
Abstract
The role of extracellular nucleotides as modulators of inflammation and cell stress is well established. One of the main actions of these molecules is mediated by the activation of purinergic receptors (P2) of the plasma membrane. P2 receptors can be classified according to two different structural families: P2X ionotropic ion channel receptors, and P2Y metabotropic G protein-coupled receptors. During inflammation, damaged cells release nucleotides and purinergic signaling occurs along the temporal pattern of the synthesis of pro-inflammatory and pro-resolving mediators by myeloid and lymphoid cells. In macrophages under pro-inflammatory conditions, the expression and activity of cyclooxygenase 2 significantly increases and enhances the circulating levels of prostaglandin E2 (PGE2), which exerts its effects both through specific plasma membrane receptors (EP1-EP4) and by activation of intracellular targets. Here we review the mechanisms involved in the crosstalk between PGE2 and P2Y receptors on macrophages, which is dependent on several isoforms of protein kinase C and protein kinase D1. Due to this crosstalk, a P2Y-dependent increase in calcium is blunted by PGE2 whereas, under these conditions, macrophages exhibit reduced migratory capacity along with enhanced phagocytosis, which contributes to the modulation of the inflammatory response and tissue repair.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Inflammation specificities and factors involved
The regulation of the inflammatory response remains a central aspect in the understanding of many pathological processes [1,2,3,4,5,6]. The three phases that characterize inflammation, i.e., initiation, extension, and repair/resolution, are controlled by a large number of factors with specific temporal and intensity patterns [7, 8]. These profiles vary between tissues and species, defining the course of the pathological process and the impact on the organisms [9]. However, despite the selectivity of many inflammatory reactions, there is an overlap in the molecular pathways involved. This diversity in the interactions between them defines specific fates in their control and the possible therapeutic interventions [10,11,12]. An example of this is the involvement of P2X7 receptor signaling in the activation of the NLRP3 inflammasome, which requires the involvement of an additional priming signal from the TLR2/4 pathway [13,14,15,16,17].
It is worth mentioning that the production of different bioactive lipids, such as prostanoids, is a common determinant in the progression of inflammatory processes (Fig. 1) [18,19,20]. The most abundant prostanoids from pro-inflammatory macrophages are synthesized after the expression of cyclooxygenase 2 (COX-2), which catalyzes the first step in the biosynthesis of prostanoids from arachidonic acid [21,22,23,24,25,26,27,28]. The end products of the COX-2 pathway are the result of additional modifications via the action of cell-specific prostaglandin synthases (Fig. 2) [29]. COX-2 is encoded by the PTGS2 gene in humans (Ptgs2 in rodents) and it is expressed in the early stages of inflammation. The transcription of the PTGS2 gene is extensively induced in many inflammatory cells and tissues, except in hepatocytes, where only after preliminary pathological changes (i.e., liver regeneration after partial hepatectomy) is the ability to express COX-2 recovered [26, 30, 31]. In the liver, this regulatory bias is only associated with hepatocytes, since Kupffer cells retain this pro-inflammatory activation [28, 32,33,34,35,36,37]. This interesting mechanism reflects the fact that, under physiological conditions, the portal blood contains pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), which do not activate COX-2 expression through cell surface receptors that recognize PAMP or DAMP [38, 39].

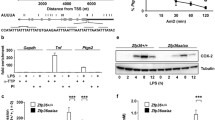
Time course of the serum levels of PGE2 and 15-deoxy-Δ.12,14-PGJ2 (15dPGJ2) in mice injected intraperitoneally with LPS. 12-month-old male mice (n = 7) received 1 mg/kg body weight of E. coli LPS (serotype 0055:B5) and the serum levels of early pro-inflammatory (PGE2) and anti-inflammatory (15dPGJ2) prostaglandins were determined using specific ELISA kits. The graph represents the mean values and shows a minimal overlapping of both prostaglandins (unpublished results from the authors). The structures of PGE2 and 15dPGJ2 are shown. 15dPGJ2 has a cyclopentenone structure which is responsible for its reactivity to perform Michael addition reactions on thiol groups from amino acids (a carbonyl group surrounded by α, β instaurations on the cyclopentenone ring; yellow stars and circle)

Schematic representation of the synthesis of PGs and pro-resolving lipids from arachidonic acid. Cyclooxygenases (COX-1 and COX-2) and lipoxygenases (5-LOX and 15-LOX) are the initial enzymes that direct the biotransformation of the arachidonic acid after activation of plasma membrane phospholipases. Red names/lines, the main pro-inflammatory molecules; blue names/lines, the main anti-inflammatory/pro-resolving molecules; in black the molecules that play a dual role in inflammation. TX, thromboxane; LT, leukotriene; LX, lipoxin; PGF2, prostaglandin F2α (also known as dinoprost); PGI2, prostacyclin; 15S-HpETE, 15-hydroperoxyicosa-5,8,11,13-tetraenoic acid
Dual role of prostaglandins in the regulation of inflammation
The prostanoids synthesized by the COX-2 pathway can act in opposite ways: they can exert pro-inflammatory actions, but they can also promote and activate anti-inflammatory mechanisms [40,41,42,43]. An example of this dual role is PGE2, which is one of the major products of the COX-2 pathway [44,45,46,47,48,49,50]. Other prostaglandins, such as prostaglandin 15-deoxy-Δ12,14-prostaglandin J2 (15dPGJ2) are potent anti-inflammatory molecules because their chemical structure contains a cyclopentenone motif (due to the presence of α,β-unsaturated carbonyl groups). This chemical structure allows for non-enzymatic reactions with cysteine residues in proteins, via Michael addition modifications (Figs. 1 and 2) [40, 51,52,53]. These Michael adducts have an impact on the enzyme activity and function of different proteins involved in the control of the inflammatory processes, such as the transcription factor NF-κB, which exerts an important activation of the pro-inflammatory response, and is inhibited by Michael addition of 15dPGJ2 [54]. In contrast, transcription factors that repress the progression of inflammation, such as the peroxisomal proliferator-activated receptor γ (PPARγ) are activated by 15dPGJ2 by this post-translational modification via Michael addition [51, 55].
Mechanisms of action of prostaglandin E2
In recent years, several groups have been interested in the role of prostanoids in the regulation of the inflammatory process. Our group focused on studying the effect of PGE2 accumulation at sites of inflammation, using cells and animal models deficient in COX-2 or expressing a transgene encoding COX-2, or by administering selective COX-2 inhibitors (called generically coxibs [56,57,58]), but maintaining the activity of COX-1, an enzyme that contributes to the synthesis of prostanoids in healthy conditions [40, 45, 50, 59, 60].
Regarding the mechanism of action, PGE2 binds to and activates specific G protein-coupled membrane receptors called E-type PGE2 receptors (EP receptors; Fig. 3). Four different receptors, EP1 to EP4, have been identified from a biochemical and pharmacological point of view [44, 61]. Interestingly, these receptors are not exclusively expressed on the plasma membrane, but also on other intracellular membranes, such as the nuclear membrane [62]. Activation of EP1 promotes the mobilization of intracellular Ca2+ stores through activation of the phosphoinositide 3-kinase pathway. This transient change in cytoplasmic Ca2+ has an impact on ionic fluxes, cellular metabolism and organelle function (i.e., mitochondria), and activates Ca2+-dependent enzymes, such as various isoforms of protein kinase C (PKC). Therefore, PGE2 induces Ca2+- and PKC-dependent effects in cells expressing EP1 [63, 64]. A relevant fact of EP1 is that the expression profile in cells is different between humans and rodents, which makes it difficult to translate the results between different species [65].

