
Abstract
Vachellia campechiana (Mill Seigler & Ebinger) is widely distributed in Mexico and is a dominant species of tailings in Huautla, in the state of Morelos, Mexico. Mining activities carried out in this region generated about 780 thousand tons of bioavailable heavy metal waste (HMs) that were deposited in the environment without any treatment. This study evaluates the bioaccumulation capacity and morphological changes of V. campechiana growing during 1 year in control or tailing substrates (treatments) under greenhouse conditions. The concentration of six HMs was also measured in roots, leaves, and seeds by atomic absorption spectrophotometry. Five metals showed a similar bioaccumulation pattern in the roots and leaves of V. campechiana grown in both substrates: Pb > Fe > Cr > Cu > Zn. The concentrations of Cr, Cu, and Pb were significantly higher in the roots and leaves of individuals growing on the exposed substrate. The presence of essential metals (Cu, Fe, Zn) was only recorded in the seeds, with similar concentrations in both treatments. Seventeen of 18 morphological characters evaluated in V. campechiana decreased in plants exposed to metals. Pb, Cu, and Fe showed a bioconcentration factor greater than one in roots and leaves. The translocation factor showed the following pattern: Cr > Cu = Pb. In conclusion, V. campechiana is a candidate species to phytoremediate environments contaminated with Pb, Cr, and Cu due to its ability to establish itself and turn into the dominant plant species in polluted sites, its ability to bioaccumulate non-essential metals in roots and leaves, and its high rate of HMs translocation.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mining generates waste in the form of gases, sewage, and/or tailings (Gutiérrez-Ruiz et al. 2007). As a result of the improper management of the waste derived from this activity, tailings have created environmental and health problems in Mexico (Mireles et al. 2012; Cortés-Jiménez et al. 2012; Mussali-Galante et al. 2013) due to their content of potentially toxic elements such as heavy metals (HMs), which harm living organisms by bioaccumulating in their tissues and damaging their genes and their ecosystems (Gutiérrez and Moreno 1995; Mussali-Galante et al. 2013).
The existence of plant species that tolerate high concentrations of metals in the soil has been reported before. These plants restrict the absorption of heavy metals and/or translocate them to the leaves, which allows them to maintain constant and relatively low concentrations of these pollutants in the aerial biomass, regardless of the concentration of metals in the soil (exclusion strategy). Other plant species actively absorb metals from the soil and accumulate them in non-toxic forms in the aerial biomass (accumulation strategy) (Brooks et al. 1977; Furini 2012; Marrero-Coto et al. 2012).
This differential response in the absorption of HMs present in the environment depends on the bioavailability of the metals, the retention capacity of the metals, the interaction between plants, roots, and metals, the plant metabolism, and the physicochemical properties of the soil, such as pH, organic matter content, and electrical conductivity (Kabata-Pendias 2000; Barceló and Poschenrieder 2003; Prieto et al. 2009; Rascio and Navari-Izzo 2011). It has been reported that the bioaccumulation of HMs by plants varies considerably across different taxonomic groups (Calow 1993).
Exposure to HMs can have various effects on plants, such as inhibiting seed germination, inhibiting root growth, causing alterations in the root biomass, hindering seedling development, causing micro-morphological alterations (stomata and trichomes), reducing the plant biomass (roots, stem, and leaves), and altering biochemical processes such as protein inhibition (Yadav 2010; Rengel et al. 2011; Tovar-Sánchez et al. 2019). Despite these adverse effects, some plant species have the ability to deal with the presence of HMs in the environment where they grow (Rascio and Navari-Izzo 2011). These species are known as accumulators and have the ability to establish themselves naturally in environments contaminated with HMs. They are fast growing species, with abundant aerial biomass, high capacity to absorb HMs from the soil, efficient translocation of HMs from root to leaf tissue, and high capacity to detoxify and retain large amounts of HMs in their leaf tissue (Rascio and Navari-Izzo 2011; Cappa and Pilon-Smits 2014; Shiqi et al. 2018). To determine if a species has these characteristics, it is necessary to evaluate how it responds to the effects of HMs when exposed to them.
The use of heavy metal accumulator plants has been a phytoremediation strategy in contaminated sites. These plants absorb the elements and remove them from mining waste, preventing their leaching to the water table. Phytoremediation uses plant species to immobilize and store HMs in different plant structures such as roots, stems, and leaves (Shiqi et al. 2018). Some indexes have been proposed in the literature that measure the capacity of plants to store HMs. For example, Yoon et al. (2006) proposed the bioconcentration factor (BCF) as a parameter that measures the efficiency of a plant species to accumulate metals from the soil in leaf tissue. They also proposed the translocation factor (FT), which indicates the efficiency with which metals are transported from the roots to the aerial parts of the plant. Using the latter index, Abhilash et al. (2009) reported that Limnocharis flava (L.) Buchenau (Limnocharitaceae) is an accumulator of cadmium (Cd).
About 450 species of plants have been recognized as accumulators of HMs, mainly Cd, nickel (Ni), copper (Cu), lead (Pb), and zinc (Zn) (Macnair 2003; Van der Ent et al. 2013). It is still important to characterize new plant species with the potential to bioaccumulate HMs and to be used in the phytoremediation of contaminated environments (Mendez and Maier 2008).
In Mexico, Huautla, located in the state of Morelos, was a mining area until 1988. It is estimated that there are in the area approximately 780,000 tons of waste in the form of tailings. These are rich in Pb, manganese (Mn), Cd, arsenic (As), Zn, Cu, iron (Fe), and chromium (Cr), all of which are bioavailable (Velasco et al. 2005; Mussali-Galante et al. 2013). Despite this, some species of plants, such as V. campechiana (Fabaceae), a shrubby species with different uses that is widely distributed in Mexico, mainly in arid and semi-arid areas, have established themselves in the Huautla area (Arce 2001; Rico 2001; Cervantes and Sotelo 2002; Armienta et al. 2008). Since it inhabits areas contaminated with HMs, V. campechiana could have characteristics that allow it to be considered as a plant with potential for phytoremediation; it has a high seed germination percentage, is fast growing, and has abundant leaf biomass. However, its possible role in the bioremediation of environments contaminated with HMs is still unknown. Therefore, the present study evaluated, under greenhouse conditions, the accumulation of HMs in the leaf and root tissue of V. campechiana individuals growing on a tailing substrate and a control substrate and the effect of this accumulation on seed germination and on the macro- and micromorphological leaf characters of the plants. The questions that guided this study were as follows: 1) Do individuals of V. campechiana growing on tailing substrate bioaccumulate HMs in root and leaf tissue? 2) Is the exposure time to HMs a factor that favors the bioaccumulation in root and leaf tissue of V. campechiana? 3) Does exposure to HMs promote changes in the macro- and micromorphological characters of this species? 4) Does the growing substrate (control, exposed) have an effect on the bioaccumulation levels of HMs and on seed germination? 5) Does the morphological and HMs bioaccumulation response (root, leaf, seeds) of V. campechiana plants chronically exposed to mine tailings make them useful for the phytoremediation of contaminated environments?
