
Abstract
Synchronisation of fruiting phenology with disperser activities is an important component of plant adaptation to seed dispersal mutualisms. Although most previous studies have exclusively evaluated the effects of fruiting phenology on seed removal rate, seed dispersal effectiveness is determined by both the number of dispersed seeds (quantitative effectiveness) and the probability of a dispersed seed becomes an adults (qualitative effectiveness). Therefore, the adaptive significance of fruiting phenology should be assessed for both components of dispersal effectiveness. This study investigates the adaptive significance of fruiting phenology for two ant-dispersed sedges, Carex lanceolata and C. tristachya. Because C. lanceolata is known to require a longer seed dispersal distance than C. tristachya, we hypothesized that C. lanceolata produces seeds when high qualitative dispersal effectiveness can be achieved, while C. tristachya bears seeds when seed disperser ants are abundant to give priority to quantitative dispersal effectiveness. Field observations of ant activity around the sedges revealed that the relative abundance of ant species with the longest seed dispersal distance, Formica japonica, was higher during the fruiting period of C. lanceolata than in that of C. tristachya, which is consistent with the requirement of C. lanceolata to achieve high qualitative effectiveness. In addition, our fruiting manipulation experiment revealed that C. tristachya fruits when the overall seed removal rate is high, resulting in high quantitative effectiveness. Overall, our results support the idea that variations in both qualitative and quantitative dispersal effectiveness should be considered to understand the adaptive significance of fruiting phenology in animal-dispersed plants.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The seed dispersal mutualism is a key interspecific interaction shaping plant regeneration worldwide (Howe and Smallwood 1982; Hanzawa et al. 1988; Herrera 2002; Lengyel et al. 2010). To maximise benefits from the seed dispersal mutualism, plants have evolved various traits including fruit size (Guimarães et al. 2008; Galetti et al. 2013), fruit colour (Lomáscolo and Schaefer 2010; Amico et al. 2011) and chemical composition (Hughes et al. 1994; Fischer et al. 2008; Whitehead et al. 2013) of rewards, and fruiting phenology synchronised with the seasonal abundance of seed dispersers (Hampe and Bairlein 2000; Oberrath and Böhning-Gaese 2002; Aizen 2003). Understanding plant adaptation to seed dispersal mutualism is crucial to predict future changes in plant communities in response to current environmental shifts (Galetti et al. 2013).
Because both abundance and species composition of disperser animals can vary seasonally (Hampe and Bairlein 2000; Oberrath and Böhning-Gaese 2002; Aizen 2003), the synchronisation of fruiting phenology with disperser activity is one of the most important dispersal adaptations in plants (Hampe and Bairlein 2000; Oberrath and Böhning-Gaese 2002; Aizen 2003). Not surprisingly, in many plants, fruiting periods coincide with periods when disperser activities are high, implying adaptations of plants to the seasonality of dispersers (Hampe and Bairlein 2000; Oberrath and Böhning-Gaese 2002; Aizen 2003). In contrast, some researchers deny the phenological adaptation of plants to seed dispersal, and they explain the phenomenon using phylogenetic constraints or adaptation to other factors, such as pollination, growth-form, and habitat (Herrera 2002). Nevertheless, in the case of ant-dispersed plants, detailed analyses controlling for both other factors demonstrated that the plant-disperser synchronisation has evolved because of the adaptation of plants to disperser seasonality (Oberrath and Böhning-Gaese 2002).
