
Abstract
As an ancient cellular co-factor ubiquitously present in all domains of life, nearly all iron-sulfur ([Fe-S]) clusters are assembled in the mitochondrion. Although multiple mitochondrion-derived signalings are known to be key players in longevity regulation, whether the mitochondrial [Fe-S] cluster assembly machinery modulates lifespan is previously unknown. Here, we find that ISCU-1, the C. elegans ortholog of the evolutionarily conserved iron-sulfur cluster (ISC) assembly machinery central protein ISCU, regulates longevity and stress response. Specifically, ISCU-1 accelerates aging in the intestine. Moreover, we identify the Nrf2 transcription factor SKN-1 and a nuclear hormone receptor NHR-49 as the downstream factors of ISCU-1. Lastly, a mitochondrial outer membrane protein phosphatase PGAM-5 appears to link ISCU-1 to SKN-1 and NHR-49 in lifespan regulation. Together, we have identified a novel function of mitochondrial ISC assembly machinery in longevity modulation and stress response.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Mitochondria modulate aging through multifaceted pathways. For example, a mild inhibition of the electron transport chain extends lifespan in multiple species [1]. Intriguingly, the mitochondrial electron transport chain signaling modulates aging in a cell-nonautonomous fashion and occurs specifically during the developmental stage [2, 3]. In addition, the free radical theory of aging suggests that the mitochondrion-derived reactive oxygen species (ROS) are a major trigger of organismal aging [4]. However, recent findings have challenged this view by showing that a mild ROS elevation can actually delay aging through mitohormesis [5]. Furthermore, various cellular stress-induced mitochondrial impairment can often trigger an adaptive transcriptional response known as the mitochondrial unfolded protein response (UPRmt). Mechanistic studies primarily conducted in C. elegans demonstrated that the UPRmt-induced signaling can also regulate lifespan in a cell-nonautonomous manner [3, 6, 7]. Lastly, both mitochondrial dynamics (fusion and fission) and mitophagy are known to modulate longevity in many species [8].
Among all essential functions of mitochondria, the biogenesis of [Fe-S] clusters is highly conserved throughout evolution [9]. Although nearly all [Fe-S] clusters are synthesized in mitochondria, it remains unknown whether the mitochondrial [Fe-S] cluster (ISC) assembly machinery modulates aging. The ISC assembly machinery is composed of at least 18 distinct proteins involved in the proper biogenesis and trafficking of [Fe-S] clusters within mitochondria [10]. Among these components, ISCS and ISCU are two central proteins with ISCS acting as a cysteine desulfurase which obtains S from cysteine and serves as a S donor for [Fe-S] cluster assembly, while ISCU being the scaffold protein to accept S and Fe atoms, assemble [Fe-S] clusters and then transfer them to recipient apoproteins [10].
In the current study, we investigated the novel functions of ISCU-1/ISCU in the aging process of C. elegans, a well-established model organism for aging research. Our findings reveal that the suppression of ISCU-1 significantly delays aging in C. elegans. Furthermore, we elucidated the nuclear factors downstream of ISCU-1 in lifespan modulation and stress responses. These results link an ancient [Fe-S] cluster biogenesis protein with longevity modulation.
Methods
Strains
All C. elegans strains were maintained at 20 °C on nematode growth medium (NGM) plates seeded with the OP50 strain of Escherichia coli as described before [11]. A list of strains used in this study is as below: Wild type: N2. SJ4100: zcIs13[hsp-6::GFP]. MGH171: sid-1(qt9);aIxIs9[vha-6p::sid-1::SL2::GFP]. WM118: rde-1(ne300);neIs9[myo-3::HA::RDE-1 + rol-6(su1006)]. NR222: rde-1(ne219);kzIs9[(pKK1260) lin-26p::NLS::GFP + (pKK1253) lin-26p::rde-1 + rol-6(su1006)]. TU3595: sid-1(pk3321);him-5(e1490);lin-15B(n744);uIs72[pCFJ90(myo-2p::mCherry) + unc-119p::sid-1 + mec-18p::mec-18::GFP]. CU5991: fzo-1(tm1133). CU6372: drp-1(tm1108). RB2547: pink-1(ok3538). VC1024: pdr-1(gk448). VC3201: atfs-1(gk3094). QV225: skn-1(zj15). LD1: ldIs7[skn-1b/c::GFP + rol-6(su1006)]. CL2166: dvIs19[(pAF15)gst-4p::GFP::NLS]. STE68: nhr-49(nr2041). AGP24: glmEx5[nhr-49p::nhr-49::gfp;myo-2p::mCherry] (a gift of Dr. Arjumand Ghazi). KU25: pmk-1(km25). PS3551: hsf-1(sy441). RUX65: daf-16(mgDf47). ZG31: hif-1(ia4). RUX713: xbp-1(zc12). AA86: daf-12(rh61rh411). DA465: eat-2(ad465). TM4941: Y45F10D.4(tm4941/nT1). RUX1009: zaxEx189[Piscu-1::SL2::mCherry;Plin-44::gfp];N2. RUX1167: zaxEx204[Pdpy-7::iscu-1::mCherry;Pdpy-7::mitoYFP;lin-44::gfp];N2. RUX1163: zaxEx201[Pges-1::iscu-1::mCherry;Pges-1::mitoYFP;lin-44::gfp];N2. RUX765: zaxIs3[Pges-1::Hyper3::SL2::mCherry;Punc-122::RFP]. RUX1027: zaxIs11[Pges-1::mitoYFP::SL2::mCherry;Plin-44::gfp]. RUX1116: zaxIs12[Piscu-1::iscu-1::SL2::mCherry;Plin-44::gfp]. RUX1325: zaxEx272[Ppgam-5::pgam-5::SL2::yfp;Plin-44::gfp].
All RNAi clones were picked from the Ahringer RNAi library except pgam-5 which was custom designed (forward: 5′-GTACCCACAGCTACTCTTGC-3′ and reverse: 5′-GACCAATATCTCCGATGGAACG-3′). All RNAi plasmids were sequence verified by DNA sequencing.
RNAi assay
Standard RNAi feeding assays were performed as described before [12, 13]. Briefly, freshly streaked single colonies of HT115 bacteria containing either empty vector L4440 (control) or RNAi plasmid were grown up overnight at 37 °C in 2X YT medium supplemented with carbenicillin (25 μg/ml). A couple of days before experiments, RNAi bacteria were seeded on RNAi NGM plates containing carbenicillin (25 μg/ml) and IPTG (1 mM). 5-fluoro-2′-deoxyuridine (FUdR) (100 μM final concentration) was added into seeded plates to prevent progeny production. Of note, iscu-1 RNAi extended lifespan with or without FUdR (Supplementary fig. 1a).
Lifespan assay
All lifespan studies were performed at 20 °C as described before [12,13,14]. In all experiments, the first day of adulthood was counted as day 1. For each lifespan assay, worms were transferred every few days to fresh 60 mm NGM plates before they stop producing progenies. Worms were censored if they crawled off the plate, exploded, bagged, or became contaminated for the accuracy of data interpretation. If not specified, adulthood-specific iscu-1 RNAi was used in all lifespan studies to avoid the interference on development. For convenience, FUdR (100 μM final concentration) was added in all lifespan assays except Supplementary figure 1a. The statistics of lifespan assays was presented in the Supplementary table 1.