Methods
Study sites
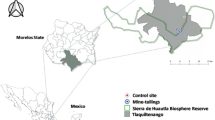
This study was conducted in the municipality of Tlaquiltenango, in the state of Morelos. Mining activities were carried out in this area until 1988, mainly extracting Pb, Zn, and Ag. For decades, the waste generated from mining was deposited in the open, without any type of treatment. Three main tailings were formed, both of which are located in the town of Huautla, south of the municipality of Tlaquiltenango, Morelos, within the Sierra de Huautla Biosphere Reserve (REBIOSH). Tailing mine 1 (T1) is the largest and is located 500 m from the town (8° 26′ 36.37″ N and 99° 01′ 26.71″ W). Tailing mine 2 (T2) is located 1000 m from the town (18° 26′ 22.62″ N and 99° 01′ 51.71″ W). Tailings are rich in metals such as Pb, Mn, Cd, As, Zn, Cu, Fe, and Cr (Velasco et al. 2005). The following control sites were chosen: Quilamula (C1 site) located at 18° 30′ 52″ N and 98° 59′ 59″ W, at an altitude of 1100 m, and Ajuchitlán (C2 site), located at 18° 27′ 52″ N and 98° 58′ 53″ W, at an altitude of 1050 m. Both sites have very similar ecological and geographical characteristics to the exposed sites (Martínez-Pacheco 2008), but have no records of mining activity or anthropogenic metal contamination (Mussali-Galante et al. 2013). Both sites are more than 6 km in linear distance from the exposed sites. In general, Tlaquiltenango presents a natural richness of mineral soils (mainly sulfur minerals) of silver and lead. The most commonly found minerals are as follows: arsenopyrite (FeAsS), galena (PbS), acantite (Ag2S), and calclacita (Cu2S) (Volke et al. 2004, 2005; Secretaria de Economía 2011). Therefore, the soils of the region are naturally rich in minerals.
Study species
V. campechiana is a shrub-like species (Fabaceae) commonly known as “cubata,” reaching up to 4 m in height, with concave spines, usually reddish when young and brown when mature, spoon-shaped, and up to 3.5 cm long and 1.3 cm wide (Arce 2001). Its fruit is white, flattened, indehiscent, and brown-reddish, with no obvious margins. Seeds of ellipsoidal shape were light brown or yellowish, without aryl (Arce 2001). The seeds show physical latency; however, when this latency is broken, the germination percentage reaches 90% (Baskin and Baskin 2004). It is a kind of secondary vegetation of tropical deciduous forests that inhabits places with sub-humid or dry climates, at altitudes ranging from 1150 to 1450 m. It is an evergreen species that blooms in May–August and produces fruit in September–May. It is used as fuel, as material for fence posts and construction, and as a growing tutor for other plants to make agricultural tools. It is also a forage and medicinal species, used for kidney and stomach problems (Rico 2001; Cervantes and Sotelo 2002).
Seed collection
Seeds of V. campechiana were collected from individuals established in the two control sites (C1 and C2) and in the two exposed sites (T1 and T2) (Fig. 1). So that the germination analysis encompassed the genetic variability of the species, the seeds from sites C1 and C2 were considered as control seeds, while the seeds from T1 and T2 were considered exposed seeds. At each site, 20 individuals were randomly selected and 20% of the seeds were collected from them (Gold et al. 2004). The seeds were transported to the laboratory, where they were cleaned and selected, removing the seeds parasitized by insects.

Geographical distribution of the two study sites at the Sierra de Huautla Biosphere Reserve, Morelos, Mexico. Control site (black-filled up-pointing triangle). Exposed site (black-filled circle)
Germination of V. campechiana and plant growing
To evaluate the germination percentage, V. campechiana seeds from the control and the exposed sites were subjected to mechanical scarification due to the presence of physical latency. Twenty-five seeds were sown in a Petri dish with agar (1%), using six replicates per treatment. This assay was monitored for 20 days.
After the seeds germinated, 72 seeds were transplanted into individual nursery polyethylene bags (15 L) with the treatment substrates (36 in tailing substrate and 36 in control substrate). Soil from Quilamula was used as control substrate; it was sieved with a stainless steel sieve number 35 (Fiicsa), with a 0.5 mm mesh, in order to obtain a particle size similar to that of the mine tailing. The mixture of residues from tailings 1 and 2 was considered the exposed substrate. All the plants were kept under greenhouse conditions; they were watered twice a day, three times a week, and the temperature ranged from 32 to 35 °C. The plants obtained were used to evaluate the bioaccumulation of HMs and measure the macro- and micromorphological characters of interest.
Evaluation of morphological characters
To assess the effect of the time of exposure to HMs on the morphological characters of V. campechiana, six individual plants were randomly selected per treatment (control and tailing substrates). Six leaves were randomly chosen from each individual, and the foliar macro- and micromorphological characters shown in Table 1 were measured every 2 months of exposition to treatments, over 1 year. The macromorphological characters were measured with a digital vernier (Stainless Hardened) and a digital scale (Acculab). For the micromorphological characteristics, a foliar epidermal impression was done using a replication technique with cyanoacrylate glue. Three slides with epidermal impressions of the abaxial part of the leaf were made per each individual plant. The slides were observed with an optical microscope (Leica) at 40X with bright field illumination (CC) and differential interference contrast (CDI). Three photomicrographs were taken at random from each slide. From the nine photomicrographs, an average of the number of stomata (E), epidermal cells (CE), and stomatal index (IE) was obtained for each individual plant. The stomatal index was calculated according to Salisbury (1968).
Concentration of heavy metals in the substrate and plant tissue of V. campechiana
A total of 10 samples of the exposed substrate were analyzed to determine the concentration of metals (Cd, Cr, Cu, Fe, Mn, Pb, and Zn). The samples were dried and sieved following the method established by the Mexican standard NMX-AA-132-SCFI-2006. This process consists in adding 50 mL of CaCl2 (0.01 M) to 10 g of substrate. The sample is kept under stirring for 24 h and centrifuged at 1500 rpm for 15 min, recovering the supernatant by filtration. The concentration of metals in the substrate samples was determined by atomic absorption spectrophotometry using the flame method (GBC 908 A).
To evaluate the concentration of metals (Cd, Cr, Cu, Fe, Mn, Pb, and Zn) in the root and leaf tissue of V. campechiana, three samples were taken from six individuals per substrate (tailing and control), every 2 months of exposure to treatments over 1 year. To evaluate the concentration of metals in seeds of V. campechiana, three samples were taken from six individuals per substrate (tailing and control). An amount of 0.25 g of each plant structure were pulverized in containers previously washed with HNO3.The samples were subjected to acid digestion in an Accelerated Reaction System Microwave (CEM® MARS-5) using 10 mL of HNO3 (70%) in Teflon pumps. The samples were dissolved and filtered in distilled water to a final volume of 50 mL until further analysis. A sample without tissue was processed simultaneously and was used as a control. The metals were analyzed by atomic absorption spectrophotometry using the flame method (GBC 908 A). The spectrophotometer was calibrated using standard solutions and known concentrations for each metal analyzed. The minimum detection limits (mg/L) of Cd, Cr, Cu, Fe, Mn, Pb, and Zn were 0.0004, 0.003, 0.001, 0.0015, 0.0015, 0.01, and 0.0005, respectively. The samples from the exposed and the control sites were processed simultaneously and in triplicate.