Previous studies examining the effects of fruiting phenology on seed dispersal success have focused on the number of dispersed seeds alone (Willson and Whelan 1993; Oberrath and Böhning-Gaese 2002; Aizen 2003). However, seed dispersal effectiveness, sensu Schupp et al. (2010), is determined not only by the number of dispersed seeds (i.e., quantitative effectiveness), but also by the probability of dispersed seeds to become a reproductive adult (i.e., qualitative effectiveness) (Schupp et al. 2010). Because qualitative effectiveness largely varies with the composition of the disperser assemblage (Hughes and Westoby 1992; Wenny and Levey 1998; Ness et al. 2004), animal-dispersed plants may not synchronise with the activities of a whole seed disperser assemblage, but specifically with the active season of favourable dispersers (Giladi 2006). Such a trait-based disperser specificity has already been demonstrated for other traits including elaiosome:seed size ratio (Leal et al. 2014) and seed maturation timing within a day (Boulay et al. 2007; Aranda-Rickert and Fracchia 2012). Moreover, Manzaneda and Rey (2009) revealed that each local population of a myrmecochore Helleborus foetidus specializes on one or a few ant species. All these studies highlight the species-specific nature of myrmecochory, as recently emphasized (Manzaneda and Rey 2009; Ness et al. 2009; Manzaneda and Rey 2012). Nevertheless, only one study by Warren et al. (2011) examined the correspondence of fruiting period with the activity of a specific disperser guild, and showed that some plant species synchronised with favourable dispersers but others did not. Warren et al. (2011) suggested that the phenological asynchrony is the maladaptive consequence of different responses to global warming between plants and ants. However, it may alternately indicate that the relative importance of quantitative and qualitative dispersal effectiveness varies among sympatric plants. Specifically, for some species, qualitative dispersal effectiveness has a stronger impact than quantitative effectiveness on overall seed dispersal effectiveness, favouring traits that increase dispersal by high-quality dispersers. For other plant species, in contrast, quantitative effectiveness has a stronger impact than qualitative effectiveness on overall seed dispersal effectiveness. For these plant species the disperser species do not vary greatly in the quality they provide. As a result, traits maximizing the number of seeds dispersed regardless of disperser species will be favoured. To explore this possibility, the relative importance between quantitative and qualitative dispersal effectiveness (i.e., dispersal requirements) should be elucidated to understand the phenological strategy of animal-dispersed plants.
This study focused on two perennial ant-dispersed sedges, Carex lanceolata Boott and C. tristachya Thunb (Cyperaceae), and examined their phenological specialisations in relation to dispersal requirements. These sedges are sympatrically distributed and both have elaiosomes facilitating seed dispersal by ants (Tanaka and Suzuki 2016; Tanaka and Tokuda 2016). However, the two sedges have different seed traits: seeds of C. lanceolata have larger elaiosomes and are more frequently dispersed by a high-quality ant species Formica japonica for a longer distance than those of C. tristachya (Tanaka and Tokuda 2016). In contrast, seeds of C. tristachya are frequently dispersed by ants such as Pheidole noda and Crematogaster osakensis, which remove the seeds but transport them for much shorter distances than F. japonica (Tanaka and Tokuda 2016). Our previous study also revealed that C. lanceolata adults suppressed the growth and reproduction of adjacent conspecific seedlings, but C. tristachya did not, which strongly suggests that C. lanceolata attracts the high-quality ant to enhance the probability of seedling isolation from conspecific adults (Tanaka and Tokuda 2016). Although seeds of C. lanceolata are highly attractive to F. japonica, they also attract low-quality ants such as P. noda and C. osakensis (Tanaka and Tokuda 2016). Notably, F. japonica is behaviourally subordinate to low-quality ants in the field (Tanaka and Suzuki 2016), so seeds discovered and recruited to earlier ants other than F. japonica generally become unavailable to F. japonica. Given these characteristics of the dispersal system, we hypothesised that C. lanceolata produces seeds when the relative abundance of F. japonica is high to achieve a greater qualitative dispersal effectiveness, and C. tristachya bears seeds when the overall assemblage of seed disperser ants is abundant to give priority to quantitative dispersal effectiveness. To test the seasonal change in qualitative dispersal effectiveness, we investigated seasonal changes in the abundance of four main disperser ant species and abiotic factors that potentially affect them. Then, we manipulated the fruiting phenology to test seasonal changes in the seed removal success, which is an indicator of quantitative dispersal effectiveness.
Materials and methods
Study organisms
Carex lanceolata is distributed widely in northeastern Asia, while C. tristachya is endemic to Japan (Katsuyama 2005). Both species grow in forests, grasslands, and roadsides. Seeds of both sedges are dispersed by two steps, i.e., gravity followed by ants.