Stress assay
For the acute extracellular iron stress assay, 40 age-synchronized adult worms were exposed to different doses of FeCl3 (0–20 mM) dissolved in NGM buffer (without agar) supplemented with OP50 bacteria food. The survival rate was counted every 10 min in the first hour of treatment and then every half an hour in the second hour. Each iron stress assay was repeated at least three times. For the oxidative stress assay, 40 age-synchronized adult worms were transferred to the OP50 bacteria-seeded NGM assay plates containing 6 mM tBuOOH and the survival rate was continuously monitored. Each oxidative stress assay was repeated at least twice. To study the temporal requirement of iscu-1 RNAi on stress resistance, iscu-1 RNAi was initiated from egg or after the L4 stage, while stress assays were conducted on day 1 (developmental stage effect) or day 6 (adulthood effect), respectively.
qRT-PCR
To quantify the mRNA levels of pgam-5, timm-17, cox-4, hmg-5, and atp-5, qRT-PCR was conducted with a CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad). Total RNA was extracted with TRI Reagent (Thermo Fisher) from a group of age-synchronized worms (treated with or without iscu-1 RNAi). The relative amount of mRNAs was analyzed by quantitative PCR (qPCR) with CYBR Green (Thermo Fisher) according to the protocol provided by the manufacturer. For normalization, act-1 (actin) was used as an internal reference and the ΔΔCt method was used to analyze the qPCR data. To measure the ratio of mitochondrial DNA copy number versus nuclear DNA copy number, we adopted a well-established protocol as described before [15]. For all our qPCR assays, we conducted 3–5 biological replicates (individual RNA extractions) and each time had three technical replicates. The qPCR results were plotted with biological replicates and each data point represented the average value of three technical replicates. qPCR primers used in this study: act-1 forward: GCTGGACGTGATCTTACTGATTACC, act-1 reverse: GTAGCAGAGCTTCTCCTTGATGTC; cox-4 forward: ACATCTACCAAGATCGTCTCG, cox-4 reverse: CAAGTTCTTCCAGTCGGCTT; timm-17 forward: GAGAGCCATGCCCATATCGTATC, timm-17 reverse: CAATTTCTTTCCTTTCGCCGCATTT; hmg-5 forward: GTCTGGAGCTGGAATGGAATC, hmg-5 reverse: CTTCGCTTCGTCTGTGTACTTCTT; atp-5 forward: ATGAGGTCAAGGTCGCTGATG, atp-5 reverse: GCTGGGAAATACTCGACGAA; pgam-5 forward: CGTCATTCGGTATTTCATCTGC, pgam-5 reverse: GACCAATATCTCCGATGGAACG; acs-2 forward: TCCGGATAAGGAGTTCTGTG, acs-2 reverse: ATTTGACGGACGTCATGGT; ech-1.2 forward: TGGATAAGCTCCAATCCGA, ech-1.2 reverse: ATTTGGATGTCAGCTCCTG. For mitochondrial DNA copy number quantification: ND1 forward: AGCGTCATTTATTGGGAAGAAGAC, ND1 reverse: AAGCTTGTGCTAATCCCATAAATGT; cox-4 forward: GCCGACTGGAAGAACTTGTC, cox-4 reverse: GCGGAGATCACCTTCCAGTA.
Brood size assay and body bending assay
To determine the brood size, each individual L4 worms were placed on a separate NGM plate and transferred to a new plate every day until they stopped laying eggs. The total amount of eggs (not including unfertilized oocytes) on each plate were counted as brood size.
To determine the body bending frequency, the locomotion of worms at different ages was monitored on NGM plates under a stereoscope. The number of body bends per minute was manually counted for worms treated with or without iscu-1 RNAi.
Oxygen consumption assay
The oxygen consumption rate (OCR) was measured with a Seahorse XFe96 Analyzer (Agilent) using a protocol described before [12]. Age-synchronized day 1 and day 10 adult worms treated with or without iscu-1 RNAi were assayed. ~10 worms were transferred into each well of a 96-well Seahorse assay plate and OCR was measured according to the standard protocol recommended by the manufacturer. Final OCR was calculated by dividing the measured OCR by the number of assayed worms.
ROS measurement using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH2)
To measure the endogenous ROS level, a membrane-permeable fluorescent ROS indicator DCFH2 (Thermo Fisher) was used to quantify worms treated with or without iscu-1 RNAi. Similar to a previously published protocol [16], RNAi-treated and age-synchronized worms (day 1 or day 10) were transferred with a density of 10 worms/well to a 96-well assay plate which contains the ROS sensor DCFH2 (25 μM in M9 buffer). After 2 h of incubation, a multi-mode plate reader was used to measure the fluorescence with the excitation wavelength of 485 nm and emission wavelength of 528 nm. The background fluorescence intensity with DCFH2 in M9 buffer was subtracted for final data analysis, and the DCFH2 ROS assay was repeated three times for each condition.
Mitochondrial morphology staining using MitoTracker Red CMXRos
To observe the morphology of mitochondria, MitoTracker Red CMXRos (Thermo Fisher) was used to stain worms using a previously published protocol [17]. The dye was added in seeded NGM plates under the dark condition at least 1 day before the experiment. A Nikon Eclipse Ti confocal microscope coupled with a Zyla-4.2 camera (Oxford Instruments) was used to take images of stained mitochondria in the anterior intestine cells.
Fluorescence microscopy
For all imaging experiments, worms were immobilized with 10 mM levamisole on a 2% agarose pad prepared on glass slides. The standard epi-fluorescence imaging was performed on an upright Olympus BX51 microscope coupled with a QImaging optiMOS sCMOS camera. For the HyPer3 ratiometric imaging of ROS, transgenic worms carrying Pges-1::HyPer3 were imaged with an inverted Olympus IX70 microscope coupled with a Sutter Lambda 10-2 optical filter changer and Lambda LS light source. Fluorescence was acquired by a Photometrics Prime 95B™ sCMOS camera in two channels corresponding to the two excitation peaks of HyPer3 (420 and 500 nm) with emission at 510 nm. The ImageJ program was used to quantify all imaging experiments.
Quantification and statistical analysis
Data analysis was performed using Prism 7 (GraphPad Software) and Sigmaplot 13 (Systat Software). Error bars were mean ± SEM. P values were determined by log-rank (Kaplan-Meier) test or unpaired two-tailed t test. P < 0.05 was considered statistically significant. Sample sizes were determined by the reproducibility of the experiments and were similar to the ones generally used by the field.
