
Abstract
To study the responses of digestive system of the freshwater crab Sinopotamon henanense to the exposure with cadmium (Cd), crabs were acutely exposed to 7.25, 14.50, and 29.00 mg/l Cd for 96 h and subchronically exposed to 0.725, 1.450, and 2.900 mg/l for 21 days. Cd bioaccumulation in the hepatopancreas and digestive tract (esophagus and intestine) was examined. Furthermore, histopathological alterations of the esophagus, midgut, hindgut, and hepatopancreas were assessed in animals from the 29.0 and 2.90 mg/l Cd treatment groups, and expression of metallothionein messenger RNA (MT mRNA) in the hepatopancreas and intestine was measured in all treatment groups. The results showed difference in the middle and high concentrations between acute and subchronic treatment groups. Cd content in digestive tract after acute 14.5 and 29.0 mg/l Cd exposure was significantly higher than that at subchronic 1.45 and 2.90 mg/l exposure, but Cd levels in hepatopancreas were not significantly different under the same condition. Acute exposure to Cd induced greater morphological damage than subchronic exposure: large areas of epithelial cells were necrotic in hepatopancreas and midgut, which detached from the basal lamina. Vacuolated muscle cells were observed in the hindgut of animals from the acute exposure group, but the changes of esophageal morphology were not obvious after acute or subchronic treatments. The expression of MT mRNA increased with increasing Cd concentration, and MT mRNA level in acute exposure groups was significantly lower when compared to the subchronic exposure groups. Higher Cd content and lower MT mRNA expression in the acutely exposed groups may be responsible for more severe damage of digestive system in these exposure groups.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cadmium (Cd) is a contaminant which is toxic to many aquatic animal species (Alazemi et al. 1996). It can be accumulated by aquatic animals through ingestion, respiration and adsorption (Hook and Fisher 2001), and it is commonly used in ecotoxicological studies (Nursita et al. 2009; Kang et al. 2012). Aquatic animals react to the exposure with Cd in various ways to reduce health effects: (1) reducing the absorption of Cd (lowering the metabolic rate) to decrease exchange with the external environmental (Barbieri 2009); (2) increasing the synthesis of Cd-chelating proteins [e.g., metallothionein (MT)] to neutralize superfluous Cd (Sparla and Overnell 1990); and (3) activating stress responses, such as superoxide dismutase (SOD) and other antioxidant defense systems (Atli and Canli 2010). When the rate of Cd accumulation surpasses the discharge and detoxification capacity of animals, this results in toxic effects through the formation of reactive oxygen species (ROS) that, in turn, result in oxidative deterioration of a variety of biological macromolecules such as lipids, proteins, and nucleic acids leading to cellular damage and initiating various pathological changes including cell death (Cao et al. 2010; Cuypers et al. 2010).
The freshwater crab Sinopotamon henanense is a species commonly found in the freshwater environment of China. Our earlier surveys on Cd accumulation in the S. henanense habitat of the Qin River showed that Cd levels could reach 10.6–14.3 mg/kg in surface sediments (Han et al. 2008). In some valleys near cadmium-rich mines, the Cd content in suspended matter has been reported to be as high as 232 mg/kg (Zhao et al. 2008). Furthermore, in some places, Cd contamination caused by human activity is much more prominent than those by the natural erosion process (Cheng et al. 2008). The effects of Cd poisoning are gradually reflected at the level of cells, tissues, and organs. The damages of organelles, cytomembrane, and organizational structure can result in dysfunctions of cells and tissues (Guardiola et al. 2013). Cd can cause different changes in the morphological structures of organs (e.g., gill and hepatopancreas, etc.) in aquatic animals, depending on its concentrations and the modes of exposure (Schuwerack and Lewis 2003; Annabi et al. 2011).
Histopathology represents a useful tool to assess the effects of pollution, particularly for sublethal and chronic effects (Cengiz and Unlu 2005). Histopathology has been widely used as a biomarker for the evaluation of the health effects of the exposure of aquatic animals to Cd (Kruatrachue et al. 2003; Ikechukwu and Ajeh 2011).
Metallothioneins (MTs) are a group of nonenzymatic proteins that are of low molecular mass (6,000–7,000 Da), cytoplasmic, single-chained, and rich in cysteine residues (up to 30 % of total amino acids). The latter provides MT with a high binding capacity for metals (Kovarova et al. 2009). One of the main mechanisms that protect organisms from damage by toxic heavy metals (e.g., Cd) is MT (Fang et al. 2010). Cd can induce the expression of the MT gene, which has been shown as a sensitive marker for detection of Cd stress in aquatic organisms (Dallinger 1994). Therefore, it has been proposed to use MT as a biomarker of environmental metal (especially Cd) pollution, using different organisms including both vertebrates (Ma et al. 2011) and invertebrates (Correia et al. 2002). For this reason MT is recognized along with a suite of other core biomarkers by the European Water Framework Directive (Amiard et al. 2006).
Under laboratory conditions, acute treatment is widely used, as the toxic effects of harmful substances on organisms can be rapidly detected in short exposure time when toxic substances are used at high doses. In natural ecosystems, however, the concentration of contaminants tend to be lower, and their toxic effects on physiological functions of aquatic animals are through long-term accumulation in the body; animals are unlikely to be exposed to steady-state acute or chronic concentration of contaminants (Thophon et al. 2003). Therefore, it is suggested that the stress response is distinct depending on toxicity test methods applied in a specific species such as acute versus chronic exposures. So far, little is known about the similarities and differences in metal bioaccumulation and morphological changes of the digestive system of crabs after acute compared to subchronic exposure to heavy metals.