Study site
The study area is an approximately 1.5 km trail on Mount Kinryu in Saga Prefecture, Kyushu, Japan (33°34′N, 130°31′E, 60–220 m a.s.l.) composed of a mosaic of broadleaved evergreen forests, Japanese cedar Cryptomeria japonica (L. f.) D. Don (Taxodiaceae) plantations, and open sites dominated by ericaceous shrubs and the large tussock grass Miscanthus sinensis Andersson (Poaceae). Based on the nearest weather station, in Saga City, the monthly mean air temperature during 1981–2010 ranged from 5.4 °C (January) to 27.8 °C (August). Annual rainfall is approximately 1900 mm, with the highest rainfall occurring in June or July (http://www.data.jma.go.jp/obd/stats/etrn/index.php).
Seasonal abundance of disperser ants
In the study area, 30 census plots (each 30 m long and 2 m wide) were established continuously along the mountain trail. Single reproductive sedge was marked randomly, but apart from other census individuals at least 5 m, in every plot in early March 2011. Because of the presence of some plots in which no individuals of focal sedge were found, the total numbers of marked individuals were 29 for C. lanceolata and 28 for C. tristachya, respectively. Ant worker density on the ground was used as an indicator of disperser abundance from 2 May 2011 when the marked sedges began fruiting, to 14 June 2011, when all marked sedges had shed their seeds. Censuses were conducted from 0900 (morning) to 1630 (dusk) on 4–5 sunny days every week. On each census day, marked individuals of each sedge species were randomly visited and ant activity was recorded for 5 min. Following the methods of previous studies (Orivel and Dejean 1999; Miura et al. 2003), slide glasses were used to assess ant activity. For each census, a slide glass (75 mm × 25 mm × 1.5 mm) was placed under an infructescence of a marked individual, and the number and species of ants that crossed the slide glass were recorded for 5 min. The presence of mature seeds on marked sedges was also recorded to confirm the fruiting periods of the two sedge species. To examine the level of yearly fluctuations in fruiting phenology, the census of fruiting period was also conducted in 2012.
To compare day and night trends of seasonal ant abundance, we also surveyed ant activity on 3–7 sunny days/week between 9 and 31 May 2013, using six individuals of each sedge species randomly selected from those marked in 2011. Diurnal and nocturnal censuses were conducted from 1100 to 1300 and from 2300 to 0100 (of the next day), respectively. To clarify the environmental correlates of seasonal ant abundance, temperature, relative humidity, and light intensity were also measured at approximately 5 cm above the slide glass using thermo-recorders (Ondotori, T and D Co., Nagano, Japan).
Seasonal ant abundance during the diurnal censuses were analysed for four major ant species, F. japonica, P. noda, C. osakensis, and Paratrechina flavipes, which account for over 90% of ants interacting with seeds of these sedges and whose seed dispersal distances were already studied in the census site (Tanaka and Tokuda 2016). Before the analysis, the visitation frequency of each ant species was summed for an early period (1 May–19 May) and a late period (20 May–14 June), respectively. The early and late periods coincide with the peak fruiting periods of C. lanceolata and C. tristachya, respectively (see “Results”). The relative abundance of F. japonica (=proportion of visitation frequency by F. japonica of the total visitation frequencies of all four major ant species) was compared between the periods using GLMM, assuming binomial errors, with the package “lme4” in R (R Core Team 2015). Furthermore, seasonal patterns in visitation frequencies of respective ant species (=visitation frequency/the number of census in each period) were analysed with LMM to interpret the seasonal patterns of F. japonica. The response variable was log-transformed before analysis to achieve normality. After calculating the median value for each period, temporal changes in environmental variables were analysed with LMM.