Results
ISCU-1 suppresses longevity
[Fe-S] cluster assembly is an essential function of mitochondria throughout evolution [9]. Since ISCU is a conserved mitochondrial protein required for the initial de novo synthesis of [Fe-S] clusters, using C. elegans as the model system, we set to examine the potential role of ISCU in longevity modulation. The C. elegans genome encodes a single ortholog of human ISCU, ISCU-1, which shares over 60% identity and ~ 90% homology in amino acid sequence with its mammalian counterpart [18, 19]. Deletion mutants of iscu-1 are maintained by a fluorescent nT1 genetic balancer (iscu-1(tm4696)/nT1 and iscu-1(tm4941)/nT1). By picking the non-fluorescent homozygous iscu-1 mutant offspring, we found that the iscu-1 deletion mutant worms were all growth arrested in the larval 1 (L1) stage before dying within a couple of days (Fig. 1a). Thus, we turned to the RNA interference (RNAi) approach to study the functional outcomes of suppressing ISCU-1 because unlike gene knockouts, RNAi typically produces hypomorphic phenotypes. Given that ISCU-1 has an essential role in development, we started iscu-1 RNAi treatment after the last larval stage (L4) to avoid the interference of ISCU-1 on development. Interestingly, initiating iscu-1 RNAi after the L4 stage (Fig. 1b) or even day 1 adulthood (Supplementary Fig. 1b) is sufficient to extend lifespan. This observation is different from the previous finding that RNAi of the mitochondrial electron transport chain signaling only promotes longevity during the developmental stage (Supplementary fig. 1c, d) [2]. In addition to the extended lifespan, adulthood-specific iscu-1 RNAi led to higher body bending frequency (Fig. 1c). Collectively, it appears that suppressing the functions of ISCU-1 during adulthood can promote longevity in C. elegans.

The [Fe-S] cluster assembly protein ISCU-1 suppresses longevity. a When the iscu-1(tm4941) mutant is balanced by an nT1 genetic balancer, the larval worms can develop normally into adulthood. However, the homozygous iscu-1 mutant worms are growth arrested in the L1 stage and die a few days later. Scale bar, 1000 μm. b To avoid the impact on development, iscu-1 RNAi is initiated after the L4 stage. iscu-1 RNAi treatment during adulthood significantly extends lifespan. p < 0.001, log-rank test. c iscu-1 RNAi treatment enhances body bending frequency. n ≥ 20, *p < 0.05, **p < 0.01, ***p < 0.001 unpaired two-tailed t test. d, e iscu-1 RNAi treatment from egg drastically promotes oxidative stress resistance in response to tBuOOH (6 mM) (d, stress assay performed at day 1), p < 0.001, log-rank test. However, adulthood-specific iscu-1 RNAi does not enhance oxidative stress resistance (e, stress assay performed at day 6). Of note, older (day 6) worms are more resistant to tBuOOH than young (day 1) worms. (f-g) iscu-1 RNAi treatment from egg significantly enhances the resistance to extracellular iron (f), p < 0.001, log-rank test. However, such a protective effect is not observed in worms treated with adulthood-specific iscu-1 RNAi (g). n ≥ 40, each experiment is repeated at least three times
Although ISCU-1 is required for normal development of both C. elegans and mice [20], a significant portion of iscu-1 RNAi-treated worms can still develop into adulthood even if iscu-1 RNAi was started from eggs (not shown), which could presumably be due to the hypomorphic nature of RNAi or maternal contribution. Notably, iscu-1 RNAi treatment from egg consistently led to greater lifespan extension compared with the adulthood-specific iscu-1 RNAi (Supplementary fig. 1b, c). Furthermore, after iscu-1 RNAi treatment, worms exhibited a shortened body length and reduced brood size (Supplementary fig. 1e, f), which are commonly observed in many long-lived mutants. Converse to the RNAi study, we generated a transgenic strain that overexpresses ISCU-1 driven by its endogenous promoter and found that ISCU-1 overexpression shortened lifespan (Supplementary fig. 1g). Thus, both RNAi and overexpression experiments support that ISCU-1 is an anti-longevity factor. In addition to ISCU-1, we examined another key component of the ISC assembly machinery, ISCS. The C. elegans cysteine desulfurase NFS-1 is the homolog of ISCS, and it shares ~ 70% sequence identity and > 90% sequence similarity with its human counterpart. Consistent with the result of iscu-1, nfs-1/iscs RNAi also significantly extended lifespan (Supplementary fig. 1h). Taken together, the ISC assembly machinery appears to suppress longevity.
ISCU-1 suppresses stress resistance during the developmental stage
Many anti-aging interventions also enhance stress resistance. Intriguingly, although iscu-1 RNAi treatment from egg drastically promoted the resistance to the oxidative stressor tert-Butyl hydroperoxide (tBuOOH) (Fig. 1d), adulthood-specific iscu-1 RNAi had no significant effect (Fig. 1e). Therefore, it seems that the developmental stage is particularly important for ISCU-1 to regulate stress resistance.
As the most common metal on earth, iron is an essential element for most, if not all, living species. On the other hand, labile iron is highly toxic to cells because of its high potency in generating ROS. Indeed, extracellular iron (FeCl3) is a potent stressor for worms and it killed worms in a dose- and duration-dependent fashion (Supplementary fig. 2a, b). Notably, the slope of the iron dose-response curve is very steep (Supplementary fig. 2b), suggesting that labile iron kills C. elegans within a narrow concentration window. In line with the oxidative stress assay, iscu-1 RNAi treatment from egg significantly enhanced survival upon iron stress (Fig. 1f) while adulthood-specific iscu-1 RNAi could not protect worms from excessive iron (Fig. 1g). In fact, adulthood-specific iscu-1 RNAi even rendered worms more susceptible to iron stress (Fig. 1g). Together, these results suggest that ISCU-1’s roles may vary between the developmental stage and adulthood stage in stress responses.
ISCU-1 modulates longevity in the intestine
After demonstrating that ISCU-1 can modulate longevity, we next set to study where it may act to modulate lifespan. By generating a transgenic reporter line that expressed a fluorescent protein mCherry driven by a 1.5-kb iscu-1 promoter, we found that ISCU-1 is mainly expressed in the intestine throughout development (Fig. 2a). In addition, ISCU-1 appears to have low expression in the anterior body wall muscle and hypodermis (Fig. 2a). Notably, the neuronal expression of ISCU-1 was undetectable.

ISCU-1 acts in the intestine to modulate lifespan. a The expression pattern of ISCU-1 is examined using a transcriptional reporter line Piscu-1::SL2::mCherry driven by a ~ 1.5 kb iscu-1 promoter. ISCU-1 is expressed throughout development, mainly in the intestine but also observed in the anterior body wall muscle and hypodermis. Scale bar, 100 μm. b The subcellular localization of ISCU-1 is investigated using translational reporter lines Pdpy-7::iscu-1::mCherry and Pges-1::iscu-1::mCherry that co-express Pdpy-7::mitoYFP and Pges-1::mitoYFP, respectively. The co-localization of ISCU-1::mCherry and mitoYFP suggests that ISCU-1 is expressed in the mitochondria. Scale bar, 10 μm. c The intestine-specific iscu-1 RNAi significantly extends lifespan. p<0.001, log-rank test. d–f The muscle- (d), hypodermis- (e), or neuron-specific (f) iscu-1 RNAi does not extend lifespan. g iscu-1 RNAi does not extend lifespan in the RNAi-deficient rde-1 mutant background. h The intestinal-specific expression of iscu-1 dsRNA is sufficient to extend lifespan. p = 0.009, log-rank test
Although [Fe-S] cluster assembly primarily occurs in the mitochondria, mammalian ISCU has been reported to exist in both mitochondria and cytosol [21]. To examine the subcellular localization of ISCU-1, we generated another transgenic reporter line co-expressing Pges-1::iscu-1::mCherry in which mCherry is fused in frame with the C terminus of ISCU-1 and Pges-1::mitoYFP in which the intestinal mitochondria are labeled by the mitochondrial outer membrane-targeting TOM20::YFP. The co-localization of ISCU-1::mCherry and mitoYFP supported that ISCU-1 is expressed in mitochondria (Fig. 2b). A similar result was also obtained for the hypodermal co-localization of ISCU-1::mCherry and mitoYFP (Fig. 2b). Therefore, ISCU-1 seems to be a mitochondrial protein in C. elegans.
