
Abstract
Purpose
Soil nutrient concentrations and stoichiometry are important indicators of plant growth, terrestrial productivity, and ecosystem functioning. Nevertheless, little is known about the vertical distribution and the environmental factors influencing the spatial patterns of different forest types under the “Grain for Green” program and the “Natural Forest Resources Protection” project in Northwest of China.
Materials and methods
We collected 114 soil profile samples within a 0–100-cm depth from black locust and Chinese pine plantations, and secondary oak forests. We determined the vertical distributions of soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), and their ratios along environmental gradients in Shaanxi Province, Northwest China.
Results and discussion
The results showed that both SOC and TN concentrations decreased exponentially within the soil profiles of the three forest types, but there was minimal change in TP. Significant differences in SOC, TN, and TP were found in the surface soil layers among the forest types. Both SOC and TN were relatively low in the N-fixing black locust plantations and TP was comparatively low in the Chinese pine plantations. The C:N:P ratios decreased with increasing soil depth for the three forest types. These ratios were comparatively high in the Chinese pine plantations, relatively low in the black locust plantations, and moderate in the oak forests. The differences in the ratios among the three forest types were more significant in surface soil than in deep soil. Precipitation was positively correlated with the concentrations of SOC and TN and the ratios of C:N:P. Temperature was negatively correlated with concentrations of SOC and TN and the ratios of C:N:P across all soil depths. A log-transformed linear C-N relationship was found for all three forest types, suggesting a well-constrained coupling between the levels of the two elements.
Conclusions
Our results demonstrated the effect of different tree species on soil C:N:P ratios and their controlling factors within soil profiles along environmental gradients. The secondary oak forest accumulated soil C and N more effectively than the plantations. The Chinese pine plantations were relatively more susceptible to P limitation. Therefore, the mechanism of different plant species on soil biogeochemical processes at the whole soil profile level must be considered when developing forest management strategies and implementing vegetation restoration projects.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
As the largest carbon (C) pool in the terrestrial biosphere, soil plays an important role in global C cycling. Soil C stores exceed those found in vegetation biomass and atmospheric reservoirs combined. Therefore, any minor change in soil organic C (SOC) will have a significant impact on atmospheric carbon dioxide (CO2) concentrations (Eswaran et al. 1993). Meanwhile, nitrogen (N) and phosphorus (P) are the key nutrients and limiting elements of terrestrial ecosystems. The availabilities of N and P in soil strongly influence primary production and C accumulation, and a tight correlation has been observed between the C and N in terrestrial biogeochemical cycles (Cleveland and Liptzin 2007). Therefore, a great deal of research worldwide has focused on the storage, pattern, and environmental controls of the spatial-temporal soil C:N:P stoichiometry across different soil types, biomes, topography, and vegetation types. These data have increased the understanding of biogeochemical cycles and the potential feedbacks of terrestrial ecosystems to global climate change (Jobbagy and Jackson 2000; Yang et al. 2007, Yang et al. 2008; Tarnocai et al. 2009; Wiesmeier et al. 2012; Bing et al. 2016; Wang et al. 2017).
Soil C:N:P stoichiometry is regulated by plant nutrient uptake, litterfall, and root exudates. Meanwhile, the physiological strategies of vegetation types to optimize use of limiting nutrients may also affect soil C, N, and P (McGroddy et al. 2004). The effects of vegetation type on soil C:N:P stoichiometry have been documented in many studies. They revealed that conifers and broadleaf trees have different impacts on soil C:N:P characteristics (Jobbagy and Jackson 2000; Chen et al. 2004; Li et al. 2005; Marin-Spiotta and Sharma 2013; Ma et al. 2015; Chai et al. 2015; Dawud et al. 2017; Feng et al. 2017). Moreover, an increasing number of studies also reported significant differences between plantations and natural/secondary forests in terms of soil C:N:P ratios (Chen et al. 2004; Li et al. 2005; Marin-Spiotta and Sharma 2013; Cao and Chen 2017). Soil C and N concentrations was lower in plantations than in natural forests (Behera and Sahani 2003; Lemenih et al. 2004; Lemma et al. 2006; Cao and Chen 2017), but higher in afforested agricultural lands than in natural forests (Wall and Hytonen 2005). A meta-analysis based on 73 published studies showed that soil C and N concentrations in plantations were 36 and 26.5% lower than those in natural forests, respectively (Liao et al. 2012).
Although large-scale soil C:N:P stoichiometry patterns have been surveyed regionally, continentally, and globally, the results have been inconsistent. This discrepancy could be explained in terms of vegetation type, climate zone, biome, soil order, and soil weathering stage (Jobbagy and Jackson 2000; Yang et al. 2007; Tarnocai et al. 2009; Tian et al. 2010; Wiesmeier et al. 2012; Bing et al. 2016; Feng et al. 2017; Wang et al. 2017). Therefore, soil C:N:P stoichiometry should be investigated under different types of plant restoration, particularly in Northwest of China which has experienced large-scale vegetation construction due to severe soil and water losses since the 1970s. The “Grain for Green” program and the “Natural Forest Resources Protection” project were launched in 1998 to restore ecological environments across Northwestern China. They have provided many ecological benefits including increases in vegetation cover (Cui et al. 2015b), biodiversity conservation (Chen and Cao 2014), and forest C sequestration (Cui et al. 2015a; Cao and Chen 2017). However, little information is available on soil C, N, and P concentrations and their stoichiometries in the secondary forests under the aegis of the “Natural Forest Resources Protection” project. In addition, relative differences in soil C:N:P between secondary forests and plantations remain to be determined.