Effects of fruiting periods on the seed removal rate
To analyse the consequences of variation in fruiting phenology on seed removal success in the two sedges, we manipulated the fruiting periods of each sedge species. From December 2011 to January 2012, eight and ten large individuals of C. lanceolata and C. tristachya, respectively, were collected along with the surrounding soil from areas within 5 km of the study site. They were transplanted into pots made of nonwoven textile fabric (35 cm diameter and 30 cm deep), and a mixture of 50% tuff loam and 50% humus was added when the surrounding soil was insufficient to fill the pot. The pots were placed in a garden on the Saga University campus and were watered every other day until the experiment.
The fruiting period of C. tristachya was advanced by warming ten preflowering individuals in a greenhouse at Saga University in March 2012, where the air temperature was maintained at a maximum of 30–35 °C during daytime and a minimum of 15–20 °C at night. The fruiting period of C. lanceolata was delayed by maintaining eight postflowering individuals at 15 °C (12L/12D) in Biotron units (NK System, Osaka) from late April to middle May 2012. After that, temperature was increased to 25 °C (12L/12D) to facilitate seed maturation. In addition, seeds were collected for the experiment from sedge individuals growing at the study site. Seeds obtained from manipulated plants did not differ in appearance from natural seeds and similarly attracted ants in preliminary observations.
Seed removal rates were estimated weekly from 3 to 23 May 2012 as follows. Twenty marked individuals of each sedge species were randomly selected and divided into two groups of ten individuals. One group was surveyed on 3 and 16 and the other on 9 and 23 May (because of the shortage of available fresh seeds, C. lanceolata was not examined on 23 May). On each census day, 20 seeds of the focal sedge species were placed on a depot near each marked individual. The depot consisted of an inverted petri dish lid (9 cm diameter and 5 mm high) covered with a polyester tea bag and fixed to the ground with marking pins. The numbers of seeds remaining on the depots were recorded after 150 min, 300 min, and 24 h. Because non-ant species seldom remove the sedge seeds (<3%, KT, unpublished data), we considered all seeds removed from the depot during the census to have been taken by ants. Because nearly all seeds were removed within 300 min on all dates (see “Results”), the number of removed seeds recorded within the first 150 min (i.e., seed removal rate/150 min) was analysed using GLMM assuming binomial errors. Date of the experiment was the fixed factor while sedge individual was the random effect.
Results
Seasonal abundance of disperser ants
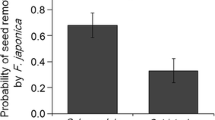
In 2011, C. lanceolata fruiting was concentrated mostly during early May (median Julian date: 127), while C. tristachya fruited mainly during late May (median Julian date: 150) extending to middle June. The fruiting dates of 2012 were very similar to those of 2011; median Julian dates of 128 and 143 for C. lanceolata and C. tristachya, respectively. The fruiting dates were significantly different between plant species for both years (P < 0.0001). The relative abundance of F. japonica was significantly lower in the late period than in the early period in both census years (Fig. 1, 2011: χ2 = 47.9, df = 1, P < 0.01, 2013: χ2 = 9.1, df = 1, P < 0.01), with the trend stronger in 2013 than in 2011 (period × year: χ2 = 5.2, df = 1, P = 0.02). Interactive effects of plant species with period were not detected (2011: χ2 = 1.5, df = 1, P = 0.23, 2013: χ2 = 0.87, df = 1, P = 0.35), indicating that seasonal patterns in the relative abundance of F. japonica did not differ between C. lanceolata and C. tristachya. Analyses of individual ant species indicated that the decrease in the relative abundance of F. japonica was due to the different seasonal patterns of F. japonica, P. noda, and C. osakensis. Specifically, F. japonica did not show significant seasonal changes in abundance (Fig. 2, χ2 = 2.7, df = 1, P = 0.10), while P. noda and C. osakensis increased from early to late periods (Fig. 2, P. noda: χ2 = 24.3, df = 1, P < 0.01, C. osakensis: χ2 = 30.6, df = 1, P < 0.01). These trends were consistent across years and plant species as indicated by the non-significant interactions between fruiting period and the latter two variables. Based on the measurement in 2013, light intensity on the ground decreased from early to late May (Fig. 3; χ2 = 4.74, df = 1, P = 0.03), while relative humidity increased during the same period (Fig. 3; χ2 = 16.90, df = 1, P < 0.01). Temperature showed no significant differences between periods (Fig. 3; χ2 = 0.001, df = 1, P = 0.97). In the 2013 nocturnal census, F. japonica was never observed, indicating that this species is strictly diurnal, while the other three ant species were active at night. In P. noda and C. osakensis, changes in the nocturnal abundance were not detected (P. noda: χ2 = 0.77, df = 1, P = 0.38, C. osakensis: χ2 = 0.56, df = 1, P = 0.46), indicating that the increase in the daytime abundance cannot be attributed to shift in daily activity schedules (i.e., shift from nocturnal to diurnal).

