase) and inhibit gluconeogenesis [22,23,24]. AMP-activated protein kinase (AMPK) is an important regulator of cell metabolism that inhibits liver glucose isogenesis. Many drugs (such as berberine and metformin) used in the treatment of type 2 diabetes can activate AMPK [25,26,27].
Euphorbia lunulata Bge, one of the species of Euphorbia L., is a perennial herb. Euphorbia L. has a wide variety and is distributed all over the world. Many species have a long history of medicinal history at home and abroad. Modern research indicates that the main active ingredients of this genus are terpenoids and some flavonoids [28]. In China, Euphorbia lunulata Bge is mainly distributed in Inner Mongolia, Shandong, Jiangsu and Hebei provinces. It has been mainly used in folk medicine to treat diseases such as asthma, gastric carcinoma and breast carcinoma [29]. According to the previous literatures on Euphorbia L. plants, diterpenoids were proved to be the active constituents and were found to have anti-tumor [30], antimicrobial [31] as well as anti-virus activities [32]. However, domestic and foreign research on Euphorbia lunulata Bge mainly focused on flavonoids [33, 34], and little research on terpenoids which may have strong pharmacological activity in Euphorbia L. A new ent-abietane-type diterpene derivative ent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide (EFLDO) and other four jatrophane-type diterpenes were isolated from Euphorbia lunulate Bge in our previous study [35]. And then, ent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide (EFLDO) was further extracted and enriched to obtain 55 mg/20.0 kg in our laboratory. The preliminary biological activities for five diterpenes against NCI-H460 and MCF-7 Hela tumor cell lines were evaluated and results indicated marked activity for EFLDO against the two cell lines (NCI-H460 IC50 = 19.5 μM; MCF-7 IC50 = 18.6 μM). In addition, other four diterpenes exhibited moderate cytotoxic activities for two cell lines with IC50 values ranging from 32.1 to 58.2 μM [35]. ent-Abietane-type diterpenes are the most abundant diterpene type of this species. In view of the strong biological activity of EFLDO (ent-abietane-type diterpene) and related literatures that reported ent-abietane-type diterpenes have hypoglycemic effects with different mechanisms of action [36,37,38], HepG2 cell line [39] is selected as a model in this study and used to study the hypoglycemic effect and mechanism of EFLDO on diabetes.
This study examined the effects of EFLDO on hyperglycemia-induced insulin signal transduction pathways. Results showed that the hyperglycemia-induced phosphorylation of IRS-1 Ser307 suppressed the activation of Akt, whereas treatment with EFLDO reduced the insulin signal blockade by improving the function of IRS-1. In addition, the results show that the effect of EFLDO is dependent on the phosphorylation of AMPK, demonstrating novel antidiabetic activity of EFLDO.
Materials and methods
Materials
ent-3α-Formylabieta-8(14),13(15)-dien-16,12β-olide (EFLDO) was extracted from the Euphorbia lunulata Bge in this lab. Dorsomorphin (Compound C, an AMPK-specific inhibitor), d-glucose and recombinant human insulin were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). AMPK, p-AMPK, glycogen synthase kinase-3β (GSK-3β), β-actin and phosphorylated (p)-GSK3β (Ser9) antibodies were from purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA) and control siRNA and siRNA against AMPK were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA; catalog No., sc-29673). IRS-1 and p-IRS-1 antibodies were purchased from BD Transduction Laboratories (Lexington, KY, USA). Akt and p-Akt antibodies (Ser473) were purchased from Ampersand Bioscience, LLC (Saranac Lake Village, NY, USA).
Methods
Cell culture
HepG2 cells were maintained in this lab and cultured in DMEM supplemented with 10% fetal bovine serum, penicillin and streptomycin (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA), and maintained in an atmosphere containing 5% carbon dioxide at 37 ℃. The cells were further cultured in serum-free medium for 24 h for further procedures.
Immunoprecipitation
Cells were lysed in RIPA buffer [1% Triton X-100, 300 mM NaCl, Tris–HCl (pH 7.4; 20 mM), 0.4 mM sodium vanadate] and centrifuged at 13,000 rpm for 15 min. Protein A/G beads were agitated gently with the IRS-1 antibody and separated by centrifugation for 30 min at 5000 rpm and 4 ℃. The supernatant (500 µg total protein) was incubated with the beads and gently agitated at 4 ℃. The beads were washed three times in cold RIPA buffer for 10 min to elute, boiled in SDS buffer at 100 ℃, and the supernatant was used for subsequent western blot analysis.