Knowing the effects of plantations and secondary forests on soil C:N:P stoichiometry helps assess soil nutrient status, biogeochemical equilibrium, and ecosystem stability. It also improves predictions for the effects of current and future changes in land use and cover on both regional and global C cycles (Bonner et al. 2013; Cao and Chen 2017). In this study, widely distributed N-fixing black locust (Robinia pseudoacacia) and indigenous Chinese pine (Pinus tabulaeformis) plantations, and secondary oak (Quercus liaotungensis) forests were selected in Shaanxi Province, Northwest China. The vertical distributions of their soil C:N:P stoichiometries were analyzed along environmental gradients from south to north regional climate. The first objective was to compare the vertical distributions and the ratios of the SOC, TN, and TP in the 0–100 cm soil profiles under the three forests. The second objective was to quantify the correlations between the spatial variations in soil C:N:P stoichiometry and selected environmental factors. The results of this study may help improve forest management and vegetation recovery.
2 Methods and materials
2.1 Study area
The study was conducted along a south to north gradient in Shaanxi Province, China (105°29′ to 111°15′ E and 31°42′ to 39°35′ N). Both mean annual temperature (18–6 °C) and mean annual precipitation (1400–320 mm) gradually declined from south to north. The vegetation type ranges from north subtropical broadleaf forest zone to warm temperate humid and semi-humid forest zone and moderate temperate arid and semi-arid desert grassland. The latest forest inventory data available for Shaanxi Province reports that the secondary oak forest, protected under the “Natural Forest Resources Protection” project, accounts for the largest proportion of forest area (50%) and timber volume (50%) in the whole province. The indigenous Chinese pine plantation also accounts for a large proportion of forested area (52%) and timber volume (51%) among all plantations. The fast-growing, N-fixing black locust has been widely planted across Shaanxi Province during the implementation of the “Grain for Green” program, with more than 70,000 ha planted over the past few decades (Qiu et al. 2010). This deciduous N-fixing species is native to North America and was introduced to China from Europe at the beginning of the twentieth century (Shan et al. 2002; Cierjacks et al. 2013; Tsunekawa et al. 2014).
2.2 Field sampling and laboratory analysis
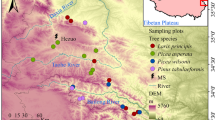
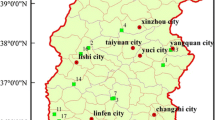
We compared soil C, N, and P stoichiometries in the large proportion of middle-age forests in N-fixing black locust and indigenous Chinese pine plantations (20–30-year-old) and in secondary oak forests (30–40-year-old) based on the forest inventory database of Shaanxi Province. These are the main types of forests covered under the “Grain for Green” program and the “Natural Forest Resources Protection” project implemented across Shaanxi. Black locust and Chinese pine plantations were planted on abandoned cultivated lands, while secondary oak forests were regenerated naturally on abandoned farmlands originally populated by native oak forests before being cleared. Furthermore, 12, 11, and 15 sampling sites were selected for black locust, Chinese pine, and oak forest, respectively, based mainly on the relative distribution of each forest types from south to north through Shaanxi (Electronic Supplementary Material, Fig. S1). All sampling sites were located far from human habitation and were neither fertilized nor irrigated. Longitude, latitude, and elevation were recorded at each site using a global positioning system (GPS).
Field soil sampling was conducted from July to August 2011. A total of 114 soil profiles were collected from 38 sites (three soil profiles per site) along the south to north gradient. Soil samples were taken from each profile at depth intervals of 0–10 cm, 10–20 cm, 20–30 cm, 30–50 cm, and 50–100 cm. Soil bulk density samples were collected using a standard 100 cm3 container and oven-dried at 105 °C for 24 h until a constant weight was obtained. Soil samples for C, N, and P analyses were air-dried for over 1 week, sieved through a 2-mm mesh, handpicked to remove plant detritus, and then ground in a ball mill. Soil organic carbon (SOC) was determined using the wet oxidation method (Nelson et al. 1996). Total N (TN) was measured using the Kjeldahl method (Bremner and Mulvaney 1982), and total P (TP) was determined using the HClO4-H2SO4 colorimetric method (Parkinson and Allen 1975).
2.3 Climate data
Datasets containing the mean annual temperature (MAT) and mean annual precipitation (MAP) for each sampling site used were obtained from the China Meteorological Data Sharing Service System (CMDSSS; http://cdc.cma.gov.cn/). We analyzed the datasets and established correlations between MAT and MAP using geographic variables (longitude, latitude, and altitude) recorded at 52 meteorological stations across the region. We then estimated the temperature and precipitation of the sampling sites based on the established regressions (Fang et al. 2001; Piao et al. 2003; Li et al. 2015). The regressions were calculated in this study as follows:
where X, Y, and H represent longitude, latitude, and elevation, respectively.
2.4 Estimation of SOC, TN, and TP
The SOC density for each soil profile was estimated from the following equation (Batjes 1996; Wiesmeier et al. 2012; Yang et al. 2007, 2008):
where SOCD i , SOC i , BD i , T i , and C i are SOC density (kg m−2), SOC (g kg−1), bulk density (g cm−3), soil thickness (cm), and volume percentage of the fraction > 2 mm at layer i, respectively. The same procedure was used to estimate the densities of TN and TP.