Statistical analysis
A two-factor analysis of variance (model I fixed effects, Zar 2010) was performed to assess the effect of the site (control or exposed), the treatment (mechanical scarification or no treatment), and interaction site × treatment on the germination of seeds of V. campechiana. A Tukey test was performed to find significant mean differences between sites and between treatments (Zar 2010). Students’ t tests were performed to evaluate the effect of the site of origin of the seeds (control and exposed) on their biomass and cover.
Two-factor analysis of variance was used to determine the effect of exposure time (60, 120, 180, 240, 300, 360 days), treatment (control and exposed), and the interaction time × treatment on the variations of 18 morphological characters (17 macro and one micro). Afterwards, a Tukey test was carried out to determine significant differences between pairs of average values of morphological characters in both treatments (Zar 2010).
The same analysis was carried out to evaluate the effect of exposure time, treatment (control and exposure), and interaction time × treatment on the accumulation of Cr, Cu, Fe, Pb, and Zn (except for Cd, which was not detected) in the roots and leaves of individuals of V. campechiana. A Tukey test was also used to determine significant differences in the average concentration of each metal over time by plant structure analyzed, for both treatments (Zar 2010).
The Mann Whitney U analysis was used to assess the effect of seed origin (control or exposed substrate) on the accumulation of HMs in the testa and embryo. All analyses were performed using the STATISTICA 8 program (StatSoft 2004).
The capacity of V. campechiana to phytoextract HMs was evaluated using two indices: the bioconcentration factor (BCF), which determines the efficiency of the plant in the accumulation of substrate metals in its tissue (Yoon et al. 2006), and the translocation factor (FT), which measures the efficiency of the plant in the transportation of metals from the root to the aerial parts (Yoon et al. 2006). These indices are calculated as follows:
where Cfoliar is the concentration of the metal in the leaf tissue, Ctailing is the bioavailable concentration in the tailing, and Croot is the concentration of the metal in the root tissue. It has been reported that if a plant has FT values > 1, the species is considered an accumulator of the analyzed metal (Yoon et al. 2006; Covarrubias and Cabriales 2017).
Results
Germination and morphological characteristics of V. campechiana seeds from the control site and the exposed site (tailing mine)
The germination experiment showed that the site of origin of the individual plants (control vs exposed) did not influence the germination percentage of the seeds. In contrast, the pre-germinative treatment did significantly change germination percentages. There was no significant effect of the interaction site × treatment on germination percentages (Table 2). The results indicate that, considering both sites of origin (control and exposed), only 12% of the seeds germinated when they did not receive pre-germinative treatment. In contrast, when the seeds were subjected to mechanical scarification, the percentage of seed germination increased to more than 90% in both sites (Table 2).
The site of origin of the individual plants did have an effect on seed biomass and surface area. The results indicate that V. campechiana seeds from the site exposed to heavy metals showed statistically lower values for both characters analyzed compared with seeds from the control site (Table 3).
Morphological and size changes in individuals of V. campechiana growing on tailing substrate or control substrate
In general, the results show that the time of exposure (t), the treatment (T), and the interaction between these two factors (t × T) had a significant effect on all the size and macro- and micromorphological characters analyzed during 360 days in individual plants of V. campechiana growing on a greenhouse under two different treatments (tailing substrate and control substrate). The treatment had no significant effect on the diameter of the petiole (DP), while the interaction between factors (t × T) had no significant effect on length (RL), dry biomass, and fresh root biomass (Table 4).
Size characters
All the evaluated characters of V. campechiana individuals grown on the control substrate showed a significant increase over time (360 days): root length (RL), stem length (SL), fresh root biomass (FRB), dry root biomass (DRB), fresh leaf biomass (FLB), dry leaf biomass (DLB). In contrast, in the plants grown on the tailing substrate, 50% of the characters (FRB, DRB, and DLB) showed a significant increase, while the remaining 50% of the characters (RL, SL, and FLB) showed no changes over time (Table 4).
The fresh and dried root biomass did not change significantly between both treatments, remaining unchanged in both treatments at the beginning and the end of the experiment. In contrast, the dry and fresh leaf biomass was significantly lower in plants grown on tailing substrate. In general, the RL did not change between treatments over time, while the SL was significantly lower in tailing substrate individuals (Table 4).
Macro-morphological characters
In the individuals of V. campechiana grown on the control substrate treatment, 54.5% of the evaluated characters had a significant increase over time (360 days): leaf blade length (LBL), petiole diameter (PD), length of the intermediate vein (LIV), leaf-shape characters such as the 1/3 apical width (1/3AW) and the 1/3 basal width (1/3BW), as well as coverage of leaf blade (CLB). Moreover, 27.3% of the characters decreased in size over time: width of the intermediate vein (WIV), length of the intermediate leaflet (LIL), and width of the intermediate leaflet WIL. The width of the leaf blade (WLB) remained more or less constant over time, while the length of the petiole (LP) showed oscillatory changes (Table 4). In the individuals established in the tailing substrate, 45.4% of the characters did not show changes over time: WLB, LP, WIV, WIL, and CLB. In contrast, 54.6% of the characters showed oscillatory changes over time: LBL, PD, LIV, 1/3BW, 1/3AW, and LIL (Table 4).
When comparing individuals from V. campechiana between treatments, the values of 63.6% of the analyzed characters decreased over time in the plants grown on the tailing substrate compared with the plants grown on the control substrate. Moreover, 27.3% of the characters showed an inverse pattern (WIV, LIL, and WIL), while the PD character did not change between treatments (Table 4).
Micro-morphological characters
With respect to micromorphology, the stomatal index (SI) decreased over time in individuals growing on the control substrate. Individuals growing on tailing substrate showed an inverse pattern. However, at the end of the treatment, there were no statistically significant differences in SI values between substrates (Table 4).
Heavy metal concentration in roots, leaves and seeds of V. campechiana
Roots
In general, the presence of Cr, Cu, Fe, Pb, and Zn was detected in the roots of individuals of V. campechiana, but Cd and Mn were not detected. The analysis of variance showed a significant effect of time (t), treatment (T), and interaction (t × T) on the bioaccumulation of four metals (Cr, Cu, Pb, and Zn) in the root of individuals of V. campechiana. In contrast, the aforementioned variables did not have a significant effect on the concentration of Fe in the roots (Table 5).
The bioaccumulation of Cr was only recorded in the roots of plants grown on tailing substrate, with a significant increase in the concentration with the time of exposure. The concentration of Cu in the roots was statistically higher in individuals growing in the control substrate, compared with those grown in tailing substrate. The concentration of Cu in the control substrate individuals remained constant over time, while plants growing in tailing substrate showed a significant increase in the concentration of Cu with the time of exposure. In general, there was no significant effect of time, treatment, and interaction (t × T) on the bioaccumulation of Fe. With respect to Pb, its concentration was significantly higher in the roots of the plants growing in tailing substrate compared with the plants growing in the control substrate. The concentration of Pb in individuals growing in the control substrate did not change significantly over time, while the roots of the plants growing in tailing substrate showed an increase in the concentration of Pb over time. Similarly, the concentration of Zn in the roots of the plants growing in tailing substrate was significantly higher than in the plants growing in the control substrate. The plants growing in the control substrate showed a significant reduction in the concentration of Zn over time; in the plants growing in the tailing substrate, the concentration remained constant (Table 5).
