
Abstract
Recently, light traps equipped with light-emitting diodes (LEDs) have been widely applied for controlling nocturnal pest moths. The oriental armyworm, known as Mythimna separata Walker, is an important insect pest in eastern Asia. The present study aimed to evaluate an influence of green light irradiation on antioxidant enzymes and light stress in M. separata adults. We determined total antioxidant capacity (T-AOC) and an activity of superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), and glutathione-S-transferase (GST) in the moth bodies according to an exposure time (0 to 180 min) of a green LED light. By our results, we suggested that the green (520 nm) LED light irradiation could induce a weak oxidative stress in M. separata adult moths, and the moths under the green light could restore the damage caused by this light stress. Additionally, we proposed that the adaptive ability to the light stress varied between the moths of different sexes. Our results may provide a theoretical and scientific basis for elucidating a reason of the phototactic behavior of nocturnal moths, including M. separata adults.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Oxidative stress is a common stress action for all aerobic organisms, and it is closely related to the increase in the production of reactive oxygen species (ROS), including superoxide anion (O2-), hydrogen peroxide (H2O2), hydroxyl radical (OH−), and singlet oxygen (1O2) as a result of electron or energy transfer to oxygen (Barbehenn et al. 2001; Zha and Lei 2012). In general, several environmental stresses, such as temperature, humidity, and light quality, significantly affect insect life because they increase the production and accumulation of ROS through complex biochemical processes in the body ( Li et al. 2015; Ali et al. 2016a, b). The ROS in very low level do not affect normal activity of cell and rather play an important role in cell signaling and defense (Kamata and Hirata 1999; Wang et al. 2001). However, when these free radicals of oxygen reach a high level, they are harmful to DNAs and proteins in the cell, and result in oxidative stress; moreover, excess oxidative stress leads to the cell death (Halliwell and Gutteridge 1999; Davies 2000). Through long-term evolution, insects have developed a suitable antioxidant system employing various antioxidant enzymes. This system counteracts and eliminates the toxicity of ROS, reduces oxidative stress, and finally, maintains the equilibrium between the production and elimination of ROS (Ahmad 1992; Felton and Summers 1995). However, the antioxidant system may not maintain the equilibrium under continuous or increasing stress conditions (Joanisse and Storey 1998; Lalouette et al. 2011). The main enzymes of the antioxidant system in insect are superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD); these enzymes catalyze oxidant radicals into oxygen and water through continuous biochemical reactions. Additionally, glutathione-S-transferase (GST) is a main enzyme, which neutralizes lipid peroxidation products or hydroperoxides to protect the cells of insect (Felton and Summers 1995; Wang et al. 2001). Logically, the activity levels of these enzymes may reflect the oxidative stress level that the insect body receives from the external environment (Meng et al. 2009; Ali et al. 2017). Some studies have shown that ultraviolet (UV) light irradiation induces the high oxidative stress in insects, thereby significantly alternating the activity levels of antioxidant enzymes, such as SOD, CAT, POD, and GST (Meng et al. 2009; Ali et al. 2017; Gao et al. 2017).
The oriental armyworm, known as Mythimna separata or Leucania separata Walker, is an important pest in eastern Asia and causes damage to more than 100 different plant species, including corn, wheat, and rice (Sharma and Davies 1983; Sharma et al. 2002; Wang et al. 2006; Lihuang et al. 2017). Zhao et al. (2016) has reported that the widespread outbreaks of the insect pest are closely related to climate change, especially increases in temperature. Control of this insect pest has been depended on synthesis chemical insecticides. However, the excessive use of the insecticides has led to the agro-ecosystem pollution and the high resistances of the insect pest to the insecticides (Sharma and Davies 1983; Yang et al. 1995). To overcome these problems, recently, some studies have been conducted to search new control method as alternative to chemical insecticides (Huang et al. 2017; Lihuang et al. 2017). The control methods using the light traps equipped with LED light source that highly attract some insect pests have been widely used for controlling agricultural and forest insect pests (Shimoda and Honda 2013; Park and Lee 2017). Our previous research demonstrated that M. separata adult moths have a high sensitivity to the green (520 nm) LED light among several wavelength LED lights (no published; Kim et al.). However, it has not been reported whether light irradiation of visible wavelength could induce oxidative stress in a phototactic insect as UV light, and the response of antioxidant enzymes in the phototactic insect body under visible light irradiation.
