
Abstract
Pontederia cordata is a heavy metal accumulator, while the heavy metal tolerance mechanisms of this plant are not well understood. Hydroponic experiments were used to assess the effects of Cd2+ on antioxidative activities, osmoregulatory substances and photosynthesis in leaves. Exposure of 5 mg L−1 Cd2+ for 7 days, the photosynthetic apparatus functioned normally and sustained a relatively high photosynthetic rate, and good growth was observed. Under 50 and 75 mg L−1 Cd2+, accelerated lipid peroxidation and increased peroxidase activity (POD; E.C.1.11.1.7) were detected, while no significant differences were observed in superoxide dismutase (SOD; E.C.1.15.1.1) and catalase (CAT; E.C.1.11.1.6) activities, as well as in lutein, ascorbic acid, and glutathione contains of leaves. Proline content increased, while soluble sugar and soluble protein contents decreased under 75 mg L−1 Cd2+. Cd2+ at different concentrations induced a reduction in carotenoid, total carotenoid, and ascorbic acid-dehydroascorbate contents. A significant increase in phytochelatin content was induced by 75 mg L−1. Chlorophyll content decreased under Cd stress and disturbed photosynthesis, causing dramatic reductions in photosynthetic parameters. Stomatal closure was responsible for a reduced photosynthetic rate under Cd2+ exposure. Cd2+ concentrations of no less than 25 mg L−1 disorganized the photosynthetic apparatus, induced the partial closure, and decreased activity of the photosystem II (PS II) reaction center, thus disturbing light conversion and utilization, thereby decreasing the photosynthetic efficiency in PS II.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Heavy-metal contamination is a severe global environmental concern. As a non-essential nutrient element, Cd is one of the common heavy-metal pollutants in the ecological environment, originating from industrial activities, the application of Cd-rich phosphate fertilizers, and the leather industry (McLaughlin et al. 1999; Groppa et al. 2012). In China, Cd concentrations in polluted water usually range from 0.7 to 40 mg L−1 and can even reach 100 mg L−1 in discharged wastewater as reported (Deng et al. 2014; Dou et al. 2017), and plenty of typical cases involved in Cd contamination also frequently occurred, and Cd concentration in edible parts was as high as 2.58 ~ 5.00 mg kg−1 in the presence of zinc (Simmons et al. 2003). Compared with other heavy metals, Cd has strong biological toxicity, is resistant to degradation, and demonstrates high chemical activity. It is easily absorbed by roots, thus constituting a threat to the health of humans and other organisms through its accumulation in the food chain and biological amplification (Grant et al. 2008; Lim et al. 2016). Many plants suffer with Cd toxicity, as their metal balance network in the cells is incapable of dealing with Cd2+ at higher concentrations (Krämer and Clemens 2005). Excessive Cd accumulation in the cells produces abundant reactive oxygen species (ROS), including hydrogen peroxide (H2O2), singlet oxygen (1O2), superoxide radicals (O2−), and hydroxyl radicals (∙OH). This leads to oxidative stress, which impairs membrane structure and function and disturbs photosynthesis and respiration, thereby resulting in the stunted growth and even death in plants (Singh and Prasad 2014; Muradoglu et al. 2015). To scavenge excess reactive oxygen species (ROS) toxic heavy metal stress, plants have evolved an array of antioxidant defensive systems consisting of enzymatic and non-enzymatic reaction systems like superoxide dismutase (SOD; E.C.1.15.1.1), peroxidase (POD; E.C.1.11.1.7), catalase (CAT; E.C.1.11.1.6), and ascorbate peroxidase (APX; E.C.1.11.1.11). SOD convert O2− to H2O2 that catalyzed to O2 and H2O by CAT and POD, thus suppressing its excessive accumulation in the cells. Simultaneously, the latter is composed of ascorbic acid (AsA), carotenoids (Cars), and glutathione (GSH), which are involved in the scavenging of ROS in cells and improve the physiological functioning of plants, reinforcing the plants for its functioning against heavy metal stress (Manging et al. 2003; Wu et al. 2004). Furthermore, proline, soluble sugars, and soluble proteins are osmoregulatory substances, the increased contents of which decrease the water potential in cells and strengthen the capacity of the cells to hold water. These are crucial mechanisms for the adaptation of plants to abiotic stresses (Munns 1988; Ghoulam et al. 2002; Farhangi-Abriza and Torabianb 2017).
As demonstrated, Cd can disturb chlorophyll biosynthesis, disorganize chloroplast structure, and decrease key enzyme activities (such as RuBisCO and NADPH) in photosynthesis, as well as inhibit photosynthetic and respiratory electron transport activities, restraining photosynthesis and plant growth (Groppa et al. 2012; Deng et al. 2014; Bashirl et al. 2015; Srivastava et al. 2017). The sensitivity of the photosynthetic apparatus to Cd varies across plant species. The Chl a content in leaves of Robinia pseudoacacia remained at a constant level when the seedlings were exposed to 250 mg L−1 Cd for 10 days, and a marked increase under increased Cd was also observed (Dezhban et al. 2015). However, Chl a in leaves of Canna indica seedlings decreased by 38.57% under 3.2 mg L−1 Cd exposure for 9 days. In recent years, Chl a fluorescence measurements have been widely used to elucidate the physiological state of the photosynthetic apparatus in plants under adverse conditions. This technique cannot damage cell integrity. Based on Chl a fluorescence, Elsholtzia argyi was demonstrated tolerance to moderate Cd, while the seedlings of Sedum alfredii and Robinia pseudoacacia are more tolerate to higher Cd concentrations (Zhou and Qiu 2005; Dezhban et al. 2015; Li et al. 2015). Pontederia cordata, which is native to the tropical and temperate regions of the new world, is mainly distributed in marshes, ditches, lakes, ponds, and shallows. It is a macrophyte with a stout rhizome, developed roots, a large biomass, and high ornamental value, and is nowadays considered to be a candidate for the revegetation and reestablishment of wetlands. Reimer and Duthie (1993) argued that P. cordata is a heavy-metal accumulator with immobilizing zinc and chromium in its roots. However, how this plant copes with heavy metal stress remains poorly understood. We thus investigated the effects of Cd at various concentrations on the physiological metabolic activities in the leaves of P. cordata with antioxidative enzymes (SOD, POD, CAT); non-enzymatic antioxidants (AsA, Car); soluble protein, soluble sugar, and proline; contents of non-protein thiol (NPT), GSH, and phytochelatins (PCs); pigments; and Chl a fluorescence measurements. The present investigation will determine the toxic effects of Cd on metabolic processes in the leaves and clarify the detoxifying mechanism of the plant to Cd stress, thus providing a theoretical reference for the application of the plant in the remediation of heavy-metal-contaminated water and wetlands.