Western blot analysis
The cells were co-treated with glucose and/or 1, 5, or 10 μM EFLDO for 24 h, then treated with 100 nM insulin for 10 min, and lysed. Total protein (50 μg) was separated by 10% SDS-PAGE and transferred to a PVDF membrane (GE Healthcare, Chicago, IL, USA). The membrane was blocked with 5% BSA for 1 h at room temperature. Then, the membrane was incubated with primary antibodies overnight, followed by the application of horseradish peroxidase-conjugated secondary antibodies for 2 h at room temperature. Visualization of the immune complex was performed with a chemiluminescence kit (GE Healthcare).
RNAi knockdown of AMPK
HepG2 cells were transfected with the siRNA against AMPK or control siRNA, using a siRNA transfection reagent (Santa Cruz Biotechnology, Inc.), according to the manufacturer’s protocol. The expression of AMPK after 30 h was analyzed via western blotting.
Results
The effect of high glucose on the tyrosine phosphorylation of IRS-1
The aim was to examine the role of hyperglycemia in the insulin pathway in the liver and to examine the phosphorylation state of IRS-1, as its phosphorylation is essential for insulin sensitivity [26]. The effect of hyperglycemia on the insulin-induced phosphorylation of IRS-1 was assessed. HepG2 cells were treated with different concentrations of glucose for 24 h, and then 100 nM insulin was added for 10 min. Insulin induces the tyrosine phosphorylation of IRS-1, whereas chronic hyperglycemia induces serine phosphorylation. As shown in Fig. 1a, the insulin-induced IRS-1 tyrosine phosphorylation was observed at higher concentrations of glucose (20 μM). However, insulin-induced IRS-1 tyrosine phosphorylation was significantly downregulated by increasing the glucose concentration to 60 μM, and the effect was dose-dependent. Analysis of the effects of hyperglycemia over time, as shown in Fig. 1b, demonstrated that the tyrosine phosphorylation of IRS-1 was evidently reduced by 33 mM glucose treatment at 12 h. Changes in the level of total IRS-1 protein were also observed after treatment, indicating that the degradation of IRS-1 was not induced.

Phosphorylation of IRS-1 induced by high glucose and insulin stimulation. a HepG2 cells were treated with the specified concentrations of d-glucose for 24 h, followed by the addition of 100 nM insulin for 10 min. b The phosphorylation of IRS-1 was detected over time in cells treated with 33 mM d-glucose. c The Akt phosphorylation levels were determined in cells treated with increasing concentrations of d-glucose for 24 h, followed by the addition of 100 nM insulin for 10 min. IRS-1 insulin receptor substrate-1, IP immunoprecipitation, IB immunoblot, p- phosphorylated
Akt is a key molecule involved in mediating the effect of insulin metabolism on downstream signaling pathways. To determine whether the insulin stimulation induced the Akt cascade, the Ser473 phosphorylation of Akt was evaluated in HepG2 cell lysates. Compared with the absence of insulin, the addition of insulin increased the phosphorylation of Akt, as shown in Fig. 1c, whereas Akt phosphorylation decreased by increasing glucose concentration (40 μM). This suggested that insulin induced the Akt cascade, while hyperglycemia decreased the effect.
Effects of EFLDO treatment on Akt phosphorylation in the presence of high glucose level
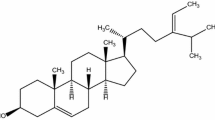
To evaluate the effect of EFLDO (Fig. 2a) on the insulin-induced phosphorylation of Akt, HepG2 cells were treated with 33 mM glucose for 24 h and then 100 nM insulin for 10 min, as shown in Fig. 2b, so different concentrations of EFLDO treatment reversed insulin-induced Akt Ser473 phosphorylation in a dose-dependent manner. Then, it was assessed whether the increase in insulin sensitivity following EFLDO treatment was due to changes in the phosphorylation of IRS-1. Using the same method-treated cells, the results demonstrated that the tyrosine phosphorylation of IRS-1 was recovered by EFLDO treatment in a dose-dependent manner (Fig. 2c). These results indicated that hyperglycemia induced IRS-1 inactivation in HepG2 cells by promoting serine phosphorylation and inhibiting tyrosine phosphorylation, whereas EFLDO reversed the effect on IRS-1 phosphorylation and enhanced the insulin-stimulated phosphorylation of Akt at Ser473.