The influence of water chemistry in modifying the uptake of metals at the gill is well understood (Luo et al. 2014; Kaya et al. 2015). However, research on similar influences within the digestive system that may modify dietary bioavailability is not well explored. The ability of digestion is one important physiological index of animal health. It could affect absorption of alimental nutrients and the growth of animals (Cui 2009). The hepatopancreas and intestine are important digestive organs of crustaceans (Strus et al. 2008). The main function of these two tissues is to digest food and the uptake of nutrients. Our earlier research (Wu et al. 2013) revealed that acute Cd exposure affected activities of digestive enzymes, thus impairing the normal digestive ability of the crab. The present study aimed to clarify the different effects of acute and subchronic Cd exposure on the digestive system of S. henanense, which will be helpful in differentiating acute/sudden from long-term Cd pollution.
Material and methods
Animals and treatments
Freshwater crabs S. henanense were purchased from the Wu Longkou Dong’an Aquatic Wholesale Market in Taiyuan. Crabs originated from a little village in Henan province without industrial pollution. Heavy metal concentrations (e.g., Cd) in water from the place where animals were collected were measured and confirmed to be low (0.0013 ± 0.0001 μg/l) by inductively coupled plasma mass spectrometry. Prior to experimentation, crabs were acclimated for 2 weeks in glass aquaria filled with tap water (aerated for 48 h, at a temperature of 16–22 °C, pH 6.8, and dissolved oxygen over 6 mg/l). Aquaria were shielded by a black plastic to reduce visual disturbance. Crabs were fed commercial fish feed (Porpoise Aquarium, China) two times a week.
After acclimation, healthy, similar-sized adult crabs (wet weight 18.7–23.6 g, carapace width 36.3–40.2 mm) were randomly divided into seven groups with 25 specimens in each group: six groups were exposed to different Cd concentrations with 2 L of CdCl2solution, and the seventh group served as a control (tap water). Exposure studies were performed in glass aquaria (50 cm × 30 cm × 25 cm). For 4-day acute exposure experiments, crabs were treated with 7.25, 14.5, or 29.0 mg/l Cd (by a sequential dilution from a 20 g/l stock solution), corresponding to 1/32, 1/16, and 1/8 of the Cds 96 h LC50 for S. henanense (Wang et al. 2008), respectively. Treatment solutions were replaced every 24 h to maintain nominal Cd concentrations. For the 21 days subchronic exposure, crabs were treated with 0.725, 1.45, and 2.90 mg/l Cd, and the treatment medium was changed every 48 h. All other conditions were kept the same as those during acclimation (temperature, pH, and dissolved oxygen). During the exposure period, crabs were fed two times a week (1 % of body weight), and the mortality and food intake of the experimental animals were observed. There was no mortality of experimental animals.
Metal determination
Concentrations of Cd were measured following the Chinese National Standard for determination of cadmium in food (GB/T5009.15-1996) with some modifications. After exposure, five crabs from each group were immobilized on ice for 15 min. Digestive tract and hepatopancreas tissues were excised and weighted (approximate 0.5 g wet weight), cut into small pieces, and excess water on surface of tissues was removed with absorbent paper. Then tissues were digested in 10 ml HNO3 (analytical grade) and 5 ml HClO4 (analytical grade) over a hot plate at about 120–150 °C, under a reflux cap. Cd concentrations of each sample were measured with an atomic absorption spectrophotometer (Shimadzu AA-6300, Japan). Standard Cd solution (national standard sample, GSB04-1721-2004, China) was used for the analysis of metal concentrations. The carrier gas was acetylene. Cd content was expressed as μg/g wet weight tissue.
Histopathological observation
Five crabs from each group were randomly selected from the control, 29.0 and 2.90 mg/l Cd exposure groups. Esophagus, midgut, hindgut, and hepatopancreas tissues were carefully excised. Then the tissues were preserved for 24 h at room temperature by direct immersion in a 0.1 M, pH 7.4, phosphate buffer with 4 % paraformaldehyde, followed by a routine histological procedure (Gurr 1962), where 4-μm-thick sections were obtained and stained with hematoxylin and eosin (H&E) for observation by light microscopy (Olympus BX51, Olympus, Tokyo, Japan). Three histological slides of each tissue were examined by light microscopy (600×); ten fields were observed in each slide (30 fields for each crab).
Following the scale suggested by Zodrow et al. (2004) and Chiodi Boudet et al. (2015), the degree of histological damage was scored according to the percentage of the total fields with histological damage found out of the total observed in the four tissues of each treatment. Scores were based on the number of fields in which histological changes were observed with (−) = no histopathology in any field, (+) = mild histopathology present in 25 % of the fields, (++) = moderate histopathology present in 25–75 % of the fields, and (+++) = severe histopathology present in >75 % of the fields.
Quantitative real-time PCR
Total RNA was extracted from 20 to 40 mg of intestine and hepatopancreas from five crabs in each group using RNeasy Mini Kit (Qiagen, Cat. No. 74104, USA), according to the manufacturer’s instructions. RNA quality was evaluated by electrophoresis on a 1 % agarose gel, and RNA concentrations as well as purity were determined by spectrophotometry (RNA/DNA Calculator, Eppendorf). First-strand complimentary DNA (cDNA) was then synthesized from 5 μg of RNA, and partial coding of sequences for MT and GAPDH was obtained using the Trans Script™ One-Step RT-PCR Super Mix (Transgene, Beijing, China), according to the manufacturer’s instructions. Specific primers were deduced from alignment of GAPDH sequences available from NCBI for different crab species using DNAMAN software (Lynnon BioSoft, Vaudreuil, Quebec, Canada). Primers for MT were according to Ma et al. (2009) and Gao et al. (2012) (Table 1).
