
Abstract
The ultraviolet light (UV-A) range of 320–400 nm is widely used as light trap for insect pests. Present investigation was aimed to determine the effect of UV light-A radiation on development, adult longevity, reproduction, and development of F1 generation of Mythimna separata. Our results revealed that the mortality of the second instar larvae was higher than the third and fourth instar larvae after UV-A radiation. As the time of UV-A irradiation for pupae prolonged, the rate of adult emergence reduced. Along with the extension of radiation time decreased the longevity of adult females and males. However, the radiation exposure of 1 and 4 h/day increased fecundity of female adults, and a significant difference was observed in a 1 h/day group. The oviposition rates of female adults in all the treatments were significantly higher than the control. In addition, UV-A radiation treatments resulted in declined cumulative survival of F1 immature stages (eggs, larvae, and pupae). After exposure time of 4 and 7 h/day, the developmental periods of F1 larvae increased significantly, but no significant effects on F1 pupal period were recorded.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Climate changes have versatile influences on consequences of abiotic stress, threatening the sustainability and productivity of agricultural systems. Abiotic and biotic stress factors have crucial role in insect life. There are numerous environmental stresses, e.g., depletion in ozone, increase in ultraviolet irradiation, loss of biodiversity and temperature, etc. UV radiation is a major environmental stress factor for many organisms (Meyer-Rochow 2000; Schauen et al. 2007). Currently, UV radiation is expecting a lot of considerations due to its possibly injurious effects on several organisms. UV proliferation can decline adult longevity, enhance oviposition rate, and reduce fecundity and fertility (Faruki et al. 2005; Zhang et al. 2011). UV stimulates ROS, which can spoil nucleic acid, membrane lipids, and proteins (Vile and Tyrrell 1995). These varieties of damages in above molecules cause substantial effects on biology such as genetic variation, cytotoxicity, and transformation in signaling cell pathways (McMillan et al. 2008).
The oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae), is an important migratory pest in eastern Asia. It has caused damage to wheat, corn, and rice crops since 1970 in China (Yang et al. 1984). Its seasonal migration has been studied in China (Chen and Bao 1987; Chen et al. 1995). During winter, this migratory pest settled in southwestern China and flies to northern China to damage summer crops. Recently, the invasion of armyworm has caused damage to crops; especially corn was damaged in neighboring Liaoning, Hebei, and the Inner Mongolia province (Ye 2012). The moths of nocturnal insects are frequently drawing intention to UV light due to their innate behavior which can be a base for their control (Wei et al. 2000; Emura and Tazawa 2004). UV radiation disturbs an insect’s fitness by limiting its ability to find a native mate, its reproduction by affecting its longevity, or disturbing the harmony of its circadian rhythms (Barlett et al. 1968; North and Holt 1968; Proverbs 1969; Van Steenyk et al. 1979; LaChance et al. 1975; Tsubaki and Bunroongsook 1990; Hasan 1995). UV light broadly used as integrated pest management (IPM) to control different kinds of insect pest populations such as lepidopteron (Antignus 2000; Steinbauer 2003; Kojima et al. 2005). UV radiation directly affects the insect behavior, developmental physiology, and biochemistry (Gunn 1998; Mackerness et al. 1999; Mazza et al. 2002). Various studied discovered that immature developmental stages are more susceptible to indirect and direct UV radiation than mature insects (Watters 1976; Tilton and Brower 1983; Shayesteh and Barthakur 1996; Zhao et al. 2007; Zhang et al. 2011), and it is also reported that UV radiation effect on living organisms is associated to cellular development. Low intensity of UV-A radiation increased the lifespan of Drosophila, and the high intensity of UV-A radiation can lead to prolonged life expectancy shortened (Zheng 2011). In the present study, we examined UV-A irradiation effect on survival, reproduction, different development stages, and as well as F1 generation of M. separata. The purpose of the present study is to estimate the effect of UV-A radiation on physiology of M. separata.