Leaves
In general, the presence of Cr, Cu, Fe, Pb, and Zn was detected in the leaves of individuals of V. campechiana, but the presence of Cd was not. The analysis of variance showed a significant effect of time, treatment, and interaction of factors on the concentration of Cr and Zn in the leaves of individuals of V. campechiana. Cu did not respond to the interaction of factors (t × T), while Fe did not respond to time of exposure or interaction (t × T), and Pb did not respond to time of exposure (Table 5).
The results indicate that the bioaccumulation Cr occurred only in the leaves of plants grown on the exposed substrate, with a significant increase in the concentration with the time of exposure. The concentration of Cu in leaf tissue was statistically higher in individuals growing in the control substrate compared with those grown in the tailing substrate. The concentration of Cu in the control substrate plants remained constant over time, while plants growing in tailing substrate showed a significant increase in Cu with time of exposure. Moreover, there was no significant effect of time and interaction of factors (t × T) on the bioaccumulation of Fe in leaf tissue. In contrast, there was a significant effect of the treatment on the bioaccumulation of Fe. The Tukey test (P < 0.05) showed significant differences in the concentration of Fe at 300 and 360 days, with higher concentrations in the plants growing in the control substrate.
With respect to Pb, its concentration was significantly higher in the leaves of plants growing in exposed substrate compared with plants growing in the control substrate. However, the concentration of this metal remained constant over time in plants growing in the control substrate. Moreover, the concentration of Pb showed a tendency to increase over time in the tailing substrate plants, it being significantly higher at 360 days.
Similarly, the concentrations of Zn in the leaves of plants growing in exposed substrate were significantly higher than in those growing in the control substrate. The concentration of Zn showed a significant decrease over time in the plants growing in the control substrate, while concentration remained constant (Table 5) in the plants growing in exposed substrate.
Seeds
Regarding the concentration of heavy metals in the seeds of V. campechiana, there was bioaccumulation of Cu, Fe, and Zn in the testa and embryo, but no bioaccumulation of Cr, Cd, and Pb.
There were no significant differences in the concentration of Cu and Fe between the testa and the embryo of the seeds from control sites, but there were significant differences in the concentration of Zn between these seed structures, with the highest concentration of this metal found in the embryo. The seeds from exposed sites did not show significant differences in the concentration of the three metals evaluated between the testa and the embryo. There were significant differences in the concentration of Cu and Zn in the seed testa between sites (control vs exposed), with Cu showing the highest concentration values in the testa of seeds from control sites. In the case of Zn, the highest values were found in the testa of seeds from the exposed sites. In the case of the embryo, no significant differences were found in the concentration of the metals evaluated in the seeds from both sites. Finally, no significant differences were found between sites in the concentration of Fe in the testa and embryo of seeds (Table 6).
The translocation factor and heavy metal enrichment in the roots and leaves of V. campechiana plants growing in substrate exposed to metals
Enrichment (BCF) of Cu, Fe, and Pb was recorded in the roots and leaves of V. campechiana plants growing in tailing substrate. In contrast, no enrichment of Cr and Zn was detected in any of the plant structures analyzed (Table 7). In roots, the metal enrichment factor showed the following pattern: Pb > Cu > Fe. In leaf tissue, the pattern of the enrichment factor was as follows: Cu > Pb > Fe (Table 7). The pattern of the translocation factor (FT) was as follows: Cr > Pb = Cu > Fe > Zn (Table 7).
Discussion
Bioaccumulation of heavy metals in V. campechiana
There are few studies that evaluate the transport and accumulation of metals in the seeds of wild plant species for ecotoxicological purposes. The present study found that V. campechiana seeds from both the control and the exposed sites accumulated only essential metals (Cu, Fe, and Zn) and that the concentration of metals in the embryo of seeds did not differ between sites. In contrast, the bioaccumulation of metals in the testa of seeds from the control site had a higher concentration of Cu and Zn. Our results are similar to those reported by Tyler and Zohlen (1998) for several herbaceous species, Achillea millefolium L. (Asteraceae), Arctium tomentosum Mill. (Asteraceae), Arenaria serpyllifolia L. (Caryophyllaceae), Cerastium semidecandrum L. (Caryophyllaceae), Filipendula ulmaria (L.) Maxim. (Rosaceae), Hypericum maculatum Crantz. (Hypericaceae), and Laserpitium latifolium (L.) (Apiaceae), the seeds of which bioaccumulate high concentrations of essential metals such as Mn, Cu, Fe, and Zn. Furthermore, the present study agrees with the study by Waters and Sankaran (2011), who reported an increase in the transport of micronutrients, mainly Fe and Zn, to the seeds of plants with nutritional potential.
It has been documented that the transport of minerals to the reproductive parts and to the seeds is carried out through the phloem (Zhang et al. 2007); in the leaves, it is carried out through the xylem. Recent studies have shown that the nicotianamine (NA) molecule transports various metals through the phloem, including Cu, Fe, and Zn, towards reproductive structures, including seeds (Grillet et al. 2014). This suggests that V. campechiana bioaccumulates and translocates essential metals such as Cu, Fe, and Zn, since these metals participate in the development of seeds and seedlings when germination begins (Stacey et al. 2008; Grillet et al. 2014).
All the analyzed metals (Cu, Fe, Zn, Cr, and Pb) bioaccumulated in the root and leaf tissue of V. campechiana, except for Cd. Three of these metals are considered essential (Cu, Fe, and Zn); the other two are considered non-essential (Cr and Pb). The accumulation pattern was as follows: Pb > Fe > Cr > Cu > Zn. It was also found that the accumulation of metals, mainly Cr and Pb, was greater in plants growing on exposed substrates than in control plants. This coincides with what has been found in other phylogenetically related plant species, such as Acacia robeorum Maslin. (Fabaceae), which has been reported to bioaccumulate metals such as Al, Fe, Mn, Cu, Zn, and Mo. These results suggest that the bioaccumulation of HMs in the leaf tissue of plant species inhabiting sites contaminated with heavy metals could be a detoxification strategy (He et al. 2012).
In the present study, only plants growing in substrate exposed to HMs accumulated Cr in root and leaf tissue, the latter being where Cr accumulated in greater amounts. The absorption of Cr through the roots may be due to the union of organic acids with the insoluble metals found in the soil, making them available for plant absorption (Panda and Choudhury 2005). The accumulation of Cr occurs when it is immobilized in vacuoles in the root cells, which makes it less toxic to the plant (Shanker et al. 2005). Moreover, the high concentration levels of Cr in the leaf tissue of V. campechiana is supported by the study conducted by Skeffington et al. (1976) on the Hordeum vulgare L. (Poaceae) species. The authors describe that Cr can enter the aerial parts of the plant through molecules that transport essential elements such as Fe, S, and P and that it can be translocated to the leaves through the xylem.