Expression of messenger RNA (mRNA) for target genes was determined using quantitative real-time PCR (qRT-PCR) using a ABI 7500 Real-Time PCR System (Applied Biosystem, Foster, CA, USA) and Quanti Fast™ SYBR Green PCR Kit (Qiagen, Cat. No. 204054, USA), according to the manufacturer’s instructions.
The qRT-PCR reaction mixture consisted of 12.5 μl SYBR Green master mix, 0.5 μl of each forward and reverse primers, 1 μl of 50× diluted cDNA template and water to adjust to 25 μl. 25 μl of reaction mixture were loaded into ABI 7500 Real-Time PCR System and subjected to the following cycling: 5 min at 95 °C to denature DNA and activate Taq polymerase, 45 cycles of 10 s at 95 °C, 34 s at 60 °C, and dissociation stage. For each primer, serial dilutions of a cDNA standard were amplified in each run to determine amplification efficiency, and two negative controls were also amplified: nonreverse transcribed total RNA as a control of contamination by genomic DNA and a template negative sample to control for any contamination of the reagent. Transcripts of the GAPDH gene as the endogenous control were quantified with each MT gene sample normalized to GAPDH content.
Statistical analysis
The 2−△△CT method was used to analyze the expression level of MT mRNA (Livak and Schmittgen 2001). Statistical analyses were performed with SPSS 17.0 software. Data distribution and the homogeneity of variance were tested using Kolmogorov–Smirnov and Levene tests, respectively. When the data satisfied the prerequisites for parametric tests [analysis of variance (ANOVA)], one-way ANOVA and Dunnet’s test were used to evaluate the significance of differences between treated and control groups. The data were expressed as means ± SD. A post hoc least significant difference (LSD) test was performed for intergroup comparisons. Histopathological scoring was examined by ANOVA followed by Tukey’s post hoc test. Probability values of P < 0.05 were considered as statistically significant.
Results
Cd bioaccumulation in the digestive system of S. henanense
Cd levels were both increased in the intestine and hepatopancreas, which were under stress by Cd (Fig. 1), and reached maximum levels in the greatest acute (29.0 mg/l) and subchronic (2.90 mg/l) exposure groups. In the medium (14.5 mg/l) and high (29.0 mg/l) acute exposure groups, Cd content in the digestive tract increased significantly compared to the subchronic groups (1.45 and 2.90 mg/l, respectively) (Fig. 1a). When compared to the 0.725 mg/l Cd subchronic exposure group, Cd concentrations in the hepatopancreas showed a higher accumulation in the low acute concentration group (7.25 mg/l) (Fig. 1b).

Effect of acute and subchronic Cd exposures on Cd accumulation in digestive tract (a) and hepatopancreas (b) of S. henanense. Columns with the same lowercase letters indicate no significant difference between groups (P > 0.05), while different lowercase letters indicate a significant difference between groups (P < 0.05, mean ± SE, N = 5)
Histopathological investigation of the digestive system of S. henanense
The degree of histopathological changes of each tissue after acute and subchronic Cd exposure is significantly different among treatment groups with acutely treated animals showing more sever effects than organisms treated subchronically. Table 2 summarizes the histopathologic observations for control and two modes treated crab with representative images of the tissues displayed in Fig. 2. The most significant differences between control and Cd-exposed animals are observed in the midgut and hepatopancreas.

Effect of acute and subchronic Cd exposure on the digestive system of S. henanense. a1–a3: Esophagus. a1 Control group, 600×, the muscle cells are packed tightly. a2 Treated with acute 29.0 mg/l Cd, 600×, parts of the epithelial cells are swelling (black arrow). a3 Treated with subchronic 2.90 mg/l Cd, 600×, histopathological alterations are not observed. b1–b3 Midgut. b1 Control group, 1,000×, shape of nucleus is regular, and microvilli structure is obvious. b2 Treated with acute 29.0 mg/l Cd, 600×, the epithelial cells are severely necrotic (black arrow). b3 Treated with subchronic 2.90 mg/l Cd, 600×, leading to cell membrane rupture and nucleus deformation, and the cavity formed between epithelial cells and basal lamina (black arrow). c1–c3 Hindgut. c1 Control group, 600×, the muscle cells are packed tightly. c2 Treated with acute 29.0 mg/l Cd, 600×, serious edema and vacuolar degeneration (black arrow). c3 Treated with subchronic 2.90 mg/l Cd, 600×, the epithelial cells and muscle cells are vacuolized and slight edema (black arrow). d1–d3 Hepatopancreas. d1 Control group, 600×, the normal columnar epithelial cells are packed tightly. d2 Treated with acute 29.0 mg/l Cd, 600×, the residue of liver tubules is not dissolving completely (black arrow). d3 Treated with subchronic 2.90 mg/l Cd group, 600×, the epithelial cells are swelling and necrotic (black arrow). Ep epithelium cells, MF muscle fiber, Mv microvilli, Bl basal lamina, c chitin, E E cells, F F cells, R R cells, B B cells)
Esophagus
The normal esophagus mucosa consists of a simple columnar epithelium cell layer, and the cells are lined up in order. Myofilaments of control crabs were clearly and regularly arranged (Fig. 2a1). In crabs exposed to 29.0 mg/l of Cd for 4 days, the esophagus maintained the normal tissue structure. However, muscle cells showed significantly slight edema and vacuolar degeneration. Some epithelia were slightly damaged with small areas of broken or swollen cells (Table 2; Fig. 2a2, black arrow). Compared to the control group, crabs exposed to 2.90 mg/l Cd for 21 days did not show significant pathologic alterations of the esophagus (Table 2; Fig. 2a3).