Materials and methods
Experimental insects
M. separata were reared at the room temperature 26 ± 2 °C, 60 ± 10 % RH, and 14:10 h L/D. An artificial diet was used to feed the larvae as mentioned in Chun (1981).
Larvae and pupae
The second, third, and fourth instar larvae and 48-h-old pupae were collected from the food medium kept in separate petri dishes. The UV-A radiation was given to larvae and pupae 1 h after the start of scotophase. The irradiation dose was approximately at 350 W/cm2. Larvae and pupae were exposed to UV-A for 0, 4, 8, and 16 min. The larval mortality was assessed up to pupation. There were 50 larvae and 30 pupae per treatment, with three replications. Irradiated pupae were kept in plastic containers for adult emergence.
Adult longevity and reproduction
UV light-A (X-series, peak emission 365 nm; Spectronics, Westbury, NY) were used in the experiment. Male and female adults were exposed to UV-A for the different period of times, i.e., 0 (control), 1, 4, and 7 h (hours) per day, until the death of males and females. Ten female–male pairs were used in each treatment with three replications of UV-A radiation. Each female–male pair was reared in a single plastic cup (10 cm 7 diameter) was tested. The plastic cups were closed with cotton net yarn and kept butter paper in glass for egg laying. Cotton net yarn and batter paper were replaced daily. The fecundity, ovipostion rate, and survival of male and female were recorded on the daily basis. The experiment was terminated until the males and females died.
Development of the F1 generation
Eggs laid by adults of the P generation were referred to as the F1 generation. Cotton yarns with eggs were sprinkled with water and then transferred to a separate plastic petri dish (7-cm diameter). One hundred fresh eggs were sampled randomly and incubated into three replications (per treatment 300 eggs) in plastic petri dishes. To estimate the hatching percentage of eggs, the newly hatched larvae were noted daily. A plastic container (3 × 3 × 4 cm) was used to rear 60 newly hatched larvae individually in three replications per treatment (total 180 larvae) to observe the larval period. The number of pupa and the timing of pupation were recorded, and 40 newly obtained pupae were transferred into new separate plastic containers with three replications per treatment (total 120 pupae). The number of the adult and timing of the emergence was logged. The experiments were performed at 26 ± 2 °C, 60 ± 10 % RH, and 14:10 h L/D photoperiod.
Statistical analysis
The software SPSS (SPSS Inc., Chicago, IL, USA) was used for data analysis. The one-way analysis of variance (ANOVA) was used for analysis of data, and means were compared Tukey HSD test at P < 0.05. Data of egg hatch, survival of larvae, and pupae were arcsine. Square-root transformed prior to analysis.
Results
Effect of UV-A radiation on larval mortality and adult emergence
The larval mortalities of second, third, and fourth instars gradually increased as UV-A radiation exposure time increased. The larval mortality of second and third instars significantly increased (F = 132.85, P < 0.001, df = 3.8; F = 45.60, P < 0.001, df = 3.8), respectively, at the exposure period of 4, 8, and 16 min UV light. It was observed that third instar larval mortality was significantly higher than fourth instar larvae (F = 32.66, P < 0.001, df = 3.8). A high mortality rate was observed in fourth instar larvae at the longest exposure time of 16 min. No significant deference was observed between control and 4-min UV irradiation. We found that earlier larval stage (second instars) were more sensitive to UV at any level of mortality counted than third and fourth instar larvae (Fig. 1).

UV-A radiation effect on different instars of Mythimna separata larvae. Values (mean ± SEM) followed by different letters within a column are significantly different by Tukey HSD test (P < 0.05)
Adult emergence of UV-A-exposed M. separata pupae significantly reduced at 8 and 16 min (F = 30.364 P < 0.001, df = 3.8). There was no significant difference between control and 4-min UV light exposure (Fig. 2).

