
Abstract
Invasive alien plant species (IAPS) are severely changing ecosystems on earth. Studying the interactions that allow IAPS to establish and spread in the new regions is crucial. Ants can disperse exotic fleshy fruits. We asked the following questions at three different sites of Chaco Mountain Forest (Córdoba, Argentina): (1) Do ants disperse diaspores of native, neonative and IAPS differentially? (2) Which is the ant species assemblage and their role in the secondary dispersal of each of the selected plant species? (3) Do ants interact in different ways with intact and manipulated fruits, and these interactions vary within plant species origin? and (4) Are diaspore traits different among the plant species considered? We selected four plant species: Celtis ehrenbergiana (native), Lantana camara (neonative), Pyracantha angustifolia and Ligustrum lucidum (IAPS). Two experiments were performed: (1) To disentangle the contribution of ants to the secondary dispersal process, and (2) To investigate the ant behavior of ground-foraging ant species when they encountered the fruits. Additionally, we measured fruit mass and the number of seeds per fruit. Ants were the main diaspore dispersers on the Chaco Mountain Forest ground. Twelve ant species interacted with the fruits; the native presented the higher number, followed by the neonative, and the two IAPS. Only Acromyrmex crassispinus and Pheidole cordyceps removed diaspores. Furthermore, the fruits differed in their mass and also in the number of seeds. Our results highlight the importance of ants and also diaspores traits in these diffuse mutualisms, and enhance their role in plant-invasive processes in subtropical ecosystems.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Humans have been modifying nature since they appeared on earth (Diamond and Ordunio 1999). However, in recent decades this process has been accelerated and the distribution of several species from different taxa worldwide has been reshaped (Sala et al. 2000; Corlett 2015). In their new distribution areas, species of some plant taxa are capable of establishing and spreading along the new landscapes (Richardson et al. 2000b). The invasive process is complex and encounters several filters at different temporal and spatial scales that the plant species have to surpass. These filters include abiotic and biotic interactions in their new habitats. For plants depending on animal vectors for pollination and dispersal, mutualisms are key to facilitate the invasive process (Richardson et al. 2000a, b; Dietz and Edwards 2006; Blackburn et al. 2011; Traveset and Richardson 2014). Nevertheless, invasive alien plant species (hereafter IAPS) are distributed along diverse regions of the world (Rejmánek and Richardson 2013; Seebens et al. 2017; Turbelin et al. 2017), causing different kinds of problems in the new ecosystems they invade (Vilà et al. 2011; Diagne et al. 2021).
Most of the research previously performed on invasive species has focused on negative interactions (e.g., Elton 1958), but it has been shown that positive interactions are also relevant for plant establishment and dispersal (e.g., mutualist interactions; Richardson et al. 2000a; Traveset and Richardson 2014). In particular, the dispersal process of diaspores (i.e., the unit of dispersal; Van der Pijl 1982) is a key component for the establishment and the spreading phases of IAPS (Theodarides and Dukes 2007). In the case of plants dispersed by animals, the possibility of successful dispersal depends on the presence of animal vectors in the new distribution areas (Richardson et al. 2000a), mainly native generalist species (Traveset and Richardson 2014). Plant species that have fleshy fruits depend on the presence of frugivorous animals to be dispersed (Cronk and Fuller 1995). In general, most studies have been performed considering the prevailing biotic dispersal syndrome and consequently the main group of organisms involved (Richardson et al. 2000a). However, two-phase dispersal processes also occur involving other secondary dispersal agents, that could also be relevant for the whole dispersal process and the establishment and spreading of IAPS in the new sites (Holmes 1990a, b; Amodeo et al. 2017).
Ants establish several relevant interactions with plants, including primary and secondary diaspore dispersal (Rico-Gray and Oliveira 2007). Myrmechocory involve plants having diaspores with particular seed structures (e.g., elaiosomes) for rewarding ant dispersers (Rico-Gray and Oliveira 2007). Myrmecochorous plants can be primary dispersed by ants, but this group of plants is also composed by diplochorous plant species (Gorb and Gorb 2003; Lengyel et al. 2010). Diplochory is a seed dispersal process with two phases, involving two or more different abiotic/biotic agents (Vander wall and Longland 2004). This two-phase process normally occurs when a primary agent, as vertebrates or processes as ballistic dispersal, remove the diaspore directly from the parent plant. Then, a secondary agent (such as ants) may complement seed dispersal, usually from the ground (Vander wall and Longland 2004). Secondary dispersal by ants involves escape from ground predation and the possibility to reach safe sites that ensure germination and establishment (Vander wall and Longland 2004). They can also perform a process known as directed dispersal, directly conducting seeds to safe sites (Schupp 1995; Richardson et al. 2000a). Small ant species can also benefit plants by removing fruit pulp, impeding fungus colonization of fruits and enhancing seed survival and germination (Oliveira et al. 1995; Leal and Oliveira 1998; Ohkawara and Akino 2005). Furthermore, previous evidence has shown that some diaspore traits (e.g., diaspore mass and seed number per diaspore) are relevant for secondary dispersal by ants. For instance, small-sized diaspores have higher chances of being removed in comparison to medium and large-sized diaspores (Pizo and Oliveira 2001; Anjos et al. 2020), while seed number can be relevant for seed dispersal effectiveness (Schupp et al. 2010). In addition, there is evidence showing that the condition of the fruit on the ground can influence subsequent interaction with ants, especially previous handling by avian frugivores (in primarily bird-dispersed species), enhancing subsequent secondary dispersal (Bieber et al. 2013; Guerra et al. 2018).