Signaling in response to PGE2 biosynthesis. High throughput biosynthesis of PGE2 is produced after the expression of COX-2 and activity of the prostaglandin E synthase. PGE2 can be exported by the cells and act as an agonist of EP1 to EP4 receptors. Each EP receptor is coupled to specific G proteins that mediate their action. In addition to these plasma membrane receptors, PGE2 can be incorporated into the cell via the PG transporter (PGT). In PGE2 synthesizing cells, the intracellular presence of this PG can act on EP receptors present at the nuclear membrane. However, the role of these nuclear EP receptors and the mechanisms of signaling are poorly characterized
The binding of PGE2 to EP2 and EP4 receptors promotes the dissociation of the Gαs/Gβγ complex from the G protein-coupled receptor. The Gαs subunit stimulates adenylate cyclase activity, which increases the intracellular levels of cyclic AMP (cAMP) and, therefore, activates the protein kinase A-dependent pathway [61]. However, EP2 and EP4 have partially non-overlapping functions: EP2 is mainly involved in smooth muscle cell relaxation, whereas EP4 activation exhibits pro- and anti-inflammatory functions ranging from vasodilation to angiogenesis, and metastasis progression [66, 67]. Unlike EP2/EP4, activation of EP3 leads to a reduction in intracellular cAMP levels [68]. These EP receptors are expressed on various cell types and provide the basis for therapeutic interventions, using selective agonists and antagonists. However, in addition to EP-mediated effects, PGE2 can exert other actions, either by accessing the cytoplasm or through binding to additional receptors, for example through purinergic signaling, although these mechanisms are less characterized, which explains the effects independent of pharmacological targeting of the EP receptors [69].
Purinergic signaling in inflammation
Inflammation involves a large number of molecules, including cytokines, chemokines, prostanoids, and extracellular nucleotides that are released during inflammation and activate myeloid and lymphoid immune cells [5, 48, 70,71,72,73]. Extracellular nucleotides (i.e. ATP and UTP) have been recognized as a new class of innate immune regulators that act through the P2 receptors and modulate the inflammatory reaction [74,75,76,77,78]. These extracellular nucleotides, which are released at sites of inflammation due to infection or cell damage, contribute to immune cell activation, including cytoskeleton reorganization, cell migration, phagocytosis and exocytosis [72]. Extracellular nucleotides also exert tissue-specific actions. For example, in the brain, they have been associated with different pathologies affecting immune cells (microglia), such as neuropathic pain; indeed, targeting extracellular nucleotide signaling is a pharmacological therapeutic tool that is being investigated in clinical trials [76, 79,80,81,82,83]. Purine and pyrimidine nucleotide receptors are involved in many neuronal and non-neuronal mechanisms: in short-term signaling, they are involved in the regulation of neurotransmission, neuromodulation of inflammation and neurosecretion, promotion of platelet aggregation and vasodilation; and in long-term actions, they are associated with cell proliferation, differentiation, motility, cell-death in development and regeneration.
Currently, the accepted P2Y receptors are P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13 and P2Y14 [84]. Among the metabotropic P2Y receptors, P2Y2, P2Y4 and P2Y6 are activated by uridine and adenine nucleotides [72, 74, 75, 78] and are coupled to phospholipase C (PLC) activation. As a consequence of the release of nucleotides into the extracellular medium, the agonistic action on P2Y receptors promotes an increase in the intracellular concentration of diacylglycerol (DAG) and inositol triphosphate (IP3), which induces the release of calcium from intracellular stores and the activation of several signaling pathways [85, 86]. P2Y receptors are expressed on various cell types and are functionally relevant in the activation of resident and circulating immune cells [5, 24, 71, 74, 87,88,89,90].
Crosstalk between PGE2 and P2 receptors in macrophages
The interaction between purinergic signaling and prostanoids has been described in different cell types. In macrophages, exposure to UTP increases the expression of COX-2, and nitric oxide synthase 2 (NOS2) under pro-inflammatory conditions [88, 91]. Macrophages can be polarized into pro-inflammatory (‘classically activated’ or M1, using microbial stimuli such as LPS, or cytokines such as IFNγ) or anti-inflammatory/pro-resolving phenotypes (‘alternatively activated’ or M2, using IL4 and/or IL13 as stimuli) [92,93,94,95,96,97,98,99,100,101,102,103]. Because macrophages can adopt different functional profiles this crosstalk between PGs and P2 signaling can contribute to the polarization of these cells. Therefore, the activation of P2 receptors helps to modulate the function of macrophages in the context of the environmental signals that govern the fate of the inflammatory response.
The presence of locally elevated concentrations of extracellular ATP promotes the activation of the P2X7 receptor, while UTP and UDP, and lower concentrations of ATP act mainly through P2Y2, P2Y4 and P2Y6, respectively [77, 78, 85, 104,105,106]. Nevertheless, the contribution of P2Y2/P2Y4 or P2Y4/P2Y6 heterodimers can also be considered in this regulatory hub [107, 108].
The signaling through the P2X7 receptor in macrophages is by far the most studied purinergic pathway. This is because P2X7 receptor activation participates in the regulation of several stress signal pathways and, more importantly, activates the NLRP3 inflammasome cascade [109,110,111]. It is well known that P2X7 activation by ATP contributes to the regulation of the innate response in macrophages: it favors the host defense against intracellular pathogens, an effect that is triggered by the release of reactive oxygen and/or nitrogen species [112, 113]. In addition to this, the activation of the NLRP3 pathway promotes the maturation of pro-inflammatory cytokines (i.e., IL-1β and IL-18), and an increase in the PGE2 levels. The pathways involved include a rise in Ca2+ influx and the activation of the MAP kinase signaling pathways [13, 16, 111].
Interestingly, the crosstalk between P2Y receptors and PGE2 has also been reported in macrophages from P2X7 receptor-deficient mice, or after inhibition of the receptor with Brilliant Blue G as well as with the receptor antagonist A 438079, which indicates that the interaction between P2Y receptors and PGE2 is independent of P2X7 receptors [24, 71, 88, 114, 115]. Furthermore, macrophages challenged with specific agonists of the P2X7 receptors did not show the inhibitory effect of PGE2 on Ca2+-mobilization [71]. Regarding the role of the polarization phenotype of macrophages on the expression levels of purinergic receptors, M1 and M2 differentiated cells exhibit similar values, both in RNA and protein levels. However, pro-inflammatory macrophages display rapid and time-dependent repression of the levels of the downstream receptor-associated phospholipase C β1 and β2 isoenzymes, which contribute to the reduced signaling dependent on P2Y receptor activation [116, 117].
The effect of extracellular ATP on the progression of the anti-inflammatory phenotype in macrophages does not involve P2Y/P2X receptor-mediated processes but rather depends on pyrophosphate ATP bonds. The pathways involved promote a reorganization of the actin cytoskeleton that favors the clustering of these actin filaments, which ultimately contribute to the clustering and organization of the NLRP3 inflammasome complex. In addition, the participation of ectonucleotidases seems to contribute to the transition of macrophages from a pro-inflammatory (M1) to an anti-inflammatory (M2) phenotype. This transition is believed to facilitate the resolution of the inflammatory reaction accomplished by macrophages [118,119,120].