Combined action of insulin stimulation, EFLDO treatment and high glucose. a The chemical structure of EFLDO. b Dose-dependent effect of EFLDO treatment on the phosphorylation of Akt in the presence of high glucose and 100 nM insulin. c The IRS-1 Ser307 phosphorylation was detected in HepG2 cells (processed with 33 mM glucose for 24 h and 100 nM insulin for 10 min) treated with different concentrations of EFLDO. EFLDOent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide, IRS-1 insulin receptor substrate-1, p phosphorylated, IP immunoprecipitation, IB immunoblot
EFLDO induces AMPK phosphorylation in the presence of high glucose level
AMPK activation is considered to be the central event of cell energy metabolism. In insulin-resistant HepG2 cells, the antidiabetic effects of certain clinical drugs are dependent on AMPK [25,26,27]. The phosphorylation of AMPK in HepG2 cells in hyperglycemic conditions following EFLDO treatment was examined. The level of AMPK phosphorylation in HepG2 cells was relatively low under high glucose conditions, but these levels increased in HepG2 cells co-treated with EFLDO in a dose-dependent manner (Fig. 3a). It has been established that the phosphorylation of Ser789 on IRS-1 leads to the activation of AMPK and an increase in PI3K activity [13]. The relationship between the inhibition of IRS-1 serine phosphorylation and the phosphorylation of AMPK was examined. The RNAi method was used to knock down the expression of AMPK in HepG2 cells. It was demonstrated that AMPK RNAi successfully reduced the expression of AMPK compared with cells transfected with a control (Fig. 3b). As afore described, in the control cells, EFLDO treatment reduced the phosphorylation of IRS-1 Ser307; however, subsequent to RNAi transfection, IRS-1 Ser307 phosphorylation remained high following EFLDO treatment (Fig. 3c). These results suggest that the activation of AMPK by EFLDO treatment may serve an important role in the protective effect of EFLDO under hyperglycemic conditions.

AMPK phosphorylation in HepG2 cells induced by EFLDO treatment in the presence of high glucose. a EFLDO treatment increased AMPK Thr172 phosphorylation in a dose-dependent manner in the presence of high glucose. b Verification of the effect of AMPK siRNA. AMPK expression was reduced in the siRNA-transfected group, compared with in the control groups. c IRS-1 Ser307 and AMPK Thr172 phosphorylation following treatment with 33 mM glucose and EFLDO. AMPK AMP-activated protein kinase, EFLDOent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide, IRS-1 insulin receptor substrate-1, p phosphorylated, IP immunoprecipitation, IB immunoblot
The metabolic effects of EFLDO are dependent on AMPK phosphorylation
It was previously demonstrated that insulin serves an important role in the regulation of glycogen synthesis [40]. As GSK-3β serves a leading role in liver glycogen synthesis, this study investigated the relationship between EFLDO treatment and GSK-3β activity. Hyperglycemia increased the expression of GSK-3β activity in HepG2 cells through inhibiting the phosphorylation of GSK-3β Ser9 (Fig. 4a). This study observed that EFLDO treatment reversed the effect on the GSK-3β serine phosphorylation level in a dose-dependent manner. Treated with LY294002, a PI3K-specific inhibitor inhibiting the activation of Akt by insulin without affecting IRS-1 phosphorylation was also observed. This indicated that the effect of EFLDO treatment may be mediated by tyrosine phosphorylation in the IRS–PI3K–Akt signaling pathway (Fig. 4b). Compound C, an AMPK-specific inhibitor, reduced the IRS-1 Ser307 phosphorylation induction by EFLDO, suggesting that the action of EFLDO may depend on the activation of AMPK. Then, AMPK siRNA knockdown was used to measure the insulin response in the cells. EFLDO treatment increased the phosphorylation of AMPK, whereas the effect of insulin on GSK3β was suspended in the AMPK knockdown group (Fig. 4c). These results demonstrate that the activation of AMPK may serve a major role in the phosphorylation of IRS-1 Ser307.