Relative abundances of Formica japonica by census period in 2011 (a) and 2013 (b), respectively. The y-axis shows the percent of visitation frequency by F. japonica of the four main disperser ants (F. japonica, Pheidole noda, Crematogaster osakensis, Paratrechina flavipes). “Early” and “Late” denotes the census periods 1 May to 19 May and 20 May to 14 June, respectively

Absolute abundances of the four main ants by census period in 2011 (a–d) and 2013 (e–h), respectively. The y-axis shows the visitation frequency by focal ant species per 5 min. Ant species are; a, e Formica japonica, b, f Pheidole noda, c, g Paratrechina flavipes, and d, h Crematogaster osakensis

Environmental variables measured at approximately 5 cm above the ground surface in the 2013 daytime census. Measured variables are; a light intensity (lx), b air temperature (°C), and c relative humidity (%)
Effects of fruiting periods on the seed removal rate
In 2013, only C. lanceolata fruited on the first experimental date (3 May), while only C. tristachya fruited on the other two experimental dates (16 and 23 May), indicating that the experimental period cover the transitional season between fruitings of C. lanceolata and of C. tristachya. All seeds were removed within 300 min regardless of the date or sedge species. The only exception was one case where a single seed remained after 300 min, but was removed within 24 h. The number of seeds removed within 150 min significantly increased over the season in both sedges (Fig. 4, C. lanceolata: χ2 = 118.11, df = 2, P < 0.01, C. tristachya: χ2 = 60.95, df = 3, P < 0.01).

Seed removal rate by date in Carex lanceolata (a) and Carex tristachya (b), respectively. The x-axis shows the number of seeds removed within the first 150 min of the experiment
Discussion
The relative abundance of F. japonica was greater in the fruiting season of C. lanceolata (early May) than in that of C. tristachya (later May to early June). Thus, the fruiting phenology of C. lanceolata facilitates seed removal by F. japonica. On the other hand, C. tristachya fruited when the relative abundance of F. japonica was low but the overall seed removal rate by the total assemblage of disperser ants was high. An alternative possibility for the association between fruiting of C. lanceolata and high relative abundance of F. japonica is that F. japonica increased their activity in response to the fruiting of their preferred foods (i.e. C. lanceolata). However, this is unlikely to be the case, because we found no significant changes in the absolute abundance of F. japonica through seasons and its high relative abundance in early May was actually caused by the relatively low frequencies of other ants. Thus, the current results support our hypotheses that both species have adaptive fruiting phenology in relation to requirements for seed dispersal: i.e., C. lanceolata promotes seed removal by F. japonica, the ant species with the longest seed dispersal distances, to increase qualitative dispersal effectiveness, while C. tristachya gives priority to removal rates rather than dispersal distance to ensure quantitative dispersal effectiveness. In addition, C. lanceolata is known to have larger elaiosomes than C. tristachya and they are highly attractive to F. japonica (Tanaka and Tokuda 2016), which supports our hypotheses.