With respect to Pb, the present study showed that its concentration in root and leaf tissue increased over time in plants growing on tailing substrate, with the highest concentration found in the leaves. Similar results have been described for other shrub-like species similar to V. campechiana; for example, Brickellia veronicifolia (Kunth) A.Gray. (Asteraceae) accumulates Cd, Cu, Ni, and Pb in its leaf tissue, with Pb showing the highest concentration (Hernández-Acosta et al. 2009). Salas-Luévano et al. (2009) found high concentrations of Pb in the leaf tissue of Buddleja scordioides Kunth. (Scrophulariaceae), Mimosa aculeaticarpa Ortega. (Fabaceae), and Acacia schaffneri (S.Watson) F.J.Herm. (Fabaceae). This could be due to the fact that Pb inhibits the transport of essential metals such as Cu, Fe, and Zn (Patra et al. 2004), which increases in concentration over time and thus its translocation to the aerial parts of the plant.
In V. campechiana, low concentration levels of Cu, Fe, and Zn were found in the root and leaf tissue. The explanation of these results could be that the absorption of Pb replaces that of Cu, Fe, and Zn, probably modifying the activity and permeability of the membranes, making them unavailable for absorption and transport within the plant (Patra et al. 2004).
Changes in the morphological characters of the roots, stems, and leaves of V. campechiana due to exposure to heavy metals
Most ecotoxicological studies analyze the effect of metals on the morphology of herbaceous species, with few studies focusing on shrub- or tree-like plants. In addition, most of these studies focus on three morphological characters: root length, root biomass, and leaf biomass (Prasad et al. 20011; Maldonado-Magaña et al. 2011). The development of effective phytoremediation strategies for contaminated sites requires the study of plant species with different life forms. Herbaceous life forms, for example, have a short life cycle that can last up to 24 months; shrubs and trees, by contrast, have a long life cycle that can last several years, and this influences their efficiency in the accumulation of HMs.
In the present study, seventeen of 18 morphological characters of V. campechiana decreased in plants exposed to HMs with respect to control plants. These results show that the HMs interfere in the plant growth, causing a reduction in the V. campechiana growth. Similar results were reported by Tovar-Sánchez et al. (2018), who observed a decrease in the morphological values of the roots and leaves of Zea mays L. (Poaceae) associated with sites contaminated by tailings in Taxco de Alarcón, GRO, Mexico. Furthermore, Hernández-Lorenzo (2015) found a decrease in the leaf morphological values of Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C.Johnst. (Fabaceae) associated with tailings in Huautla, MOR, Mexico. In other shrub-like species [Salix viminalis L. (Salicaceae), Caesalpinia pulcherrima (L.) Sw. (Fabaceae), Albizia lebbeck (L.) Benth. (Fabaceae), Acacia holosericea Cunn. ex G.Don. (Fabaceae), Leucaena leucocephala (Lam.) de Wit. (Fabaceae) and Vachellia farnesiana (L.) Wight & Arn. (Fabaceae)] exposed to Cr and Pb, there have been reports of a decrease in root length and leaf biomass (Panda and Patra 2000; Prasad et al. 2001; Iqbal et al. 2001; Suseela et al. 2002; Shanker 2003; Shanker et al. 2005; Maldonado-Magaña et al. 2011). Some studies have reported that the reduction in the plant morphological characters may be due to the fact that HMs could interfere with metabolic processes and are potentially toxic. Resulting in growth abnormalities (Bini et al. 2012) as a weak plant growth, yield depression, and may be accompanied by disorders in plant metabolism such as reduction of the meristematic zone (Maleci et al. 2001). In addition, other studies have found that nonessential HMs such as Cr and Pb inhibit the mitosis process in root cells, reducing the extension of this tissue. For example, it has been reported that nonessential HMs such as Cr and Pb inhibit the mitosis process in root cells, reducing the extension of this tissue. Prasad et al. (2001) reported a toxic effect of Cr, which inhibited primary root growth and suppressed the growth of new secondary roots. Studies conducted by Shanker et al. (2005) found that Cr inhibits root growth in Caesalpinia pulcherrima (L.) Sw. (Fabaceae), Triticum aestivum L. (Poaceae), and Vigna radiata (L.) R.Wilczek. (Fabaceae) Furthermore, the absorption of Pb causes a decrease in the rate of root growth and affects the root branching pattern (Maldonado-Magaña et al. 2011). For example, in Picea abies (L.) H.Karst. (Pinaceae), the development of secondary roots is particularly sensitive to exposure to Pb (Godbold and Kettner 1991; DalCorso 2012).
Changes in the leaf morphology of plant species that are exposed to pollution with HMs can be explained by the biochemical changes that form part of the adaptive mechanisms developed by plants to tolerate or bioaccumulate these metallic elements in their tissues (Meharg 1994). The bioaccumulation of HMs in leaf tissue triggers a detoxification mechanism in which metals bind to a ligand (chelation) such as sulfhydryl, phosphate, carboxyl, and hydroxyl groups and to peptides such as phytochelatins and metallothioneins (Rauser 1995; Cobbett 2000). The HMs are surrounded by ligands, forming a complex that is immersed in a chemical interaction that keeps it in electronic balance while it is transferred to inactive cell compartments, mainly vacuoles (Yong-Eui et al. 2004; Yang et al. 2005; Rodríguez-Serrano et al. 2008; Lin and Aarts 2012).
In the present study, the stomatal index decreased in individuals of V. campechiana grown in substrate exposed to HMs. Similar results were reported for P. laevigata exposed to HMs (Hernández-Lorenzo 2015). Thakur (1990) mentioned that the decrease in the number of stomata per unit area (mm2) prevents excessive perspiration in plants and increases stomatal resistance. Although few studies have focused on the stomata of plants exposed to metals, it has been reported that occlusive cells are sensitive to chemical stress, so that the position and number of stomata can change or a mechanism for fall of leaves could be developed to protect the plant against the effects of metals (Zimmermann 2001).
With respect to the seeds of V. campechiana, the size and biomass of the seeds was smaller in the sites exposed to metals. However, germination was not affected, with similar values in both treatments. This could be due to the fact that seeds from sites exposed to HMs only accumulated essential elements (Cu, Fe, and Zn), which are involved in the germination process and the development of seedlings.