EFLDO enhanced the insulin-stimulated GSK3β activation in HepG2 cells. a The GSK3β Ser9 phosphorylation in HepG2 cells was detected in cells treated with increasing concentrations of EFLDO. b The IRS-1 Ser307 phosphorylation level in HepG2 cells was detected in cells treated with increasing concentrations of EFLDO. c IRS-1 Ser307 phosphorylation, AMPK Thr172 phosphorylation and GSK3β Ser9 were detected in the presence of high glucose, EFLDO treatment and with AMPK siRNA. EFLDOent-3α-formylabieta-8(14),13(15)-dien-16,12β-olide, GSK3β glycogen synthase kinase 3β, IRS-1 insulin receptor substrate-1, AMPK AMP-activated protein kinase, IB immunoblot
Discussion
Irregular insulin signaling pathways have been reported in diabetes and several diabetic animal models [14]. This study examined the insulin signal response in hyperglycemic conditions following EFLDO treatment. The results suggest that EFLDO has insulin-sensitizing activity. It has shown that exposure to increased glucose results in IRS-1 serine phosphorylation. This is in accordance with a previous study [11]. In addition, EFLDO treatment markedly reversed the effect of hyperglycemia. The effect of EFLDO was dose-dependent. Finally, it was shown that the inhibition of AMPK activation prevents the inhibition of IRS-1 Ser307 phosphorylation by EFLDO. Therefore, EFLDO may stimulate the insulin sensitivity of liver cells through multiple signal transduction pathways. These data indicate a novel molecular mechanism for EFLDO to reduce insulin resistance under hyperglycemic conditions.
High IRS-1 serine phosphorylation leads to insulin-induced blockade of IRS-1 tyrosine phosphorylation. It has been reported that serine phosphorylation of IRS-1 is critical in the development of insulin resistance. In the present study, we found that EFLDO treatment significantly reduced hyperglycemia-induced phosphorylation of IRS-1 Ser307. Previous studies have shown that IRS-1 is a target of c-Jun N-terminal kinase (JNK) and protein kinase C (PKC); hyperglycemia may also activate JNK and PKC. The activation of PKC increases the phosphorylation of IRS-1 Ser307, which blocks the activation of the downstream Akt signaling pathway.
Another observation in the present study was the inhibitory effect of EFLDO on AMPK-mediated IRS-1 serine phosphorylation. In the current research, EFLDO was demonstrated to inhibit the phosphorylation of IRS-1 Ser307, whereas this effect could be blocked by inhibiting AMPK. We further identified that EFLDO activates AMPK. AMPK can be activated by multiple signaling pathways, including those of calcium/calmodulin-dependent protein kinase (CaMKK) and serine/threonine kinase 11 (LKB1) [27].
References
Qi X, Li L, Yang G, Liu J, Li K, Tang Y, Liou H, Boden G (2007) Circulating obestatin levels in normal subjects and in patients with impaired glucose regulation and type 2 diabetes mellitus. Clin Endocrinol 66:593–597
Earle KE, Archer AG, Baillie JE (1989) Circulating and excreted levels of chromium after an oral glucose challenge: influence of body mass index, hypoglycemic drugs, and presence and absence of diabetes mellitus. Am J Clin Nutr 49:685–689
Del Prato S (2009) Role of glucotoxicity and lipotoxicity in the pathophysiology of Type 2 diabetes mellitus and emerging treatment strategies. Diabet Med J Br Diabet Assoc 26:1185–1192
Bedard K, Strecko J, Theriault K, Bedard J, Veyrat-Durebex C, Gaudreau P (2008) Effects of a high-glucose environment on the pituitary growth hormone-releasing hormone receptor: type 1 diabetes compared with in vitro glucotoxicity. Am J Physiol Endocrinol Metab 294:E740–751
Lindmark S, Buren J, Eriksson JW (2006) Insulin resistance, endocrine function and adipokines in type 2 diabetes patients at different glycaemic levels: potential impact for glucotoxicity in vivo. Clin Endocrinol 65:301–309