The increase in the activities of two ant species, P. noda and C. osakensis, was associated with an increase in relative humidity and a decrease in light intensity. This suggests that the difference in seasonal abundance might be attributed to ant species-specific tolerances to drought and/or solar radiation, as shown by previous studies conducted in other regions (Briese and Macauley 1980; Cros et al. 1997). In general, subordinate species have wider environmental tolerances than dominants, possibly to avoid harmful interactions with dominants through temporal niche partitions (Briese and Macauley 1980; Cros et al. 1997; Aranda-Rickert and Fracchia 2012). Our results are consistent with this, because ant species with relatively low interference competitive ability, i.e., F. japonica and P. flavipes (Tanaka and Suzuki 2016), did not show significant temporal patterns. Thus, interspecific competition among ants can lead to the temporal niche partitions, which ultimately enable the phenological specialisations in ant-dispersed plants.
Different degrees of synchronisation with specific disperser guilds have already been reported in North American ant-dispersed plant species (Warren et al. 2011). Such variations in synchrony may be the result of the failure in some plant species to synchronise with dispersers due to climatic changes following global warming (Warren et al. 2011). However, because selective pressures acting on dispersal can vary with different factors, such as spatiotemporal environmental heterogeneity (Green 1983; North et al. 2011) and the intensity of intraspecific competition (North et al. 2011), the variations may alternately reflect different dispersal requirements among plants. In our study, fruiting seasons of the two sympatric sedge species were different even though they share the same disperser guild, while, as mentioned earlier, the fruiting phenology is considered to be adaptive for both species.
Seed dispersal effectiveness is determined by the multiplication of the number of seeds dispersed (quantity component) and the probability that a dispersed seed produces a new adult (quality component) (Schupp et al. 2010). However, most previous studies have only measured the quantitative effectiveness when evaluating the selection pressure for seed dispersal traits (Willson and Whelan 1993; Oberrath and Böhning-Gaese 2002; Galetti et al. 2013), which might lead to biased results. The consequences of the phenological differences between C. lanceolata and C. tristachya for dispersal effectiveness would not have been detected in our study if the temporal changes in disperser species compositions, the correlate of qualitative dispersal effectiveness, were neglected. The importance of considering qualitative dispersal effectiveness is in line with recent findings on myrmecochory that show the disproportionate attraction of high-quality ants by virtue of large diaspore (Takahashi and Itino 2012) or elaiosome size (Leal et al. 2014), and the seed maturation timing within a day (Boulay et al. 2007). Together with these previous findings, our results highlight the importance of considering both the quantitative and qualitative components of dispersal effectiveness when assessing the fitness consequences of seed traits. They also suggest that the negative consequence of global warming on phenological synchronisation was underestimated in studies that focused on quantitative dispersal effectiveness alone.
An unclarified point of our study is actual seed removal frequencies by respective ant species. Several studies on invasive ants have found a seasonal shift in dietary preference (Stein et al. 1990; Abbott et al. 2014), possibly reflecting the temporal changes in dietary needs of colonies (Stein et al. 1990). If the preference for sedge seeds shows the opposite seasonal pattern with abundance, our conclusion based solely on the abundance of ants might be affected. However, we consider that this phenomenon does not apply in our case, because seed removal probability of F. japonica upon discovery is already high (9 in 14) in early May (Tanaka and Tokuda 2016), indicating that both seed attractiveness to and relative abundance of F. japonica is high in early May. Similarly, seed removal probability by low-quality ants is quite high in the later period (Tanaka and Tokuda 2016), also showing the coincidence between seed attractiveness and relative abundance. Although precise quantification of the seasonal change in qualitative seed dispersal effectiveness needs additional data on actual seed removal, our conclusion, i.e., qualitative dispersal effectiveness decreases from May to June, will be stable.
Our study supports the idea that plants in which qualitative dispersal effectiveness is the primary determinant of overall seed dispersal effectiveness have fruiting phenologies which are more specialised on particularly effective disperser guilds. Although assessing the generality of our conclusion awaits further studies, recognition that sympatric plant species can differently synchronise with disperser assemblages is important to predict the impact of future climate change on plant communities. For example, plant species requiring synchronization with high-quality dispersers may be more vulnerable to changes in disperser phenology due to global warming than plant species without such requirements. If this is the case, global warming will lead to the increased dominance of generalist plants. To predict the actual consequences of disruption in disperser-plant synchronisation, more knowledge about the functional relationship between fruiting phenology, temporal patterns of dispersal effectiveness, and dispersal requirements of plants is required.