2.5 Statistical analysis
One-way ANOVAs were run with post hoc to determine whether the distributions of SOC, TN, TP, and the stoichiometric ratios of the three forest types differed significantly. A partial correlation analysis was used to examine the relationships among SOC, TN, and TP and the various environmental drivers, latitude, longitude, elevation, temperature, and precipitation (Wang et al. 2004). The level of statistical significance was set at P < 0.05. All analyses were performed using SPSS (version 20, IBM Crop., Armonk, NY, USA).
3 Results
3.1 Vertical distribution of SOC, TN, and TP
In all three forest types, SOC and TN concentrations decreased exponentially with soil depth in the top 0–1 m layer (P < 0.001; Fig. 1a, b). The SOC and TN concentrations were higher in the secondary oak forests than in both Chinese pine and black locust plantations within the first 30 cm (P < 0.001), but there were no significant differences in SOC and TN among the three forest types below 30 cm. In contrast, there was minimal change in soil TP with increasing depth for the three forest types (Fig. 1c). The soil TP concentrations in the Chinese pine plantations were significantly lower than in the black locust plantations and the oak forests along the soil profile (P < 0.001, Fig. 1c). On the other hand, the soil TP concentrations in N-fixing black locust plantations were not higher than those of the secondary oak forests.

Vertical distribution of the SOC (a), TN (b), and TP (c) concentrations for the three forest types
The SOC and TN densities for the three forest types also decreased within soil depth (the last two depth intervals converted to the equivalent of 10 cm thickness; Figs. 2 and 3). The SOC densities of the N-fixing black locust plantations were significantly lower than those of the Chinese pine plantations and secondary oak forests. They did not, however, significantly differ between the secondary oak forests and the Chinese pine plantations in the upper 30 cm layer (Fig. 2). The soil TN densities were significantly higher in the secondary oak forests than these of black locust and Chinese pine plantations at 0–10 cm and 10–20 cm (Fig. 3). The soil TP densities of the N-fixing black locust plantations were significantly higher than those of the non-N-fixing forest types. The soil TP densities of the secondary oak forests, however, were significantly higher than that of the highest density in the Chinese pine plantation within the top 30. Nevertheless, little difference in soil TP density was found between the oak forests and the black locust plantations at depth intervals of 30–50 cm and 50–100 cm (Fig. 4). Approximately 50% of the SOC and TN stocks in the upper 1 m soil layer were stored within the top 30-cm depth interval, while the top 50-cm depth interval was stored about half of the total TP stocks in all three forest types (Figs. 2, 3, and 4).

Vertical distribution of the densities (a–c) and the proportional distribution of SOC (d–f) in black locust plantations (a, d), Chinese pine plantations (b, e), and secondary oak forests (c, f). Different letters denote significant differences among forest types (LSD test, P < 0.05). Error bars represent standard deviation of the means

Vertical distribution of the densities (a–c) and the proportional distributions of TN (d–f) in black locust plantations (a, d), Chinese pine plantations (b, e), and secondary oak forests (c, f). Different letters denote significant differences among forest types (LSD test, P < 0.05). Error bars represent standard deviation of the means

Vertical distribution of the densities (a–c) and the proportional distribution of TP (d–f) in black locust plantations (a, d), Chinese pine plantations (b, e), and secondary oak forests (c, f). Different letters denote significant differences among forest types (LSD test, P < 0.05). Error bars represent standard deviation of the means
3.2 Vertical distributions of SOC, TN, and TP stoichiometric ratios
The soil C:N, C:P, and N:P ratios all significantly decreased (P < 0.001) with increasing soil depth (Fig. 5). The C:N ratio varied only slightly along the profile with a coefficient variation of 11.17% compared to that of the C:P and N:P ratios. The ratios of C:N, C:P, and N:P were significantly higher in the Chinese pine plantations than those in the N-fixing black locust plantations at each depth interval (Fig. 5). Moreover, the C:N:P values in the secondary oak forests were in between those for the black locust and Chinese pine plantations. The differences among the three forest types were more significant in surface soil than in deep soil (Fig. 5).

Vertical distribution of C:N (a), C:P (b), and N:P (c) ratios for black locust plantations, Chinese pine plantations, and secondary oak forests. Different letters denote significant differences among these three forest types (LSD test, P < 0.05). Error bars represent standard deviation of the means
3.3 Environmental controls of SOC, TN, and TP, and their spatial variations
The SOC, TN, and TP concentrations and their ratios exhibited significant spatial variations (Table 1). TP increased with latitude and decreased with longitude and elevation. All the correlations between TP and the geographic variables weakened with increasing soil depth. In contrast, the SOC and TN were significantly correlated with elevation at all soil depths but not with longitude or latitude (Table 1). The C:N, N:P, and C:P ratios were positively correlated with longitude and elevation but negatively correlated with latitude. The correlations were significant at almost each soil interval for N:P and C:P, but only weakly significant at the top 30 cm for C:N (Table 1).
Temperature and precipitation had strong impacts on soil TP, C:N, N:P, and C:P (Table 2). Temperature had a positive effect on soil TP whereas precipitation had a negative effect upon it. Conversely, N:P and C:P ratios were negatively correlated with temperature and positively correlated with precipitation (Table 2).