Where does ISCU-1 act to modulate longevity? To address this question, we applied the well-established tissue-specific RNAi approach to knockdown the expression of iscu-1 in the intestine, muscle, hypodermis, and neuron, respectively [22]. Specifically, the rde-1 mutant is known to be resistant to RNAi and RDE-1 functions in a cell-autonomous fashion [22]. Thus, tissue-specific expression of the wild-type rde-1 cDNA in the rde-1 mutant background limits RNAi sensitivity to a specific tissue. Using this approach, we found that only the intestine-specific iscu-1 RNAi could extend lifespan whereas iscu-1 RNAi in other tissues had no effect on longevity (Fig. 2c–f). As a negative control, iscu-1 RNAi in the rde-1 mutant background (without wild-type rde-1 cDNA rescue) also failed to extend lifespan (Fig. 2g).
As the lifespan extension induced by intestine-specific iscu-1 RNAi is relatively small, we used another sid-1-mediated tissue-specific RNAi approach to provide further evidence that ISCU-1 modulates longevity in the intestine. Sid-1 encodes a channel protein for double-strand RNA (dsRNA) entry in C. elegans. Thus, although the sid-1(qt9) mutant is defective for systemic RNAi, tissue-specific expression of dsRNA will allow for cell-autonomous RNAi [23]. As expected, iscu-1 dsRNA driven by the intestine-specific ges-1 promoter significantly extended lifespan (Fig. 2h). Collectively, our results suggest that ISCU-1 may modulate longevity in the intestine.
ISCU-1 regulates cellular redox status
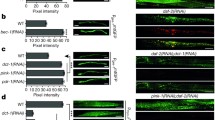
Due to their intrinsic chemical and electronic features, [Fe-S] clusters are highly sensitive to cellular redox status. We thus decided to examine whether ISCU-1 could modulate cellular redox status in C. elegans. Using the genetically encoded ratiometric fluorescent H2O2 sensor HyPer3 [24], we were able to measure the H2O2 level in live worms. Interestingly, iscu-1 RNAi treatment significantly decreased the intracellular H2O2 level (Fig. 3a, b). In line with the result obtained with Hyper3, we also used another well-established ROS sensor 2′,7′-dichlorofluorescin (DCFH2) [25], and found that iscu-1 RNAi-treated worms contained less ROS (Supplementary fig. 2c, d). Furthermore, worms treated with iscu-1 RNAi consumed significantly less oxygen (Supplementary fig. 2e). Therefore, it is possible that ISCU-1 might suppress longevity through elevated ROS generation. To test it, we added a well-established antioxidant vitamin C (10 mM) to the medium. Consistent with previous reports [26, 27], vitamin C significantly extended lifespan in wild-type worms (Fig. 3c). However, vitamin C no longer promoted longevity in iscu-1 RNAi-treated worms (Fig. 3d), indicating that iscu-1 RNAi and vitamin C might extend lifespan through a common pathway.

ISCU-1 modulates reactive oxygen species in C. elegans. a, b iscu-1 RNAi treatment significantly reduces the endogenous H2O2 level as revealed by the genetically encoded H2O2 indicator HyPer3. a Sample images. b Summary of imaging experiments as shown in (a). n ≥ 40, ***p < 0.001, unpaired two-tailed t test. Scale bar, 100 μm. c, d Although the antioxidant vitamin C significantly extends lifespan in wild-type animals (c), it does not promote longevity in iscu-1 RNAi-treated worms (d). p < 0.001 (WT vs WT + vitamin C), log-rank test
Mitochondrial fission underlies the anti-longevity effect of ISCU-1
As a central component of the mitochondrial [Fe-S] cluster assembly machinery, does ISCU-1 modulate mitochondrial dynamics? To examine it, we used a transgenic line Pges-1::mitoYFP in which the intestinal mitochondria are labeled by the yellow fluorescent protein (YFP). Consistent with a previous report [28], aged worms have more mitochondria than young worms (Fig. 4a, b). More interestingly, iscu-1 RNAi treatment significantly elevated the mitochondrial level (Fig. 4a, b and supplementary fig. 3). To complement our imaging experiment, we also quantified mitochondria amount by measuring the ratio of mitochondrial DNA copy number versus nuclear DNA copy number (mtDNA/nuDNA) with a qPCR method as described before [15]. In line with the imaging results, knockdown of iscu-1 increased the mitochondrial copy number (Fig. 4c). Together, these results suggest that ISCU-1 might suppress mitochondrial biogenesis. In support of this notion, our qPCR assay demonstrated that iscu-1 RNAi enhanced the expression levels of multiple mitochondrial biogenesis genes including cox-4, timm-17, hmg-5, and atp-5 (Fig. 4d) [28].

ISCU-1 regulates mitochondrial dynamics in C. elegans. a, b The intestinal mitochondria are labeled by TOM20::YFP driven by an intestine-specific promoter Pges-1. Day 10 worms exhibit a higher intestinal mitochondrial level compared with day 1 worms. More interestingly, iscu-1 RNAi significantly elevates the total mitochondrial level in the intestine. a Sample images. b Quantification of imaging experiments as shown in (a). n ≥ 15, *p < 0.05, **p < 0.01, ***p < 0.001, unpaired two-tailed t test. Scale bar, 100 μm. c The relative mitochondrial DNA copy number versus nuclear DNA copy number are quantified using qPCR. Consistent with imaging results, iscu-1 RNAi significantly elevates the mitochondrial level. n = 3, **p < 0.01, ****p < 0.0001, unpaired two-tailed t test. d iscu-1 RNAi elevates the expression level of multiple mitochondrial biogenesis genes. n = 4, *p < 0.05, **p < 0.01; ***p < 0.001, unpaired two-tailed t test. e Mitochondrial fusion is not required for the iscu-1 RNAi-induced lifespan extension as iscu-1 RNAi still significantly extends lifespan in the fusion-defective fzo-1(tm1133) mutant. p < 0.001, log-rank test. f Mitochondrial fission is required for the iscu-1 RNAi-induced lifespan extension as the fission-defective drp-1(tm1108) mutant abolishes the prolongevity effect of iscu-1 RNAi. g, h Mitophagy is not required for the iscu-1 RNAi-induced lifespan extension as iscu-1 RNAi treatment still significantly extends lifespan in the pink-1(ok3538) (g) and pdr-1(gk448) (h) mutants. p < 0.001, log-rank test
Mitochondrial dynamics is actively regulated by fusion and fission, which are both linked to lifespan regulation in C. elegans [29,30,31]. Mitochondrial outer membrane fusion is mediated by the membrane-anchored Mitofusin (FZO-1 in C. elegans) while inner membrane fusion is mediated by Opa1 [32]. By contrast, fission is mediated by a cytosolic dynamin-related protein Drp1 (DRP-1 in C. elegans) [32]. Interestingly, the fission-defective drp-1(tm1108) mutant, but not fusion-defective fzo-1(tm1133) mutant, completely suppressed the iscu-1 RNAi extended lifespan (Fig. 4e, f), supporting that mitochondrial fission is required for ISCU-1 in lifespan modulation.