Potential use of V. campechiana as a phytoremediation species in sites contaminated with metals
V. campechiana is a shrub species that is distributed in disturbed places such as sites contaminated by HMs. This species commonly shares its habitat with V. farnesiana, a taxonomically close species that has been reported to accumulate HMs as As, Cu, Mn, Ni, Pb, V, and Zn (Maldonado-Magaña et al. 2011; Alcantara-Martinez et al. 2016; Cervantes-Ramírez et al. 2018). However, the phytoremediation potential of V. campechiana is still unknown. In general, it has been documented that the plant species used to phytoremediate environments contaminated by HMs belong to the following families: Asteraceae, Brassicaceae, Caryophyllaceae, Flacourtiaceae, Lamiaceae, Poaceae, Violaceae, and Euphorbiaceae (Prasad 2003; Mahar et al. 2016), these being mostly herbaceous species. The results obtained in the present study suggest that V. campechiana has phytoremediation potential for sites contaminated by Cr, Cu, and Pb. The reasons that justify this suggestion are as follows: 1) V. campechiana is an accumulator species of Cr, Cu, and Pb. Olguín and Sánchez-Galván (2012) and Ali et al. (2013) propose that a plant can be considered an accumulator if its translocation factor (TF) is equal to or greater than 1. In the present study, in spite of V. campechiana growing under experimental conditions, it showed TF values greater than one for Cr (2.24), Pb (1.23), and Cu (1.22). These levels are similar to the TF found in species considered as accumulators that grow directly in contaminated environments such as Gentiana pennelliana Fernald [(Gentianaceae) (TF Zn = 1.2)], Cyperus esculentus L [(Cyperaceae) (TF Pb = 1.6, TF Zn = 1.1)], Phyla nodiflora (L.) Greene [(Verbenaceae) (TF Cu = 12.0, TF Zn = 1.1)], Rubus fruticosus L [(Rosaceae) (TF Cu = 5.6), Sesbania herbacea (Mill.) McVaugh [(Fabaceae) (TF Cu = 4.0)], and Limnocharis flava (L.) Buchenau [(Alismataceae) (TF Cd = 1.3)] (Yoon et al. 2006; Abhilash et al. 2009). 2) The bioaccumulation factor (BCF) for Cu in roots and leaves was 7.6 and 39.4, respectively. The study by Lin et al. (2003) supports the results obtained in the present work. They reported bioaccumulation of Cu in Helianthus annuus L. (Asteraceae), with a concentration that was 2 to 10 times higher than the concentration of Cu in the soil. With respect to Pb, it showed a BCF in roots of 15.1 and of 17.4 in leaf tissue. 3) In V. campechiana, the accumulation of Cr and Pb increased over time (12 months) in leaf tissue; thus, long-term studies could record a higher accumulation of Cr and Pb. 4) Seeds do not accumulate nonessential metals such as Cr and Pb, and seed germination is not affected by exposure to heavy metals. Unlike other species such as Sanvitalia procumbens Lam. (Asteraceae), the germination percentage of seeds from sites exposed to HMs decreases compared with the germination percentage of seeds from control sites (Rosas-Ramírez 2018). 5) Throughout the study (12 months), no mortality was recorded in the plants exposed to HMs. 6) The root biomass of V. campechiana plants was not significantly affected by the bioaccumulation of heavy metals.
Conclusion
In the present study V. campechiana chronically exposed to mine tailings in greenhouse/experimental condition showed high levels of HMs translocation, its ability to bioaccumulate non-essential metals in roots and leaves and changes in the macro- and micromorphological characters. These findings suggest that this plant may be a suitable candidate for use in phytoremediation studies in contaminated environments mainly for Cr, Cu, and Pb. Future studies in situ are necessary for complementing the role of V. campechiana as a phytoremediation plant. Also, we consider that conducting ecotoxicological studies on new plant species with different life forms provides useful information to arrive at effective phytoremediation strategies for soils contaminated with HMs. A detailed characterization of the effects of these xenobiotics on the exposed plants would allow the selection of the species whose macro- and micro morphological characters are affected the least by exposure to HMs. Furthermore, different species can extract different metals, so it is necessary to combine plant species to develop more effective strategies that can be used in environments polluted with more than one HMs.
References
Abhilash PC, Pandey VC, Srivastava P, Rakesh PS, Chandran S, Singh N, Thomas AP (2009) Phytofiltration of cadmium from water by Limnocharis flava (L.) Buchenau grown in free-floating culture system. J Hazard Mater 170:791–797. https://doi.org/10.1016/j.jhazmat.2009.05.035
Alcantara-Martinez N, Guizar S, Rivera-Cabrera F, Anicacio-Acevedo BE, Buendia-Gonzalez L, Volke-Sepulveda T (2016) Tolerance, arsenic uptake, and oxidative stress in Acacia farnesiana under arsenate-stress. Int J Phytoremediation 18:671–678. https://doi.org/10.1080/15226514.2015.1118432
Ali H, Khan E, Sajad MA (2013) Phytoremediation of heavy metals-concepts and applications. Chemosphere 91:869–881. https://doi.org/10.1016/j.chemosphere.2013.01.075
Arce MDLR (2001) El género Acacia (leguminosae, mimosoideae) en el Estado de Oaxaca, México. An Jard Bot Madr 58(2):251–275
Armienta MA, Ongley LK, Rodríguez R, Cruz O, Mango H, Villaseñor G (2008) Arsenic distribution in mesquite (Prosopis laevigata) and huizache (Acacia farnesiana) in the Zimapán mining area, México. Geochem Explor Environ Anal 8:191–197
Barceló J, Poschenrieder C (2003) Phytoremediation: principles and perspectives. Contrib Sci 2(3):333–344
Baskin JM, Baskin CC (2004) A classification system for seed dormancy. Seed Sci Res 14:1–16
Bini C, Wahsha M, Fontana S, Maleci L (2012) Effects of heavy metals on morphological characteristics of Taraxacum officinale Web growing on mine soils in NE Italy. J Geochem Explor 123:101–108. https://doi.org/10.1016/j.gexplo.2012.07.009
Brooks RR, Lee J, Reeves RD, Jaffré (1977) Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants. J Geochem Explor 7:49–57. https://doi.org/10.1016/0375-6742(77)90074-7
Calow P (1993) General principles and overview. In: Calow P (ed) Handbook of ecotoxicology. Blackwell Scientific Publications, London, pp 1–5
Cappa JJ, Pilon-Smits EAH (2014) Evolutionary aspects of elemental hyperaccumulation. Plant 239:267–275. https://doi.org/10.1007/s00425-013-1983-0
Cervantes SMA, Sotelo BME (2002) Guías técnicas para la propagación sexual de 10 especies latifoliadas de selva baja caducifolia en el estado de Morelos 30:1–35