3.4 Relationships between SOC, TN, and TP concentrations
The TP concentrations were positively correlated with TN, but significantly correlated with SOC only at 0–30 cm (Table 3). There were, however, significant correlations between SOC and TN for all three forest types and all soil depths (Table 3, Fig. 6). In addition, an isometric relationship (the slopes of log-transformed C and N correlations were not statistically different from 1) between SOC and TN was observed in Chinese pine plantations at 20–100 cm and in oak forests in the top 20 cm, respectively (Table 4). In contrast, no isometric relationship was found for black locust plantations. Nevertheless, the slopes of the stoichiometric C-N relationship did not significantly differ among the three forest types (P > 0.05).

Relationship between log-transformed C and N at different soil depths for black locust plantations, Chinese pine plantations, and oak forests. a 0–10 cm. b 10–20 cm. c 20–30 cm. d 30–50 cm. e 50–100 cm. f 0–100 cm. All fitting equations are significant (P < 0.05)
4 Discussion
4.1 Vertical distributions of SOC, TN, and TP concentrations and ratios
In this study, the vertical distributions of the SOC, TN, and TP concentrations and their ratios were investigated in three different types of forest along a south to north gradient in Shaanxi Province, Northwest China. The results showed that, except for TP, the soil nutrient concentrations decreased along the soil profile. These results were consistent with findings from previous studies conducted in plantations on the Loess Plateau (Zhang et al. 2013; Song et al. 2016), subalpine forests of the eastern Tibetan Plateau (Feng et al. 2017), and other land use types (i.e., shrub, crop, and desert) worldwide (Jobbagy and Jackson 2000; Wang et al. 2004; Kahle et al. 2010; Chai et al. 2015). The decreasing trend with soil depth for SOC and TN was determined by the input pattern of organic matter in the soil, such as plant root and litter, which gradually decreased down the vertical profile. On the other hand, the soil TP profile pattern was mainly attributed to the parent material located in the bottom of the soil, which provided the major sources of P (Walker and Adams 1958). Long-term weathering of the parent material, together with an accumulation of P in the surface soil layer through translocation by plants, ultimately resulted in a relatively stable vertical TP distribution (Tian et al. 2010). Consequently, C:P and N:P declined much faster than C:N with increasing soil depth. This observation was consistent with the results reported in a nationwide synthesis of data compiled from many studies conducted in China (Tian et al. 2010). The soil surface layer (0–30 cm) interacts directly with the atmosphere and is the most sensitive to land-use changes (Batjes 1996; Wang et al. 2004). Differences in SOC and TN concentrations and their densities among the three forest types occur mainly within the 0–30 cm surface soil layers.
N-fixing species have been widely proven to increase the soil SOC and TN concentrations, N stores, N cycling and availability, and primary productivity in the recovery of degraded mining lands, deforested lands, and agroforestry systems (Deans et al. 1999; Uselman et al. 2000; Rice et al. 2004; Macedo et al. 2008; Wang et al. 2010). For example, soil organic matter and N concentrations in the 0–5 cm soil layer were 40–50% and 20–50% higher in the N-fixing forests than in the non-N-fixing forests, respectively, in the restoration of degraded lands of southern China (Wang et al. 2010). However, the current study and previous studies indicated that SOC and TN concentrations and densities in the soils under N-fixing black locust stands were lower than those under other species (Tateno et al. 2007; Song et al. 2016). Tateno et al. (2007) described high litter decomposition rates and low litter and soil organic layers inputs in the black locust plantations of the Loess Plateau. In addition, the fast litter decomposition rates of the black locust plantation can also be accounted for by the significantly lower soil C:N ratios in the black locust plantations compared to those under other forest types, especially in the surface soil layer (Bui and Henderson 2013; Zhao et al. 2015). A meta-analysis of 74 publications on the influence of land-use changes on soil C stocks revealed that soil C stocks in N-fixing tree species were not significantly higher than those under other tree species (Guo and Gifford 2002). The secondary oak forests in our field investigations had larger and thicker litter layers than those in the other forest types. Consequently, the soil under secondary oak forest had higher SOC and TN concentrations and densities than those under the black locust and Chinese pine plantations. Therefore, secondary oak forests were more effective at accumulating C and N than black locust and Chinese pine plantations.
Soil TP patterns are affected by the interactions among climate, vegetation type, and parent material (Sundqvist et al. 2014; Zhou et al. 2016). It is generally accepted that soil P concentrations in most regions of China, especially the Loess Plateau, are below the global average, resulting from low P inputs by weathering and strong losses by soil erosion and water runoff (Han et al. 2005; Zheng and Shangguan 2007; Zhou et al. 2016). In our study, we observed that soil TP concentration and density were significantly lower in the coniferous Chinese pine plantations than in the two broad-leaved forest types, which is mainly a result of more acidity in the foliage and litter of coniferous trees. Moreover, the concentrations of other P forms (i.e., the diluted HCl-extractable inorganic P) were significantly lower in coniferous forests than those in broadleaf forests of Chile and Southwest China (Thomas et al. 1999; Zhou et al. 2016). In addition, the soil N:P ratios in the Chinese pine plantations were higher than those under the other two forest types which implied P limitation for growth of Chinese pine in the study area. Therefore, the synergistic effects of soil nutrient limitation with aggravating water deficiency conditions induced by global warming may threaten the recovery of Chinese pine plantations. These facts should be taken into consideration when planning forest management and ecological restoration.