Ahmed F, Waslien C, Al-Sumaie MA, Prakash P, Allafi A (2013) Trends and risk factors of hyperglycemia and diabetes among Kuwaiti adults: National Nutrition Surveillance Data from 2002 to 2009. BMC Public Health 13:103
Tiano JP, Delghingaro-Augusto V, Le May C, Liu S, Kaw MK, Khuder SS, Latour MG, Bhatt SA, Korach KS, Najjar SM, Prentki M, Mauvais-Jarvis F (2011) Estrogen receptor activation reduces lipid synthesis in pancreatic islets and prevents beta cell failure in rodent models of type 2 diabetes. J Clin Investig 121:3331–3342
Kanat M, Serin E, Tunckale A, Yildiz O, Sahin S, Bolayirli M, Arinc H, Dirican A, Karagoz Y, Altuntas Y, Celebi H, Oguz A (2009) A multi-center, open label, crossover designed prospective study evaluating the effects of lipid lowering treatment on steroid synthesis in patients with Type 2 diabetes (MODEST Study). J Endocrinol Invest 32:852–856
Jiang F, Li S, Pan L, Jia C (2015) Association of the G1057D polymorphism in insulin receptor substrate 2 gene with type 2 diabetes mellitus: a meta-analysis. J Diabet Complicat 29:731–736
Xu J, Lin S, Yin J (2015) Effect of gastric bypass operation on expressions of adipic insulin receptor and insulin receptor substrate-1 in rats with type 2 diabetes mellitus. Zhonghua Wei Chang Wai Ke Za Zhi 18:65–68
Zhang Y, Sun CM, Hu XQ, Zhao Y (2014) Relationship between melatonin receptor 1B and insulin receptor substrate 1 polymorphisms with gestational diabetes mellitus: a systematic review and meta-analysis. Sci Rep 4:6113
Oliveira JM, Rebuffat SA, Gasa R, Gomis R (2014) Targeting type 2 diabetes: lessons from a knockout model of insulin receptor substrate 2. Can J Physiol Pharm 92:613–620
De Blasio MJ, Huynh K, Qin C, Rosli S, Kiriazis H, Ayer A, Cemerlang N, Stocker R, Du XJ, McMullen JR, Ritchie RH (2015) Therapeutic targeting of oxidative stress with coenzyme Q10 counteracts exaggerated diabetic cardiomyopathy in a mouse model of diabetes with diminished PI3K(p110alpha) signaling. Free Radical Biol Med 87:137–147
Qi Z, Xu Y, Liang Z, Li S, Wang J, Wei Y, Dong B (2015) Baicalein alters PI3K/Akt/GSK3beta signaling pathway in rats with diabetes-associated cognitive deficits. Int J Clin Exp Med 8:1993–2000
Andrade Ferreira I, Akkerman JW (2005) IRS-1 and vascular complications in diabetes mellitus. Vitam Horm 70:25–67
Aytug S, Reich D, Sapiro LE, Bernstein D, Begum N (2003) Impaired IRS-1/PI3-kinase signaling in patients with HCV: a mechanism for increased prevalence of type 2 diabetes. Hepatology 38:1384–1392
Gelaleti RB, Damasceno DC, Salvadori DM, Marcondes JP, Lima PH, Morceli G, Calderon IM, Rudge MV (2015) IRS-1 gene polymorphism and DNA damage in pregnant women with diabetes or mild gestational hyperglycemia. Diabetol Metab Syndr 7:30
Alharbi KK, Khan IA, Abotalib Z, Al-Hakeem MM (2014) Insulin receptor substrate-1 (IRS-1) Gly927Arg: correlation with gestational diabetes mellitus in Saudi women. Biomed Res Int 2014:146495
Fatchiyah F, Christian N, Soeatmadji D (2013) Reducing IRS-1 activation cause mutation of tyrosine kinase domain hINSR gene on type-2 diabetes mellitus patients. Bioinformation 9:853–857
Liu H, Ou S, Xiao X, Zhu Y, Zhou S (2015) Diabetes worsens ischemia-reperfusion brain injury in rats through GSK-3beta. AM J Med Sci 350:204–211
Liu W, Hao J, Zhu L, Li F, Liu Q, Liu S, Zhao S, Li H, Duan H (2013) Phospho-GSK-3beta is involved in the high-glucose-mediated lipid deposition in renal tubular cells in diabetes. Int J Biochem Cell Biol 45:2066–2075
Kim YK, Lee GS, Jung EM, Hyun SH, Hwang WS, Jeung EB (2012) Generation of fibroblasts overexpressing liver-specific PEPCK in a miniature pig model of human type 2 diabetes mellitus. Mol Med Rep 6:45–50
Samuel VT, Beddow SA, Iwasaki T, Zhang XM, Chu X, Still CD, Gerhard GS, Shulman GI (2009) Fasting hyperglycemia is not associated with increased expression of PEPCK or G6Pc in patients with Type 2 Diabetes. P Natl Acad Sci USA 106:12121–12126
Cadoudal T, Fouque F, Benelli C, Forest C (2008) Glyceroneogenesis and PEPCK-C: pharmacological targets in type 2 diabetes. Med Sci 24:407–413