Adult emergences (%) of Mythimna separata from UV-irradiated pupae. Values (mean ± SEM) followed by different letters within a column are significantly different by Tukey HSD test (P < 0.05)
Effect of UV-A radiation on adult longevity
A significant decrease in female longevity was observed when female adults were exposed to UV-A radiation for 1, 4, and 7 h/day (F = 50.261, P < 0.001, df = 3.8). There was a significant decrease in male longevity found when male adults were expose to UV-A radiation for 7 h/day (F = 6.025, P < 0.01, df = 3.8), while no significant difference was seen in male longevity between 1 and 4 h/day UV radiation as compared to control group (Fig. 3a, b).

Effect of UV-A radiation on adult longevity of Mythimna separata, for a females, b males. Error bars indicate SEM. Bars with different letters are significantly different by Tukey HSD test (P < 0.05)
Effect of UV-A radiation on reproduction
UV-A radiation effect on reproduction of M. separata from the exposure of 1 and 4 h/day resulted in a fecundity increase as compared to control. We found a significant difference after the exposure of 1 h/day (F = 6.208, P < 0.017, df = 3.8). Females laid fewer eggs compared to control in the oviposition period, but statistically, the difference was not significant when they were exposed to UV-A radiation for 7 h/day. Oviposition rate was significantly affected by UV-A radiation (F = 11.089, P < 0.003, df = 3.8). The rate of oviposition was increased as the exposure time increased, and it reached to a maximum at 7 h/day as compared to all other groups (Table 1).
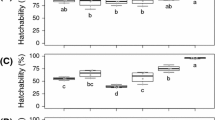
Effect of the UV-A radiation on development of the F1 generation
Data on the effect of UV-A radiation indicated a significant decrease on 7 h/day in F1 eggs (F = 6.606, P < 0.01, df = 3.8), and it was not significantly different for the survival of F1 larval (F = 2.528, P = 0.13, df = 3.8). A significant decrease was observed on 4 h/day in F1 pupal survival (F = 10.377, P < 0.004, df = 3.8). UV-A radiation exposure resulted a significant reduction in cumulative survival as compared to the control group for a period of 1, 4, and 7 h/day (Table 2).
The larval and pupal developmental periods of F1 M. separata was also affected by UV-A radiation. It was found that F1 larval developmental period increased in contrast to control, as the exposure of UV-A radiation was given to adults. At the duration of 4 and 7 h/day, a significant (F = 39.787, P < 0.001, df = 3.8) interruption was seen in the development of F1 larvae. The larvae of F1 took longest time to attain the pupal stage for the longest UV-A exposure time (7 h/day). There was no significant (F = 0.854, P = 0.50, df = 3.8) effect seen on the developmental period of F1 pupae. The emergence time for the adult stage for both F1 pupae in the control and all the treated groups were same (Table 3).
Discussion
UV-A radiation has a strong capability to affect the physiology of insects. Our results showed that UV-A radiation has more harmful effect on younger larvae as compared to older larvae. Similar results were also reported by (Qureshi et al. 1970; Faruki et al. 2005). Mortality of the irradiated larvae was directly related to the dose of radiation, i.e., mortality gradually increased as the dose of radiation increased. Recently, it is found that larval mortality was completely associated with radiation doses (Pandır and Guven 2014; Guven et al. 2015). Insect mortality ratio enhances with increasing UV irradiation time. It may be due to the protein and fat level in the insect body. These molecules are less in earlier larval instar and higher in later instar. These molecules may have the role in immune defense system, so they hinder the UV light effect in later stages. Therefore, mortality is higher in earlier larval instar as compared to later instars. Adult emergence of UV-exposed M. separata pupae decreased as the time of irradiation increased. A significant reduction in adult emergence from irradiated pupae was observed (Hasan et al. 1998; Faruki et al. 2007). In our results, longer exposure time highly affected pupal stage. The possible reason is that pupal stage is static and UV irradiation directly enters and disturbs key physiologically processes (Tilton and Brower 1983).