In the present study, we attempted to evaluate the influence of green (520 nm) light irradiation on antioxidant enzymes and oxidative stress in M. separata adult moths, and to theoretically analyze the potential relationship between the insect phototactic behavior and light stress based on previous studies and our results.
Materials and methods
Insects
M. separata adults were collected from Hubei Insect Resources Utilization and Sustainable Pest Management Key Laboratory, College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, China. The experimental insects reared at the temperature 25 ± 2 °C, with 65 ± 10% relative humidity (RH) under a photoperiod of 14L/10D in the special insect rearing room. An artificial diet as described by Be (1981) are used in larval stages of the insect. Males and females were selected based on the morphological characteristics of the abdominal terminal segment of pupae. Virgin males and females were separated into different special insect adult rearing cases (50 × 50 × 50 cm) and reared using 10% honey solution as adult feed. During the insect rearing period, all insects were maintained with no exposure to any chemicals.
Treatment of green LED light irradiation
Prior to the experiment, 3-day-old male and female adult moths were selected and individually placed in a cylindrical glass tube (diameter 2 cm; length 8 cm). The moths were divided into five groups; each group contained 15 adults and was maintained in darkness for 2 h. Moths of each group were exposed to green LED light illumination (peak emission 520 ±3 nm; luminance intensity 200~250 lux; luminance irradiance 29.8 ± 2 μW/cm2) for 0, 30, 60, 120, and 180 min, respectively. After light irradiation, three adults were selected from each group and immediately frozen in liquid nitrogen and stored at − 80°C until an analysis. Each treatment group was repeated in triplicate. The green LED lights (5 W; It is used in the light trap as a light source) used in the experiment were purchased from Shenzhen New Photoelectric Technology Co., Ltd. (China). The wavelength and irradiance of the green LED light were measured by a spectrometer (USB4H11915, Ocean Optics Co. Ltd., USA). The digital lux meter (GM1020-CH-00) used in the experiment was purchased from Shenzhen Jumaoyuan Science and Technology Co., Ltd. (China).
Sample preparation and measuring a protein concentration
Each adult moth was homogenized using a tissue grinder (JXFSTPRP-24; Shanghai Jingxin Industrial Development Co., Ltd. China) in 0.9% saline solution at a ratio of 0.9 ml of normal saline to 0.1 g body weight. The prepared homogenate was centrifuged at 2500 rpm for 10 min at 4 °C. The supernatant was collected to be used for subsequent analysis. The protein concentrations of all samples were measured by using an available BCA protein concentration assay kit (Beijing Dingguo Changsheng Biotechnology Co., Ltd, China).
Assay of total antioxidant capacity and antioxidant enzymes
The total antioxidant capacities (T-AOC) of samples were assayed according to the manufacturer’s protocol from an available assay kit (Nanjing Jiancheng Bioengineering Institute, China). The assay principle was based on the ability to reduce Fe3+ in a pool of antioxidant substances from a supernatant sample. In the assay, the antioxidant in a sample was used as a reductant in the redox-linked colorimetric reaction, reduced Fe2+ makes a relatively stable complex compound with porphyrin substance, which can measured at 520 nm. One unit of T-AOC was defined as the amount necessary to increase the absorbance by 0.01 per minute per mg protein at 37 °C. The T-AOC was noted as unit per milligram protein.
The activities of several antioxidant enzymes, including SOD, CAT, POD, and GST, were determined by using available assay kits (Nanjing Jiancheng Bioengineering Institute, China) based on the manufacturer’s protocols of these kits.
SOD activity was assayed spectrophotometrically at 550 nm by employing the reaction system of xanthine and xanthine oxidase. One unit of the SOD activity was stipulated as the amount of enzyme required to bring about a half inhibition of the xanthine and xanthine oxidase reaction system in 1 mg protein per ml enzyme extraction. The SOD activity was noted as unit per milligram protein.
CAT activity was resolved by measuring the decrease in the absorbance at 405 nm in response to the decomposition of H2O2. One unit of the CAT activity was defined as the amount of enzyme required for the composition of 1 μmol of H2O2 per second per mg protein. The CAT activity was expressed as unit per milligram protein.