Materials and methods
Plant material and growth conditions
Rhizomes of P. cordata were collected in late autumn 2017 from Tianjing Aquatic Plant Gardens, Hangzhou, Zhejiang Province, and were cultivated at the National Landscape Architecture Experimental Teaching Demonstration Center, Nanjing Forestry University. Tillers from the rhizomes cultivated for 2 months in the following spring were selected and adaptively cultivated in 2 L 1/2 Hoagland’s solution in a greenhouse for 20 days. The plants were then transferred to 2 L 1/2 Hoagland’s solution containing 0, 5, 25, 50, and 75 mg L−1 Cd (calculated as ion) supplied with CdCl2·2.5H2O and cultivated for 7 days. There were totally 45 plants with three replications in each treatment. During the treatments, the nutrition was supplemented to the initial volume with 1/2 Hoagland’s solution. For determining the various parameters, 1 ~ 2 mature leaves from the apex of the plants were used.
Determination of thiobarbituric acid reactive substance content
Fresh leaves were immersed in 20-mL deionized water and kept for 24 h at room temperature before initial conductivity (C1) was measured. The samples were then maintained at 100 °C for 20 min. Final conductivity (C2) was measured after the samples had cooled down to room temperature. Membrane leakage was expressed as relative electron conductivity.
Thiobarbituric acid reactive substance (TBARS) content was assayed according to Li (2000), and the absorbance at 600, 532, and 450 nm was determined using an ultraviolet visible spectrophotometer (Lambda 25, Perkin-Elmer, Waltham, USA).
Determination of antioxidative activity
Fresh leaves were collected from each treatment for antioxidant enzyme activity determination. All enzyme activities were measured with an ultraviolet visible spectrophotometer (UV-VIS) (Lambda 25, Perkin-Elmer, Waltham, USA). SOD activity was assayed by monitoring the inhibition of nitroblue tetrazolium photochemical reduction (Beyer and Fridovich 1987). One unit of SOD activity was defined as the amount of enzyme required for 50% inhibition of the reduction rate of NBT under light, determined at 560 nm (Aebi 1984). CAT activity was assayed by measuring the rate of continuous decomposition of hydrogen peroxide (H2O2) according to the method of Aebi (1984). POD activity was assayed according to the method of Upadhyaya et al. (1985). One unit of POD activity was defined as the H2O2 decomposition per minute. Increases in the absorbance as a result of guaiacol were assayed at 470 nm.
AsA and AsA-DHA contents were determined according to the method of Kampfenkel et al. (1995): fresh leaves were homogenized with 6% trichloroacetic acid, and the absorbance at 525 nm was determined.
Fresh leaves were immersed in a mixture of ethanol and acetone (1:1, V/V) in the dark for Car extraction. Car, Lutein, and Car T contents were determined using the UV-VIS. The absorbance at 470, 474, 485, 642.5, 649, and 663 nm was according to the method of Xu et al. (2013).
Proline, soluble sugar, and protein content analysis
Fresh leaves were homogenized with 3% trichloroacetic acid (w/v), and the absorbance at 520 nm was determined using an UV-VIS spectrophotometer according to the method of Bates et al. (1973).
Soluble sugar and soluble protein contents were determined according to the method of anthrone colorimetry and Coomassie brilliant blue staining, respectively, and the absorbance at 630 and 595 nm was separately determined using the UV-VIS according to the method of Bradford (1976).
Determination of PC content
Non-protein thiol content was determined according to the method of Rama and Prasad (1998), and fresh leaves were homogenized with 5% sulfosalicylic acid. GSH content was measured in accordance with the procedure of Anderson (1985), and the leaves were grounded into a homogenate with ethylene diamine tetraacetic acid-trichloroacetic acid (EDTA-TCA). The absorbance at 412 nm was determined using the UV-VIS. The PC content was calculated according to the method of Bhargava et al. (2005).
Determination of photosynthetic parameters
Fresh leaves were homogenized with 80% alkaline acetone (containing 1% ammonium hydroxide, V/V) to extract chlorophyll precursors, and the absorbance at 575, 590, and 628 nm was determined with Hodgins and VanHuystee’s method (1986).
Fresh leaves were immersed in a mixture of ethanol and acetone (1:1, V/V) in the darkness for Chl extraction. Chl contents were determined in line with the method of Arnon (1949), and determination of the absorbance at 645 nm and 663 nm was carried out.
Photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), intercellular CO2 concentration (Ci), and vapor pressure deficit (VPD) in leaves of the plants were recorded at 9:00–11:00 on a sunny day by using a portable photosynthetic system (CIRAS-2, PP System, UK).
A Handy PEA chlorophyll fluorimeter (Hansatech Instruments Limited, Norfolk, UK) was applied to assay Chl a fluorescence parameters of each treatment on the basis of Li et al. (2015) with minor modifications. Leaves were exposed to a saturated pulsed light of 3000 mmol·m−2·s−1 for 1 s after having been kept in darkness for 30 min. Parameters including minimal fluorescence (Fo), maximal fluorescence (Fm), and maximal efficiency of PSII (Fv/Fm) were obtained from the specific measurements assayed by the chlorophyll fluorimeter.