Meng et al. (2009) reported that ultraviolet light could induce oxidative stress in adult insects. These radiations can upset the working capacity of proteins and augment the resultant product of protein oxidation in adult of M. separata. These processes decreased longevity and enhanced the oviposition rate (Zhang et al. 2011). Similar results were found in the present study in which the adult longevity was shortened, and rate of oviposition was elevated in M. separata. UV-A radiation extremely affected the biological parameters of P generation. Fecundity was increased after 1 and 4 h/day UV-A radiation as compared to control groups. But, significant difference was noted after 1 h/day groups. Fecundity was lesser than control at longer exposure time 7 h/day, but there was no significant difference. The oviposition rate of female adult in all UV-A radiation treatment groups were significantly increased as compared to control groups. After UV-A radiation, why fecundity and oviposition rate was increased remains unknown. Reason could be as the theoretically life history forecasts that some traits may exchange one another, particularly when each trait is vigorously expensive (Zera and Harshman 2001; Zera and Denno 1997). In common life history theory, it is a crucial prophecy that reproduction sustains a survival cost (Williams 1966; Roff 1992). Consequently, we presume that there is an exchange in energy sharing between the longevity and reproduction of M. separata adults under UV stress. Previously, it was known in lepidopteron moths that they have the ability to enhance their antioxidant defense system in response to toxic effects of ultraviolet radiation (Meng et al. 2009). It seems that this reaction is possibly expensive for the organism in terms of energy. In response to UV-A exposure, reproductive costs and antioxidant defenses are developed, so life history theory forecasts that existence capability must be faded to a certain limit (Holloway et al. 1990; de Jong and van Noordwijk 1992), and it was seen in the reduction of longevity in M. separata adult. In our experimental condition, UV-A radiation on hatched eggs and pupae survival of F1 generation significantly differed while there was no significant difference observed in larval survival. Present findings indicate that cumulative survival of F1 generation control was comparatively higher than UV-A radiation-treated groups. We observed that UV radiation disturbs the F1 generation, and longer time UV radiation can reduce the cumulative survival of F1 immature stages. The results revealed that significant difference was not found in developmental periods of F1 pupae, while significant change was seen in the developmental periods of F1 larvae between UV-A exposure and control insects. F1 larvae of UV-A-treated adults took longer time to become pupae. This result revealed that UV-A radiation has harmful effects on the larval development period of F1 generation. UV radiation can be used as an ecological stress factor in insects Meng et al. (2009, 2010). Many insects have the ability to prolong their developmental stages to prevent themselves from adverse effects of environmental conditions for their survival, which is the strategy of organisms to compensate themselves according to the environmental conditions for the protection of harmful effect on growth (Gintenreiter et al. 1993; van Ooik et al. 2007).
Conclusion
The present investigation revealed that UV-A radiation had high effect on survival, reproduction, different development stages, and F1 generation of M. separata. Earlier instar larvae of M. separata are more sensitive to UV radiation than later instar larvae. Adult emergence of UV-A-exposed M. separata pupae reduced as the time of irradiation increased. UV radiation reduces adult longevity and increases fecundity and oviposition rate of M. separata. It also affected the developmental stage of M. separata F1 generation.