POD activity was determined spectrophotometrically at 420 nm by catalyzing the oxidation in H2O2 presence of a substrate. One unit of POD activity was described as the amount that catalyzes 1 μg substrate per minute per mg protein under 37 °C condition. The POD activity was noted as unit per milligram protein.
GST activity was evaluated by spectrophotometric method at 412 nm based on the ability of chemical combination of glutathione (GSH) and 1-choro-2,4-dinitrobenzene (CDNB) by using CDNB as a substrate. One unit of the GST activity was characterized as the amount that catalyzes the chemical combination of GSH and CDNB per minute per mg protein under 37 °C condition. The GST activity was expressed as unit per milligram protein.
Statistical analysis
We performed one-way analysis of variance (ANOVA) with the software SPSS (SPSS Inc., Chicago, IL, USA) to compare the level of T-AOC and the activities of several antioxidant enzymes in same sexual insect bodies according to different light exposure times. Significant differences were analyzed using Tukey’s HSD test (P < 0.05). Significant differences between male and female bodies according to different light exposure times were analyzed by paired t test. Values of all graphs are expressed as the means and standard errors (mean ± SEM).
Results
Influence on T-AOC
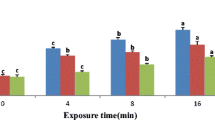
No significant changes in total antioxidant capacities of male moths and female moths were observed after exposure to green LED light irradiation for 30 min (Fig. 1). However, significant increases in males were recorded for exposure times of 60 and 120 min, but significant difference was not recorded for an exposure time of 180 min compared to the control (0 min) (F = 11.138, df = 4,10, p = 0.001). Meanwhile, total antioxidant capacities of females for an exposure time of 60 and 120 min were significantly higher compared with 180 min and the control (0 min), and total antioxidant capacities of females for an exposure time of 60 min was significantly higher than 120 min, but the 180-min group has no significant difference compared to the control (F = 17.853; df = 4.10; p = 0.000). Additionally, a significant difference between males and females occurred at 60 min (t = − 5.465; df = 2; p = 0.032) and 180 min (t = − 4.677; df = 2; p = 0.043) groups, respectively.

Influence of green LED light irradiation on T-AOC in Mythimna separata adults according to different exposure times. The value of each bar expresses a mean of the replicate experiments, and the error bar indicates the S.E. Different letters above each bar denote significant differences according to the Tukey HSD test (p < 0.05; capital letters and lowercase letters display significant differences in males and females, respectively). Different signs above each line express significant differences according to the paired t test (n.s., not significant; *p < 0.05)
Influence on SOD activity
When the males and females were exposed to green LED light, significant differences in SOD activities of males compared to the control (0 min) were not recorded for an exposure time of 0 to 120 min, but a significant decrease was recorded for 180 min compared with 30 and 60 min (F = 4.200; df = 4,10; p = 0.030) (Fig. 2). Additionally, a significant decrease in females was recorded for 180 min compared to 120 min and the control (0 min) (F = 5.823; df = 4,10; p = 0.011). Meanwhile, a significant difference between male and female moths was seen at 30 min (t = 4.603; df = 2; p = 0.044), 60 min (t = 8.127; df = 2; p = 0.015), and 180 min (t = 5.866; df = 2; p = 0.028), respectively.

Influence of green LED light irradiation on SOD activity in Mythimna separata adults according to different exposure times. The value of each bar expresses a mean of the replicate experiments, and the error bar indicates the S.E. Different letters above each bar denote significant differences according to the Tukey HSD test (p < 0.05; capital letters and lowercase letters display significant differences in males and females, respectively). Different signs above each line express significant differences according to the paired t test (n.s., not significant; *p < 0.05)
Influence on CAT activity
The CAT activities in males and females did not significantly change until an exposure time of 30 min, but significant increases compared with the control appeared at 60 and 120 min, respectively (Fig. 3). A significant decrease in males was recorded for 180 min compared with 60 and 120 min, but there was no a significant difference in contrast with the control (0 min) (F = 16.251; df = 4,10; p = 0.000). Additionally, a significant increase compared with the control among females appeared at 60 and 120 min, and there was no significant difference between 180 min and the control (0 min) (F = 9.153; df = 4,10; p = 0.002). Meanwhile, a marginal significant difference between males and females was recorded for 180 min (t = − 3.961; df = 2; p = 0.058).