4.2 Relationships between SOC, TN, TP, and environmental factors
Our analysis indicated significant effects of climate on soil nutrient concentrations and their ratios. Both SOC and TN concentrations were positively correlated with precipitation and negatively correlated with temperature, but not significantly. These results were consistent with those obtained from Chinese (Wu et al. 2003; Wang et al. 2004; Yang et al. 2007), American (Epstein et al. 2002), and global (Jobbagy and Jackson 2000) datasets. The SOC was determined from the balance between plant biomass production input and the microbiological decomposition output (Post et al. 1982; Davidson and Janssens 2006). Increased precipitation may promote plant growth, thereby increasing soil C and N inputs in the form of litterfall and fine roots. Increases in temperature would accelerate the mineralization of soil organic matter and decrease both SOC and N (Fang et al. 2001; Wu et al. 2003; Yang et al. 2007; Chai et al. 2015). However, the combined effects of temperature and precipitation on SOC and TN are complex and not simply the sum of the individual influences of each factor (Jobbagy and Jackson 2000; Davidson and Janssens 2006). In our study area, both precipitation and temperature declined gradually from south to north, while precipitation was the main variable affecting SOC. Consequently, the SOC storage also decreased from south to north similarly to the precipitation gradients. The overall SOC storage in China is inversely proportional to temperature and directly proportional to precipitation. The correlation between SOC and temperature was stronger than that between SOC and precipitation (Wang et al. 2004). In contrast, both precipitation and temperature significantly affect soil TP. Increased temperature accelerates the weathering of parent materials where the soil P originates, and an increase in precipitation facilitates the leaching of P from the soil. Therefore, soil TP increases with latitude and decreases with longitude because of the precipitation gradients in these two spatial variables. Soil TP also decreases with elevation since temperature falls with rising altitude. In our study, the N concentration increased and the P concentration decreased with increasing elevation (Table 3). The results of our study corroborate those previous reports which also stated that C and P concentrations, litter decomposition rates, and microbial N and P uptake and release all decline with increasing elevation (Sundqvist et al. 2014).
A linear log-transformed soil C-N relationship was found in all three forest types, suggesting a closely linked interaction between SOC and TN. Since the slope of the C-N stoichiometric relationships was not statistically different from 1.0, the SOC and TN were scaled isometrically in both the Chinese pine and oak forests. Similar soil C-N isometric patterns were also documented in a global forest meta-analysis (Cleveland and Liptzin 2007; Yang and Luo 2011). These trends may reflect the isometric C-N stoichiometry of plants from which soil C and N originate (McGroddy et al. 2004; Cleveland and Liptzin 2007). Certain studies, however, found no isometric C-N stoichiometry in plants, which complicated our understanding of the interaction between plants and soils (Yang and Luo 2011). This isometric stoichiometry was not examined between soil N (or C) and P at every depth in this study, although many studies have indicated a tightly linked relationship between N and P in plant leaves (Han et al. 2005; Reich and Oleksyn 2004; Reich et al. 2010) and fine roots (Yuan et al. 2011). A well-constrained soil N:P was noted in the Chinese (Tian et al. 2010) and global (Cleveland and Liptzin 2007) datasets. In both cases, only the surface (0–10 cm) had relatively consistent N:P ratios. Decoupling soil N (or C) and P in deeper soil layers is the result of differences in the main sources of these nutrients. The SOC was highly correlated with soil organic P and C, whereas N and P were moderately associated with the soil organic matter (Walker and Adams 1958).
5 Conclusions
Large-scale ecological restoration projects have been conducted in Northwestern China using different plant species, which have exerted profound effects on the dynamics and patterns of soil nutrients. Our field investigation examined the vertical distributions of soil C, N, and P concentrations and their ratios for the three major forest types in the region. We found that both SOC and TN decreased but TP only slightly decreased along the soil depth profile for all three forests. The concentrations and storage of SOC and TN in N-fixing black locust plantations were not higher than those in the indigenous Chinese pine plantations or the secondary oak forests. Precipitation and temperature had significant effects on SOC, TN, and TP concentrations and their ratios. SOC and TN varied directly with precipitation and inversely with temperature. In contrast, TP is less significantly affected by climate since its source is primarily the parent material in the deeper soil strata. Since they grew mainly in arid and semi-arid nature of the region, Chinese pine plantations had lower soil TP densities and were more severely limited in terms of nutrient and water availability than the black locust and oak forests. Consequently, Chinese pine should be given particular attention when developing ecological restoration projects in the region.