Statistical analysis
All data are mean values and standard deviations (SDs). The data were statistically analyzed using one-way analysis of variance in SPSS (version 19.0). P < 0.05 was employed to indicate statistical significance.
Results
Relative electron conductivity
As demonstrated in Appendix Fig. 6, the 50 and 75 mg L−1 treatments induced significantly increased relative electron conductivity by 43.58 and 87.03% (P < 0.05) when compared with the control, indicating that Cd2+ at higher concentrations can amplify leaf membrane permeability in P. cordata.
Variations in TBARS contents
TBARS, a by-product of membrane lipid peroxidation, represents the degree of plant cell membrane damage induced by adverse conditions. The TBARS content in the leaves of P. cordata increased with the Cd2+ concentration in the solution, and a significant increase was only observed at 75 mg L−1 Cd2+ compared with that of the control (Appendix Fig. 7), demonstrating that the highest Cd2+ concentration accelerated membrane lipid peroxidation.
Variations in antioxidative activities and phytochelatin contents
Compared with the control, Cd2+ at various concentrations significantly decreased Car contents by 11.37 ~ 50.41% (Table 1), and a similar variation tendency in total Car (Car T) content was also observed (P < 0.05). However, no obvious variations in the content of lutein, AsA, and AsA-DHA in the leaves were induced by Cd2+ (Table 1).
Compared with the control, no significant variations in SOD and CAT activities (Table 1) in the leaves of P. cordata were detected under 5 ~ 75 mg L−1 Cd2+ for 7 days(P > 0.05). Cd2+ increased the POD activity in the leaves, and a marked increase by 4.45 times was induced by 50 mg L−1 Cd2+, which was also demonstrated in the highest Cd2+ concentration (Table 1).
Dates are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05.
As indicated in Table 1, as the Cd2+ concentration increased, a marked increase in non-protein thiol contents in the leaves was observed. A prominent increase of 0.44% was induced by 75 mg L−1 Cd2+ in comparison with the control, which is similar to the response of phytochelatins (P < 0.05). Conversely, GSH content was relatively stable across all treatments (P > 0.05).
Variations in proline, soluble sugar, and protein contents
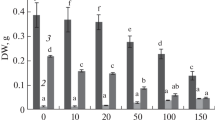
As shown in Fig. 1 a and b, Cd2+ no more than 50 mg L−1 did not alter soluble protein and soluble sugar contents (P > 0.05), which were dramatically decreased by 0.27 and 42.76% induced by 75 mg L−1 Cd2+ when compared with those in control, respectively (P < 0.05). Proline content increased with Cd2+ addition (Fig. 1c), and a significant increase of 1.27 ~ 2.84% in its content was observed under 50 and 75 mg L−1 Cd2+ (P < 0.05).

Variations in soluble protein, soluble sugar, and free proline contents of P. cordata leaves with various Cd2+ concentrations in the solution exposed for 7 days. Dates are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05
Variations in photosynthetic parameters
The content of protoporphyrin IX decreased notably by 20.22 ~ 33.96% compared with the control under 25 ~ 75 mg L−1 Cd2+ for 7 days (Table 2). With Cd2+ addition, Mg protoporphyrin IX content in the leaves decreased by 9.48 ~ 50.81%, which is similar to that observed with NADPH-protochlorophyllide (Pchlide) content (P < 0.05).
Cd2+ at various concentrations induced a significant reduction in Chl a content by 8.32 ~ 50.58% (P < 0.05), which was in accordance with Chl b and Chl T when compared with that in the control (Table 2). However, obvious variations in Chl a/b were not observed, as shown in Table 2.
Table 2 shows that a distinct reduction in Pn by 22.82 ~ 86.20%, Tr by 50.00 ~ 73.40%, Gs by 73.51 ~ 93.35%, and Ci by 29.03 ~ 36.74% in the leaves was differentially induced by Cd2+ at 5 ~ 75 mg L−1 compared with that in the control (P < 0.05), while VPD and Ls increased by 76.94 ~ 149.93% and 237.47 ~ 296.07%, which was dependent on the Cd2+ concentration in the solution. Simultaneously, we also noted that Cd2+ at 5 mg L−1 resulted in a prominent increase of 36.39%, and at the highest level led to an obvious reduction of 50.46% in water use efficiency (WUE) (P < 0.05). Compared with the control, Cd2+ at higher concentrations (≧ 25 mg L−1) significantly reduced CO2 use efficiency (CUE) by 37.33 ~ 81.45% (P < 0.05).
Dates are mean ± SD of three independent experiments. Different small and capital letters in the same row indicate significant differences at P < 0.05.
As indicated in Fig. 2a, Fo increased with Cd2+ concentration in the solution, and a significant increase was observed under 50 and 75 mg L−1 Cd2+ (P < 0.05), which was parallel to that observed in Vj, Vi dV/dTo, and dVG/dTo (Fig. 2d–g). The highest Cd2+ concentration induced a significant reduction in Fm by 21.41% (Fig. 2b), as well as in Fv (Fig. 2c), and a similar trend was also demonstrated by Fv/Fm (Fig. 3a). Under increased Cd2+ concentration, Fv/Fo and Fm/Fo declined (Fig. 3b, c), demonstrating a marked reduction by 17.29 ~ 106.03% and 14.51 ~43.19% under 50 ~ 75 mg L−1 Cd2+, respectively (P < 0.05). Furthermore, we also demonstrated Cd2+ at higher concentrations (≧ 25 mg L−1) induced a significant reduction in PIabs by 35.60 ~ 79.59% (Fig. 2d) and PItotal by 30.11 ~ 77.85% (Fig. 2e) compared with that in the control.