Influence of green LED light irradiation on CAT activity in Mythimna separata adults according to different exposure times. The value of each bar expresses a mean of the replicate experiments, and the error bar indicates the S.E. Different letters above each bar denote significant differences according to the Tukey HSD test (p < 0.05; capital letters and lowercase letters display significant differences in males and females, respectively). Different signs above each line express significant differences according to the paired t test (n.s., not significant; #p < 0.1)
Influence on POD activity
No significant changes in POD activities of males and females were observed after exposure to green LED light irradiation for an exposure time of 30 min (Fig. 4). However, the POD activities in males were significantly increased at 60 and 120 min, and there was a significant decrease at 180 min compared with 60 and 120 min, but the 180-min group was not significantly changed compared to the control (0 min) (F = 17.531; df = 4,10; p = 0.000). A significant increase among females was observed for 60 min, and there were no significant difference among other exposure times (F = 6.979; df = 4.10; p = 0.006). Meanwhile, significant differences and marginal significant difference between males and females occurred at 30 min (t = 4.407; df = 2; p = 0.048), 120 min (t = 24.560; df = 2; p = 0.002), and 180 min (t = 4.185; df = 2; p = 0.053) groups, respectively.

Influence of green LED light irradiation on POD activity in Mythimna separata adults according to different exposure times. The value of each bar expresses a mean of the replicate experiments and the error bar indicates the S.E. Different letters above each bar denote significant differences according to the Tukey HSD test (p < 0.05; capital letters and lowercase letters display significant differences in males and females, respectively). Different signs above each line express significant differences according to the paired t test (n.s., not significant; #p < 0.1; *p < 0.05)
Influence on GST activity
The GST activity of the 30-min group in males was significantly lower than other exposure time groups and the control (0 min), and in addition to the 30-min group, the other groups had no significant change compared with the control (F = 7.037; df = 4,10; p = 0.006) (Fig. 5). Additionally, the 30- and 60-min groups in females were significantly decreased compared with the other groups and the control, and there was no significant change between the other groups compared with the control (F = 71.531; df = 4,10; p = 0.000). Meanwhile, significant differences between males and females were recorded for 30 min (t = 4.237; df = 2; p = 0.049), 60 min (t = 4.624; df = 2; p = 0.044), 120 min (t = 10.239; df = 2; p = 0.009), and 180 min (t = 6.964; df = 2; p = 0.020).

Influence of green LED light irradiation on GST activity in Mythimna separata adults according to different exposure times. The value of each bar expresses a mean of the replicate experiments, and the error bar indicates the S.E. Different letters above each bar denote significant differences according to the Tukey HSD test (p < 0.05; capital letters and lowercase letters display significant differences in males and females, respectively). Different signs above each line express significant differences according to the paired t test (n.s., not significant; *p < 0.05; **p < 0.01)
Discussion
Some studies have substantiated that green LED lights as light trap light source are useful to control pest moths in agriculture (Cho and Lee 2012; Yang et al. 2012; Shimoda and Honda 2013; Park and Lee 2016, 2017). However, it is not yet clear why many insects, including moths, fly unconsciously toward a light source of special wavelength, including a green light. UV light irradiation as an environmental stress induces the production of ROS in aerobic organisms, such as insects and animals ( Rebollar et al. 2006; Schauen et al. 2007; Yang et al. 2010; Gao et al. 2017). Previous studies have shown that UV light irradiation induces a relatively high oxidative stress in several insects, including M. separata (Ali et al. 2017), Helicoverpa armigera (Meng et al. 2009), Spodoptera litura (Karthi et al. 2014), Serica orientalis, and Anomala corpulenta (Gao et al. 2017). However, there have been few the studies on insect oxidative stress under monochromatic or visual wavelength light irradiation condition. The network of enzymatic antioxidant systems plays an important role in protecting cells from produced ROS in insect and animal bodies, along with non-enzymatic systems (Felton and Summers 1995; Ali et al. 2017). To evaluate the influence of green LED light irradiation on the oxidative stress in a phototactic moth, we measured T-AOC and the activities of the antioxidant enzymes, including SOD, CAT, POD, and GST in M. separata adult moths under green light irradiation.