Interestingly, unlike naïve and M2 polarized macrophages, M1 cells do not display the inhibitory effect of PGE2 on Ca2+ mobilization [24, 71]. These polarization specificities were observed in both rodent and human macrophages. As for the mechanism by which M1 macrophages fail to show this PGE2-dependent P2Y desensitization, it has been shown to occur at least two hours after the pro-inflammatory challenge. This suggests that this is not the result of the rapid signaling elicited after TLR4 and/or pro-inflammatory cytokine receptors engagement, but rather is due to secondary events in the signaling process. From a mechanistic point of view, the sustained response to P2Y receptors in the presence of PGE2, as occurs in M1 macrophages ensures the activity of the purinergic signaling in the early steps of inflammation [71, 88, 91, 115, 121]. As an extension, in platelets, a cross-desensitization between ADP and the thromboxane receptor signaling has been reported [122, 123]. All of these interactions play an important role in several inflammatory and degenerative disorders, such as multiple sclerosis, amyotrophic lateral sclerosis and Alzheimer´s disease [124,125,126]. Indeed, in these pathologies, extracellular ATP exerts pro-inflammatory actions that cause the release of cytokines and the production of PG. Interestingly, this modulation could play an important role in the anti-inflammatory effects of PGE2.
A relevant aspect in this context of the heterogeneity of P2Y/P2X receptors is the possible crosstalk between the P2X and P2Y receptor families [127,128,129]. An example is the synergism between both families in the activation of dendritic cells, which are necessary for the efficient initiation of immune responses [130]. In addition to antigens, the presence of P2 agonists released by necrotic cells results in a synergistic activation and maturation of dendritic cells, and therefore, in more efficient signaling in T cells, leading to increased expression of pro-inflammatory mediators and adhesion molecules.
Molecular mechanisms involved in PGE2-P2Y receptor crosstalk
The pathways involved in the crosstalk between P2Y receptors and PGE2 on macrophages have been established using biochemical (inhibitors and activators of signal transduction pathways), pharmacological (mainly through the use of agonists and antagonists of the EP and P2Y receptors) and genetic (cells lacking P2X7 receptor or COX-2; expressing a COX-2 transgene or expressing different constructs of the proteins that participate in the signal-transduction pathways) approaches [24, 71, 88, 115, 131, 132]. Based on the data from these different strategies it was concluded that PKD1 phosphorylation at S916 is a necessary condition to suppress PGE2-dependent UTP-mediated Ca2+-mobilization. In contrast, selective inhibition of PKD1 is sufficient to attenuate the effect of PGE2 on P2Y signaling. PKDs are ubiquitously expressed and regulate various cellular processes, including oxidative stress, gene expression, cell survival, vesicle trafficking and, interestingly, P2X7 signaling, although their precise function in macrophages remains poorly characterized. Analysis devoted to identifying the PKD isoform(s) involved in this P2Y crosstalk showed that PKD1, which is regulated by extracellular ligands in macrophages, is specifically targeted [24, 71]. Furthermore, overexpression of PKD1 reduced the effect of UTP on Ca2+ mobilization but when a vector encoding a catalytically inactive kinase of PKD1 was expressed, the response to UTP persisted and the inhibitory effect of PGE2 was abolished (Fig. 4) [71].

Crosstalk between PGE2 and P2Y receptors in macrophages. Pro-inflammatory macrophages express high levels of COX-2 that promote a rapid increase in PGE2 synthesis and release. In pro-inflammatory macrophages (M1-type), PGE2 is unable to affect the signaling of P2Y receptors. However, naïve, resting, or alternatively activated macrophages (M2-type) exhibit an impaired P2Y receptor signaling that results in a blockade of Ca2+-dependent mobilization. This inhibitory effect of PGE2 depends on the activities of PKD1 and PKCε and interferes with the different pathways modulated by the transient increase in Ca2+ due to P2Y agonists. In cerebellar astrocytes the EP3 receptor is also involved. Red lines and arrows, pro-inflammatory pathways; blue lines and arrows, resting and anti-inflammatory pathways. PKC, protein kinase C; PKD, protein kinase D; PKD-DN, a dominant-negative form of PKD; AKT, protein kinase B; P, phosphorylation
In fact, an association of PKD1 with TLR9 and, in general, with the MyD88-dependent pro-inflammatory innate immune responses has been described [133, 134]. Additionally, PKCδ activation has been reported to act as an upstream PKD1 activation step. However, transfection of macrophages with constitutively active PKCδ constructs did not mimic the effects of PGE2 on UTP-dependent Ca2+ mobilization. However, expression in macrophages of a constitutively active PKCε, but not of other classical, new, or atypical PKCs, was sufficient to mimic the effects of PGE2 on P2Y receptors in terms of Ca2+ mobilization [71].
Regarding the role of macrophage polarization in this PGE2-P2Y crosstalk, naïve and anti-inflammatory/pro-resolving (M2) macrophages show this inhibitory interaction, but it was not observed in those that were polarized to M1 pro-inflammatory cells. Under these M1 conditions, PGE2-dependent phosphorylation of PKD1 at S916 is not observed, while naïve and M2 macrophages exhibit this PKD1 phosphorylation [24, 71]. This phosphorylation of PKD1 at S916 has been reported to correspond to a fully activated PKD1. Moreover, activation of PKD1 has been associated with the response to upstream PKCs and/or activation of G-proteins and various receptor-associated tyrosine kinases [135]. The PGE2-dependent activation of PKD1 promotes DAGs release not only at the plasma membrane level but also from other compartments, such as the endoplasmic reticulum and the Golgi apparatus. Interestingly, PKD activation plays a role in the crosstalk between P2Y and P2X receptors (Fig. 4). In line with this, P2X4 receptor signaling favors the activation of phospholipase A2 (PLA2) and, in turn, the supply of substrates for COX-2 and, therefore, the increase in the release of PGE2 that participates in the intercellular crosstalk between P2X and P2Y receptors [107, 136].
The regulation of P2Y activity in macrophages, which involves the participation of PGE2, has functional implications in the basic biological responses of these cells, such as metabolic activation and migration. In this regard, cell migration contributes to normal development and differentiation. Recent data indicate that extracellular nucleotides can regulate the migration and attachment activities of “professional phagocytes” (macrophages, neutrophils and microglia) and other cell types (i.e., fibroblasts, endothelial cells, neurons and keratinocytes) [137,138,139]. From a functional point of view, it has been shown that PGE2 inhibits P2Y-dependent macrophage migration, even in the presence of other chemoattractants. These chemotactic actions are common for several P2Y receptors, such as P2Y2, P2Y4, and P2Y6 [140,141,142]. These observations are consistent with the fact that P2 receptors participate in a wide range of phagocytic and chemotactic actions, as described for P2Y2,4,6 receptors in the phagocytosis of apoptotic bodies by microglial cells. In addition to these signaling mechanisms, the EP3 receptors have been involved in the impairment of Ca2+-mobilization by PGE2 in cerebellar astrocytes [88].
Interestingly, PGE2 promotes the internalization of P2Y4 in fibroblasts transfected with COX-2, an effect that is suppressed after the inhibition of COX-2 with the coxib DFU [24]. Moreover, the blockade in Ca2+-mobilization by PGE2 has an important consequence in terms of the activation of different signaling pathways in fibroblasts, including activation of various PKCs and the energetic metabolism via activation of AMP-dependent protein kinase (AMPK) and inhibition of acetyl-CoA carboxylase (ACC) [24, 71]. Again, this regulatory network is suppressed when fibroblasts are in an inflammatory environment. Recent trends in tissue repair of inflammatory lesions have focused on the interaction between stromal cells, such as macrophages and fibroblasts. Based on these observations, it can be proposed that targeting the stromal microenvironment is likely to be an important and promising strategy for future anti-inflammatory and pro-resolution therapies.