Variations in fluorescence yield of P. cordata leaves with Cd2+ exposure for 7 days. Dates are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05

Variations in PSII activity of P. cordata leaves with Cd2+ exposure for 7 d. Dates are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05
ABS/RC increased with Cd2+ concentration in the solution, and a conspicuous increase of 73.90% was induced by the highest Cd2+ concentration (Fig. 4a). A similar change was observed in DIo/RC (Fig. 4b) and TRo/RC (Fig. 4c) in comparison with the control. ETo/RC demonstrated a consistent increase by 9.71 ~ 14.60% under 5 ~ 50 mg L−1 Cd2+ (P < 0.05) and then decreased to control levels at the highest Cd2+ concentration (Fig. 4d). A significant increase in REo/RC was found only when the plant was exposed to 5 mg L−1(Fig. 4e). Under 5 mg L−1 Cd2+ addition, ETo/CSo (Fig. 5d) and REo/CSo (Fig. 5e) reached maximum levels (273.75; 97.89), respectively. Concurrently, ABS/CSo (Fig. 5a), DIo/CSo (Fig. 5b), and TRo/CSo (Fig. 5c) increased with Cd2+ concentration in the solution.

Variations in light absorption, transmission, and allocation in PSII per RC of P. cordata leaves with Cd2+ exposure for 7 days. Dates are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05

Variations in light absorption, transmission, and allocation in PSII per CS of P. cordata leaves with Cd2+ exposure for 7 days. Dates are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05
Discussion
Even at a low concentration, Cd2+ can affect physiological metabolic processes in plants (Seregin and Ivanov 2001), as has been demonstrated in Azolla pinnata (Prasad and Singh 2011), Lactuca sativa (Dias et al. 2013), and Ceratopteris pteridoides (Deng et al. 2014). This is also evident in our study, and a significant decline in Chl content was induced by 5 mg L−1 Cd2+ (Table 2), clearly indicating that Cd2+ disturbed chloroplast proliferation (Chen et al. 2008). Somashekaraiah et al. (1992) demonstrated that the membrane lipid peroxidation induced by Cd2+ decreased the Chl content in the leaves of Phaseolus vulgaris seedlings. Nevertheless, a significant increase in TBARS content under 5 mg L−1 Cd2+ (Appendix Fig. 7) was not found in the present study, suggesting that Cd2+ inhibits chlorophyll biosynthesis probably by suppressing magnesium ion chelation with protoporphyrin IX, as demonstrated by the reduced Mg protoporphyrin IX and Pchlide content (Table 2). Consequently, we speculated that Cd2+ probably damages Chl rather than the plasma membrane, as reported in other studies (Sytar et al. 2013; Nanda and Agrawal 2016). However, Cd2+ at higher concentrations (≧ 50 mg L−1) not only decreased Chl content (Table 2) but also increased membrane permeability (Appendix Fig. 6) and accelerated peroxidation (Appendix Fig. 7) in the leaves of P. cordata. We concurrently observed premature senescence symptoms, including chlorosis and withering of the leaves, indicating unbalanced activated oxygen metabolism and excessive ROS in the cells of the leaves. Similar results were found in cotton (Farooq et al. 2016) and Solanum melongena (Singh and Prasad 2014). Chloroplasts, like mitochondria, are considered to be the main organelles for ROS formation. Kato and Shimizu (1985) argued that metal ions can inhibit electron transfer in photosystem (PS) II, inducing the formation of excited chlorophyll, thereby resulting in ROS formation and lipid peroxidation, which probably decreases the content and instauration of fatty acids in the chloroplast membrane (Vassilev et al. 2004; Djebali et al. 2005; Sytar et al. 2013).
Enzymatic reaction systems constitute an important defense mechanism of a plant against heavy metal stress. As reported, Cd2+ can increase POD activity in leaves of a plant, such as cotton (Farooq et al. 2016) and Solanum melongena (Singh and Prasad 2014), as demonstrated in the present study (Table. 1). A slight increase in CAT (Table 1) and ascorbate peroxidase activities in leaves of P. cordata under heavy metal stress was also demonstrated in a previous study (Ma et al. 2020), which can alleviate lipid peroxidation (Mahmud et al. 2018). CAT displays a low affinity for H2O2 in plant cells and cannot effectively remove H2O2 by itself, and therefore AsA-GSH components (such as APX) are required for the elimination of H2O2 (Iqbal et al. 2010; Nanda and Agrawal 2016). Non-enzymatic reaction systems, consisting of AsA, GSH, and Car, also play a vital role in plant defense to heavy metals. GSH is a substrate for phytochelatin biosynthesis, and both are rich in thiol, which has a high affinity for Cd2+. Furthermore, GSH can also prevent damage to the cellular components by ROS. As demonstrated by Grupta et al. (2010), GSH contents in the shoots of Sedum alfredii cultivated with Pb2+ significantly increased by 51.61 ~ 403.23% compared with the control, demonstrating that GSH has an essential function in Pb2+ detoxification. An opposite trend was found in rice leaves under exposure to Cd2+ (Hsu and Kao 2007). In the present study, the content of NPT and PCs increased with Cd2+ concentration in the solution, whereas the GSH content did not change, as also demonstrated in Indian mustard in a study by Bashirl et al. (2015). This suggests that PCs could have a more important role in the detoxification of P. cordata leaves to Cd2+ than to GSH, as PCs can chelate Cd2+ to a stable complex in the vacuole, thus disturbing the combination of Cd2+ with macromolecular substances. Insufficient redox activity in the leaf cells of P. cordata induced by Cd2+ could be responsible for the lack of significant increase in GSH content. Furthermore, GSH is involved in AsA biosynthesis, and AsA participates in the response of plants to heavy metal stress by eliminating ROS in the cells. A significant reduction in AsA content induced by Cd2+ has been demonstrated in many plants, such as Hordeum vulgare (Wu et al. 2004) and Indian mustard (Bashirl et al. 2015), and this could result from the excessive AsA consumed in order to remove ROS. In our present investigation, no obvious alteration in AsA content was observed (Table 1), perhaps indicating that moderate amounts of AsA were responsible for ROS elimination, which is conducive for the maintenance of relatively stable oxidation resistance in cells. Furthermore, Cars will consume excessive excitation energy in PS II and remove ROS, thus protecting the photosynthetic membrane and preventing Chl molecule destruction, which would improve the adaptive capacity of plants to adverse conditions. We demonstrated significant reductions in the content of Car by 11.37 ~ 50.41% and Car T by 11.13 ~ 40.35% in the leaves of P. cordata under 5 ~ 75 mg L−1 Cd2+ (Table 1). Similar results were demonstrated in Arabidopsis thaliana (Peñalver et al. 2012) and Artemisia annua (Li et al. 2012), clearly demonstrating that Cd2+ impairs oxidation resistance in plant cells.