T-AOC is known as a widely used tool for assessing redox and an index representing a total antioxidant capacity on an organism (Ghiselli et al. 2000; Sashidhara et al. 2011). The present study confirmed that the exposure of M. separata adult moths to green LED light for 60 min significantly increased T-AOC compared to the control (0 min), long exposure times from 60 to 180 min gradually decreased, but T-AOC values of the control and 180 min were not a significantly different. Additionally, T-AOC values in males and females revealed significant differences at 60 and 180 min, respectively. It suggests that T-AOC in the moths adapts to eliminate an oxidative stress due to free radical formation under green light irradiation condition, and T-AOC levels in male and female under visible light irradiation are different.
A significant increase in the activity of antioxidant enzymes, including SOD, CAT, POD, and GST, indicates formation of high amount of ROS in an organism; it means a sign of oxidative stress (Foyer et al. 1994). Whereas, if these activities of four enzymes are significantly reduced in comparison with a control (in normal condition) under continuous oxidative stress conditions, it means a damaging stage of cells due to high amount of ROS (Halliwell and Gutteridge 1999; Ali et al. 2016b). Some studies have showed that the activities of these antioxidant enzymes in some moths under UV light irradiation are significantly raised up to a certain exposure time, but these activities for an increasing exposure time are sharply reduced compared with a control (normal stage) (Meng et al. 2009; Karthi et al. 2014; Ali et al. 2017). Additionally, a proteomic and molecular analysis of H. armigera adult moths after exposure to UV light suggested that UV light induces an oxidative stress in H. armigera adult moths (Meng et al. 2010; Wang et al. 2012).
A main function of SOD is to catalyze the dismutation of superoxide radicals produced by oxidative stress into H2O2 (Ahmad et al. 1991). Based on present research, an activity of SOD in M. separata male moths exposed to green light irradiation did not reveal any significant difference compared with the control, but a significant decrease in females was recorded for 180 min. However, a numerical increase of the activity of SOD in males at 30 and 60 min, and in females at 120 min, revealed an increasing production of superoxide radicals.
CAT as a light-sensitive antioxidant enzyme plays an important role in catalyzing the dismutation of H2O2 into water and oxygen (Lesser 2006). Some studies have shown that an expression of CAT gene is associated with longevity of insects, a level of CAT in insects may express oxidative stress degree (Orr and Sohal 1994; Karthi et al. 2014). In the present research, an activity of CAT in the male and female moths exposed to green light irradiation for 60 and 120 min was significantly increased compared to the control; it suggests that a high level of CAT in exposed moths is due to protect DNA from oxidative stress. Meanwhile, CAT based on the present result was lower level than SOD level, reflecting that except for CAT, other enzymes, such as POD, may participate in the eliminating process of H2O2.
An activity level of POD is associated with the increase in stress tolerance (Mathews et al. 1997). In our results, significant increases of an activity level of POD recorded for 60 and 120 min in male and for 60 min in female, and the changing tendency in the activity levels of POD according to increasing exposure time was similar to that of CAT. We thus suspected that the high level of POD in the moths exposed to green light serves as a protective mechanism against DNA damage, and POD works in cooperating with CAT to scavenge H2O2.
GST is a crucial enzyme in the insect antioxidant system, and it can metabolize lipid peroxidation products together with POD, which also breaks down H2O2 (Jia et al. 2011). In our results, GST activity significantly decreased at 30 min in male and at 30 to120 min in female. We supposed that a lipid peroxidation by green light irradiation already occurred before the exposure time of 30 min; it suggests that green light may result in a lipid peroxidation in the moths.
Ali et al. (2017) has reported that when M. separata moths are exposed to UV light irradiation for prolonged exposure time (120 min), T-AOC, SOD, CAT, POD, and GST in the moths significantly decrease compared with the control (0 min). The two previous studies also found the similar results in H. armigera adult moths (Meng et al. 2009) and S. litura adult moths (Karthi et al. 2014). These findings suggest that UV light irradiation for a long time may damage cell in the moth bodies to cause cell deaths. However, our finding showed that except for SOD in female, the activity levels of these antioxidant enzymes at the longest exposure time (180 min) were not significantly different compared to the control, suggesting green light irradiation induces the oxidative stress in M. separata moths, but the moths may restore the ability to manage the oxidative stress and free radicals associated with this irradiation. In other words, the oxidative stress degree of the green light irradiation in the moths is lower than that of UV light irradiation. Comorosan et al. (2009) has suggested that a green light causes a protective effect on DNA denatured by UV light at a cellular level. Zhang et al. (2017) has proposed that the artificial boosts of ROS can extend the life span of the H. armigera pupae by using components of insulin-signaling pathway. When different developmental stages of Drosophila melanogaster were exposed to light irradiation of different wavelengths, green light irradiation resulted in high survival rate compared with the control (Hori et al. 2014).