References
Batjes NH (1996) Total carbon and nitrogen in the soils of the world. Eur J Soil Sci 47(2):151–163. https://doi.org/10.1111/j.1365-2389.1996.tb01386.x
Behera N, Sahani U (2003) Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil. For Ecol Manag 174(1-3):1–11. https://doi.org/10.1016/S0378-1127(02)00057-9
Bing HJ, Wu YH, Zhou J, Sun HY, Luo J, Wang JP, Yu D (2016) Stoichiometric variation of carbon, nitrogen, and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of eastern Tibetan Plateau. J Soils Sediments 16(2):405–416. https://doi.org/10.1007/s11368-015-1200-9
Bonner MTL, Schmidt S, Shoo LP (2013) A meta-analytical global comparison of aboveground biomass accumulation between tropical secondary forests and monoculture plantations. For Ecol Manag 291:73–86. https://doi.org/10.1016/j.foreco.2012.11.024
Bremner J, Mulvaney C (1982) Nitrogen-total, methods of soil analysis. Part 2. Chemical and microbiological properties, p 595–624
Bui EN, Henderson BL (2013) C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 373(1-2):553–568. https://doi.org/10.1007/s11104-013-1823-9
Cao Y, Chen Y (2017) Ecosystem C:N:P stoichiometry and carbon storage in plantations and a secondary forest on the Loess Plateau, China. Ecol Eng 105:125–132. https://doi.org/10.1016/j.ecoleng.2017.04.024
Chai H, Yu GR, He NP, Wen D, Li J, Fang JP (2015) Vertical distribution of soil carbon, nitrogen, and phosphorus in typical Chinese terrestrial ecosystems. Chin Geogr Sci 25(5):549–560. https://doi.org/10.1007/s11769-015-0756-z
Chen Y, Cao Y (2014) Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China. Ecol Eng 73:238–245. https://doi.org/10.1016/j.ecoleng.2014.09.055
Chen CR, Xu ZH, Mathers NJ (2004) Soil carbon pools in adjacent natural and plantation forests of subtropical Australia. Soil Sci Soc Am J 68(1):282–291. https://doi.org/10.2136/sssaj2004.2820
Cierjacks A, Kowarik I, Joshi J, Hempel S, Ristow M, von der Lippe M, Weber E (2013) Biological Flora of the British Isles: Robinia pseudoacacia. J Ecol 101(6):1623–1640. https://doi.org/10.1111/1365-2745.12162
Cleveland CC, Liptzin D (2007) C:N:P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85(3):235–252. https://doi.org/10.1007/s10533-007-9132-0
Cui GY, Chen YM, Cao Y (2015a) Temporal-spatial pattern of carbon stocks in forest ecosystems in Shaanxi, Northwest China. Plos One 10(11):e0142753. https://doi.org/10.1371/journal.pone.0142753
Cui GY, Chen YM, Cao Y, Chun-Chun AN (2015b) Analysis on carbon stock distribution patterns of forest ecosystems in Shaanxi Province. Chin J Plant Ecol 39:333–342
Davidson EA, Janssens IA (2006) Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440(7081):165–173. https://doi.org/10.1038/nature04514
Dawud SM, Raulund-Rasmussen K, Ratcliffe S, Domisch T, Finer L, Joly FX, Hattenschwiler S, Vesterdal L (2017) Tree species functional group is a more important driver of soil properties than tree species diversity across major European forest types. Funct Ecol 31(5):1153–1162. https://doi.org/10.1111/1365-2435.12821
Deans JD, Diagne O, Lindley DK, Dione M, Parkinson JA (1999) Nutrient and organic-matter accumulation in Acacia senegal fallows over 18 years. For Ecol Manag 124(2-3):153–167. https://doi.org/10.1016/S0378-1127(99)00063-8
Epstein HE, Burke IC, Lauenroth WK (2002) Regional patterns of decomposition and primary production rates in the US Great Plains. Ecology 83:320–327
Eswaran H, Vandenberg E, Reich P (1993) Organic-carbon in soils of the world. Soil Sci Soc Am J 57(1):192–194. https://doi.org/10.2136/sssaj1993.03615995005700010034x
Fang JY, Piao SL, Tang ZY, Peng CH, Wei J (2001) Interannual variability in net primary production and precipitation. Science 293(5536):1723–11723. https://doi.org/10.1126/science.293.5536.1723a
Feng DF, Bao WK, Pang XY (2017) Consistent profile pattern and spatial variation of soil C/N/P stoichiometric ratios in the subalpine forests. J Soils Sediments 17(8):2054–2065. https://doi.org/10.1007/s11368-017-1665-9
Guo LB, Gifford RM (2002) Soil carbon stocks and land use change: a meta analysis. Glob Chang Biol 8(4):345–360. https://doi.org/10.1046/j.1354-1013.2002.00486.x
Han WX, Fang JY, Guo DL, Zhang Y (2005) Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol 168(2):377–385. https://doi.org/10.1111/j.1469-8137.2005.01530.x
Jobbagy EG, Jackson RB (2000) The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol Appl 10(2):423–436. https://doi.org/10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2
Kahle P, Baum C, Boelcke B, Kohl J, Ulrich R (2010) Vertical distribution of soil properties under short-rotation forestry in Northern Germany. J Plant Nutr Soil Sci 173(5):737–746. https://doi.org/10.1002/jpln.200900230