In summary, the translation of basic studies on the interactions between prostaglandin synthesis and the signaling through P2Y and P2X receptors in the immune system to clinical trials can result in the development of new therapeutic options to modulate the course of inflammatory diseases.
Data availability
Data and comments that support this study are available from the corresponding authors upon request.
Abbreviations
- cAMP:
-
Cyclic AMP
- DAMPs:
-
Damage-Associated Molecular Patterns
- 15dPGJ2 :
-
15-Deoxy-Δ.12,14-Prostaglandin J2
- DAG:
-
Diacylglycerol
- DFU:
-
5,5-Dimethyl-3-(3-fluorophenyl)-4-(4-methylsulphonyl)phenyl-2(5H)-furanone
- IFNγ:
-
Interferon-γ
- IL:
-
Interleukin
- LPS:
-
Lipopolysaccharide
- MyD88:
-
Myeloid Differentiation Primary Response 88
- NLRP3:
-
NLR Family Pyrin Domain Containing 3
- NF-κB:
-
Nuclear Factor κB
- PAMPs:
-
Pathogen-Associated Molecular Patterns
- PL:
-
Phospholipase
- PG:
-
Prostaglandin
- PGE2 :
-
Prostaglandin E2
- EP:
-
Prostaglandin E2 Receptor
- PKC:
-
Protein kinase C
- PKD:
-
Protein kinase D
- P2:
-
Purinergic Receptors
- TLR:
-
Toll-Like Receptor
References
Fullerton JN, Gilroy DW (2016) Resolution of inflammation: a new therapeutic frontier. Nat Rev Drug Discov 15:551–567. https://doi.org/10.1038/nrd.2016.39
Chen L, Deng H, Cui H et al (2017) Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 9:7204–7218. https://doi.org/10.18632/oncotarget.23208
Bours MJ, Swennen EL, Di Virgilio F et al (2006) Adenosine 5’-triphosphate and adenosine as endogenous signaling molecules in immunity and inflammation. Pharmacol Ther 112:358–404. https://doi.org/10.1016/j.pharmthera.2005.04.013
Chiurchiu V, Leuti A, Maccarrone M (2018) Bioactive Lipids and Chronic Inflammation: Managing the Fire Within. Front Immunol 9:38. https://doi.org/10.3389/fimmu.2018.00038
Idzko M, Ferrari D, Eltzschig HK (2014) Nucleotide signalling during inflammation. Nature 509:310–317. https://doi.org/10.1038/nature13085
Nathan C (2002) Points of control in inflammation. Nature 420:846–852. https://doi.org/10.1038/nature01320
Goldberg EL, Dixit VD (2015) Drivers of age-related inflammation and strategies for healthspan extension. Immunol Rev 265:63–74. https://doi.org/10.1111/imr.12295
Boscá L, González-Ramos S, Prieto P et al (2015) Metabolic signatures linked to macrophage polarization: from glucose metabolism to oxidative phosphorylation. Biochem Soc Trans 43:740–744. https://doi.org/10.1042/BST20150107
Uguccioni M, Teixeira MM, Locati M, Mantovani A (2017) Editorial: Regulation of Inflammation, Its Resolution and Therapeutic Targeting. Front Immunol 8:415. https://doi.org/10.3389/fimmu.2017.00415
Pajares MI, Rojo A, Manda G et al (2020) Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 9:1687. https://doi.org/10.3390/cells9071687
Germano G, Mantovani A, Allavena P (2011) Targeting of the innate immunity/inflammation as complementary anti-tumor therapies. Ann Med 43:581–593. https://doi.org/10.3109/07853890.2011.595732
Giglio RV, Pantea Stoian A, Al-Rasadi K et al (2021) Novel Therapeutical Approaches to Managing Atherosclerotic Risk. Int J Mol Sci 22:4633. https://doi.org/10.3390/ijms22094633
Song N, Liu Z-S, Xue W et al (2017) NLRP3 Phosphorylation Is an Essential Priming Event for Inflammasome Activation. Mol Cell 68:185-197.e6. https://doi.org/10.1016/j.molcel.2017.08.017
Kelley N, Jeltema D, Duan Y, He Y (2019) The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int J Mol Sci 20:3328. https://doi.org/10.3390/ijms20133328
Jiang H, He H, Chen Y et al (2017) Identification of a selective and direct NLRP3 inhibitor to treat inflammatory disorders. J Exp Med 214:3219–3238. https://doi.org/10.1084/jem.20171419
Yang Y, Wang H, Kouadir M et al (2019) Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis 10:128. https://doi.org/10.1038/s41419-019-1413-8
Li N, Zhou H, Wu H et al (2019) STING-IRF3 contributes to lipopolysaccharide-induced cardiac dysfunction, inflammation, apoptosis and pyroptosis by activating NLRP3. Redox Biol 24:101215. https://doi.org/10.1016/j.redox.2019.101215
Serhan CN, Levy BD (2018) Resolvins in inflammation: emergence of the pro-resolving superfamily of mediators. J Clin Invest 128:2657–2669. https://doi.org/10.1172/JCI97943
Lehmann C, Homann J, Ball AK et al (2015) Lipoxin and resolvin biosynthesis is dependent on 5-lipoxygenase activating protein. FASEB J 29:5029–5043. https://doi.org/10.1096/fj.15-275487
Buckley CD, Gilroy DW, Serhan CN (2014) Proresolving lipid mediators and mechanisms in the resolution of acute inflammation. Immunity 40:315–327. https://doi.org/10.1016/j.immuni.2014.02.009
Ulmann L, Hirbec H, Rassendren F (2010) P2X4 receptors mediate PGE2 release by tissue-resident macrophages and initiate inflammatory pain. EMBO J 29:2290–2300. https://doi.org/10.1038/emboj.2010.126
Parkinson JF (2006) Lipoxin and synthetic lipoxin analogs: an overview of anti-inflammatory functions and new concepts in immunomodulation. Inflamm Allergy Drug Targets 5:91–106. https://doi.org/10.2174/187152806776383125
Francés DE, Motiño O, Agrá N et al (2015) Hepatic cyclooxygenase-2 expression protects against diet-induced steatosis, obesity, and insulin resistance. Diabetes 64:1522–1531. https://doi.org/10.2337/db14-0979
Pimentel-Santillana M, Través PG, Pérez-Sen R et al (2014) Sustained Release of Prostaglandin E 2 in Fibroblasts Expressing Ectopically Cyclooxygenase 2 Impairs P2Y-Dependent Ca 2+ -Mobilization. Mediators Inflamm 2014:1–9. https://doi.org/10.1155/2014/832103
Frances DE, Ingaramo PI, Mayoral R et al (2013) Cyclooxygenase-2 over-expression inhibits liver apoptosis induced by hyperglycemia. J Cell Biochem 114:669–680. https://doi.org/10.1002/jcb.24409
Goren N, Cuenca J, Martin-Sanz P, Boscá L (2004) Attenuation of NF-?B signalling in rat cardiomyocytes at birth restricts the induction of inflammatory genes. Cardiovasc Res 64:289–297. https://doi.org/10.1016/j.cardiores.2004.06.029