References
Antignus Y (2000) Manipulation of wavelength-dependent behaviour of insects: an IPM tool to impede insects and restrict epidemics of insect-borne viruses. Virus Res 71:213–220
Barlett AC, Hooker PA, Hardee DD (1968) Behaviour of irradiated boll weevils. I. Feeding, attraction, mating and mortality. J Econ Entomol 61:1677–1680
Chen RL, Bao XZ (1987) Research on the migration of oriental armyworm in China and a discussion of management strategy. Insect Sci Appl 8:571–572
Chen RL, Sun YJ, Wang SY, Zhai BP, Bao XZ (1995) Migration of the oriental armyworm Mythimna separata in East Aisa in relation to weather and climate. I. Northeastern China. In: Drake VA, Gatehouse AG (eds) Insect migration: tracking resource in space and time. Cambridge University Press, Cambridge, pp 93–104
Chun BF (1981) A new artificial diet for army worm. Acta Entomol Sin 24:379–383
De Jong G, Van Noordwijk AJ (1992) Acquisition and allocation of resources: genetic (co) variances, selection, and life histories. Am Nat 139:749–770
Emura K, Tazawa S (2004) The development of the eco-engineering insect control technology-physical control of insect behavior using artificial lights. Eco-engineering 16:237
Faruki SI, Das DR, Khatun S (2005) Effects of UV-radiation on the larvae of the lesser mealworm, Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae) and their progeny. J Biol Sci 5:444–448
Faruki SI, Das DR, Khan AR, Khatu M (2007) Effects of ultraviolet (254nm) irradiation on egg hatching and adult emergence of the flour beetles Tribolium castaneum, T confusum and the almond moth Cadra cautella. J Insect Sci 7:1–6
Gintenreiter S, Ortel J, Nopp HJ (1993) Effects of different dietary levels of cadmium, lead copper, and zinc on the vitality of the forest pest insect Lymantria disparL. (Lymantriidae, Lepid). Arch Environ Contam Toxicol 25:62–66
Gunn A (1998) The determination of larval phase coloration in the African armyworm Spodoptera exempta and its consequences for thermoregulation and protection from UV light. Entomol Exp Appl 86:125–133
Guven E, Pandir D, Bas H (2015) UV radiation-induced oxidative stress and DNA damage on Mediterranean flour moth, Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) larvae. Turk J Entomol 39:23–33
Hasan M (1995) Comparative sensitivity of Tribolium spp. To gamma irradiation throughout ontogeny. PhD Thesis, Newcastle Univ, UK, pp-261
Hasan M, Jahan MS, Khan AR (1998) Effect of ultraviolet irradiation on the Uzi-fly, Exorista sorbillans Wiedmann, an endoparasitoid of the silkworm, L. Int J Trop Insect Sci 18:87–91
Holloway GJ, Sibly RM, Povey SR (1990) Evolution in toxin-stressed evironments. Funct Ecol 4:289–294
Kojima Y, Aoyagi K, Yasue T (2005) Effect of lithium ion addition on afterglow time of green-emitting Ce3 + and Pr3 + codoped CaS phosphor by black light irradiation. J Lumin 115:13–18
LaChance LE, Richard RD, Proshold FI (1975) Radiation response in pink boll worm: a comparative study of sperm bundle production, Sperm transfer, and oviposition response elicited by native and laboratory reared males. Environ Entomol 4:321–324
Mackerness SAH, Surplus SL, Blake P, John CF, Buchanan-Wollaston V, Jordan BR, Thomas B (1999) Ultraviolet-B-induced stress and changes in gene expression in Arabidopsis thaliana: role of signalling pathways controlled by jasmonic acid, ethylene and reactive oxygen species. Plant Cell Environ 22:1413–1423
Mazza CA, Izaguirre MM, Zavala J, Scopel AL, Ballare CL (2002) Insect perception of ambient ultraviolet-B radiation. Ecol Lett 5:722–726
McMillan TJ, Leatherman E, Ridley A, Shorrocks J, Tobi SE, Whiteside JR (2008) Cellular effects of long wavelength UV light (UVA) in mammalian cells. J Pharm Pharmacol 60:969–976
Meng JY, Zhang CY, Zhu F, Wang XP, Lei CL (2009) Ultraviolet light-induced oxidative stress: effects on antioxidant response of Helicoverpa armigera adults. J Insect Physiol 55:588–592
Meng JY, Zhang CY, Lei CL (2010) A proteomic analysis of Helicoverpa armigera adults after exposure to UV light irradiation. J Insect Physiol 56:405–411
Meyer-Rochow VB (2000) Risks, especially for the eye, emanating from the rise of solar UV-radiation in the Arctic and Antarctic regions. Int J Circ Health 59:38–51