Ropelle ER, Pauli JR, Fernandes MF, Rocco SA et al (2016) Expression of concern. A central role for neuronal AMP-activated protein kinase (AMPK) and mammalian target of rapamycin (mTOR) in high-protein diet-induced weight loss. Diabetes 65:1122–1123
Wall CE, Yu RT, Atkins AR, Downes M, Evans RM (2016) Nuclear receptors and AMPK: can exercise mimetics cure diabetes? J Mol Endocrinol 57:R49–58
Kjobsted R, Pedersen AJ, Hingst JR, Sabaratnam R, Birk JB, Kristensen JM, Hojlund K, Wojtaszewski JF (2016) Intact regulation of the AMPK signaling network in response to exercise and insulin in skeletal muscle of male patients with type 2 diabetes: illumination of AMPK activation in recovery from exercise. Diabetes 65:1219–1230
Ding AW, Zhang L (2008) Progress on research of the Euphorbia L. Chin J Tradit Chin Med 26(11):2433–2435
Shi HM, Williams ID, Sung HH, Zhu HX, Ip NY, Min ZD (2005) Cytotoxic diterpenoids from the roots of Euphorbia ebracteolata. Planta Med 71(04):349–354
Madureira AM, Gyemant N, Ascenso JR, Abreu PM, Molnar J, Ferreira MJU (2006) Euphoportlandols A and B, tetracylic diterpene polyesters from Euphorbia portlandica and their anti-MDR effects in cancer cells. J Nat Prod 69(6):950–953
Sudhakar M, Rao CV, Rao PM, Raju DB, Venkateswarlu Y (2006) Antimicrobial activity of Caesalpinia pulcherrima, Euphorbia hirta and Asystasia gangeticum. Fitoterapia 77(5):378–380
Kong LY, Li Y, Wu XL, Min ZD (2002) Cytotoxic diterpenoids from Euphorbia pekinensis. Planta Med 68(3):249–252
Zhao M, Wu S, Li J, Tang WX, Wang JI, Zhang SJ (2014) Chemical constituents from Euphorbia lunulata. China J Chin Mater Med 39(12):2289–2294
Zhang WJ, Weng LJ, Yi LT, Geng D (2016) Chemical constituent in whole herb of Euphorbia lunula. Chin Tradit Herbal Drugs 47(4):554–558
Liu C, Liao ZX, Liu SJ, Qu YB, Wang HS (2014) Two new diterpene derivatives from Euphorbia lunulata Bge and their anti-proliferative activities. Fitoterapia 96:33–38
Long C, Ock KM, Hee SJ, Soon KI, Ye KN et al (2012) Abietane diterpenoids of Rosmarinus officinalis and their diacylglycerol acyltransferase-inhibitory activity. Food Chem 132(4):1775–1780
Bajpai VK, Park YH, Na MK, Kang SC (2015) α-Glucosidase and tyrosinase inhibitory effects of an abietane type diterpenoid taxoquinone from Metasequoia glyptostroboides. BMC Complement Altern Med 15:1–6
Ninon Etsassala GER, Waryo TT, Iwuoha EI, Badmus JA, Marnewick JL, Cupido CN, Hussein AA (2019) Alpha-glucosidase and alpha-amylase inhibitory activities of novel abietane diterpenes from Salvia africana-lutea. Antioxidants 8(10):421
Qu YB, Liao ZX, Liu C, Wang XZ, Zhang J (2018) EFLDO induces apoptosis in hepatic cancer cells by caspase activation in vitro and suppresses tumor growth in vivo. Biomed Pharmacother 100:407–416
Liang L, Guo WH, Esquiliano DR, Asai M, Rodriguez S, Giraud J, Kushner JA, White MF, Lopez MF (2010) Insulin-like growth factor 2 and the insulin receptor, but not insulin, regulate fetal hepatic glycogen synthesis. Endocrinology 151:741–747
Acknowledgements
This work was supported by the National Natural Science Foundation of China (41976109), the Qinghai key R & D and transformation project (Qinghai science and Technology Department) (Grant no. 2017-NK-C25). The Open Project of Qinghai Key Laboratory of Qinghai-Tibet Plateau Biological Resources (Grant no. 2017-ZJ-Y10). The Priority Academic Program Development of Jiangsu Higher Education Institutions (Grant no. 1107047002). Key Laboratory for Chemistry and Molecular Engineering of Medicinal Resources (Guangxi Normal University) (Grant no. CMEMR2016-B06).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hou, P., Qu, Y., Liao, Z. et al. A diterpene derivative enhanced insulin signaling induced by high glucose level in HepG2 cells. J Nat Med 74, 434–440 (2020). https://doi.org/10.1007/s11418-019-01384-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11418-019-01384-7