Callejas NA, Fernández-Martínez A, Castrillo A et al (2003) Selective inhibitors of cyclooxygenase-2 delay the activation of nuclear factor kappa B and attenuate the expression of inflammatory genes in murine macrophages treated with lipopolysaccharide. Mol Pharmacol 63:671–677. https://doi.org/10.1124/mol.63.3.671
Hortelano S (2007) Animal models for the study of liver regeneration: role of nitric oxide and prostaglandins. Front Biosci 12:13. https://doi.org/10.2741/2045
Castro-Sanchez L, Agra N, Llorente Izquierdo C et al (2013) Regulation of 15-hydroxyprostaglandin dehydrogenase expression in hepatocellular carcinoma. Int J Biochem Cell Biol 45:2501–2511. https://doi.org/10.1016/j.biocel.2013.08.005
Prieto P, Jaén RI, Calle D et al (2019) Interplay between post-translational cyclooxygenase-2 modifications and the metabolic and proteomic profile in a colorectal cancer cohort. World J Gastroenterol 25:433. https://doi.org/10.3748/wjg.v25.i4.433
Jaén RI, Prieto P, Casado M et al (2018) Post-translational modifications of prostaglandin-endoperoxide synthase 2 in colorectal cancer: An update. World J Gastroenterol 24:5454–5461. https://doi.org/10.3748/wjg.v24.i48.5454
Mayoral R, Mollá B, Flores JM et al (2008) Constitutive expression of cyclo-oxygenase 2 transgene in hepatocytes protects against liver injury. Biochem J 416:337–346. https://doi.org/10.1042/BJ20081224
Martín-Sanz P, Casado M, Boscá L et al (2017) Cyclooxygenase 2 in liver dysfunction and carcinogenesis: Facts and perspectives. World J Gastroenterol 23:3572–3580. https://doi.org/10.3748/wjg.v23.i20.3572
Martín-Sanz P, Callejas NA, Casado M et al (1998) Expression of cyclooxygenase-2 in foetal rat hepatocytes stimulated with lipopolysaccharide and pro-inflammatory cytokines. Br J Pharmacol 125:1313–1319. https://doi.org/10.1038/sj.bjp.0702196
Casado M, Callejas NA, Rodrigo J et al (2001) Contribution of cyclooxygenase-2 to liver regeneration after partial hepatectomy. FASEB J 15:2016–2018. https://doi.org/10.1096/fj.01-0158fje
Martin-Sanz P, Mayoral R, Casado M et al (2010) COX-2 in liver, from regeneration to hepatocarcinogenesis: what we have learned from animal models? World J Gastroenterol 16:1430–1435. https://doi.org/10.3748/wjg.v16.i12.1430
Motino O, Agra N, Brea Contreras R et al (2016) Cyclooxygenase-2 expression in hepatocytes attenuates non-alcoholic steatohepatitis and liver fibrosis in mice. Biochim Biophys Acta 1862:1710–1723. https://doi.org/10.1016/j.bbadis.2016.06.009
Delgado C, Ruiz-Hurtado G, Gómez-Hurtado N et al (2015) NOD1, a new player in cardiac function and calcium handling. Cardiovasc Res 106:375–386. https://doi.org/10.1093/cvr/cvv118
Tang D, Kang R, Coyne CB et al (2012) PAMPs and DAMPs: signal 0s that spur autophagy and immunity. Immunol Rev 249:158–175. https://doi.org/10.1111/j.1600-065X.2012.01146.x
Scher JU, Pillinger MH (2009) The anti-inflammatory effects of prostaglandins. J Investig Med 57:703–708. 10.231/JIM.0b013e31819aaa76
Moro K, Nagahashi M, Ramanathan R et al (2016) Resolvins and omega three polyunsaturated fatty acids: Clinical implications in inflammatory diseases and cancer. World J Clin Cases 4:155. https://doi.org/10.12998/wjcc.v4.i7.155
Serhan CN (2014) Pro-resolving lipid mediators are leads for resolution physiology. Nature 510:92–101. https://doi.org/10.1038/nature13479
Batista-Gonzalez A, Vidal R, Criollo A, Carreño LJ (2020) New Insights on the Role of Lipid Metabolism in the Metabolic Reprogramming of Macrophages. Front Immunol 10:1–7. https://doi.org/10.3389/fimmu.2019.02993
O’Callaghan G, Houston A (2015) Prostaglandin E2 and the EP receptors in malignancy: possible therapeutic targets? Br J Pharmacol 172:5239–5250. https://doi.org/10.1111/bph.13331
Yuan C, Smith WL (2015) A cyclooxygenase-2-dependent prostaglandin E2 biosynthetic system in the Golgi apparatus. J Biol Chem 290:5606–5620. https://doi.org/10.1074/jbc.M114.632463
Madrigal JLM, Moro MA, Lizasoain I et al (2003) Induction of Cyclooxygenase-2 Accounts for Restraint Stress-Induced Oxidative Status in Rat Brain. Neuropsychopharmacology 28:1579–1588. https://doi.org/10.1038/sj.npp.1300187
Callejas NA (2001) Expression of cyclooxygenase-2 promotes the release of matrix metalloproteinase-2 and -9 in fetal rat hepatocytes. Hepatology 33:860–867. https://doi.org/10.1053/jhep.2001.23002
Ito M, Matsuoka I (2008) Regulation of purinergic signaling by prostaglandin E2 in murine macrophages. J Pharmacol Sci 107:443–450. https://doi.org/10.1254/jphs.08087fp
Xu J, Chalimoniuk M, Shu Y et al (2003) Prostaglandin E2 production in astrocytes: regulation by cytokines, extracellular ATP, and oxidative agents. Prostaglandins Leukot Essent Fat Acids 69:437–448
Brea R, Motiño O, Francés D et al (2018) PGE2 induces apoptosis of hepatic stellate cells and attenuates liver fibrosis in mice by downregulating miR-23a-5p and miR-28a-5p. Biochim Biophys Acta - Mol Basis Dis 1864:325–337. https://doi.org/10.1016/j.bbadis.2017.11.001
Millán O, Rico D, Peinado H et al (2006) Potentiation of tumor formation by topical administration of 15-deoxy-delta12,14-prostaglandin J2 in a model of skin carcinogenesis. Carcinogenesis 27:328–336. https://doi.org/10.1093/carcin/bgi213
Oliva JL, Pérez-Sala D, Castrillo A et al (2003) The cyclopentenone 15-deoxy-Δ 12,14 -prostaglandin J 2 binds to and activates H-Ras. Proc Natl Acad Sci 100:4772–4777. https://doi.org/10.1073/pnas.0735842100
Hortelano S, Castrillo A, Alvarez AM, Boscá L (2000) Contribution of Cyclopentenone Prostaglandins to the Resolution of Inflammation Through the Potentiation of Apoptosis in Activated Macrophages. J Immunol 165:6525–6531. https://doi.org/10.4049/jimmunol.165.11.6525
Das S, Kashyap N, Kalita S, et al (2020) A brief insight into the physicochemical properties of room-temperature acidic ionic liquids and their catalytic applications in C C bond formation reactions. pp 1–98
Castrillo A, Díaz-Guerra MJ, Hortelano S et al (2000) Inhibition of IkappaB kinase and IkappaB phosphorylation by 15-deoxy-Delta(12,14)-prostaglandin J(2) in activated murine macrophages. Mol Cell Biol 20:1692–1698. https://doi.org/10.1128/mcb.20.5.1692-1698.2000
Consalvi S, Biava M, Poce G (2015) COX inhibitors: a patent review (2011–2014). Expert Opin Ther Pat 25:1357–1371. https://doi.org/10.1517/13543776.2015.1090973