In addition to fusion and fission, mitophagy plays an important role in mitochondrial homeostasis. As PTEN-induced kinase 1 (PINK1) and Parkin are important regulators of mitophagy [33], we next tested the potential involvement of C. elegans PINK1 (PINK-1) and Parkin (PDR-1) in the ISCU-1-modulated aging process. Interestingly, iscu-1 RNAi still significantly extended lifespan in the pink-1(ok3538) and pdr-1(gk448) mutant worms (Fig. 4g, h), arguing against the involvement of mitophagy. Therefore, among major regulators of mitochondrial dynamics, fission underlies the anti-longevity effect of ISCU-1.
Transcription factors downstream of ISCU-1 in lifespan modulation
Perturbation of mitochondria can often trigger the adaptive UPRmt which is known to modulate aging cell nonautonomously [3, 6, 7]. Indeed, iscu-1 RNAi treatment is a potent inducer of UPRmt as revealed by the well-established UPRmt reporter Phsp-6::GFP (Fig. 5a, b) [34]. Since the previous genome-wide RNAi screen for regulators of UPRmt did not identify iscu-1 [35], our current study revealed iscu-1 as a novel regulator of UPRmt. The bZip transcription factor ATFS-1 is a major player downstream of UPRmt [34]. Thus, we reasoned that ATFS-1 may act downstream of ISCU-1 to modulate lifespan. To our surprise, iscu-1 RNAi still extended lifespan in the atfs-1(gk3094) deletion mutant (Fig. 5c), indicating that ATFS-1 does not mediate ISCU-1’s function in lifespan regulation.

ISCU-1 modulates longevity through Nrf2 transcription factor SKN-1 and nuclear hormone receptor NHR-49. a, b iscu-1 RNAi strongly induces the expression of Phsp-6::gfp, a fluorescent reporter for UPRmt. a Sample images. b Quantification of imaging experiments as shown in (a). n ≥ 70, ***p < 0.001, unpaired two-tailed t test. Scale bar, 100 μm. c ATFS-1 is not required for the iscu-1 RNAi-induced lifespan extension as iscu-1 RNAi still extends lifespan in the atfs-1(gk3094) deletion mutant background. p = 0.004, log-rank test. d ISCU-1 modulates longevity through SKN-1 as iscu-1 RNAi no longer extends lifespan in the skn-1(zj15) mutant background. e ISCU-1 modulates longevity through NHR-49 as iscu-1 RNAi no longer extends lifespan in the nhr-49(nr2041) mutant background. f, g The skn-1(zj15) (f) and nhr-49(nr2041) (g) mutants largely suppress the iscu-1 RNAi-promoted oxidative stress resistance. h, i iscu-1 RNAi no longer enhances the iron resistance in the skn-1(zj15) (h) and nhr-49(nr2041) (i) mutant backgrounds
As most aging-related signaling pathways converge on several key transcription factors and nuclear hormone receptors [1, 36, 37], we next used a candidate gene approach and screened a host of aging-related transcription factors and nuclear hormone receptors, including daf-16 (Supplementary fig. 4b), pqm-1 (Supplementary fig. 4c), hsf-1 (Supplementary fig. 4d), hif-1 (Supplementary fig. 4e), xbp-1 (Supplementary fig. 4f), skn-1 (Fig. 5d), pha-4 (Supplementary fig. 4g), daf-12 (Supplementary fig. 4h), and nhr-49 (Fig. 5e). Among them, skn-1(zj15) and nhr-49(nr2041) were the only mutants that abolished the iscu-1 RNAi extended lifespan (Fig. 5d, e). Notably, PHA-4 acts downstream of the dietary restriction pathway to promote longevity [38]. Although iscu-1 RNAi only triggered a relatively modest but significant lifespan extension in the pha-4 RNAi-treated worms, iscu-1 RNAi still greatly extended lifespan in the dietary-restricted eat-2(ad465) mutant (Supplementary fig. 4i), suggesting that ISCU-1 does not modulate aging through the dietary restriction pathway.
In addition to the lifespan assay, we performed oxidative stress and iron stress assays to examine whether SKN-1 and NHR-49 also mediate ISCU-1’s function in stress responses. While iscu-1 RNAi treatment greatly enhanced the resistance to tBuOOH and extracellular iron (Fig. 1d, f), the skn-1(zj15) and nhr-49(nr2041) mutants largely suppressed stress resistance (Fig. 5f–i), supporting that SKN-1 and NHR-49 are also involved in ISCU-1’s role in stress resistance. Taken together, SKN-1 and NHR-49 seem to act downstream of ISCU-1 in lifespan modulation and stress response.
ISCU-1 suppresses the nuclear activities of SKN-1 and NHR-49
After finding that the skn-1 and nhr-49 mutants could suppress ISCU-1’s role in aging and stress resistance, we reasoned whether ISCU-1 can modulate the activities of these two transcription factors. gst-4 is a well-established target gene of SKN-1 and the reporter line dvIs19[(pAF15)gst-4p::GFP::NLS] is commonly used to monitor the SKN-1 activity [39, 40]. Using this reporter line, we found that iscu-1 RNAi significantly enhanced the expression of Pgst-4::GFP (Fig. 6a, b), indicating that ISCU-1 suppresses SKN-1. Furthermore, we performed the qPCR assay and probed two target genes of NHR-49, acs-1 and ech-1.2 [41,42,43]. Consistent with the reporter result of SKN-1, iscu-1 RNAi elevated the expression levels of acs-1 and ech-1.2 (Fig. 6c), supporting that ISCU-1 also suppresses NHR-49.

ISCU-1 suppresses SKN-1 and NHR-49 without changing their subcellular localization patterns. a, b iscu-1 RNAi significantly induces the expression of Pgst-4::gfp, a fluorescent reporter for the activation of SKN-1. a Sample images. b Quantification of imaging experiments as shown in (a). n ≥ 71, ***, p < 0.001, unpaired two-tailed t test. Scale bar, 100 μm. c iscu-1 RNAi enhances the expression levels of NHR-49 target genes acs-2 and ech-1.2 as revealed by the qPCR assay. n = 4, *p < 0.05, ****p<0.0001, unpaired two-tailed t test. d, e iscu-1 RNAi does not cause major changes in the subcellular distribution patterns of SKN-1 or NHR-49 as revealed by their translational reporter lines ldIs7[skn-1b/c::GFP + rol-6(su1006)] and glmEx5[nhr-49p::nhr-49::gfp;myo-2p::mCherry], respectively. Scale bar, 100 μm
Many aging-related transcription factors are present in both the nucleus and cytoplasm. Longevity modulators may regulate these downstream transcription factors by promoting their nuclear translocation [44]. To our surprise, as revealed by the ldIs7[skn-1b/c::GFP + rol-6(su1006)] and glmEx5[nhr-49p::nhr-49::gfp;myo-2p::mCherry] translational reporter lines, we did not observe major changes in the subcellular distribution patterns of either SKN-1 or NHR-49 after worms were treated with iscu-1 RNAi (Fig. 6d, e). It should be noted that the nuclear activity of DAF-16/FOXO, another major transcription factor involved in longevity, can also be regulated by multiple aging-related factors (e.g., SMK-1, HCF-1, EAK-7, and the TRPA-1 pathway) without causing its nuclear accumulation [13, 45,46,47]. Thus, it is possible that ISCU-1 may only regulate the functions of SKN-1 and NHR-49 that are already present in the nucleus.