Midgut
Under control conditions, the basal lamina of normal midgut could be clearly seen. Epithelial cells of the control group were in the original columnar form and packed tightly. The nuclei are spherical or ovoid, and typical and developed microvilli of the epithelium cells are clearly observed (Fig. 2b1). After 4-day exposure to 29.0 mg/l Cd, the morphological structure of midgut was significantly altered compared to controls (Table 2; Fig. 2b2, black arrow): epithelial cells were obviously damaged and almost completely separated from the basal lamina. The cell membrane was ruptured, the cytoplasm leaked out of the cells, and the nuclei were fragmented and scattered in the lumen (Fig. 2b2, black arrow). Compared with the acute treatment group, the degree of damages in the midgut of subchronic groups was not significant (Table 2). Some parts of the cell membrane were swollen and deformed, and the cell boundaries were fuzzy or disappeared. Microvilli were not well defined. The nucleuses became lightly stained and fragmented, and some epithelial cells were separated from the basal lamina (Fig. 2b3, black arrow). However, there was no significant evidence of necrosis.
Hindgut
No recognizable changes were observed in the hindgut of the control crabs throughout the course of the experiment. The epithelial lining of the hindgut was of mostly ciliated columnar type with scattered mucus-secreting goblet cells. Nuclei were ellipsoidal in shape, and they are situated in the center of the cells (Fig. 2c1). The hindgut of crabs exposed to 29.0 mg/l of Cd resulted in significant tissue degeneration and necrosis: most epithelial cells showed edema, dilatation of interstitial space, and marked vacuolar degeneration; the muscle cells showed severe edema and vacuolar degeneration (Table 2; Fig. 2c2, black arrow). Compared to the control group, slight alterations were observed in the histology of the hindgut of crabs after 21 days exposure to 2.90 mg/l Cd. However, swelling, vacuolization in epithelial cells and muscle cells were significantly appeared (Table 2; Fig. 2c3, black arrow).
Hepatopancreas
In the control group, the hepatopancreas was shaped like a circle or an ellipse and the inner surface was irregular but continuous. Various types of epithelial cells were arranged in neat rows and showed cell-specific typical pattern (Fig. 2d1). The morphological structure of the hepatic ducts was disorganized in the acute Cd treatment group. Vacuole areas appeared in epithelial cells, and significant proportions of cells were necrotic. Some epithelial cells of hepatic ducts were even dissolved completely, and cell debris was scattered along the basal lamina (Table 2; Fig. 2d2, black arrow). After 21 days, crabs from the 2.90 mg/l Cd treatment group showed swollen or necrotic epithelial cells, some of which had ruptured cytomembranes. Vacuole areas appeared. Karyorrhexis and necrosis were also detected (Table 2; Fig. 2d3, black arrow). These alters were significantly different from control group.
MT induction in digestive system of S. henanense after Cd exposure
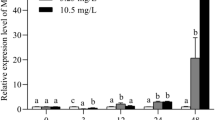
MT mRNA expression was significantly increased in both intestine (Fig. 3a) and hepatopancreas (Fig. 3b) with increasing Cd concentration after being acutely and subchronically exposed to Cd. MT abundances in crabs from the acute groups were significantly lower than those of animals from the subchronic groups at medium and high Cd concentrations (P < 0.05).

Effects of Cd on MT mRNA expression in intestine (a) and hepatopancreas (b) of S. henanense. Columns with the same lowercase letters indicate no significant difference between groups (P > 0.05), while different lowercase letters indicate a significant differences between groups (P < 0.05, mean ± SE, N = 5)
Discussion
The function of the crustacean digestive tract is to digest food, to absorb nutrition, and to excrete food residues (Barker and Gibson 1978). The hepatopancreas is the most important digestive gland and detoxification organ for crustaceans (Lou et al. 2010). Aquatic organisms can accumulate Cd through ingestion of food or water by the digestive system (Bais and Lokhande 2012). Therefore, in addition to the gills, the digestive system is one of the primary target organs of heavy metal pollution. The results of Cd accumulation in the digestive tract and hepatopancreas of crabs after acute and subchronic treatment showed that Cd levels in each treatment group were significantly higher than those in the control group. However, compared with the subchronic treatment groups, Cd levels in hepatopancreas were not significantly different in the middle- and high-concentration acute groups, and we hypothesized that Cd accumulation in hepatopancreas might have reached saturation after exposure to 14.5 and 29.0 mg/l Cd. The hepatopancreas is one of the most important organs that plays an important role in heavy metal detoxification. Thus, it is the key site of Cd accumulation in crustaceans (Kaoud and Eldahshan 2010). Cd accumulation in tissues of organisms can activate the corresponding detoxification mechanisms, such as an increase in the expression of MT, which is related to Cd excretion and detoxification and, therefore, functions in lowering Cd toxicity (Klaassen et al. 2009; Martinez-Finley and Aschner 2011). When the in vivo Cd concentration exceeds the MT-binding capacity, saturation is reached. Thus, extra Cd will bind to macromolecules like enzymes or other proteins, and triggers a toxic response in organisms (Ma et al. 2008). In the present study, acute Cd treatment led to elevated expression of MT in hepatopancreas, which binds to Cd as a response and reaches saturation fast. This possibly resulted in Cd accumulation to maximum tissue levels. In contrast, during subchronic exposures, crabs might have gradually adapted to Cd pollution. The study of Silvestre et al. (2005) showed that the Cd accumulated in the digestive organ of the Chinese crab Eriocheir sinensis could have gradually been transferred to other tissues such as the muscle, carapace, etc. This may be the reason that the Cd accumulation of digestive tract in the acute treatment groups was higher than that in the subchronic groups (Silvestre et al. 2005).