In our previous study, we found that the green (520 nm) LED light results in a higher phototactic response more than UV light in M. separata moths (not published; Kim et al.), which may be due to the possible reason that green light irradiation induces a weaker oxidative stress compared to UV light irradiation in the moths. Similarly, a previous study has reported that trapping effect of a light trap equipped with a light source that induces a weaker light stress is superior to that of a light trap that induces a stronger light stress in cockchafers (Gao et al. 2017).
Additionally, significant differences of the activities of these antioxidant enzymes between the male and the female moths according to an exposure time were revealed, suggesting that the metabolizing ability of antioxidant system varies according to the sex of moths; thereby, the photosensitivities of male and female moths are different from each other. However, the above-mentioned findings still require more scientific verifications.
Conclusions
The present research demonstrated that green (520 nm) LED light irradiation could induce a weak oxidative stress, and the moths under the green light could restore the damage caused by this light stress. Additionally, we suggested that the adaptive ability to the oxidative stress varied between the moths of different sexes. Our results may provide a theoretical and scientific basis for elucidating a reason of the phototactic behavior of nocturnal moths, including M. separata.
References
Ahmad S (1992) Biochemical defence of pro-oxidant plant allelochemicals by herbivorous insects. Biochem Syst Ecol 20(4):269–296. https://doi.org/10.1016/0305-1978(92)90040-K
Ahmad S, Duval DL, Weinhold LC, Pardini RS (1991) Cabbage looper antioxidant enzymes: tissue specificity. Insect Biochem 21(5):563–572
Ali A, Rashid M, Huang Q, Lei C (2016a) Effect of UV-A radiation as an environmental stress on the development, longevity, and reproduction of the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae). Environ Sci Pollut Res 23(17):17002–17007. https://doi.org/10.1007/s11356-016-6865-0
Ali A, Rashid MA, Huang QY, Wong C, Lei CL (2016b) Response of antioxidant enzymes in Mythimna separata (Lepidoptera: Noctuidae) exposed to thermal stress. B Entomol Res 107(3):382–390. https://doi.org/10.1017/S0007485316001000
Ali A, Rashid MA, Huang QY, Lei CL (2017) Influence of UV-A radiation on oxidative stress and antioxidant enzymes in Mythimna separata (Lepidoptera: Noctuidae). Environ Sci Pollut Res 24(9):8392–8398. https://doi.org/10.1007/s11356-017-8514-7
Barbehenn RV, Bumgarner SL, Roosen EF, Martin MM (2001) Antioxidant defenses in caterpillars: role of the ascorbate-recycling system in the midgut lumen. J Insect Physiol 47:349–357. https://doi.org/10.1016/S0022-1910(00)00125-6
Be FC (1981) New artificial diet for the army worm Leucania separata Walker. Acta Entomol Sin 24(4):379–382 (in Chinese)
Cho KS, Lee HS (2012) Visual preference of diamondback moth, Plutella xylostella, to light-emitting diodes. J Korean Soc Appl Biol Chem 55:681–684. https://doi.org/10.1007/s13765-012-2116-3
Comorosan S, Kappel W, Constantinescu I, Gheorghe M, Ionescu E, Pîrvu C, Cinca S, Cristache L (2009) Green light effects on biological systems: a new biophysical phenomenon. J Biol Phys 35(3):265–277. https://doi.org/10.1007/s10867-009-9164-x
Davies KJ (2000) Oxidative stress, antioxidant defenses, and damage removal, repair, and replacement systems. IUBMB Life 50:279–289. https://doi.org/10.1080/15216540051081010
Felton GW, Summers CB (1995) Antioxidant systems in insects. Arch Insect Biochem Physiol 29:187–197. https://doi.org/10.1002/arch.940290208
Foyer CH, Descourvières P, Kunert KJ (1994) Protection against oxygen radicals: an important defence mechanism studied in transgenic plants. Plant Cell Environ 17(5):507–523