North DT, Holt GG (1968) Inherited sterility in Progeny of irradiated male cabbage loopers. J Econ Entomol 61:928–931
Pandır D, Guven E (2014) Effect of microwave radiation on stored product pest Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) larvae. Turk J Entomol 38(2):135–142
Proverbs MD (1969) Sterilization and insect control. Annu Rev Entomol 14:81–102
Qureshi ZA, Wilbur DA, Mills RB (1970) Irradiation of early instar of the Angoumois moth. J Econ Entomol 63:1241–1247
Roff DA (1992) In: Roff DA (ed) The evolution of life histories: theory and analysis. Chapman & Hall, New York, pp 384–385
Schauen M, Hornig-Do HT, Schomberg S, Herrmann G, Wiesner RJ (2007) Mitochondrial electron transport chain activity is not involved in ultraviolet A (UVA)-induced cell death. Free Radic Biol Med 42:499–509
Shayesteh N, Barthakur NN (1996) Mortality and behavior of two stored-product insect species during microwave irradiation. J Stored Prod Res 32:239–246
Steinbauer MJ (2003) Using ultra-violet light traps to monitor autumn gum moth, Mnesampela private (Lepidoptera: Geometridae), in south-east Australia. Aust For 66:279–286
Tilton EW, Brower JH (1983) Radiation effects of on arthropods. In: Josephson ES, Peterson MS, Boca R (eds) Preservation of food by ionizing radiation, vol 11. CRC Press Inc, Boca Raton, pp 269–316
Tsubaki Y, Bunroongsook S (1990) Sexual competitive ability of mass-reared male sand mate preference in wild females: their effect on eradication of melonflies. Appl Entomol Zool 25:457–466
Van Ooik T, Rantala MJ, Saloniemi I (2007) Dietmediated effects of heavy metal pollution on growth and immune response in the geometrid moth Epirrita autumnata. Environ Pollut 145:348–354
Van Steenyk RA, Henneberry TJ, Ballmer GR, Wolf WW, Sevacherian V (1979) Mating competitiveness of laboratory culture sandsterilized Pink boll worm for use in a sterile moth release programme. J Econ Entomol 72:502–550
Vile GF, Tyrrell RM (1995) UVA radiation-induced oxidative damage to lipids and proteins in vitro and in human skin fibroblasts is dependent on iron and singlet oxygen. Free Radic Biol Med 18:721–730
Watters FL (1976) Microwave radiation for control of Tribolium confusum in wheat and flour. J Stored Prod Res 12:19–25
Wei GS, Zhang QW, Zhou MZ, Wu WG (2000) Studies on the phototaxis of Helicoverpa armigera. Acta Biophys Sin 16:89–95
Williams GC (1966) Natural selection, the costs of reproduction, and a refinement of Lack’s principle. Am Nat 100:687–690
Yang YG, He DY, Zhu SM, Yang PZ, Zho YS (1984) Studies on the generations and migration of armyworm Mythimna separata (Walker) in Yunnan province. Zool Res 1:1–20
Ye PY (2012) Farmers battle armyworms. http://www.globaltimes.cn/content/726757.shtml
Zera AJ, Denno RF (1997) Physiology and ecology of dispersal polymorphism in insects. Annu Rev Entomol 42:207–231
Zera AJ, Harshman LG (2001) The physiology of lifehistory trade-offs in animals. Annu Rev Ecol Syst 32:95–126
Zhang CY, Meng JY, Wang XP, Lei CL (2011) Effects of UV-A exposures on longevity and reproduction in Helicoverpa armigera, and on the development of its F1 generation. Insect Sci 18:697–702
Zhao S, Qiu C, Xiong S, Cheng X (2007) A thermal lethal model of rice weevils subjected to microwave irradiation. J Stored Prod Res 43:430–434
Zheng JL (2011) Effect of UVA radiation on biological fitness and antioxidant system of fruit fly, Drosophila Melanogaster. M.S. thesis, Huazhong Agricultural University, Wuhan, China
Acknowledgments
The research was supported by the National Department Public Benefit (Agriculture) Research Foundation (201403031) and the National Natural Science Foundation of China (no. 30871639).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article

Cite this article
Ali, A., Rashid, M.A., Huang, Q.Y. et al. Effect of UV-A radiation as an environmental stress on the development, longevity, and reproduction of the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae). Environ Sci Pollut Res 23, 17002–17007 (2016). https://doi.org/10.1007/s11356-016-6865-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-016-6865-0