In our experiment, bioaccumulation reached the highest level after exposure to 29.0 and 2.90 mg/l Cd in the acute and chronic exposure groups, respectively, which may have resulted in multiple physiological and pathological changes. Histopathological analysis showed that there were significant increases in the prevalence of changes of morphological structure of tissues in the 29.0 mg/l acute Cd group compared to the 2.90 mg/l subchronic exposure group. Damages to the hepatopancreas were more severe than those in the other three tissues. Among the different tissues, the esophagus was the most resistant to Cd toxicity (Table 2), the reason for which may be that the esophagus is the primary digestive organ and has no further digestion function and detoxification function (Jiang and Yan 2009).
After acute 29.0 mg/l Cd treatment, a large number of midgut epithelial cells were necrotic and separated from the basal lamina. In contrast, in the subchronic Cd treatment group, the injury to the midgut was milder than that of the acute group, but epithelial cells lost their normal morphology (Table 2). As an important digestive organ in crustaceans, the midgut is not only involved in the synthesis and secretion of a variety of digestive enzymes and in effective food digestion but also involved in the absorption of nutrients (Li et al. 1994, 2008). Furthermore, the epithelial cells of midgut are covered by microvilli that enlarge the area for digestion and absorption (Copenhaver et al. 1971). Therefore, morphological changes such as loss of membrane microvilli, cytoplasmic hypervacuolization and epithelial necrosis of the midgut in both Cd treatment groups are expected to impact its normal digestive function.
There are several ridges formed by the intestinal wall towards the luminal stenosis in the hindgut providing the spaces for formation and storage of feces. A muscular layer then supports the discharge of feces (Barker and Gibson 1977). After acute and chronic exposure to Cd, the observed histological changes of the hindgut led to a dilation of epithelial cells and muscle cells, which contained more vacuoles. Tanhan et al. (2005) reported a decrease in ciliary length or microvilli in esophagus, stomach, rectum, and gill of Babylonia areolata under Cd stress. The same holds for an enlargement of vacuoles by fusion of vesicles, and vacuoles in the cells might result from the interaction of heavy metals with membranes. These reactions might induce changes in the composition, fluidity, and stability of the membranes. The intensified fusion between small and large vacuoles resulted in an increase of large vacuoles after intoxication. Tanhan et al. (2005) also suggested that the increase of fluid content might cause some of the symptoms of poisoning.
The hepatopancreas is a very important organ for crustaceans. While it can reduce or even eliminate the toxic effects of external chemicals to crustacean, on the other side, it has also been shown to be vulnerable to harmful substances including inorganics and organics (Liu et al. 2013; Stará et al. 2014). It is, therefore, very sensitive to external environmental changes (Johnston et al. 1998). In the present study, acute Cd treatment led to serious injuries of the hepatopancreas, and necrosis was observed in large areas of the hepatic tubules. Similar results were also found in hepatic tubules of other aquatic animals, such as Lamellidens marginalis, which were acutely exposed to Cd (Yasmeen et al. 2012). In the subchronic Cd treatment groups, large areas of vacuoles appeared in the hepatopancreas. The formation of intracellular vacuoles may be a mechanism to counteract poisoning by isolating Cd in these vacuoles (Coombs and George 1978). After acute and chronic exposure to Cd, morphology of the hepatopancreas of crabs was altered to different degrees, which was likely to affect the digestion, absorption, secretion, and detoxification process, leading to a decrease in metabolism, a reduction of crab activities or even death of the organisms.
MT is related to metabolism of trace elements and has been proposed as an intracellular antioxidant by sequestering reactive metals and inactivating hydroxyl radicals and superoxide radicals (Nordberg 1998). It is also involved in the metabolism of Cd, such as absorption, transport, accumulation, and excretion, and has a high affinity to Cd. After exposure to Cd, the damage to the intestine and hepatopancreas were more severe than that to the esophagus. These two tissues were analyzed for MT mRNA differential expression. The results of the experiment showed that MT mRNA became significantly induced in both intestine and hepatopancreas of crabs. With increasing Cd concentrations, MT mRNA expression was increased, which provides a high binding capacity for Cd (Xiang et al. 2013; Vincent-Hubert et al. 2014). Furthermore, MT mRNA levels in the acutely 29.0 mg/l exposed groups were significantly lower in both tissues compared to those in the subchronically 2.90 mg/l exposed groups. This can be explained by a delay between Cd accumulation and MT gene expression in tissues. The induction of MT is one of the primary responses in animals after chronic Cd treatment (Moltedo et al. 2000; Trinchella et al. 2006). However, Goto and Wallace (2007) showed that only 36.3 % of Cd was chelated by heat stable proteins (MT) when Cd was accumulated in the body of the polychaete (Capitella capitata) after 50 μg/l Cd stress for 1 week. In the present study, the concentration of the treatment was higher than that of the study of Goto and Wallace (2007), which indicated that more toxic effects were induced by excess Cd. Therefore, although the increase of MT expression was used to chelate Cd in cells, injuries of the digestive system by unbound Cd could not be prevented and eliminated.