Lemenih M, Olsson M, Karltun E (2004) Comparison of soil attributes under Cupressus lusitanica and Eucalyptus saligna established on abandoned farmlands with continuously cropped farmlands and natural forest in Ethiopia. For Ecol Manag 195(1-2):57–67. https://doi.org/10.1016/j.foreco.2004.02.055
Lemma B, Kleja DB, Nilsson I, Olsson M (2006) Soil carbon sequestration under different exotic tree species in the southwestern highlands of Ethiopia. Geoderma 136(3-4):886–898. https://doi.org/10.1016/j.geoderma.2006.06.008
Li Y, Xu M, Zou XM, Shi PJ, Zhang YQ (2005) Comparing soil organic carbon dynamics in plantation and secondary forest in wet tropics in Puerto Rico. Glob Chang Biol 11(2):239–248. https://doi.org/10.1111/j.1365-2486.2005.00896.x
Li ZQ, Yang L, Lu W, Guo W, Gong XS, Xu J, Yu D (2015) Spatial patterns of leaf carbon, nitrogen stoichiometry and stable carbon isotope composition of Ranunculus natans CA Mey. (Ranunculaceae) in the arid zone of northwest China. Ecol Eng 77:9–17. https://doi.org/10.1016/j.ecoleng.2015.01.010
Liao CZ, Luo YQ, Fang CM, Chen JK, Li B (2012) The effects of plantation practice on soil properties based on the comparison between natural and planted forests: a meta-analysis. Glob Ecol Biogeogr 21(3):318–327. https://doi.org/10.1111/j.1466-8238.2011.00690.x
Ma J, Bu RC, Liu M, Chang Y, Qin Q, Hu YM (2015) Ecosystem carbon storage distribution between plant and soil in different forest types in Northeastern China. Ecol Eng 81:353–362. https://doi.org/10.1016/j.ecoleng.2015.04.080
Macedo MO, Resende AS, Garcia PC, Boddey RM, Jantalia CP, Urquiaga S, Campello EFC, Franco AA (2008) Changes in soil C and N stocks and nutrient dynamics 13 years after recovery of degraded land using leguminous nitrogen-fixing trees. For Ecol Manag 255(5-6):1516–1524. https://doi.org/10.1016/j.foreco.2007.11.007
Marin-Spiotta E, Sharma S (2013) Carbon storage in successional and plantation forest soils: a tropical analysis. Glob Ecol Biogeogr 22(1):105–117. https://doi.org/10.1111/j.1466-8238.2012.00788.x
McGroddy ME, Daufresne T, Hedin LO (2004) Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial redfield-type ratios. Ecology 85(9):2390–2401. https://doi.org/10.1890/03-0351
Nelson DW, Sommers LE, Sparks D, Page A, Helmke P, Loeppert R, Soltanpour P, Tabatabai M, Johnston C, Sumner M (1996) Total carbon, organic carbon, and organic matter. Methods of soil analysis. Part 3—chemical methods, p 961–1010
Parkinson J, Allen S (1975) A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun Soil Sci Plant Anal 6(1):1–11. https://doi.org/10.1080/00103627509366539
Piao SL, Fang JY, Zhou LM, Guo QH, Henderson M, Ji W, Li Y, Tao S (2003) Interannual variations of monthly and seasonal normalized difference vegetation index (NDVI) in China from 1982 to 1999. J Geophys Res-Atmos 108(D14):4401. https://doi.org/10.1029/2002JD002848
Post WM, Emanuel WR, Zinke PJ, Stangenberger AG (1982) Soil carbon pools and world life zones. Nature 298(5870):156–159. https://doi.org/10.1038/298156a0
Qiu LP, Zhang XC, Cheng JM, Yin XQ (2010) Effects of black locust (Robinia pseudoacacia) on soil properties in the loessial gully region of the Loess Plateau, China. Plant Soil 332(1-2):207–217. https://doi.org/10.1007/s11104-010-0286-5
Reich PB, Oleksyn J (2004) Global patterns of plant leaf N and P in relation to temperature and latitude. PNAS 101(30):11001–11006. https://doi.org/10.1073/pnas.0403588101
Reich PB, Oleksyn J, Wright IJ, Niklas KJ, Hedin L, Elser JJ (2010) Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes. Proc Roy Soc B-Biol Sci 277(1683):877–883. https://doi.org/10.1098/rspb.2009.1818
Rice SK, Westerman B, Federici R (2004) Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen-cycling in a pine-oak ecosystem. Plant Ecol 174(1):97–107. https://doi.org/10.1023/B:VEGE.0000046049.21900.5a
Shan C, Liang Z, Hao W (2002) Review on growth of locust and soil water in Loess Plateau. Acta Botan Boreali-Occiden Sin 23:1341–1346
Song BL, Yan MJ, Hou H, Guan JH, Shi WY, Li GQ, Du S (2016) Distribution of soil carbon and nitrogen in two typical forests in the semiarid region of the Loess Plateau, China. Catena 143:159–166. https://doi.org/10.1016/j.catena.2016.04.004
Sundqvist MK, Wardle DA, Vincent A, Giesler R (2014) Contrasting nitrogen and phosphorus dynamics across an elevational gradient for subarctic tundra heath and meadow vegetation. Plant Soil 383(1-2):387–399. https://doi.org/10.1007/s11104-014-2179-5
Tarnocai C, Canadell JG, Schuur EAG, Kuhry P, Mazhitova G, Zimov S (2009) Soil organic carbon pools in the northern circumpolar permafrost region. Glob Biogeochem Cycle 23:2607–2617
Tateno R, Tokuchi N, Yamanaka N, Du S, Otsuki K, Shimamura T, Xue Z, Wang S, Hou Q (2007) Comparison of litterfall production and leaf litter decomposition between an exotic black locust plantation and an indigenous oak forest near Yan’an on the Loess Plateau, China. For Ecol Manag 241(1-3):84–90. https://doi.org/10.1016/j.foreco.2006.12.026