Vosooghi M, Amini M (2014) The discovery and development of cyclooxygenase-2 inhibitors as potential anticancer therapies. Expert Opin Drug Discov 9:255–267. https://doi.org/10.1517/17460441.2014.883377
Garcia Rodriguez LA, Cea-Soriano L, Tacconelli S, Patrignani P (2013) Coxibs: pharmacology, toxicity and efficacy in cancer clinical trials. Recent Results Cancer Res 191:67–93. https://doi.org/10.1007/978-3-642-30331-9_4
Cebola I, Custodio J, Muñoz M et al (2015) Epigenetics override pro-inflammatory PTGS transcriptomic signature towards selective hyperactivation of PGE2 in colorectal cancer. Clin Epigenetics 7:74. https://doi.org/10.1186/s13148-015-0110-4
Kirkby NS, Chan MV, Lundberg MH et al (2013) Aspirin-triggered 15-epi-lipoxin A4 predicts cyclooxygenase-2 in the lungs of LPS-treated mice but not in the circulation: implications for a clinical test. Faseb J 27:3938–3946. https://doi.org/10.1096/fj.12-215533
Sugimoto Y, Narumiya S (2007) Prostaglandin E receptors. J Biol Chem 282:11613–11617. https://doi.org/10.1074/jbc.R600038200
Bhattacharya M, Peri KG, Almazan G et al (1998) Nuclear localization of prostaglandin E2 receptors. Proc Natl Acad Sci USA 95:15792–15797. https://doi.org/10.1073/pnas.95.26.15792
Narumiya S (2007) Physiology and pathophysiology of prostanoid receptors. Proc Japan Acad Ser B 83:296–319. https://doi.org/10.2183/pjab.83.296
Tanaka Y, Furuyashiki T, Momiyama T et al (2009) Prostaglandin E receptor EP1 enhances GABA-mediated inhibition of dopaminergic neurons in the substantia nigra pars compacta and regulates dopamine level in the dorsal striatum. Eur J Neurosci 30:2338–2346. https://doi.org/10.1111/j.1460-9568.2009.07021.x
Markovič T, Jakopin Ž, Dolenc MS, Mlinarič-Raščan I (2017) Structural features of subtype-selective EP receptor modulators. Drug Discov Today 22:57–71. https://doi.org/10.1016/j.drudis.2016.08.003
Regan JW (2003) EP2 and EP4 prostanoid receptor signaling. Life Sci 74:143–153. https://doi.org/10.1016/j.lfs.2003.09.031
Vleeshouwers W, van den Dries K, de Keijzer S et al (2021) Characterization of the Signaling Modalities of Prostaglandin E2 Receptors EP2 and EP4 Reveals Crosstalk and a Role for Microtubules. Front Immunol 11:613286. https://doi.org/10.3389/fimmu.2020.613286
Reader J, Holt D, Fulton A (2011) Prostaglandin E2 EP receptors as therapeutic targets in breast cancer. Cancer Metastasis Rev 30:449–463. https://doi.org/10.1007/s10555-011-9303-2
Meves H (2006) The Action of Prostaglandins on Ion Channels. Curr Neuropharmacol 4:41–57. https://doi.org/10.2174/157015906775203048
Myrtek D, Muller T, Geyer V et al (2008) Activation of human alveolar macrophages via P2 receptors: coupling to intracellular Ca2+ increases and cytokine secretion. J Immunol 181:2181–2188
Través PG, Pimentel-Santillana M, Carrasquero LMG et al (2013) Selective impairment of P2Y signaling by prostaglandin E2 in macrophages: Implications for Ca 2+ -dependent responses. J Immunol 190:4226–4235. https://doi.org/10.4049/jimmunol.1203029
Marques-da-Silva C, Burnstock G, Ojcius DM, Coutinho-Silva R (2010) Purinergic receptor agonists modulate phagocytosis and clearance of apoptotic cells in macrophages. Immunobiology 216:1–11. https://doi.org/10.1016/j.imbio.2010.03.010
Vitiello L, Gorini S, Rosano G, la Sala A (2012) Immunoregulation through extracellular nucleotides. Blood 120:511–518. https://doi.org/10.1182/blood-2012-01-406496
Kong Q, Quan Y, Tian G et al (2021) Purinergic P2 Receptors: Novel Mediators of Mechanotransduction. Front Pharmacol 12:671809. https://doi.org/10.3389/fphar.2021.671809
Boeynaems J-M, Communi D, Gonzalez NS, Robaye B (2005) Overview of the P2 Receptors. Semin Thromb Hemost 31:139–149. https://doi.org/10.1055/s-2005-869519
Inoue K (2022) Overview for the study of P2 receptors: From P2 receptor history to neuropathic pain studies. J Pharmacol Sci 149:73–80. https://doi.org/10.1016/j.jphs.2022.04.003
Abbracchio MP, Burnstock G, Boeynaems JM et al (2006) International Union of Pharmacology LVIII: update on the P2Y G protein-coupled nucleotide receptors: from molecular mechanisms and pathophysiology to therapy. Pharmacol Rev 58:281–341. https://doi.org/10.1124/pr.58.3.3
Ralevic V, Burnstock G (1998) Receptors for purines and pyrimidines. Pharmacol Rev 50:413–492
Tsuda M, Beggs S, Salter MW, Inoue K (2013) Microglia and intractable chronic pain. Glia 61:55–61. https://doi.org/10.1002/glia.22379
Norenberg W, Hempel C, Urban N et al (2011) Clemastine potentiates the human P2X7 receptor by sensitizing it to lower ATP concentrations. J Biol Chem 286:11067–11081. https://doi.org/10.1074/jbc.M110.198879
Raouf R, Chabot-Dore AJ, Ase AR et al (2007) Differential regulation of microglial P2X4 and P2X7 ATP receptors following LPS-induced activation. Neuropharmacology 53:496–504. https://doi.org/10.1016/j.neuropharm.2007.06.010
Xia M, Zhu Y (2011) Signaling pathways of ATP-induced PGE2 release in spinal cord astrocytes are EGFR transactivation-dependent. Glia 59:664–674. https://doi.org/10.1002/glia.21138
Beggs S, Trang T, Salter MW (2012) P2X4R(+) microglia drive neuropathic pain. Nat Neurosci 15:1068–1073. https://doi.org/10.1038/nn.3155
Jacobson KA, Delicado EG, Gachet C et al (2020) Update of P2Y receptor pharmacology: IUPHAR Review 27. Br J Pharmacol 177:2413–2433. https://doi.org/10.1111/bph.15005
Burnstock G (2012) Purinergic signalling: Its unpopular beginning, its acceptance and its exciting future. BioEssays 34:218–225. https://doi.org/10.1002/bies.201100130
Dal Ben D, Buccioni M, Lambertucci C et al (2015) Purinergic P2X receptors: Structural models and analysis of ligand-target interaction. Eur J Med Chem 89:561–580. https://doi.org/10.1016/j.ejmech.2014.10.071
Leon-Otegui M, Gomez-Villafuertes R, Diaz-Hernandez JI et al (2011) Opposite effects of P2X7 and P2Y2 nucleotide receptors on alpha-secretase-dependent APP processing in Neuro-2a cells. FEBS Lett 585:2255–2262. https://doi.org/10.1016/j.febslet.2011.05.048