References
Abbott KL, Green PT, O’Dowd DJ (2014) Seasonal shifts in macronutrient preferences in supercolonies of the invasive yellow crazy ant Anoplolepis gracilipes (Smith, 1857) (Hymenoptera: Formicidae) on Christmas Island, Indian ocean. Austral Entomol 53:337–346
Aizen MA (2003) Influences of animal pollination and seed dispersal on winter flowering in a temperate mistletoe. Ecology 84:2613–2627
Amico GC, Rodriguez-Cabal MA, Aizen MA (2011) Geographic variation in fruit colour is associated with contrasting seed disperser assemblages in a south-Andean mistletoe. Ecography 34:318–326
Aranda-Rickert A, Fracchia S (2012) Are subordinate ants the best seed dispersers? Linking dominance hierarchies and seed dispersal ability in myrmecochory interactions. Arthropod Plant Interact 6:297–306
Boulay R, Carro F, Soriguer RC, Cerdá X (2007) Synchrony between fruit maturation and effective dispersers’ foraging activity increases seed protection against seed predators. Proc R Soc B 274:2515–2522
Briese DT, Macauley BJ (1980) Temporal structure of an ant community in semi-arid Australia. Austral J Ecol 5:121–134
Cros S, Cerdá X, Retana J (1997) Spatial and temporal variations in the activity patterns of Mediterranean ant communities. Ecoscience 4:269–278
Fischer RC, Richter A, Hadacek F, Mayer V (2008) Chemical differences between seeds and elaiosomes indicate an adaptation to nutritional needs of ants. Oecologia 155:539–547
Galetti M, Guevara R, Côrtes MC, Fadini R, Matter SV, Leite AB, Labecca F, Ribeiro T, Carvalho CS, Collevatti RG, Pires MM, Guimarães PR Jr, Brancalion PH, Ribeiro MC, Jordano P (2013) Functional extinction of birds drives rapid evolutionary changes in seed size. Science 340:1086–1090
Giladi I (2006) Choosing benefits or partners: a review of the evidence for the evolution of myrmecochory. Oikos 112:481–492
Green DS (1983) The efficacy of dispersal in relation to safe site density. Oecologia 56:356–358
Guimarães PR Jr, Galetti M, Jordano P (2008) Seed dispersal Anachronisms: rethinking the fruits extinct megafauna ate. PLoS One 3:e1745
Hampe A, Bairlein F (2000) Modified dispersal-related traits in disjunct populations of bird-dispersed Frangula alnus (Rhamnaceae): a result of its quaternary distribution shifts? Ecography 23:603–613
Hanzawa FM, Beattie AJ, Culver DC (1988) Directed dispersal: demographic analysis of an ant-seed mutualism. Am Nat 131:1–13
Herrera CM (2002) Seed dispersal by vertebrates. In: Herrera CM, Pellmyr O (eds) Plant-animal interactions. Blackwell, USA, pp 185–208
Howe HF, Smallwood J (1982) Ecology of seed dispersal. Annu Rev Ecol Evol 13:201–228
Hughes L, Westoby M (1992) Fate of seeds adapted for dispersal by ants in Australian sclerophyll vegetation. Ecology 73:1285–1299
Hughes L, Westoby M, Jurado E (1994) Convergence of elaiosomes and insect prey: evidence from ant foraging behaviour and fatty acid composition. Funct Ecol 8:358–365
Katsuyama T (2005) Carex of Japan. Bun-ichi Co., Japan
Leal LC, Neto MCL, de Oliveira AFM, Andersen AN, Leal IR (2014) Myrmecochores can target high-quality disperser ants: variation in elaiosome traits and ant preferences for myrmecochorous Euphorbiaceae in Brazilian Caatinga. Oecologia 174:493–500
Lengyel S, Gove AD, Latimer AM, Majer JD, Dunn RR (2010) Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey. Perspect Plant Ecol Evol Syst 12:43–55
Lomáscolo SB, Schaefer HM (2010) Signal convergence in fruits: a result of selection by frugivores? J Evol Biol 23:614–624