PMK-1 and PGAM-5 act downstream of ISCU-1 in longevity regulation
[Fe-S] cluster assembly occurs in mitochondria while both NHR-49 and SKN-1 modulate aging in the nucleus. How is ISCU-1 coupled with nuclear transcription factors? Notably, the stress-inducible p38 MAPK PMK-1 is well established to link cellular stress to SKN-1 activation [48]. Thus, we reasoned that PMK-1 might act as a signal transducer between ISCU and SKN-1. Indeed, the pmk-1(km25) deletion mutant abolished iscu-1 RNAi-induced lifespan extension (Fig. 7a), indicating that the stress-inducible p38 MAPK PMK-1 may act downstream of ISCU-1.

PMK-1 and PGAM-5 transduce the longevity signal downstream of ISCU-1. a The pmk-1(km25) mutant abolishes the iscu-1 RNAi-induced lifespan extension. b pgam-5 RNAi suppresses the iscu-1 RNAi treatment-induced lifespan extension. c, d A mitochondria dye MitoTracker™ Red CMXRos is used to observe the mitochondrial morphology with or without iscu-1 RNAi treatment. The knockdown of ISCU-1 seems to promote the fragmentation of mitochondria as more mitochondrial puncta and less mitochondrial network are observed after iscu-1 RNAi. Scale bar, 10 μm. e, h Similar to iscu-1 RNAi, the overexpression of PGAM-5 also promotes the fragmentation of mitochondria (f). In the drp-1 mutant background, significantly less mitochondrial puncta are observed (g), and the drp-1 mutant suppresses the mitochondrial morphology phenotype of PGAM-5 overexpression (h). Scale bar, 10 μm. i A working model of the underlying mechanism by which ISCU-1 modulates longevity
The mitochondrial outer membrane Serine/Threonine protein phosphatase PGAM5 (PGAM-5 in C. elegans) is an important regulator of multiple necrotic death pathways and aging [49, 50]. Very interestingly, a subset of SKN-1 directly binds to PGAM-5, by which SKN-1 can sense mitochondrial function and stress [51]. Moreover, PGAM5 recruits and activates the mitochondrial fission factor Drp1 [49] and DRP-1/Drp1 is required for the iscu-1 RNAi extended lifespan (Fig. 4f). Lastly, PGAM5 has been shown to directly activate the apoptosis signal-regulating kinase 1 (ASK1) and its downstream p38 MAPK [52]. Based on all of this evidence, we hypothesized that PGAM-5 might act downstream of ISCU-1. Consistent with this notion, pgam-5 RNAi greatly suppressed the lifespan extension induced by iscu-1 RNAi (Fig. 7b). Moreover, we performed mitochondrial morphology staining experiments, and found that compared with the control condition (Fig. 7c, e) both iscu-1 RNAi and PGAM-5 overexpression promoted mitochondrial puncta formation (Fig. 7d, f), suggesting that ISCU-1 and PGAM-5 may act in a common pathway. In the fission-defective drp-1 mutant background, significantly less mitochondrial puncta were observed (Fig. 7g). Importantly, the drp-1 mutant suppressed the mitochondrial morphology phenotype of PGAM-5 overexpression (Fig. 7h). Thus, DRP-1 seems to act downstream of PGAM-5 in the ISCU-1 pathway. Taken together, PMK-1 and PGAM-5 may transduce the longevity signal downstream of ISCU-1 (Fig. 7i). On the other hand, more future studies are clearly required to dissect the genetic pathway that links ISCU-1 to the nuclear hormone receptor NHR-49.
Discussion
In the present study, using C. elegans as a model organism, we investigated the novel roles of mitochondrial ISC machinery in aging and stress response. Interestingly, while ISCU-1 is required for larval development, it shortens lifespan during adulthood. Thus, as discovered for several longevity-related signaling molecules [53, 54], ISCU-1 may represent another example of antagonistic pleiotropy - genes beneficial early in life at the cost of aging. On the other hand, although iscu-1 RNAi from egg protects worms from Fe3+ stress, adulthood-specific iscu-1 RNAi renders worms more sensitive (Fig. 1f, g). Thus, it seems that ISCU-1 may have distinct roles in stress responses during different life stages. It is possible that ISCU-1 might differentially regulate gene expressions in larva and adult. Notably, during temperature modulation of aging DAF-16 and TRPA-1 act in larva to shorten lifespan but extend lifespan in adulthood, and the underlying mechanism also involves differential gene expressions in larva and adult [55].
An interesting observation of our current study is that while adulthood-specific RNAi against ISCU-1 extends lifespan, it does not promote stress resistance (Fig. 1b, e, g). In this regard, the longevity role of ISCU-1 appears to be dissociated from its role in stress defense. Notably, although longevity and stress resistance are often positively correlated, much evidence has shown that they are physiologically distinct and can be uncoupled [56,57,58]. For example, the C. elegans transcription elongation and splicing factor TCER-1 promotes lifespan but widely represses stress resistance [57]. Additionally, genetic disruption of the glycerol-3-phosphate dehydrogenases gpdh-1 or gpdh-2 can also extend lifespan but exhibit decreased stress resistance [58]. Thus, stress resistance may not be simply treated as a surrogate of longevity.
Due to their biochemical nature, [Fe-S] clusters are intrinsically sensitive to intracellular ROS. In this regard, many [Fe-S] cluster-containing proteins may in principle act as ROS sensors. Notably, ROS modulates aging in a complex manner. Our data showed that iscu-1 RNAi treatment greatly reduced the ROS level, suggesting that ISCU-1 might accelerate aging through increased ROS generation. The biogenesis of [Fe-S] clusters is a complicated process. Although the initial assembly of [Fe-S] clusters occurs in the mitochondria through ISC machinery, maturation of [Fe-S] cluster-containing proteins mainly happens in the cytoplasm through the cytosolic iron-sulfur protein assembly (CIA) machinery. It remains to be determined whether other ISC machinery components or CIA machinery also play a role in aging and stress response.
Mitochondrion-derived signaling has a great impact on aging. A mild inhibition of mitochondrial electron transport chain extends lifespan in multiple species [36]. Very interestingly, the electron transport chain signaling modulates longevity cell nonautonomously and both neuron- and intestine-specific RNAi of electron transport chain can extend lifespan [3]. By contrast, ISCU-1 is primarily expressed in the intestine and neuronal RNAi of iscu-1 does not affect longevity (Fig. 2). Furthermore, the electron transport chain is known to regulate lifespan during the developmental stage and initiating RNAi of electron transport chain components during adulthood does not extend lifespan (Supplementary fig. 1d) [2]. This is also different from ISCU-1 as the adulthood-specific iscu-1 RNAi significantly promotes longevity (Fig. 1b and Supplementary fig. 1b). Notably, in a genetic screen of genes essential for growth and development, it was found that inactivation of several components of mitochondrial electron transport chain (e.g., atp-2, atp-3, phi-37 and nuo-1) in the adulthood stage also extend lifespan [59]. Thus, mitochondrial signaling appears to be able to modulate aging during both the developmental stage and adulthood.