Gao Y, Li G, Li K, Lei C, Huang Q (2017) Comparison of the trapping effect and antioxidant enzymatic activities using three different light sources in cockchafers. Environ Sci Pollut Res 24(36):27855–27861. https://doi.org/10.1007/s11356-017-0388-1
Ghiselli A, Serafini M, Natella F, Scaccini C (2000) Total antioxidant capacity as a tool to assess redox status: critical view and experimental data. Free Radic Biol Med 29(11):1106–1114. https://doi.org/10.1016/S0891-5849(00)00394-4
Halliwell B, Gutteridge JMC (1999) Free radicals in biology and medicine. Oxford University Press, Oxford
Hori M, Shibuya K, Sato M, Saito Y (2014) Lethal effects of short-wavelength visible light on insects. Sci Rep 4(7383):1–6. https://doi.org/10.1038/srep07383
Huang J, Hua HQ, Zhang F, Li YX (2017) Suitability assessment of three Trichogramma species in the control of Mythimna separata (Lepidoptera: Noctuidae). J Appl Entomol 142:131–140. https://doi.org/10.1111/jen.12414
Jia FX, Dou W, Hu F, Wang JJ (2011) Effects of thermal stress on lipid peroxidation and antioxidant enzyme activities of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Fla Entomol 94(4):956–963
Joanisse DR, Storey KB (1998) Oxidative stress and antioxidants in stress and recovery of cold-hardy insects. Insect Biochem Mol Biol 28(1):23–30. https://doi.org/10.1016/S0965-1748(97)00070-2
Kamata H, Hirata H (1999) Redox regulation of cellular signalling. Cell Signal 11(1):1–14. https://doi.org/10.1016/S0898-6568(98)00037-0
Karthi S, Sankari R, Shivakumar MS (2014) Ultraviolet-B light induced oxidative stress: effects on antioxidant response of Spodoptera litura. J Photochem Photobiol B 135:1–6. https://doi.org/10.1016/j.jphotobiol.2014.04.008
Lalouette L, Williams CM, Hervant F, Sinclair BJ, Renault D (2011) Metabolic rate and oxidative stress in insects exposed to low temperature thermal fluctuations. Comp Biochem Physiol A Physiol 158:229–234. https://doi.org/10.1016/j.cbpa.2010.11.007
Lesser MP (2006) Oxidative stress in marine environments: biochemistry and physiological ecology. Annu Rev Physiol 68:253–278. https://doi.org/10.1146/annurev.physiol.68.040104.110001
Li M, Liu XM, Wang Y, Wang XP, Zhou LL, Si SY (2015) Effect of relative humidity on entomopathogens infection and antioxidant responses of the beet armyworm, Spodoptera exigua (Hübner). Afr Entomol 22(3):651–659
Lihuang K, Zhang Z, Kim K, Huang Q, Lei C (2017) Antennal and behavioral responses of Mythimna separata (Walker) to three plant volatiles. Environ Sci Pollut Res 24:24953–24964. https://doi.org/10.1007/s11356-017-0140-x
Mathews MC, Summers CB, Felton GW (1997) Ascorbate peroxidase: a novel antioxidant enzyme in insects. Arch Insect Biochem Physiol 34(1):57–68
Meng JY, Zhang CY, Zhu F, Wang XP, Lei CL (2009) Ultraviolet light-induced oxidative stress: effects on antioxidant response of Helicoverpa armigera adults. J Insect Physiol 55(6):588–592. https://doi.org/10.1016/j.jinsphys.2009.03.003
Meng JY, Zhang CY, Lei CL (2010) A proteomic analysis of Helicoverpa armigera adults after exposure to UV light irradiation. J Insect Physiol 56(4):405–411. https://doi.org/10.1016/j.jinsphys.2009.11.015
Orr WC, Sohal RS (1994) Extension of life-span by overexpression of superoxide dismutase and catalase in Drosophila melanogaster. Science 263(5150):1128–1130. https://doi.org/10.1126/science.8108730
Park JH, Lee HS (2016) Phototactic behavior 10: phototactic behavioral effects of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) adults to different light-emitting diodes of seven wavelengths. J Appl Biol Chem 59(2):95–98. https://doi.org/10.3839/jabc.2016.017
Park JH, Lee HS (2017) Phototactic behavioral response of agricultural insects and stored-product insects to light-emitting diodes (LEDs). Appl Biol Chem 60(2):137–144. https://doi.org/10.1007/s13765-017-0263-2