Recent evidence shows that highly invasive species are more dependent on mutualisms than previously expected (Moyano et al. 2020). It is very important to consider the relevance of diffuse mutualisms (i.e., a focal species depends on multiple agents) (Janzen 1985; Richardson et al. 2000a), such as seed-dispersal mutualisms. For instance, native ants in the genera Anoplolepis and Pheidole disperse seeds of the IAPS Acacia cyclops A. Cunn. ex G. Don and A. saligna (Labill.) H.L. Wendl. in Western Cape (South Africa) at short distances, rendering the diaspores less vulnerable to rodent predation (Holmes 1990a, b). Native and invasive ant species contribute to the spread of Chelidonium majus L. seeds, a fast-growing European perennial herb, introduced into North America (Prior et al. 2014). Ants can also disperse fleshy fruits of IAPS, such as the shrub Murraya paniculata (L.) Jack by Acromyrmex niger in Brazil (Smith, 1858) (Pikart et al. 2011). In Argentina, some studies have shown that native ant species disperse seeds of IAPS (Sanhueza et al. 2008; Pirk and Lopez de Casenave 2017; Ortiz et al. 2021). Despite the wide distribution of the IAPS in some regions, including South America, knowledge of secondary dispersal by ants of fleshy diaspores is incipient and the consequences of the invasive potential of these plant species is not well known (Giorgis and Tecco 2014). The selection of plants with different origins (i.e., native, exotic-invasive, and neonative) would allow us to analyze if the native ant assemblage selects or not diaspores according to the history of the interaction. Neonative designates plants that have recently expanded their range as a consequence of environmental changes generated by human-activities (Essl et al. 2019). Neonative plants have an eco-evolutionary history more similar to native species compared to IAPS, and this could result in a gradient of preferences (i.e., secondary dispersal) between ant and diaspores of plants with different origins.
In the present work we ask the following questions: (1) Do ants disperse diaspores of native, neonative and invasive alien plant species differentially? (2) Which is the ant species assemblage and their role in the secondary dispersal of each of the selected plant species? (3) Do ants interact in different ways with intact and manipulated fruits (i.e., simulating previous manipulation by birds), and these interactions vary within plant species origin? and (4) Are diaspore traits (i.e., diaspore mass and number of seeds per fruit) different among the plant species considered? Considering the history of interaction, we expect a higher number of ants interacting and removing fruits of the native plant species, but this would be constrained by diaspore mass. Regarding the fruit type, we expect a higher number of interactions with previously manipulated fruits independent of plant origin.
Materials and methods
Study site
We performed our study near the localities of Río Ceballos and Salsipuedes in Córdoba Province (central Argentina), during summer months (November 2017-March 2018). We selected three different sites corresponding to the Chaco Mountain Forest phytogeographical region (31° 09′ S and 64° 17′ W; 640–753 m a.s.l.; Cabrera 1976; Oyarzabal et al. 2018). We deliberately selected sites where the neonative species and the two IAPS were already present. In a regional context (i.e., several vegetation units of the Chaco Mountain Forest), the neonative species and also the two IAPS are in high abundance and therefore causing several ecological problems (Hoyos et al. 2010; Ferreras et al. 2015, 2019; Whitworth-Hulse et al. 2020).
The vegetation of the study area is a mosaic of physiognomies, from grasslands and low open shrublands to locally closed forests, with a total cover between 80 and 100%. Tree species such as Lithraea molleoides (Vell.) Engl. (Anacardiaceae), Condalia buxifolia Reissek (Rhamnaceae) and Zanthoxylum coco Gillies ex Hook. f. & Arn. (Rutaceae) together with some shrubs and vines are among the dominant species (Giorgis et al. 2017; Cabido et al. 2018). An open shrub layer is formed by Croton lachnostachyus Baill. (Euphorbiaceae) and other species; in the herb layer, the grasses Oplismenus hirtellus (L.) P. Beauv., Setaria oblongata (Griseb.) Parodi and Gouinia latifolia (Griseb.) Vasey var. latifolia (Poaceae) occur with the highest frequency (Cabido et al. 2018). Mean annual precipitation between October and May is 750 mm (Luti et al. 1979; Moglia and Giménez 1998).
Plant species
We selected four plant species: one native, one neonative (Richardson 2001; Essl et al. 2019) and two considered IAPS for this region (Giorgis and Tecco 2014; Giorgis et al. 2017; Cabido et al. 2018). All of them are woody species with fleshy fruits primarily dispersed by birds (Ferreras et al. 2008; Ponce et al. 2012; Vergara-Tabares et al. 2016; Díaz Vélez et al. 2017; Chari et al. 2020).