Paniagua-Herranz L, Gil-Redondo JC, Queipo MJ et al (2017) Prostaglandin E2 Impairs P2Y2/P2Y4 receptor signaling in cerebellar astrocytes via EP3 receptors. Front Pharmacol 8:e937. https://doi.org/10.3389/fphar.2017.00937
Stachon P, Heidenreich A, Merz J et al (2017) P2X7 Deficiency Blocks Lesional Inflammasome Activity and Ameliorates Atherosclerosis in Mice. Circulation 135:2524–2533. https://doi.org/10.1161/circulationaha.117.027400
Stachon P, Geis S, Peikert A et al (2016) Extracellular ATP Induces Vascular Inflammation and Atherosclerosis via Purinergic Receptor Y2 in Mice. Arter Thromb Vasc Biol 36:1577–1586. https://doi.org/10.1161/atvbaha.115.307397
Chen BC, Lin WW (2000) Pyrimidinoceptor potentiation of macrophage PGE(2) release involved in the induction of nitric oxide synthase. Br J Pharmacol 130:777–786. https://doi.org/10.1038/sj.bjp.0703375
Palmieri EM, Gonzalez-Cotto M, Baseler WA et al (2020) Nitric oxide orchestrates metabolic rewiring in M1 macrophages by targeting aconitase 2 and pyruvate dehydrogenase. Nat Commun 11:698. https://doi.org/10.1038/s41467-020-14433-7
Tawakol A, Singh P, Mojena M et al (2015) HIF-1α and PFKFB3 mediate a tight relationship between proinflammatory activation and anerobic metabolism in atherosclerotic macrophages. Arterioscler Thromb Vasc Biol 35:1463–1471. https://doi.org/10.1161/ATVBAHA.115.305551
Biswas SK, Mantovani A (2012) Orchestration of metabolism by macrophages. Cell Metab 15:432–437. https://doi.org/10.1016/j.cmet.2011.11.013
Blagih J, Jones RG (2012) Polarizing macrophages through reprogramming of glucose metabolism. Cell Metab 15:793–795. https://doi.org/10.1016/j.cmet.2012.05.008
Ruytinx P, Proost P, Van Damme J, Struyf S (2018) Chemokine-Induced Macrophage Polarization in Inflammatory Conditions. Front Immunol 9:1930. https://doi.org/10.3389/fimmu.2018.01930
Zhu L, Yang T, Li L et al (2014) TSC1 controls macrophage polarization to prevent inflammatory disease. Nat Commun 5:4696. https://doi.org/10.1038/ncomms5696
Povo-Retana A, Mojena M, Boscá A et al (2021) Graphene Particles Interfere with Pro-Inflammatory Polarization of Human Macrophages: Functional and Electrophysiological Evidence. Adv Biol 5:2100882. https://doi.org/10.1002/adbi.202100882
Colin S, Chinetti-Gbaguidi G, Staels B (2014) Macrophage phenotypes in atherosclerosis. Immunol Rev 262:153–166. https://doi.org/10.1111/imr.12218
Dai L, Bhargava P, Stanya KJ et al (2017) Macrophage alternative activation confers protection against lipotoxicity-induced cell death. Mol Metab 6:1186–1197. https://doi.org/10.1016/j.molmet.2017.08.001
Funes SC, Rios M, Escobar-Vera J, Kalergis AM (2018) Implications of macrophage polarization in autoimmunity. Immunology 154:186–195. https://doi.org/10.1111/imm.12910
Galván-Peña S, O’Neill LAJ (2014) Metabolic reprogramming in macrophage polarization. Front Immunol 5:1–6. https://doi.org/10.3389/fimmu.2014.00420
Shapouri-Moghaddam A, Mohammadian S, Vazini H et al (2018) Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol 233:6425–6440. https://doi.org/10.1002/jcp.26429
Young MT (2010) P2X receptors: dawn of the post-structure era. Trends Biochem Sci 35:83–90. https://doi.org/10.1016/j.tibs.2009.09.006
Illes P, Müller CE, Jacobson KA et al (2021) Update of P2X receptor properties and their pharmacology: IUPHAR Review 30. Br J Pharmacol 178:489–514. https://doi.org/10.1111/bph.15299
North RA (2002) Molecular physiology of P2X receptors. Physiol Rev 82:1013–1067. https://doi.org/10.1152/physrev.00015.2002
Klaver D, Thurnher M (2021) Control of Macrophage Inflammation by P2Y Purinergic Receptors. Cells 10:1098. https://doi.org/10.3390/cells10051098
Nishimura A, Sunggip C, Oda S et al (2017) Purinergic P2Y receptors: Molecular diversity and implications for treatment of cardiovascular diseases. Pharmacol Ther 180:113–128. https://doi.org/10.1016/j.pharmthera.2017.06.010
Csoka B, Nemeth ZH, Toro G et al (2015) Extracellular ATP protects against sepsis through macrophage P2X7 purinergic receptors by enhancing intracellular bacterial killing. Faseb J 29:3626–3637. https://doi.org/10.1096/fj.15-272450
Baron L, Gombault A, Fanny M et al (2015) The NLRP3 inflammasome is activated by nanoparticles through ATP, ADP and adenosine. Cell Death Dis 6:e1629. https://doi.org/10.1038/cddis.2014.576
Zhou J, Zhou Z, Liu X et al (2021) P2X7 Receptor-Mediated Inflammation in Cardiovascular Disease. Front Pharmacol 12:e654425. https://doi.org/10.3389/fphar.2021.654425
Ferrari D, Pizzirani C, Adinolfi E et al (2006) The P2X7 receptor: a key player in IL-1 processing and release. J Immunol 176:3877–3883
Adinolfi E, Capece M, Franceschini A et al (2015) Accelerated tumor progression in mice lacking the ATP receptor P2X7. Cancer Res 75:635–644. https://doi.org/10.1158/0008-5472.can-14-1259
Zhang Y, Pop IL, Carlson NG, Kishore BK (2011) Genetic deletion of the P2Y2 receptor offers significant resistance to development of lithium-induced polyuria accompanied by alterations in PGE2 signaling. Am J Physiol Ren Physiol 302:F70–F77. https://doi.org/10.1152/ajprenal.00444.2011
Berenbaum F, Humbert L, Bereziat G, Thirion S (2003) Concomitant recruitment of ERK1/2 and p38 MAPK signalling pathway is required for activation of cytoplasmic phospholipase A2 via ATP in articular chondrocytes. J Biol Chem 278:13680–13687. https://doi.org/10.1074/jbc.M211570200
Chiang C-Y, Veckman V, Limmer K, David M (2012) Phospholipase Cγ-2 and Intracellular Calcium Are Required for Lipopolysaccharide-induced Toll-like Receptor 4 (TLR4) Endocytosis and Interferon Regulatory Factor 3 (IRF3) Activation. J Biol Chem 287:3704–3709. https://doi.org/10.1074/jbc.C111.328559
Zhu L, Jones C, Zhang G (2018) The Role of Phospholipase C Signaling in Macrophage-Mediated Inflammatory Response. J Immunol Res 2018:1–9. https://doi.org/10.1155/2018/5201759
Merz J, Nettesheim A, von Garlen S et al (2021) Pro- and anti-inflammatory macrophages express a sub-type specific purinergic receptor profile. Purinergic Signal 17:481–492. https://doi.org/10.1007/s11302-021-09798-3
Giuliani AL, Sarti AC, Di Virgilio F (2021) Ectonucleotidases in Acute and Chronic Inflammation. Front Pharmacol 11:619458. https://doi.org/10.3389/fphar.2020.619458