Manzaneda AJ, Rey PJ (2009) Assessing ecological specialization of an ant–seed dispersal mutualism through a wide geographic range. Ecology 90:3009–3022
Manzaneda AJ, Rey PJ (2012) Geographical and interspecific variation and the nutrient-enrichment hypothesis as an adaptive advantage of myrmecochory. Ecography 35:322–332
Miura R, Doi M, Yoshino M (2003) Distribution and myrmecochory of Veronica polita subsp. lilacina observed in the area around Kyoto University. J Weed Sci Tech 48:140–142
Ness JH, Bronstein AN, Holland JN (2004) Ant body size predicts dispersal distance of ant-adapted seeds: implications of small-ant invasions. Ecology 85:1244–1250
Ness JH, Morin DF, Giladi I (2009) Uncommon specialization in a mutualism between a temperate herbaceous plant guild and an ant: are Aphaenogaster ants keystone mutualists? Oikos 118:1793–1804
North A, Cornell S, Ovaskainen O (2011) Evolutionary responses of dispersal distance to landscape structure and habitat loss. Evolution 65:1739–1751
Oberrath R, Böhning-Gaese K (2002) Phenological adaptation of ant-dispersed plants to seasonal variation in ant activity. Ecology 83:1412–1420
Orivel J, Dejean A (1999) Selection of epiphyte seeds by ant-garden ants. Ecoscience 6:51–55
R Core Team (2015) R: a language and environment for statistical computing, version 3.2.3. R Foundation for Statistical Computing, Vienna, http://www.R-project.org/
Schupp EW, Jordano P, Gómez JM (2010) Seed dispersal effectiveness revisited: a conceptual review. New Phytol 188:333–353
Stein MB, Thorvilson HG, Johnson JW (1990) Seasonal changes in bait preference by red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae). Florida Entom 73:117–123
Takahashi S, Itino T (2012) Larger seeds are dispersed farther: the long-distance seed disperser ant Aphaenogaster famelica prefers larger seeds. Sociobiology 59:1401–1411
Tanaka K, Suzuki N (2016) Interference competition among disperser ants affects their preference for seeds of an ant-dispersed sedge Carex tristachya (Cyperaceae). Plant Species Biol 31:11–18
Tanaka K, Tokuda M (2016) Seed dispersal distances by ant partners reflect preferential recruitment patterns in two ant-dispersed sedges. Evol Ecol 30:943–952
Warren RJ II, Bahn V, Bradford MA (2011) Temperature cues phenological synchrony in ant-mediated seed dispersal. Global Change Biol 17:2444–2454
Wenny DG, Levey DJ (1998) Directed seed dispersal by bellbirds in a tropical cloud forest. Proc Nat Acad Sci 95:6204–6207
Whitehead SR, Jeffrey CS, Leonard MD, Dodson CD, Dyer LA, Bowers MD (2013) Patterns of secondary metabolite allocation to fruits and seeds in Piper reticulatum. J Chem Ecol 39:1373–1384
Willson MF, Whelan CJ (1993) Variation of dispersal phenology in a bird-dispersed shrub, Cornus drummondii. Ecol Monogr 63:151–172
Acknowledgements
We thank Hidezumi Ohashi for his help in field observations.
Author information
Authors and Affiliations
Contributions
KT and MT conceived the ideas and designed methodology. KT collected and analysed the data. KT and MT wrote the manuscript. Both authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
All authors do not have any conflict of interest.
About this article

Cite this article
Tanaka, K., Tokuda, M. Phenological specialisation of two ant-dispersed sedges in relation to requirements for qualitative and quantitative dispersal effectiveness. Ecol Res 32, 677–684 (2017). https://doi.org/10.1007/s11284-017-1479-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-017-1479-7