According to our results, morphological damages to the intestine and hepatopancreas in the acute groups were more severe than in subchronic groups, which is consistent with the results of Cd accumulation. This may be due to the high Cd concentrations applied in the acute groups. It is difficult for crabs to adapt to the surrounding Cd-polluted environment in a short time, and water-borne Cd was taken into their bodies through digestion and respiration and accumulated to a degree that exceeded the MT-binding capacity. Extra Cd can lead to the generation of reactive oxygen species, causing lipid peroxidation and cell oxidative damage (Souid et al. 2013), as well as injuries to tissues and organs. While in the subchronic groups, crabs were provided with lower Cd concentrations at long-term exposure than acute groups, Cd absorbed by the intestine was gradually transferred to other detoxification tissues like hepatopancreas, and the detoxification mechanisms of organisms were likely to have slowly adapted to the toxic insult. It is assumed that this adaptation represents a metal resistance mechanism (Sevcikova et al. 2011). Therefore, high concentrations during the acute Cd exposure might be more toxic to crabs. It has to be investigated whether this holds for other aquatic organisms as well.
Conclusion
The present study demonstrated that Cd-induced damages in the organs of the digestive system in S. henanense also caused histopathological changes as well as a differential expression of MT mRNA. General changes in all organs were mostly similar and increased from subchronic to acute Cd exposure.
References
Alazemi BM, Lewi JW, Andrews EB (1996) Gill damage in the freshwater fish Gnathonemus petersii (family: Mormyridae) exposed to selected pollutants: an ultrastructural study. Environ Technol 17:225–238
Amiard JC, Amiard-Triquet C, Barka S, Pellerin J, Rainbow PS (2006) Metallothioneins in aquatic invertebrates: their role in metal detoxification and their use as biomarkers. Aquat Toxicol 76:160–202
Annabi A, Messaoudi I, Kerkeni A, Said K (2011) Cadmium accumulation and histological lesion in mosquetofish (Gambusia affinis) tissues following acute and chronic exposure. Int J Environ Res 5(3):745–756
Atli G, Canli M (2010) Response of antioxidant system of freshwater fish Oreochromis niloticus to acute and chronic metal (Cd, Cu, Cr, Zn, Fe) exposures. Ecotoxicol Environ Saf 73:1884–1889
Bais UE, Lokhande MV (2012) Effect of cadmium chloride on histopathological changes in the freshwater fish Ophiocephalus striatus (Channa). Int J Zool Res 8(1):23–32
Barbieri E (2009) Effects of zinc and cadmium on oxygen consumption and ammonium excretion in pink shrimp (Farfantepenaeus paulensis, Pérez-Farfante 1967, Crustacea). Ecotoxicology 18:312–318
Barker PL, Gibson R (1977) Observations on the feeding mechanism, structure of the gut, and digestive physiology of the European lobster Homarus gammarus (L.) (Decapoda: Nephropidae). J Exp Mar Biol Ecol 26(3):297–324
Barker PL, Gibson RJ (1978) Observations on the structure of the mouthparts, histology of the alimentary tract, and digestive physiology of the mud crab Scylla serrata (Decapoda: Portunidae). J Exp Mar Biol Ecol 58:175–191
Cao L, Huan W, Liu JH, Yin XB, Dou SZ (2010) Accumulation and oxidative stress biomarkers in Japanese flounder larvae and juveniles under chronic cadmium exposure. Comp Biochem Physiol C 151:386–392
Cengiz EI, Unlu E (2005) Sublethal effects of commercial delta merthrin on the structure of the gill, liver and gut tissues of mosquitofish, Gambusia affinis: a microscopic study. Environ Toxicol Pharmacol 21:246–253
Cheng HX, Zhao CD, Zhuang GM, XiaWL LYH, Yang K, Nie HF (2008) Reconstruction of the regional soil pollution history by heavy metals in Taihu lake drainage area: taking Pb and Cd as examples. Earth Sci Front 5:167–178
Chiodi Boudet LN, Polizzi P, Romero MB, Robles A, Marcovecchio JE, Gerpe MS (2015) Histopathological and biochemical evidence of hepatopancreatic toxicity caused by cadmium in white shrimp, Palaemonetes argentines. Ecotoxicol Environ Saf 113:231–240
Coombs J, George SG (1978) Mechanisms of immobilization and detoxification of metals in marine organisms. In: McLusky DS, Berry AJ (eds) Physiology and behavior of marine organisms [M]. Pergamon Press, London, pp 179–187
Copenhaver WM, Bunge RP, Bunge MB (1971) Bailey’s textbook of histology [M], 16th edn, Figs. 16–40. Willams & Wilkins, Baltimore, p 446
Correia AD, Livingstone DR, Costa MH (2002) Effects of water-borne copper on metallothionein and lipid peroxidation in the marine amphipod Gammarus locusta. Mar Environ Res 54:357–360
Cui F (2009) Study on cadmium enrichment in aquatic animals and its effect on growth and digestive enzyme activities. Thesis for Master of Science. Nanjing Agricultural University, Nanjing