Cervantes-Ramírez LT, Ramírez-López M, Mussali-Galante P, Ortiz-Hernández ML, Sánchez-Salinas E, Tovar-Sánchez E (2018) Heavy metal biomagnification and genotoxic damage in two trophic levels exposed to mine tailings: a network theory approach. Rev Chil Hist Nat 91:1–13. https://doi.org/10.1186/s40693-018-0076-7
Cobbett CS (2000) Phytochelatin biosynthesis and function in heavy-metal detoxification. Curr Opin Plant Biol 3:211–216. https://doi.org/10.1016/S1369-5266(00)80067-9
Cortés-Jiménez EV, Mugica-Álvarez V, González-Chávez MCA, Carrillo-González R, Gordillo MM, Mier MV (2012) Natural revegetation of alkaline tailing heaps at Taxco, Guerrero, México. Int J Phytoremediation 15:127–141. https://doi.org/10.1080/15226514.2012.683208
Covarrubias SA, Cabriales JJP (2017) Contaminación ambiental por metales pesados en México: Problemática y estrategias de fitorremediación. Rev Int Contam Ambie 33:7–21. https://doi.org/10.20937/RICA.2017.33.esp01.01
DalCorso G (2012) Heavy metal toxicity. In: Furini A (ed) Plant and heavy metals. Springer Science & Business Media, Pisa, pp 1–25
Furini A (2012) Plants and heavy metals. Springer Science & Business Media, Pisa
Godbold DL, Kettner C (1991) Lead influences root growth and mineral nutrition of Picea abies seedlings. J Plant Physiol 139:95–99. https://doi.org/10.1016/S0176-1617(11)80172-0
Gold KP, León-Lobos Y, Way M (2004) Manual de recolección de semillas de plantas silvestres para conservación a largo plazo y restauración ecológica. Instituto de Investigaciones Agropecuarias, Centro Regional de Investigación Intihuasi, La Serena Boletín INIA 10:62
Grillet L, Mari S, Schmidt W (2014) Iron in seeds–loading pathways and subcellular localization. Front Plant Sci 4:535. https://doi.org/10.3389/fpls.2013.00535
Gutiérrez ME, Moreno M (1995) Los residuos de la minería mexicana. In: Garfias FJ, Ayala I, Weber LB (eds) Taller para el desarrollo sustentable: residuos peligrosos. México, pp 37–44
Gutiérrez-Ruiz M, Romero F, González-Hernández G (2007) Suelos y sedimentos afectados por la dispersión de jales inactivos de sulfuros metálicos en la zona minera de Santa Bárbara, Chihuahua, México. Rev Mex Cienc Geol 24(2):170–184
He H, Bleby TM, Veneklaas EJ, Lambers H, Kuo J (2012) Morphologies and elemental compositions of calcium crystals in phyllodes and branchlets of Acacia robeorum (Leguminosae: Mimosoideae). Ann Bot 109:887–896. https://doi.org/10.1093/aob/mcs004
Hernández-Acosta E, Mondragón-Romero E, Cristobal-Acevedo D, Rubiños-Panta JE, Robledo-Santoyo E (2009) Vegetación, residuos de mina y elementos potencialmente tóxicos de un jal de Pachuca, Hidalgo, México. Rev Chapingo Ser Cie 15(2):109–114
Hernández-Lorenzo B (2015) Análisis de la anatomía y morfología de Prosopis laevigata, por acumulación de metales pesados en la sierra de Huautla, Morelos. Facultad de Ciencias Biológicas, Universidad Autónoma del Estado de Morelos, Cuernavaca
Iqbal MZ, Saeeda S, Muhammad S (2001) Effects of chromium on an important arid tree (Caesalpinia pulcherrima) of Karachi city, Pakistan. Ekol Bratislava 20:414–422
Kabata-Pendias A (2000) Trace elements in soils and plants. CRC Press, Boca Raton
Lin YF, Aarts MG (2012) The molecular mechanism of zinc and cadmium stress response in plants. Cell Mol Life Sci 69(19):3187–3206. https://doi.org/10.1007/s00018-012-1089-z
Lin J, Jiang W, Liu D (2003) Accumulation of copper by roots, hypocotyls, cotyledons and leaves of sunflower (Helianthus annuus L.). Bioresour Technol 86:151–155. https://doi.org/10.1016/S0960-8524(02)00152-9
Macnair MR (2003) The hyperaccumulation of metals by plants. Adv Bot Res 40:63–105. https://doi.org/10.1016/S0065-2296(05)40002-6
Mahar A, Wang P, Ali A, Awasthi MK, Lahori AH, WangQ ZZ (2016) Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: a review. Ecotoxicol Environ Saf 126:111–121. https://doi.org/10.1016/j.ecoenv.2015.12.023
Maldonado-Magaña A, Favela-Torres E, Rivera-Cabrera F, Volke-Sepulveda TL (2011) Lead bioaccumulation in Acacia farnesiana and its effect on lipid peroxidation and glutathione production. Plant Soil 339:377–389. https://doi.org/10.1007/s11104-010-0589-6
Maleci L, Bini C, Paolillo A (2001) Chromium (III) uptake by Calendula arvensis L. and related phytotoxicity. Proc. VI ICOBTE, Guelph, On., p. 384 (abstract)
Marrero-Coto J, Amores-Sánchez I, Coto-Pérez O (2012) Fitorremediación, una tecnología que involucra a plantas y microorganismos en el saneamiento ambiental. ICIDCA 46(3):52–61
Martínez-Pacheco M (2008) Evaluación de los efectos genotóxicos de metales presentes en el agua de bebida de la población de Huautla, Morelos. Facultad de ciencias. Universidad Nacional Autónoma de México, México
Meharg AA (1994) Integrated tolerance mechanisms: constitutive and adaptive plant responses to elevated metal concentrations in the environment. Plant Cell Environ 17:989–993
Mendez MO, Maier RM (2008) Phytoremediation of mine tailings in temperate and arid environments. Environ Sci Biotechnol 7:47–59. https://doi.org/10.1007/s11157-007-9125-4
Mireles F, Davila JI, Pinedo JL, Reyes E, Speakman RJ, Glascock MD (2012) Assessing urban soil pollution in the cities of Zacatecas and Guadalupe, Mexico by instrumental neutron activation analysis. Microchem J 103:158–164
Mussali-Galante P, Tovar-Sánchez E, Valverde M, del Castillo ER (2013) Biomarkers of exposure for assessing environmental metal pollution: from molecules to ecosystems. Rev Int Contam Ambie 29(1):117–140
Olguín EJ, Sánchez-Galván G (2012) Heavy metal removal in phytofiltration and phycoremediation: the need to differentiate between bioadsorption and bioaccumulation. New Biotechnol 30(1):3–8. https://doi.org/10.1016/j.nbt.2012.05.020
Panda SK, Choudhury S (2005) Chromium stress in plants. J Plant Physiol 17(1):95–102. https://doi.org/10.1590/S1677-04202005000100008
Panda SK, Patra HK (2000) Nitrate and ammonium ions effect on the chromium toxicity in developing wheat seedlings. Proc Natl Acad Sci India 70:75–80
Patra M, Bhowmik N, Bandopadhyay B, Sharma A (2004) Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ Exp Bot 52:199–223. https://doi.org/10.1016/j.envexpbot.2004.02.009
Prasad MNV (2003) Phytoremediation of metal-polluted ecosystems: hype for commercialization. Russ J Plant Physl 50(5):686–701
Prasad MNV, Greger M, Landberg T, Acacia nilotica L (2001) Bark removes toxic elements from solution: corroboration from toxicity bioassay using Salix viminalis L. in hydroponic system. Int J Phytoremediation 3(1):289–300. https://doi.org/10.1080/15226510108500060
Prieto J, Ramírez GC, Román A, Prieto F (2009) Contaminación y fitotoxicidad en plantas por metales pesados provenientes de suelos y agua. Tro Subtro Agroecosyst 10:29–44
Rascio N, Navari-Izzo F (2011) Heavy metal hyperaccumulating plants: how and why do they do it? And what makes them so interesting? Plant Sci 180:169–181