Taken together, in the present study, we identified a novel function of the mitochondrial [Fe-S] cluster assembly protein ISCU-1 in longevity modulation and stress response. Moreover, we found that the nuclear factors SKN-1 and NHR-49 act downstream of ISCU-1. These findings establish ISCU-1 as a novel mitochondrion-derived signaling molecule that modulates aging and stress response. Since ISCU-1 and [Fe-S] cluster assembly machinery are highly conserved throughout evolution, our results may provide important insights into various human diseases due to mutations in key factors required for [Fe-S] cluster biogenesis, transport, and maturation.
References
Kenyon CJ. The genetics of ageing. Nature. 2010;464(7288):504–12.
Dillin A, et al. Rates of behavior and aging specified by mitochondrial function during development. Science. 2002;298(5602):2398–401.
Durieux J, Wolff S, Dillin A. The cell-non-autonomous nature of electron transport chain-mediated longevity. Cell. 2011;144(1):79–91.
Beckman KB, Ames BN. The free radical theory of aging matures. Physiol Rev. 1998;78(2):547–81.
Ristow M, Schmeisser S. Extending life span by increasing oxidative stress. Free Radic Biol Med. 2011;51(2):327–36.
Berendzen KM, et al. Neuroendocrine coordination of mitochondrial stress signaling and proteostasis. Cell. 2016;166(6):1553–1563.e10.
Zhang Q, et al. The mitochondrial unfolded protein response is mediated cell-non-autonomously by retromer-dependent Wnt signaling. Cell. 2018;174(4):870–883.e17.
Sebastian D, Palacin M, Zorzano A. Mitochondrial dynamics: coupling mitochondrial fitness with healthy aging. Trends Mol Med. 2017;23(3):201–15.
Lill R, Muhlenhoff U. Iron-sulfur protein biogenesis in eukaryotes: components and mechanisms. Annu Rev Cell Dev Biol. 2006;22:457–86.
Johnson DC, et al. Structure, function, and formation of biological iron-sulfur clusters. Annu Rev Biochem. 2005;74:247–81.
Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974;77(1):71–94.
Xiao R, et al. RNAi interrogation of dietary modulation of development, metabolism, behavior, and aging in C. elegans. Cell Rep. 2015;11(7):1123–33.
Xiao R, et al. A genetic program promotes C. elegans longevity at cold temperatures via a thermosensitive TRP channel. Cell. 2013;152(4):806–17.
Zhang B, et al. Brain-gut communications via distinct neuroendocrine signals bidirectionally regulate longevity in C. elegans. Genes Dev. 2018;32(3–4):258–70.
Rooney JP, et al. PCR based determination of mitochondrial DNA copy number in multiple species. Methods Mol Biol. 2015;1241:23–38.
Yoon DS, Lee MH, Cha DS. Measurement of intracellular ROS in Caenorhabditis elegans using 2 ',7 '-dichlorodihydrofluorescein diacetate. Bio-Protocol. 2018;8(6):e2774.
Han SM, et al. Secreted VAPB/ALS8 major sperm protein domains modulate mitochondrial localization and morphology via growth cone guidance receptors. Dev Cell. 2012;22(2):348–62.
Ast T, et al. Hypoxia rescues frataxin loss by restoring iron sulfur cluster biogenesis. Cell. 2019;177(6):1507–1521.e16.
Kim W, et al. OrthoList 2: a new comparative genomic analysis of human and Caenorhabditis elegans genes. Genetics. 2018;210(2):445–61.
Nordin A, et al. Tissue-specific splicing of ISCU results in a skeletal muscle phenotype in myopathy with lactic acidosis, while complete loss of ISCU results in early embryonic death in mice. Hum Genet. 2011;129(4):371–8.
Tong WH, Rouault T. Distinct iron-sulfur cluster assembly complexes exist in the cytosol and mitochondria of human cells. EMBO J. 2000;19(21):5692–700.
Qadota H, et al. Establishment of a tissue-specific RNAi system in C. elegans. Gene. 2007;400(1–2):166–73.
Winston WM, Molodowitch C, Hunter CP. Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science. 2002;295(5564):2456–9.
Bilan DS, et al. HyPer-3: a genetically encoded H(2)O(2) probe with improved performance for ratiometric and fluorescence lifetime imaging. ACS Chem Biol. 2013;8(3):535–42.
Bromme HJ, et al. DCFH2 interactions with hydroxyl radicals and other oxidants--influence of organic solvents. Exp Gerontol. 2008;43(7):638–44.
De Magalhaes Filho CD, et al. Visible light reduces C. elegans longevity. Nat Commun. 2018;9(1):927.
Dallaire A, et al. Expression profile of Caenorhabditis elegans mutant for the Werner syndrome gene ortholog reveals the impact of vitamin C on development to increase life span. BMC Genomics. 2014;15:940.
Palikaras K, Lionaki E, Tavernarakis N. Coordination of mitophagy and mitochondrial biogenesis during ageing in C. elegans. Nature. 2015;521(7553):525–8.
Weir HJ, et al. Dietary restriction and AMPK increase lifespan via mitochondrial network and peroxisome remodeling. Cell Metab. 2017;26(6):884–896.e5.
Yang CC, et al. The dynamin-related protein DRP-1 and the insulin signaling pathway cooperate to modulate Caenorhabditis elegans longevity. Aging Cell. 2011;10(4):724–8.
Zhang Y, et al. Neuronal TORC1 modulates longevity via AMPK and cell nonautonomous regulation of mitochondrial dynamics in C. elegans. Elife. 2019;8:e49158.
Westermann B. Mitochondrial dynamics in model organisms: what yeasts, worms and flies have taught us about fusion and fission of mitochondria. Semin Cell Dev Biol. 2010;21(6):542–9.
Nguyen TN, Padman BS, Lazarou M. Deciphering the molecular signals of PINK1/Parkin mitophagy. Trends Cell Biol. 2016;26(10):733–44.
Nargund AM, et al. Mitochondrial import efficiency of ATFS-1 regulates mitochondrial UPR activation. Science. 2012;337(6094):587–90.
Bennett CF, et al. Activation of the mitochondrial unfolded protein response does not predict longevity in Caenorhabditis elegans. Nat Commun. 2014;5:3483.
Riera CE, et al. Signaling networks determining life span. Annu Rev Biochem. 2016;85:35–64.
Antebi A. Nuclear receptor signal transduction in C. elegans. WormBook. 2015:1–49. https://doi.org/10.1895/wormbook.1.64.2.
Panowski SH, et al. PHA-4/Foxa mediates diet-restriction-induced longevity of C. elegans. Nature. 2007;447(7144):550–5.
Steinbaugh MJ, et al. Lipid-mediated regulation of SKN-1/Nrf in response to germ cell absence. Elife. 2015;4:e07836.
Robida-Stubbs S, et al. TOR signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO. Cell Metab. 2012;15(5):713–24.
Van Gilst MR, et al. Nuclear hormone receptor NHR-49 controls fat consumption and fatty acid composition in C. elegans. PLoS Biol. 2005;3(2):e53.
Van Gilst MR, Hadjivassiliou H, Yamamoto KR. A Caenorhabditis elegans nutrient response system partially dependent on nuclear receptor NHR-49. Proc Natl Acad Sci U S A. 2005;102(38):13496–501.