Osmotic adjustment substances, including proline, soluble sugars, and proteins, function in maintaining the osmotic potential of a plant cell. We found that Cd2+ at the highest concentration induced a marked reduction in soluble protein and soluble sugar contents (Fig. 1a, b) by inhibiting their accumulation and accelerating their decomposition, which would result in declined capacity of the cells in the P. cordata leaves to hold water. Soluble sugars are unstructured carbohydrates and the products of photosynthesis, and thus suppressed photosynthesis will lead to decreased soluble sugar content, which would not be sufficient for the cellular defense and repair mechanisms for sugar. Soluble protein can combine with available metal ions in plant cells, alleviating their cytotoxicity (Wei et al. 2016). However, higher Cd2+ concentrations will increase proteolytic enzyme activity and aggravate membrane lipid peroxidation (Appendix Fig. 7), leading to accelerated proteolysis (Chiraz et al. 2003). Proline is a stabilizer of cell membrane and some large molecules, as well as a scavenger of ROS, which can protect the integrity of cell structure and function. As confirmed in the present study, the proline content in the cells of P. cordata leaves increased with Cd2+ concentrations, as has been observed in tomato (Khateeb and Al-Qwasemeh 2014), indicating that proline plays a significant role in protecting plant cells from heavy metal toxicity.
Photosynthesis is considered to be a crucial process of plant growth and biomass formation and is sensitive to heavy metals. Reduced photosynthetic activity under Cd2+ is common (Farooq et al. 2016; Liu et al. 2017). This could result from inhibited Chl biosynthesis (Chen et al. 2008), reduced photosynthetic efficiency (Chu et al. 2018), photosynthetic enzyme activity (Deng et al. 2014), and disturbed water and nutrition balance in plants (Dias et al. 2013; Zhang et al. 2014). In our present study, Cd2+ at various concentrations induced a reduction in Pn of P. cordata compared with the control (Table 2). Deng et al. (2014) demonstrated a significant decline in Pn in Ceratopteris pteridoides under exposure to 1.12 mg L−1 Cd2+ for 7 days, as was also found in Zea mays under 2.5 mg L−1 Cd2+ (Wang et al. 2009). However, Pn in rice leaves was not altered under exposure to 11.24 mg L−1 Cd2+ for 7 days (Xu et al. 2005), indicating that P. cordata has a relatively stronger photosynthetic tolerance to Cd2+. With increased Cd2+ concentrations, Gs and Ci declined, while Ls increased, demonstrating that stomatal limitation was probably responsible for the reduced photosynthesis in P. cordata (Farquar and Sharkey 1982). Stomatal sensitivity to saturated vapor pressure deficit (VPD) is enhanced to different degrees in plants cultivated with Cd2+ at various concentrations, thereby displaying varied degrees of stomatal closure in the leaves. Leaf water use efficiency (WUE) represents the basic efficiency of a plant to consume water and form dry matter, reflecting the adaptive capacity of a plant to adversities. In the present study, 5 mg L−1 Cd2+ led to a significant increase in WUE compared with that in the control, demonstrating that transpiration was more strongly inhibited by Cd2+ than photosynthesis, which could effectively avoid excessive water loss and thus improve the adaptive capacity of P. cordata to 5 mg L−1 Cd2+. However, we showed a significant reduction in WUE by 50.46% under the highest Cd2+ concentration, and Deng et al. (2014) demonstrated a similar result in Ceratopteris pteridoides, revealing damaged physiological metabolism in the leaves and a decreased ability of the cells to hold water. This is demonstrated by the reduced soluble protein and soluble sugar contents in the present study (Fig. 1a, b), which would be unfavorable to photosynthesis in P. cordata (Table 2). Furthermore, CUE in the leaves of P. cordata declined with increased Cd2+ concentration in the solution, which negatively impacted plant growth, as demonstrated by the reduced biomass in the late period of the test. Additionally, Cd2+ could also induce a marked reduction in RuBisCO activity in the dark reactions, thus disturbing normal photosynthesis (Dias et al. 2013).
Variations in Chl a fluorescence parameters indicate changes in the photosynthetic apparatus efficiency of P. cordata leaves under Cd2+ stress (Fig. 2, 3, 4, and 5). Fo represents minimal fluorescence, and the increase in its value is related to PS II reaction center partial closure or inactivation. In the present study, significant increases in Fo were induced by 50 and 75 mg L−1 Cd2+ (Fig. 2a), as demonstrated by the increased dV/dTo, dVG/dTo (Fig. 2f, g). Similar results were also reported in Z. mays (Tanyolaç et al. 2007) and R. pseudoacacia (Dezhban et al. 2015), which indicated inhibited acceptor sides of PS II, accordingly increasing excitation energy loss during the transfer of excitation energy from PS II antenna pigments to the reaction center. Variations in Fv/Fm can determine variations in PS II photochemical activity, and the highest Cd2+ concentration led to a significant increase in Fv/Fm (Fig. 3a) resulting from the decline in Fm (Fig. 2b), which probably indicated damaged structure of the chloroplast thylakoid membrane, thereby inhibiting electron transfer (Monnet et al. 2001). This was demonstrated by the reduced ETo/RC and ETo/CSo (Figs. 4d and 5d). Baszynski et al. (1980) and Siedleska and Baszynski 1993 argued that thylakoid degradation induced by Cd2+ will intensively inhibit electron transfer in PS II. Furthermore, a reduction in chlorophyll content, as demonstrated in our present study, could be due to inhibition of electron transfer by suppressing the capture efficiency of excitation energy (Vassilev and Yordanov 1997). Declined Fv/Fo and Fm/Fo as evidenced by our study (Fig. 3b, c) clearly indicates that Cd2+ induces PS II inactivation and thus reduces photosynthetic efficiency as reported earlier (Xing et al. 2010). Moreover, impaired PS II increased specific activity psarameters, including ABS/RC, DIo/RC, TRo/RC, ETo/RC, ABS/CSo, DIo/CSo, and TRo/CSo, were observed in Elsholtzia argyi (Li et al. 2015) and Solanum melongena (Singh and Prasad 2015) with Cd2+ exposure. A significant increase in REo/RC was only observed under 5 mg L−1 Cd2+ (Fig. 4d). Compared with Fv/Fm, performance indexes, such as PIabs and PItotal found to be more sensitive to Cd2+ that more accurately indicate the activity of the PS II reaction center in plants. In the present study, Cd2+ at higher concentrations (≧ 25 mg L−1) markedly declined PIabs and PItotal (Fig. 3d, e), which was consistent with the findings in Solanum melongena (Singh and Prasad 2015), indicating reduced activity of the PS II reaction center.