Rauser WE (1995) Phytochelatins and related peptides. Structure, biosynthesis, and function. Plant Physiol 109:1141
Rengel M, Gil F, Montaño J (2011) Crecimiento y dinámica de acumulación de nutrientes en caña de azúcar. I macronutrientes. Bioagro 23:5–8
Rico ML (2001) El género Acacia (Leguminosae, Mimosoideae) en el Estado de Oaxaca, México. An Jard Bot Madr 58:251–302
Rodríguez-Serrano M, Martínez-de la Casa N, Romero-Puertas M, Del Río L, Sandalio L (2008) Toxicidad del cadmio en plantas. Ecosistemas 17:139–146
Rosas-Ramírez ME (2018) Relación entre la bioacumulación de metales pesados y la concentración de clorofila en Sanvitalia procumbens. Facultad de Ciencias Biológicas. Universidad Autónoma del Estado de Morelos, Cuernavaca
Salas-Luévano MA, Manzanares-Acuña E, Letechipía-de León C, Vega-Carrillo HR (2009) Tolerant and hyperaccumulators autochthonous plant species from mine tailing disposal sites. J Exp Sci Asia 23(1):27–32
Salisbury FT (1968) Las plantas vasculares: forma y funciones. Herrero Hermanos Sucesores, México
Secretaria de Economía (2011) Panorama Minero del estado de Morelos. Servicio Geológico Mexicano, serie panorama minero de los estados, Pachuca
Shanker AK (2003) Physiological, biochemical and molecular aspects of chromium toxicity and tolerance in selected crops and tree species. PhD Thesis, Tamil Nadu Agricultural University, Coimbatore, India
Shanker AK, Cervantes C, Loza-Tavera H, Avudainayagam S (2005) Chromium toxicity in plants. Environ Int 31:739–753. https://doi.org/10.1016/j.envint.2005.02.003
Shiqi L, Yang B, Kou Y, Zeng J, Wang R, Xiao Y, Li F, Lu Y, Mu Y, Zhao C (2018) Assessing the difference of tolerance and phytoremediation potential in mercury contaminated soil of a non-food energy crop, Helianthus tuberosus L. (Jerusalem artichoke). Peer J 6:1–18. https://doi.org/10.7717/peerj.4325
Skeffington RA, Shewry PR, Peterson PJ (1976) Chromium uptake and transport in barley seedlings (Hordeum vulgare L.). Plant 132:209–214
Stacey MG, Patel A, McClain WE, Mathieu M, Remley M, Rogers EE, Gassmann W, Blevins DG, Stacey G (2008) The Arabidopsis AtOPT3 protein functions in metal homeostasis and movement of iron to developing seeds. Plant Physiol 146:589–601
StatSoeft, Inc (2004) STATISTICA (data analysis software system), version 7. www.statsoft.com
Suseela MR, Sinha S, Singh S, Saxena R (2002) Accumulation of chromium and scanning electron microscopic studies in Scirpus lacustris L. treated with metal and tannery effluent. Bull Environ Contam Toxicol 68:540–548. https://doi.org/10.1007/s001280288
Thakur P (1990) Different physiological responses of tomato (Lucopersicom esculetum Mill.) cultivars to drought. Acta Physiol Plant 12:175–182
Tovar-Sánchez E, Cervantes-Ramírez T, Castañeda-Bautista J, Gómez-Arroyo S, Ortiz-Hernández L, Sánchez-Salinas E, Mussali-Galante P (2018) Response of Zea mays to multimetal contaminated soils: a multibiomarker approach. Ecotoxicology 27:1161–1177. https://doi.org/10.1007/s10646-018-1974-9
Tovar-Sánchez E, Suarez-Rodríguez R, Ramírez-Trujillo A, Valencia-Cuevas L, Hernández-Plata I, Mussali-Galante P (2019). The use of biosensors for biomonitoring environmental metal pollution, Biosensors for Environmental Monitoring, Toonika Rinken and Kairi Kivirand, IntechOpen, https://doi.org/10.5772/intechopen.84309. Available from: https://www.intechopen.com/books/biosensors-for-environmental-monitoring/the-use-of-biosensors-for-biomonitoring-environmental-metal-pollution
Tyler G, Zohlen A (1998) Plant seeds as mineral nutrient resource for seedlings—a comparison of plants from calcareous and silicate soils. Ann Bot 81:455–459. https://doi.org/10.1006/anbo.1997.0581
Van der Ent A, Baker AJ, Reeves RD, Pollard AJ, Schat H (2013) Hyperaccumulators of metal and metalloid trace elements: facts and fiction. Plant Soil 362:319–334. https://doi.org/10.1007/s11104-012-1287-3
Velasco TJA, de la Rosa PDA, Ramírez IME, Volke ST (2005) Evaluación de tecnologías de remediación para suelos contaminados con metales. Etapa II. secretaria del medio ambiente y recursos naturales, México, pp 1–36
Volke ST, Velsco TA, De la Rosa PA, Solórzano OG (2004) Evaluaciones de tecnologías de remediación para suelos contaminados con metales. Etapa I. Secretaria de Medio Ambiente y Recursos Naturales, México
Volke ST, Velsco TA, De la Rosa PA, Solórzano OG (2005) Evaluaciones de tecnologías de remediación para suelos contaminados con metales. Etapa II. Secretaria de Medio Ambiente y Recursos Naturales, México
Waters BM, Sankaran RP (2011) Moving micronutrients from the soil to the seeds: genes and physiological processes from a biofortification perspective. Plant Sci 180:562–574. https://doi.org/10.1016/j.plantsci.2010.12.003
Yadav SK (2010) Heavy metals toxicity in plants: an overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S Afr J Bot 76:167–179. https://doi.org/10.1016/j.sajb.2009.10.007
Yang X, Feng Y, He Z, Stoffella PJ (2005) Molecular mechanisms of heavy metal hyperaccumulation and phytoremediation. J Trace Elem Med Biol 18:339–353. https://doi.org/10.1016/j.jtemb.2005.02.007
Yong-Eui C, Harada E, Gwang-Hoon K, Eui-Soo Y, Sano H (2004) Distribution of elements on tobacco trichomes and leaves under cadmium and sodium stresses. J Plant Biol 47:75–82
Yoon J, Cao X, Zhou Q, Ma LQ (2006) Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci Total Environ 368:456–464. https://doi.org/10.1016/j.scitotenv.2006.01.016
Zar JH (2010) Biostatistical analysis. Prentice-Hall, Upper Saddle River
Zhang Z, Rengel Z, Meney K (2007) Removal of nutrients from secondary-treated municipal wastewater in wetland microcosms using ornamental plant species. Int J Environ Waste Manag 1:363–375
Zimmermann M (2001) Adaptaciones de plantas a estrés abiótico que les permiten vivir y prosperar en diferentes condiciones ambientales. Rev Creces:1–9
Acknowledgments
Also, we would like to thank Rosalind Pearson Hedge for her comments and English edition that improved our manuscript.
Funding
This research was supported by a CONACyT scholarship grant to M.S.M. (Grant: 307350).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible editor: Elena Maestri
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Santoyo-Martínez, M., Mussali-Galante, P., Hernández-Plata, I. et al. Heavy metal bioaccumulation and morphological changes in Vachellia campechiana (Fabaceae) reveal its potential for phytoextraction of Cr, Cu, and Pb in mine tailings. Environ Sci Pollut Res 27, 11260–11276 (2020). https://doi.org/10.1007/s11356-020-07730-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-020-07730-7