Fredens J, et al. Quantitative proteomics by amino acid labeling in C. elegans. Nat Methods. 2011;8(10):845–7.
Landis JN, Murphy CT. Integration of diverse inputs in the regulation of Caenorhabditis elegans DAF-16/FOXO. Dev Dyn. 2010;239(5):1405–12.
Wolff S, et al. SMK-1, an essential regulator of DAF-16-mediated longevity. Cell. 2006;124(5):1039–53.
Alam H, et al. EAK-7 controls development and life span by regulating nuclear DAF-16/FoxO activity. Cell Metab. 2010;12(1):30–41.
Li J, et al. Caenorhabditis elegans HCF-1 functions in longevity maintenance as a DAF-16 regulator. PLoS Biol. 2008;6(9):e233.
Blackwell TK, et al. SKN-1/Nrf, stress responses, and aging in Caenorhabditis elegans. Free Radic Biol Med. 2015;88(Pt B):290–301.
Wang Z, et al. The mitochondrial phosphatase PGAM5 functions at the convergence point of multiple necrotic death pathways. Cell. 2012;148(1–2):228–43.
Borch Jensen M, et al. PGAM5 promotes lasting FoxO activation after developmental mitochondrial stress and extends lifespan in Drosophila. eLife. 2017;6:e26952.
Paek J, et al. Mitochondrial SKN-1/Nrf mediates a conserved starvation response. Cell Metab. 2012;16(4):526–37.
Takeda K, et al. Mitochondrial phosphoglycerate mutase 5 uses alternate catalytic activity as a protein serine/threonine phosphatase to activate ASK1. Proc Natl Acad Sci U S A. 2009;106(30):12301–5.
Blagosklonny MV. Revisiting the antagonistic pleiotropy theory of aging: TOR-driven program and quasi-program. Cell Cycle. 2010;9(16):3151–6.
Ezcurra M, et al. C.elegans eats its own intestine to make yolk leading to multiple senescent pathologies. Curr Biol. 2018;28(20):3352.
Zhang B, et al. Environmental temperature differentially modulates C. elegans longevity through a thermosensitive TRP channel. Cell Rep. 2015;11(9):1414–24.
Dues DJ, et al. Uncoupling of oxidative stress resistance and lifespan in long-lived isp-1 mitochondrial mutants in Caenorhabditis elegans. Free Radic Biol Med. 2017;108:362–73.
Amrit FRG, et al. The longevity-promoting factor, TCER-1, widely represses stress resistance and innate immunity. Nat Commun. 2019;10(1):3042.
Dues DJ, et al. Resistance to Stress can be experimentally dissociated from longevity. J Gerontol A Biol Sci Med Sci. 2019;74(8):1206–14.
Curran SP, Ruvkun G. Lifespan regulation by evolutionarily conserved genes essential for viability. PLoS Genet. 2007;3(4):e56.
Acknowledgements
We appreciate the Caenorhabditis Genetic Center, which is supported by the National Institutes of Health Office of Research Infrastructure Programs (P40 OD010440), Dr. Arjumand Ghazi (University of Pittsburgh), and the National BioResource Project (Tokyo, Japan) for providing strains.
Funding
This work was supported by grants from the National Institute on Aging (AG063766 and AG028740), the American Cancer Society (RSG-17-171-01-DMC), and the American Federation for Aging Research (AFAR).
Author information
Authors and Affiliations
Contributions
R.X. and Y.S. conceived and designed the study; Y.S. performed most experiments with the assistance of G.Y., K.C., S.C., M.O., S.M.H., and C.L.; R.X. and Y.S. analyzed and interpreted results; R.X. and Y.S. wrote the manuscript. All authors discussed the results and commented on the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
Supplementary Fig. 1 ISCU-1 suppresses longevity during adulthood. (a) iscu-1 RNAi extends lifespan in the absence of FUdR. p=0.004, log-rank test. (b) Initiating iscu-1 RNAi from day 1 adulthood is sufficient to significantly extend lifespan. p<0.001 (vector RNAi vs iscu-1 RNAi), log-rank test. (c) RNAi of iscu-1 or cyc-1 (an essential component of the mitochondrial electron transport chain) from egg leads to significant lifespan extension. p<0.001, log-rank test. (d) Adulthood-specific RNAi of iscu-1, but not cyc-1, significantly promotes longevity, suggesting that ISCU-1 and CYC-1 may have different temporal actions in lifespan regulation. p<0.001 (vector RNAi vs iscu-1 RNAi), log-rank test. (e) Worms treated with iscu-1 RNAi are shorter than the ones treated with control vector RNAi. n≥27, NS, not significant; ***, p<0.001, unpaired two-tailed t test. (f) Worms treated with iscu-1 RNAi produce fewer progenies than the ones treated with control vector RNAi. n≥6, ***, p<0.001, unpaired two-tailed t test. (g) ISCU-1 overexpression driven by its endogenous promoter significantly shortens lifespan. WT denotes wild type. p<0.001, log-rank test. (h) Similar to iscu-1, the knockdown of the C. elegans ISCS nfs-1 also significantly extends lifespan. p<0.001, log-rank test. Supplementary Fig. 2 ISCU-1 in stress responses. (a) Extracellular iron kills C. elegans in a dose- and duration-dependent manner. n≥40, each experiment is repeated at least twice. (b) The dose-response curve of extracellular iron that kills worms within 24 hours. (c-d) iscu-1 RNAi-treated worms (c: treated till day 1 adulthood; d: treated till day 10 adulthood) contain less ROS as revealed by the ROS dye DCFH2. ***, p<0.001, unpaired two-tailed t test. (e) Worms treated with iscu-1 RNAi consume less oxygen than the ones treated with control vector RNAi. n≥12, NS, not significant; ***, p<0.001, unpaired two-tailed t test. Supplementary Fig. 3 iscu-1 RNAi increases the mitochondria amount. Confocal images of the intestinal mitochondria (labeled by Pges-1::mitoYFP) with or without iscu-1 RNAi treatment. iscu-1 RNAi elevates the mitochondrial amount. Scale bar, 10 μm. Supplementary Fig. 4 Genetic dissection of the aging-related nuclear factors downstream of ISCU-1. (a) iscu-1 RNAi treatment significantly extends lifespan in the wild type (WT) worms. p<0.001, log-rank test. (b-h) iscu-1 RNAi treatment still significantly extends lifespan in the daf-16(mgDf47) (b), hsf-1(sy441) (d), hif-1(ia4) (e), xbp-1(zc12) (f), daf-12(rh61rh411) (g) mutants as well as pqm-1 RNAi (c)- and pha-4 RNAi (h)-treated worms, indicating that these aging-related factors are not required for iscu-1 RNAi to delay aging. p<0.001, log-rank test. (i) iscu-1 RNAi treatment greatly extends lifespan in the dietary-restricted eat-2(ad465) mutant, supporting that ISCU-1 does not regulate longevity through dietary restriction pathway. p<0.001, log-rank test. (PDF 314 kb)
ESM 2
(DOCX 46 kb)
About this article

Cite this article
Sheng, Y., Yang, G., Casey, K. et al. A novel role of the mitochondrial iron-sulfur cluster assembly protein ISCU-1/ISCU in longevity and stress response. GeroScience 43, 691–707 (2021). https://doi.org/10.1007/s11357-021-00327-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11357-021-00327-z