Conclusion
Under exposure to 5 mg L−1 Cd2+ for 7 days, P. cordata displayed good morphophysiological adaptability and good growth with no symptoms of toxicity. We demonstrated that 5 mg L−1 Cd2+ firstly damaged the Chl rather than the plasma membrane in the leaves. Additionally, the declined Chl content, stomatal conductance, and the capacity of the cells to hold water, as well as the increased oxidative stress in the P. cordata leaves, were probably responsible for the photoinhibition induced by Cd2+ at higher concentrations. POD, proline, and PCs alleviated the Cd2+ toxicity in the leaf cells of P. cordata. The reduction in photosynthetic efficiency could mainly have resulted from partial closure and inactivation, restrained acceptor sides of the PS II reaction center, and inhibited electron transfer.
Abbreviations
- PSII:
-
Photosystem II
- F o :
-
Minimal fluorescence in dark-adapted leaves
- F m :
-
Maximal fluorescence in dark-adapted leaves
- Fv/fm :
-
Maximum quantum efficiency of PSII
- Fv/Fo :
-
The potential activity of PSII
- Fm/Fo :
-
Electron transfer in PSII
- dV/dTo :
-
Net close rate of reaction center at 300 μs
- dVG/dTo :
-
Net close rate of reaction center at 100 μs
- V j :
-
Relatively variable fluorescence at J point
- V i :
-
Relatively variable fluorescence at I point
- ABS/RC:
-
Absorption flux (of antenna Chlorophylls) per reaction center
- TRo/RC:
-
Trapped energy flux (leading to QA reduction) per reaction center
- DIo/RC:
-
Dissipate flux per reaction center
- ETo/RC:
-
Trapped energy flux for electron transfer per reaction center
- RE/RC:
-
Electron flux reducing end electron acceptors at the PSI acceptor side per reaction center
- ABS/CSo :
-
Absorption flux in cross section
- TRo/CSo :
-
Trapped energy flux in cross section
- ETo/CSo :
-
Electron transport flux in cross section
- DIo/CSo :
-
Dissipate flux in cross section
- REo/CSo :
-
Electron flux reducing end electron acceptors at the PSI acceptor side in cross section
- RC/CSo :
-
Density of active reaction centers (QA-reducing PSII reaction centers)
- PIabs :
-
Performance index for energy conservation from photons absorbed by PSII to the reduction of intersystem electron acceptors
- PItotal :
-
Performance index for energy conservation from photons absorbed by PSII to the reduction of PSI end acceptors
References
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Anderson ME (1985) Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol 113:548–555
Arnon DI (1949) Copper enzyme in isolated chloroplast. Polyphenoloxidases in Beta vulgaris. Plant Physiol 24:1–16
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Bashirl H, Ibrahim MM, Bagheril R, Ahmadl J, Arif IA, Baigl MA, Qureshi MI (2015) Influence of sulfur and cadmium on antioxidants, phytochelatins and growth in Indian mustard. AoB Plant 7:plv001
Baszynski T, Wajda L, Krol M, Wolinska D, Krupa Z, Tukendorf A (1980) Photosynthetic activities of cadmium-treated tomato plants. Physiol Plant 48:365–370
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207
Beyer WF, Fridovich I (1987) Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal Biochem 161:546–553
Bhargava P, Srivastava AK, Urmil S, Rai LC (2005) Phytochelatin plays a role in UV-B tolerance in N2-fixing cyanobacterium Anabaena doliolum. J Plant Physiol 162:1220–1225
Chen F, Wang F, Zhang GP, Wu FB (2008) Identification of barley varieties tolerant to cadmium toxicity. Biol Trace Elem Res 121:171–179
Chiraz C, Houda G, Habib GM (2003) Nitrogen metabolism in tomato plants under cadmium stress. J Plant Nutr 26:1617–1634
Chu J, Zhu F, Chen X, Liang H, Wang R, Wang X, Huang X (2018) Effects of cadmium on photosynthesis of Schima superba young plant detected by chlorophyll fluorescence. Environ Sci Pollut Res 25:10679–10687
Deng G, Li M, Li H, Yin L, Li W (2014) Exposure to cadmium causes declines in growth and photosynthesis in the endangered aquatic fern (Ceratopteris pteridoides). Aquat Bot 112:23–32
Dezhban A, Dhirvany A, Attarod M, Delshad M, Matinizadeh M, Khoshnevis M (2015) Cadmium and lead effects on chlorophyll fluorescence, chlorophyll pigments and proline of Robinia pseudoacacia. J Forestry Res 26:323–329
Dias MC, Monteiro C, Moutinho-Pereira J, Correia C, Gonçalves B, Santos C (2013) Cadmium toxicity affects photosynthesis and plant growth at different levels. Acta Physiol Plant 35:1281–1289
Djebali W, Zarrouk M, Brouquisse R, EI Kahoui S, Limam F, Ghorbel MH, Chaïbi W (2005) Ultrastructure and lipid alterations induced by cadmium in tomato (Lycopersicon esculentum) chloroplast membranes. Plant Biol 7:358–368
Dou M, Zhao P, Wang Y, Li G (2017) Health risk assessment of cadmium pollution emergency for urban populations in Foshan City, China. Environ Sci Pollut Res 24:8071–8086
Farhangi-Abriza S, Torabianb S (2017) Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotox Environ Safe 137:64–70
Farooq MA, Ali S, Haneed A, Bharwana SA, Rizwan M, Ishaque W, Farid M, Mahmood K, Iqbal Z (2016) Cadmium stress in cotton seedlings: physiological, photosynthesis and oxidative damages alleviated by glycinenetaine. S Afr J Bot 104:61–68