Cuypers A, Plusquin M, Remans T, Jozefczak M, Keunen E, Gielen H, Opdenakker K, Nair AR, Munters E, Artois TJ, Nawrot T, Vangronsveld J, Smeets K (2010) Cadmium stress: an oxidative challenge. Biometals 23:927–940
Dallinger T (1994) Invertebrate organisms as biological indicators of heavy metal pollution. Appl Biochem Biotechnol 48:27–31
Fang Y, Yang HS, Wang TM, Liu BZ, Zhao HL, Chen MY (2010) Metallothionein and superoxide dismutase responses to sublethal cadmium exposure in the clam Mactra veneriformis. Comp Biochem Physiol C 151(3):325–33
Gao AB, Wang L, Yuan H (2012) Expression of metallothionein cDNA in a freshwater crab, Sinopotamon yangtsekiense, exposed to cadmium. Exp Toxicol Pathol 64(3):253–258
Goto D, Wallace WG (2007) Interaction of Cd and Zn during uptake and loss in the polychaete Capitella capitata: whole body and subcellular perspectives. J Exp Mar Biol Ecol 352(1):65–77
Guardiola FA, Cuest A, Meseguer J, Martínez S, Martínez-Sánchez MJ, Pérez-Sirvent C, Esteban MA (2013) Accumulation, histopathology and immune toxicological effects of waterborne cadmium on gilthead seabream (Sparus aurata). Fish Shellfish Immunol 35(3):792–800
Gurr E (1962) Staining animal tissues: practical and theoretical, 2nd edn. Leonard Hill (Hooks), London
Han T, Wang Q, Wang L (2008) Ecological investigation of freshwater crab and river pollution in basin of Qinhe River. Sichuan J Zool 27:804–806
Hook SE, Fisher NS (2001) Reproductive toxicity of materials in calanoid copepods. Mar Biol 138:1131–1140
Ikechukwu EL, Ajeh EA (2011) Histopathological alterations in the liver and lungs of Hoplobatrachus occipitalis exposed to sub lethal concentrations of cadmium. Aust J Basic Appl Sci 5(11):1062–1068
Jiang YH, Yan SF (2009) Histological and histochemical studies on the digestive tract of Panulirus stimpsoni. J Jimei Univ (Nat Sci) 14(3):1–7
Johnston DJ, Alexander CG, Yellowlees D (1998) Epithelial cytology and function in the digestive gland of Thenus orientalis (Decapoda: Scyllaridae). J Crust Biol 18:271–278
Kang XJ, Mu SM, Li WY, Zhao N (2012) Toxic effects of cadmium on crabs and shrimps. Toxici Drug Test. ISBN: 978-953-51-0004-1
Kaoud HA, Eldahshan AR (2010) Bioaccumulation of cadmium in the freshwater prawn Macrobrachium rosenbergii. Nat Sci 8(4):157–168
Kaya ED, Söyüt H, Beydemir Ş (2015) The toxicological impacts of some heavy metals on carbonic anhydrase from gilthead sea bream (Sparus aurata) gills. Environ Toxicol Pharmacol 39(2):825–832. doi:10.1016/j.etap.2015.01.021
Klaassen CD, Liu J, Diwan BA (2009) Metallothionein protection of cadmium toxicity. Toxicol Appl Pharmacol 238:215–220
Kovarova J, Kizek R, Adam V, Harustiakova D, Celechovska O, Svobodova Z (2009) Effect of cadmium chloride on metallothionein levels in carp. Sensors 9:4789–4803
Kruatrachue M, Rangsayatorn N, Pokethitiyook P, Upatham ES, Singhakaew S (2003) Histopathological changes in the gastrointestinal tract of fish, Puntius gonionotus, fed on dietary cadmium. Bull Environ Contam Toxicol 71:561–569
Li TW, Su XR, Zhang F (1994) Histology, histochemistry of the digestive system in blue crab Portunus trituberculatus (Decapoda, Portunidae). J Liaoning Normal Univ (Nat Sci) 17(3):230–237
Li CL, Cao FJ, Huang XH, Liu CW (2008) Histological observation on alimentary system of Panulirus homarus with light and scanning electron microscopy. J Trop Oceanogr 27(3):72–78
Liu DM, Yang J, Wang L (2013) Cadmium induces ultrastructural changes in the hepatopancreas of the freshwater crab Sinopotamon henanense. Micron 47:24–32
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real time quantitative PCR and the 2 (-Delta DeltaC (T)) method. Methods 25:402–408
Lou D, Yang JF, Xie H, Chen JG (2010) A preliminary study of comparative histology of major organs from Portunus trituberculatus and Sinopotamon. J Mar Sci 28(3):72–78
Luo LZ, Ke CH, Guo XY, Shi B, Huang MQ (2014) Metal accumulation and differentially expressed proteins in gill of oyster (Crassostrea hongkongensis) exposed to long-term heavy metal-contaminated estuary. Fish Shellfish Immunol 38(2):318–329
Ma WL, Wang L, He YJ, Yang T (2008) Cadmium accumulation and metallothionein biosynthesis in the fresh-water crab Sinopotamon henanense. Acta Sci Circumst 28(6):1192–1197
Ma WL, Yan T, He YJ, Wang L (2009) Purification and cDNA cloning of a cadmium-binding metallothionein from the freshwater crab Sinopotamon henanense. Arch Environ Contam Toxicol 56:747–753
Ma Y, Lin J, Zhang C, Ren Y, Lin J (2011) Cd(II) and As (III) bioaccumulation by recombinant Escherichia coli expressing oligomeric human metallothioneins. J Hazard Mater 185(2–3):1605–1608
Martinez-Finley EJ, Aschner M (2011) Revelations from the nematode Caenorhabditis elegans on the complex interplay of metal toxicological mechanisms. J Toxicol 5(3):745–756
Moltedo O, Verde C, Capasso A, Parisi E, Remondelli P, Bonatti S, Alvarez-Hernandez X, Glass J, Alvino CG, Leone A (2000) Zinc transport and metallothionein secretion in the intestinal human cell line Caco-2. J Biol Chem 275:31819–31825