Thomas SM, Johnson AH, Frizano J, Vann DR, Zarin DJ, Joshi A (1999) Phosphorus fractions in montane forest soils of the Cordillera de Piuchue, Chile: biogeochemical implications. Plant Soil 211(2):139–148. https://doi.org/10.1023/A:1004686213319
Tian HQ, Chen GS, Zhang C, Melillo JM, Hall CAS (2010) Pattern and variation of C:N:P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98(1-3):139–151. https://doi.org/10.1007/s10533-009-9382-0
Tsunekawa A, Liu G, Yamanaka N, Du S (2014) Restoration and development of the degraded Loess Plateau. Springer, China. https://doi.org/10.1007/978-4-431-54481-4
Uselman SM, Qualls RG, Thomas RB (2000) Effects of increased atmospheric CO2, temperature, and soil N availability on root exudation of dissolved organic carbon by a N-fixing tree (Robinia pseudoacacia L.) Plant Soil 222(1/2):191–202. https://doi.org/10.1023/A:1004705416108
Walker TW, Adams AFR (1958) Studies on soil organic matter: I. Influence of phosphorus content of parent materials on accumulations of carbon, nitrogen, sulfur, and organic phosphorus in grassland soils. Soil Sci 85(6):307–318. https://doi.org/10.1097/00010694-195806000-00004
Wall A, Hytonen J (2005) Soil fertility of afforested arable land compared to continuously. Plant Soil 275(1-2):247–260. https://doi.org/10.1007/s11104-005-1869-4
Wang S, Huang M, Shao X, Mickler RA, Li K, Ji J (2004) Vertical distribution of soil organic carbon in China. Environ Manag 33:S200–S209
Wang FM, Li ZA, Xia HP, Zou B, Li NY, Liu J, Zhu WX (2010) Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Sci Plant Nutr 56(2):297–306. https://doi.org/10.1111/j.1747-0765.2010.00454.x
Wang QD, Song JM, Cao L, Li XG, Yuan HM, Li N (2017) Distribution and storage of soil organic carbon in a coastal wetland under the pressure of human activities. J Soils Sediments 17(1):11–22. https://doi.org/10.1007/s11368-016-1475-5
Wiesmeier M, Sporlein P, Geuss U, Hangen E, Haug S, Reischl A, Schilling B, von Lutzow M, Kogel-Knabner I (2012) Soil organic carbon stocks in southeast Germany (Bavaria) as affected by land use, soil type and sampling depth. Glob Chang Biol 18(7):2233–2245. https://doi.org/10.1111/j.1365-2486.2012.02699.x
Wu HB, Guo ZT, Peng CH (2003) Distribution and storage of soil organic carbon in China. Glob Biogeochem Cycle 17:67–80
Yang Y, Luo Y (2011) Carbon: nitrogen stoichiometry in forest ecosystems during stand development. Glob Ecol Biogeogr 20(2):354–361. https://doi.org/10.1111/j.1466-8238.2010.00602.x
Yang YH, Mohammat A, Feng JM, Zhou R, Fang JY (2007) Storage, patterns and environmental controls of soil organic carbon in China. Biogeochemistry 84(2):131–141. https://doi.org/10.1007/s10533-007-9109-z
Yang YH, Fang JY, Tang YH, Ji CJ, Zheng CY, He JS, Zhu BA (2008) Storage, patterns and controls of soil organic carbon in the Tibetan grasslands. Glob Chang Biol 14(7):1592–1599. https://doi.org/10.1111/j.1365-2486.2008.01591.x
Yuan ZY, Chen HYH, Reich PB (2011) Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nat Commun 2:344. https://doi.org/10.1038/ncomms1346
Zhang C, Liu GB, Xue S, Sun CL (2013) Soil organic carbon and total nitrogen storage as affected by land use in a small watershed of the Loess Plateau, China. Eur J Soil Biol 54:16–24. https://doi.org/10.1016/j.ejsobi.2012.10.007
Zhao FZ, Sun J, Ren CJ, Kang D, Deng J, Han XH, Yang GH, Feng YZ, Ren GX (2015) Land use change influences soil C, N, and P stoichiometry under ‘Grain-to-Green Program’ in China. Sci Rep 5:10195
Zheng S, Shangguan Z (2007) Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China. Trees Struct Funct 21(3):357–370. https://doi.org/10.1007/s00468-007-0129-z
Zhou J, Wu YH, Bing HJ, Yang ZJ, Wang JP, Sun HY, Sun SQ, Luo J (2016) Variations in soil phosphorus biogeochemistry across six vegetation types along an altitudinal gradient in SW China. Catena 142:102–111. https://doi.org/10.1016/j.catena.2016.03.004
Funding
This research was supported by National Key R&D Program of China (2016YFC0501703), CAS “Light of West China” Program, and the National Nature Science Foundation of China (No. 41371506 and 41201088).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible editor: Chengrong Chen
Electronic supplementary material
Fig. S1
Sampling sites of the three forest types in Shaanxi Province (JPEG 1222 kb)
Rights and permissions
About this article

{kind=link}
Cite this article
Cao, Y., Zhang, P. & Chen, Y. Soil C:N:P stoichiometry in plantations of N-fixing black locust and indigenous pine, and secondary oak forests in Northwest China. J Soils Sediments 18, 1478–1489 (2018). https://doi.org/10.1007/s11368-017-1884-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-017-1884-0