Farquar GD, Sharkey TD (1982) Stomatal conductance and photosynthesis. Annu Rev Plant Physiol 33:317–345
Ghoulam C, Foursy A, Fares K (2002) Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ Exp Bot 47:39–50
Grant CA, Clarke JM, Duguid S, Chaney RL (2008) Selection and breeding of plant cultivars to minimize cadmium accumulation. Sci Total Environ 390:301–310
Groppa MD, Ianuzzo MP, Rosales EP, Vázquez SC, Benavides MP (2012) Cadmium modulates NADPH oxidase activity and expression in sunflower leaves. Biol Plant 56:167–171
Grupta DK, Huang HG, Yang XE, Razafindrabe BHN, Inouhe M (2010) The detoxification of lead in Sedum alfredii H. is not related to phytochelatins but the glutathione. J Hazard Mater 177:437–444
Hodgins RR, Huystee RBV (1986) Rapid simultaneous estimation of protoporphyrin and Mg-porphyrins in higher plants. J Plant Physiol 125:311–323
Hsu YT, Kao CH (2007) Cadmium-induced oxidative damage in rice leaves is reduced by polyamines. Plant Soil 291:27–37
Iqbal N, Masood A, Nazar R, Syeed S, Khan NA (2010) Photosynthesis, growth and antioxidant metabolism in mustard (Brassica juncea L.) cultivars differing in cadmium tolerance. Agric Sci China 9:519–527
Kampfenkel K, Vanmontagu M, Inze D (1995) Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal Biochem 225:165–167
Kato M, Shimizu S (1985) Chlorophyll metabolism in higher plants VI. Involvement of peroxidase in chlorophyll degradation. Plant Cell Physiol 26:1291–1301
Khateeb W, Qwasemeh HA (2014) Cadmium, copper and zinc toxicity effects on growth, proline content and genetic stability of Solanum nigrum L., a crop wild relative for tomato; comparative study. Physiol Mol Biol Plants 20:31–39
Krämer U, Clemens S (2005) Functions and homeostasis of zinc, copper, and nickel in plants. In: Tamas MJ, Martnoia E. Molecular Biology of Metal Homeostasis and Detoxification. Springer Berlin Heidelberg
Li HS (2000) Principles and techniques of plant physiological biochemical experiment. Higher Education Press, Beijing
Li S, Yang W, Yang T, Chen Y, Nia W (2015) Effects of cadmium stress on leaf chlorophyll fluorescence and photosynthesis of Elsholtzia argyi-a cadmium accumulating plant. Int J Phytoremediat 17:85–92
Li X, Zhao MX, Guo LP, Huang LQ (2012) Effect of cadmium on photosynthetic pigments, lipid peroxidation, antioxidants, and artemisinin in hydroponically grown Artemisia annua. J Environ Sci 24:1511–1518
Lim TY, Lee ST, Cho HG, Kim SO (2016) Comparison of human health risk assessment of heavy metal contamination from two abandoned metal mines using metal mine-specific exposure parameters. J Environ Impact Assess 25:414–431
Liu H, Zhang C, Wang J, Zhou C, Feng H, Mahajan MD, Han X (2017) Influence and interaction of iron and cadmium on photosynthesis and antioxidative enzymes in two rice cultivars. Chemosphere 171:240–247
Ma S, Xin J, Chen Y, Xin H, Tian R (2020) Effects of copper on antioxidant enzyme activities, antioxidant and non-protein thiol content in Pontederia cordata’s leaves. Pratacul Sci 37:459–468
Mahmud JA, Hasanuzzaman M, Nahar K, Borhannuddinbhuyan MHM, Fujita M (2018) Insights into citric acid-induced cadmium tolerance and phytoremediation in Brassica juncea L.: coordinated functions of metal chelation, antioxidant defense and glyoxalase systems. Ecotoxicol Environ Saf 147:990–1001
Manging WJ, Flager RB, Frenkel MA (2003) Assessing plant response to ambient ozone: growth ozone-sensitive loblolly pine seeding treated with ethylenediurea or sodium erythorbate. Environ Pollut 126:73–81
McLaughlin MJ, Parker DR, Clarke JM (1999) Metals and micronutrients: food safety issues. Field Crop Res 60:143–163
Monnet F, Vailant N, Vernay P, Coudret A, Sallanon H, Hitmi A (2001) Relationship between PSII activity, CO2 fixation, and Zn, Mn, and Mg contents of Lolium perenne under zinc stress. J Plant Physiol 158:1137–1144
Munns R (1988) Why measure osmotic adjustment? Aust J Plant Physiol 15:717–726
Muradoglu F, Gundogdu M, Ercisli S, Tarik E, Balta F, Jaafar HZE, Zia-Ul-Haq M (2015) Cadmium toxicity affects chlorophyll a and b content, antioxidant enzyme activities and mineral nutrient accumulation in strawberry. Biol Res 48:11
Nanda R, Agrawal V (2016) Elucidation of zinc and copper induced oxidative stress, DNA damage and activation of defense system during seed germination in Cassia angustifolia Vahl. Environ Exp Bot 125:31–41
Peñalver AM, Graña E, Reigosa MJ, Moreiras AMS (2012) The early response of Arabidopsis thaliana to cadmium- and copper-induced stress. Environ Exp Bot 78:1–9
Prasad SM, Singh A (2011) Metabolic responses of Azolla pinnata to cadmium stress: photosynthesis, antioxidative system and phytoremediation. Chem Ecol 276:543–555
Rama DS, Prasad MNV (1998) Copper toxicity in Ceratophyllum demersum L. (Coontail), a free floating macrophyte: response of antioxidant enzymes and antioxidants. Plant Sci 138:157–165