Nordberg M (1998) Metallothioneins: historical review and state of knowledge. Talanta 46:243–254
Nursita AI, Singh B, Lees E (2009) Cadmium bioaccumulation in Proisotoma minuta in relation to bioavailability in soils. Ecotoxicol Environ Saf 72:1767–1773
Schuwerack PM, Lewis JW (2003) The mode of action of acute and chronic concentrations of waterborne Cd in the digestive gland of the acclimated infested freshwater crab (Potamonautes warreni). Cell Tissue Res 312:249–263
Sevcikova M, Modra H, Slaninova A, Svobodova Z (2011) Metals as a cause of oxidative stress in fish: a review. Vet Med-Czech 56(11):537–546
Silvestre F, Duchêne C, Trausch G, Devos P (2005) Tissue-specific cadmium accumulation and metallothionein-like protein levels during acclimation process in the Chinese crab Eriocheir sinensis. Comp Biochem Physiol C 140(1):39–45
Souid G, Souayed N, Yaktiti F, Maaroufi K (2013) Effect of acute cadmium exposure on metal accumulation and oxidative stress biomarkers of Sparus aurata. Ecotoxicol Environ Saf 89:1–7
Sparla AM, Overnell J (1990) The binding of cadmium to crab cadmium metallothionein. Biochem J 267:539–540
Stará A, Kouba A, Velíšek J (2014) Effect of chronic exposure to prometryne on oxidative stress and antioxidant response in red swamp crayfish (Procambarus clarkii). Biomed Res Int 2014:680131
Strus J, Klepal W, Repina J, Tusek-Znidaric M, Milatovic M, Pipan Z (2008) Ultrastructure of the digestive system and the fate of midgut during embryonic development in Porcellio scaber (Crustacea: Isopoda). Arthropod Struct Dev 37(4):287–298
Tanhan P, Sretarugsa P, Pokethitiyook P, Kruatrachue M, Upatham ES (2005) Histopathological alterations in the edible snail, Babylonia areolata (Spotted Babylon), in acute and sub-chronic cadmium poisoning. Environ Toxicol 20:142–149
Thophon S, Kruatrachue M, Upatham ES, Pokethitiyook P, Sahaphong S, Jaritkhuan S (2003) Histopathological alterations of white seabass, Lates calcarifer, in acute and subchronic cadmium exposure. Environ Pollut 121(3):307–320
Trinchella F, Riggio M, Filosa S, Volpe MG, Parisi E, Scudiero R (2006) Cadmium distribution and metallothionein expression in lizard tissues following acute and chronic cadmium intoxication. Comp Biochem Physiol C 144(3):272–278
Vincent-Hubert F, Châtel A, Gourlay-Francé C (2014) Metallothionein mRNA induction is correlated with the decrease of DNA strand breaks in cadmium exposed zebra mussels. Mutat Res-Gen Tox En 766:10–15
Wang L, Yan B, Liu N, Li YQ, Wang Q (2008) Effects of cadmium on glutathione synthesis in hepatopancreas of freshwater crab, Sinopotamon yangtsekiense. Chemosphere 74:51–56
Wu H, Xuan RJ, Li YJ, Zhang XM, Wang Q, Wang L (2013) Effects of cadmium exposure on digestive enzymes, antioxidant enzymes and lipid peroxidation in the freshwater crab Sinopotamon henanense. Environ Sci Pollut Res 20:4085–4092
Xiang DF, Zhu JQ, Jin S, Hu YJ, Tan FQ, Yang, WX (2013) Expression and function analysis of metallothionein in the testis of Portunus trituberculatus exposed to cadmium. Aquat Toxicol 140–141: 1–10
Yasmeen S, Suryawanshi GD, Mane UH (2012) Histopathological changes in the hepatopancreas of Lamellidens marginalis exposed to cadmium in winter. Int Multidiscip Res J 2(6):41–43
Zhao CD, Chen FR, Chen XR, Zhao HC, Xia WL, Nie HF, Kong M, Liu F, Yang K (2008) A methodology of tracking sources of cadmium anomalies and their quantitative estimation in the Yangtze River basin. Earth Sci Front 15:179–193
Zodrow JM, Stegeman JJ, Tanguay RL (2004) Histological analysis of acute toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in zebrafish. Aquat Toxicol 66:25–38
Acknowledgments
This work was supported by the National Nature Science Foundation (No. 31272319) and the Overseas Returnee Research Fund in Shanxi Province (No. 201015). The authors are thankful to Prof. Hans-Uwe Dahms and Shaoqin Li for critical amendments to the manuscript.
Ethical statement
On behalf of, and having obtained permission from all the authors, I declare that:
-
a)
The material has not been published in whole or in part elsewhere;
-
b)
The paper is not currently being considered for publication elsewhere;
-
c)
All authors have been personally and actively involved in substantive work leading to the report and will be jointly and individually responsible for its content themselves; and
-
d)
All relevant ethical safeguards have been met in relation to animal experimentation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Markus Hecker
Rights and permissions
About this article

Cite this article
Wu, H., Li, Y., Lang, X. et al. Bioaccumulation, morphological changes, and induction of metallothionein gene expression in the digestive system of the freshwater crab Sinopotamon henanense after exposure to cadmium. Environ Sci Pollut Res 22, 11585–11594 (2015). https://doi.org/10.1007/s11356-015-4419-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-015-4419-5