Deaglio S, Robson SC (2011) Ectonucleotidases as Regulators of Purinergic Signaling in Thrombosis, Inflammation, and Immunity. Adv Pharmacol 11:301–332. https://doi.org/10.1016/B978-0-12-385526-8.00010-2
Degagne E, Grbic DM, Dupuis AA et al (2009) P2Y2 receptor transcription is increased by NF-kappa B and stimulates cyclooxygenase-2 expression and PGE2 released by intestinal epithelial cells. J Immunol 183:4521–4529. https://doi.org/10.4049/jimmunol.0803977
Kunapuli S (2003) Platelet purinergic receptors. Curr Opin Pharmacol 3:175–180. https://doi.org/10.1016/S1471-4892(03)00007-9
Kunapuli SP, Daniel JL (1998) P2 receptor subtypes in the cardiovascular system. Biochem J 336:513–523
Cieślak M, Wojtczak A (2018) Role of purinergic receptors in the Alzheimer’s disease. Purinergic Signal 14:331–344. https://doi.org/10.1007/s11302-018-9629-0
Godoy PA, Ramírez-Molina O, Fuentealba J (2019) Exploring the Role of P2X Receptors in Alzheimer’s Disease. Front Pharmacol 10:1330. https://doi.org/10.3389/fphar.2019.01330
Yiangou Y, Facer P, Durrenberger P et al (2006) COX-2, CB2 and P2X7-immunoreactivities are increased in activated microglial cells/macrophages of multiple sclerosis and amyotrophic lateral sclerosis spinal cord. BMC Neurol 6:12. https://doi.org/10.1186/1471-2377-6-12
Seo DR, Kim SY, Kim KY et al (2008) Cross talk between P2 purinergic receptors modulates extracellular ATP-mediated interleukin-10 production in rat microglial cells. Exp Mol Med 40:19. https://doi.org/10.3858/emm.2008.40.1.19
Horioka M, Ceraudo E, Lorenzen E et al (2021) Purinergic Receptors Crosstalk with CCR5 to Amplify Ca2+ Signaling. Cell Mol Neurobiol 41:1085–1101. https://doi.org/10.1007/s10571-020-01002-1
Chaves MM, Canetti C, Coutinho-Silva R (2016) Crosstalk between purinergic receptors and lipid mediators in leishmaniasis. Parasit Vectors 9:489. https://doi.org/10.1186/s13071-016-1781-1
Erb L, Weisman GA (2012) Coupling of P2Y receptors to G proteins and other signaling pathways. Wiley Interdiscip Rev Membr Transp Signal 1:789–803. https://doi.org/10.1002/wmts.62
Brambilla R, Burnstock G, Bonazzi A et al (1999) Cyclo-oxygenase-2 mediates P2Y receptor-induced reactive astrogliosis. Br J Pharmacol 126:563–567. https://doi.org/10.1038/sj.bjp.0702333
Xing M, Post S, Ostrom RS et al (1999) Inhibition of phospholipase A2-mediated arachidonic acid release by cyclic AMP defines a negative feedback loop for P2Y receptor activation in Madin-Darby canine kidney D1 cells. J Biol Chem 274:10035–10038
Park J-E, Kim Y-I, Yi A-K (2008) Protein Kinase D1: A New Component in TLR9 Signaling. J Immunol 181:2044–2055. https://doi.org/10.4049/jimmunol.181.3.2044
Park J-EE, Kim Y-II, Yi A-KK (2009) Protein Kinase D1 Is Essential for MyD88-Dependent TLR Signaling Pathway. J Immunol 182:6316–6327. https://doi.org/10.4049/jimmunol.0804239
Kolczynska K, Loza-Valdes A, Hawro I, Sumara G (2020) Diacylglycerol-evoked activation of PKC and PKD isoforms in regulation of glucose and lipid metabolism: a review. Lipids Health Dis 19:113. https://doi.org/10.1186/s12944-020-01286-8
Firestein BL, Xing M, Hughes RJ et al (1996) Heterogeneity of P2u- and P2y-purinergic receptor regulation of phospholipases in MDCK cells. Am J Physiol Physiol 271:F610–F618. https://doi.org/10.1152/ajprenal.1996.271.3.F610
Corriden R, Insel PA (2012) New insights regarding the regulation of chemotaxis by nucleotides, adenosine, and their receptors. Purinergic Signal 8:587–598. https://doi.org/10.1007/s11302-012-9311-x
Ledderose C, Liu K, Kondo Y et al (2018) Purinergic P2X4 receptors and mitochondrial ATP production regulate T cell migration. J Clin Invest 128:3583–3594. https://doi.org/10.1172/JCI120972
Lee BH, Hwang DM, Palaniyar N et al (2012) Activation of P2X(7) Receptor by ATP Plays an Important Role in Regulating Inflammatory Responses during Acute Viral Infection. PLoS One 7:e35812. https://doi.org/10.1371/journal.pone.0035812
Wu L-J, Vadakkan KI, Zhuo M (2007) ATP-induced chemotaxis of microglial processes requires P2Y receptor-activated initiation of outward potassium currents. Glia 55:810–821. https://doi.org/10.1002/glia.20500
O’Grady SM (2012) Purinergic signaling and immune cell chemotaxis. Focus on “The UDP-sugar-sensing P2Y 14 receptor promotes Rho-mediated signaling and chemotaxis in human neutrophils.” Am J Physiol Physiol 303:C486–C487. https://doi.org/10.1152/ajpcell.00184.2012
Koizumi S, Shigemoto-Mogami Y, Nasu-Tada K et al (2007) UDP acting at P2Y6 receptors is a mediator of microglial phagocytosis. Nature 446:1091–1095. https://doi.org/10.1038/nature05704
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work has been supported by: Ministerio de Ciencia, Investigación y Universidades, Agencia Estatal de Investigación https://doi.org/10.13039/501100011033 (PID2020-113238RB-I00 and PID2019-108977RB-I00), Comunidad de Madrid Programa Biociencias (P2022-BMD-7223), and Centro de Investigación Biomédica en Red en Enfermedades Cardiovasculares (CB16/11/00222) from the Instituto de Salud Carlos III (co-financed by the European Development Regional Fund “A Way to Achieve Europe”).
Author information
Authors and Affiliations
Contributions
A.P-R. and L.B. conceived, researched, wrote, provided funding, discussed with all authors and prepared the final version; C.D. and P.M-S. provided intellectual input and contributed to the analysis of prostanoids signaling; S.S-G., C.A-L., R.L-V. and P.P. provided intellectual input and revised the manuscript.
Corresponding authors
Ethics declarations
Ethical approval
This is a review article of already published work and does not include unpublished studies on either animals or humans.
Informed consent
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Povo-Retana, A., Sánchez-García, S., Alvarez-Lucena, C. et al. Crosstalk between P2Y receptors and cyclooxygenase activity in inflammation and tissue repair. Purinergic Signalling 20, 145–155 (2024). https://doi.org/10.1007/s11302-023-09938-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11302-023-09938-x