Reimer P, Duthie HC (1993) Concentrations of zinc and chromium in aquatic macrophytes from the Sudbury and muskoka regions of Ontario, Canada. Environ Pollut 79:261–265
Seregin IV, Ivanov VB (2001) Physiological aspects of cadmium and lead toxic effects on higher plants. Russ J Plant Physiol 48:523–544
Siedleska A, Baszynski T (1993) Inhibition of electron transport flow around photosystem I in chloroplasts of Cd-treated maize plants is due to Cd-induced iron deficiency. Physiol Plant 87:199–202
Simmons RW, Pongsakul P, Chaney RL, Saiyasitpanich D, Klinphoklap S, Nobuntou W (2003) The relative exclusion of zinc and iron from rice grain in relation to rice grain cadmium as compared to soybean: implications for human health. Plant Soil 257:163–170
Singh S, Prasad SM (2014) Growth, photosynthesis and oxidative responses of Solanum melongena L. seedlings to cadmium stress: mechanism of toxicity amelioration by kinetin. Sci Hortic 176:1–10
Singh S, Prasad SM (2015) IAA alleviates cd toxicity on growth, photosynthesis and oxidative damages in eggplant seedlings. Plant Growth Regul 77:87–98
Somashekaraiah BV, Padmaja K, Prasad ARK (1992) Phytotoxicity of cadmium ions on germinating seedlings of mung bean (Phaseolus vulgaris): involvement of lipid peroxides in chlorophyll degradation. Physiol Plant 85:85–89
Srivastava RK, Rajpoot R, Pandey P, Rani A, Dubey RS (2017) Cadmium alters mitochondrial, membrane potential, inhibits electron transport chain activity and induces callose deposition in rice seedlings. J Plant Growth Regul 37:1–10
Sytar O, Kumar A, Sato T, Latowski D, Kuczynska P, Strzalka K, Prasad M (2013) Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol Plant 35:985–999
Tanyolaç D, Ekmekçi Y, Ünalan Ş (2007) Changes in photochemical and antioxidant enzyme activities in maize (Zea mays L.) leaves exposed to excess copper. Chemosphere 67:89–98
Upadhyaya A, Sankhla D, Davis TD, Sankhla N, Smith BN (1985) Effect of paclobutrazol on the activities of some enzymes of activated oxygen metabolism and lipid peroxidation in senescing soybean leaves. J Plant Physiol 121:453–461
Vassilev A, Lidon F, Scotti P, Da Graca M, Yordanov I (2004) Cadmium-induced changes in chloroplast lipids and photosystem activities in barley plants. Biol Plant 48:153–156
Vassilev A, Yordanov I (1997) Reductive analysis of factors limiting growth of cadmium-treated plants: a review. Bul J Plant Physiol 23:114–133
Wang H, Zhao SC, Liu RL, Zhou W, Jin JY (2009) Changes of photosynthetic activities of maize (Zea mays L.) seedlings in response to cadmium stress. Photosynthetica 47:277–283
Wu FB, Chen F, Wei K, Zhang GP (2004) Effect of cadmium on free amino acid, glutathione and ascorbic acid concentrations in two barley genotypes (Hordeum vulgare L.) differing in cadmium tolerance. Chemosphere 57:447–454
Xing W, Huang W, Liu G (2010) Effect of excess iron and copper on physiology of aquatic plant Spirodela polyrrhiza (L.) Schleid. Environ Toxicol 25:103–112
Xu YM, Wei YH, Zeng JY (2013) Comparative analysis on the content of carotenoid in the leaves of the three general plants. J Guangxi Normal Uni Nat 30:18–19
Xu H, Weng X, Mao W, Yang Y (2005) Effects of cadmium stress on photosynthesis, chlorophyll fluorescence characteristics and excitation energy distribution in leaves of rice. Chinese J Rice Sci 19: 338–342
Zhang X, Gao B, Xia H (2014) Effect of cadmium on growth, photosynthesis, mineral nutrition and metal accumulation of bana grass and vetiver grass. Ecotoxicol Environ Saf 106:102–108
Zhou W, Qiu B (2005) Effects of cadmium hyperaccumulation on physiological characteristics of Sedum alfredii Hance (Crassulaceae). Plant Sci 169 (4):737–745
Acknowledgments
The research was supported by the National Natural Science Foundation of China (No. 30972408), China Postdoctoral Science Foundation (2020M671509), A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Qing Lan Project. We also thank the Advanced Analysis and Testing Center (AATC) of Nanjing Forestry University for their kind support and LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Gangrong Shi
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendix 1

Variations in membrane permeability of P. cordata leaves with various Cd2+ concentrations in the solution exposed for 7 days. Dates are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05
Appendix 2

Variations in TBARS content of P. cordata leaves with various Cd2+ concentrations in the solution exposed for 7 days. Date are mean ± SD of three independent experiments. Different small letters indicate significant differences at P < 0.05
Rights and permissions
About this article

Cite this article
Xin, Jp., Ma, S., Zhao, C. et al. Cadmium phytotoxicity, related physiological changes in Pontederia cordata: antioxidative, osmoregulatory substances, phytochelatins, photosynthesis, and chlorophyll fluorescence. Environ Sci Pollut Res 27, 41596–41608 (2020). https://doi.org/10.1007/s11356-020-10002-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-020-10002-z