Rebollar E, Valadez-Graham V, Vázquez M, Reynaud E, Zurita M (2006) Role of the p53 homologue from Drosophila melanogaster in the maintenance of histone H3 acetylation and response to UV-light irradiation. FEBS Lett 580(2):642–648. https://doi.org/10.1016/j.febslet.2005.12.083
Sashidhara KV, Singh SP, Srivastava A, Puri A (2011) Identification of the antioxidant principles of Polyalthia longifolia var. pendula using TEAC assay. Nat Prod Res 25(9):918–926
Schauen M, Hornig-Do HT, Schomberg S, Herrmann G, Wiesner RJ (2007) Mitochondrial electron transport chain activity is not involved in ultraviolet A (UVA)-induced cell death. Free Radic Biol Med 42(4):499–509. https://doi.org/10.1016/j.freeradbiomed.2006.11.016
Sharma HC, Davies JC (1983) The oriental armyworm, Mythimna separata (Wal.) distribution, biology and control: a literature review. Miscellaneous Rep 59:1–24
Sharma HC, Sullivan DJ, Bhatnagar VS (2002) Population dynamics and natural mortality factors of the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae), in South-Central India. Crop Prot 2(9):721–732
Shimoda M, Honda KI (2013) Insect reactions to light and its applications to pest management. Appl Entomol Zool 48:413–421. https://doi.org/10.1007/s13355-013-0219-x
Wang Y, Oberley LW, Murhammer DW (2001) Antioxidant defense systems of two lipidopteran insect cell lines. Free Radic Biol Med 30(11):1254–1262. https://doi.org/10.1016/S0891-5849(01)00520-2
Wang GP, Zhang QW, Ye ZH, Luo LZ (2006) The role of nectar plants in severe outbreaks of armyworm Mythimna separata (Lepidoptera: Noctuidae) in China. Bull Entomol Res 96(5):445–455
Wang Y, Wang L, Zhu Z, Ma W, Lei C (2012) The molecular characterization of antioxidant enzyme genes in Helicoverpa armigera adults and their involvement in response to ultraviolet-A stress. J Insect Physiol 58(9):1250–1258. https://doi.org/10.1016/j.jinsphys.2012.06.012
Yang C, Gong G, Tan F, You Z (1995) Preliminary studies on monitoring and mechanisms of the insecticide resistance in Mythimna separata. Plant Prot 21(3):2–5 (in Chinese)
Yang LH, Hai H, Wang JJ (2010) Antioxidant responses of citrus red mite, Panonychus citri (McGregor) (Acari: Tetranychidae), exposed to thermal stress. J Insect Physiol 56(12):1871–1876. https://doi.org/10.1016/j.jinsphys.2010.08.006
Yang JY, Kim MG, Lee HS (2012) Phototactic behavior: attractive effects of Spodoptera litura (Lepidoptera: Noctuidae), tobacco cutworm, to high-power light-emitting diodes. J Korean Soc Appl Biol Chem 55(6):809–811. https://doi.org/10.1007/s13765-012-2172-8
Zha YP, Lei CL (2012) Effects of ultrasound-stress on antioxidant enzyme activities of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). J Agric Urban Entomol 28(1):34–41
Zhang XS, Wang T, Lin XW, David LD, Xu WH (2017) Reactive oxygen species extend insect life span usingcomponents of the insulin-signaling pathway. Proc Natl Acad Sci U S A 114(37):7832–7840. https://doi.org/10.1073/pnas.1711042114
Zhao Z, Sandhu HS, Fang O, Feng G (2016) Landscape changes have greater effects than climate changes on six insect pests in China. Sci China Life Sci 59(6):627–633. https://doi.org/10.1007/s11427-015-4918-0
Funding
This study was supported by the National Key Research and Development Program of China (2017YFD0200900) and the National Department Public Benefit (Agriculture) Research Foundation (201403031).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article

Cite this article
Kim, KN., Yun, CN., Sin, UC. et al. Green light and light stress in moth: influence on antioxidant enzymes in the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae). Environ Sci Pollut Res 25, 35176–35183 (2018). https://doi.org/10.1007/s11356-018-3415-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-3415-y