The native species Celtis ehrenbergiana (Klotszch) Liebm. (Cannabaceae), is a thorny tree producing fruits from November to April. This species is usually found in dry forests and scrubs from the southern USA to central Argentina (Berg and Dahlberg 2001; Cabido et al. 2018). The shrub species Lantana camara L. (Verbenaceae) has a fruiting phenology between January and May (Díaz Vélez et al. 2017), and is considered a neonative species (sensu Essl et al. 2019) for this region (Chaco Mountain Forest), because it is originally from other phytogeographical regions of South America (Richardson 2001). These two species have evidence that ants have interacted with their fruits (Grilli and Galetto 2009; Osunkoya and Perrett 2011; Palacio et al. 2020). Moreover, there is no record of human introduction of L. camara in the southern region of the continent (central Argentina), so probably it colonized this region due to human-induced changes of the biophysical environment (Essl et al. 2019). However, nowadays this plant species is commonly used as ornamental. In addition, other authors have considered this plant species as a native invader species because it is native from Argentina (northeast) but it became an invader in the Córdoba Province (Giorgis and Tecco 2014; Torres and Galetto 2014).
The two IAPS are Pyracantha angustifolia (Franch.) C.K. Schneid. (Rosaceae) (hereafter IAPS-P) and Ligustrum lucidum W.T. Aiton (Oleaceae) (hereafter IAPS-L). The former is a thorny evergreen shrub native to Asia that produces fruits throughout the whole year, mainly in autumn and winter months (Tecco et al. 2006). Ligustrum lucidum is an evergreen and shade-tolerant Chinese tree. The fruiting period of IAPS-L in low Chaco Mountain Forest ranges from May to the end of November (Ferreras et al. 2008). Both IAPS have been introduced as ornamental and are widely used in landscaping and gardening (Novara 1993; Giorgis and Tecco 2014). In the Chaco Mountain Forest of Córdoba, IAPSL was introduced at the beginning of the twentieth century, whereas the first occurrence of IAPS-P has been recorded in 1984 (Delucchi 1991; Hoyos et al. 2010; Giorgis et al. 2011). Afterwards, they spread throughout the region (Giorgis et al. 2017; Cabido et al. 2018). Ligustrum lucidum has evidence that ants have interacted with their fruits (Ferreras et al. 2008). Furthermore, while most native species disperse in the warm and rainy season, mostly from November to April, both IAPS disperse their fruits also in the cold and dry season (Gurvich et al. 2005; Tecco et al. 2013).
Fruit harvest and measurements
To perform the experiments described in the following sections, we collected mature fruits from at least five different plant individuals on the previous day of the beginning of the experiments. In this way, we created fruit pools from each species that were maintained in the fridge between 3 and 5 °C prior to the experiments. We selected 100 diaspores from the fruit pool of each plant species to determine fresh diaspore mass using a precision balance (0.0001 g) in the laboratory. It has been previously documented that fruit variables (i.e., fruit size and fruit mass) are highly correlated (Flörchinger et al. 2010). Additionally, for all species we counted the number of seeds per fruit from 100 fruits.
Caging experiment
In order to disentangle the contribution of ants to the secondary dispersal process of the selected plant species, we designed a paired experiment: some fruits were exposed to all potential dispersers and others were excluded from vertebrates. Vertebrates were excluded from diaspores using wire cages (15 × 10 × 10 cm; 1.5 cm mesh) that impeded vertebrates to make contact with fruits but allowed ants to freely go in and out. Each experimental pair consisted of a set of 10 intact fruits on white filter paper placed directly on the ground inside a wire cage (experimental treatment: vertebrates excluded), and an exposed set of diaspores placed outside the cage, 10 m away (open control: ants and vertebrates allowed) (Figs. 1a; 2). Previous studies showed that the filter paper apparently has neither effect on ant behavior nor on other potential seed dispersers (e.g., Byre and Levey 1993; Pizo and Oliveira 2000; Passos and Oliveira 2002). One transect of each kind was placed in each selected site with five paired stations (i.e., experimental treatment and open control) for each plant species. Fruits were set out at 09:00 a.m. and after 24 h, we recorded the number of fruits removed in each experimental category. We settled the experiment during a 24 h period, based on evidence from previous studies using a similar approach (e.g., Passos and Oliveira 2002; Christianini et al. 2007; Christianini and Oliveira 2010; Camargo et al. 2016). A given fruit was considered removed if it was not found within 30 cm of its original location. Before establishing the transects, all fruits from any of the selected plant species were removed from the area. Also, it is important to mention that during the experiment, apart from ants no invertebrate species were seen at caged fruits, nor rodents at exposed ones.

Conceptual diagram showing the two experiments performed: a caging experiment showing a paired station with fruits on white filter paper, inside wire cages (left) and exposed (right) and b fruit manipulation experiment showing a paired station with fruits on white filter paper, intact (left) and manipulated (right) simulating previous manipulation by avian frugivores. Each transect contained five paired stations (separated by 10 m) for each plant species; it is important to consider that the experiments were carried out with ten fruits per species, only five are shown schematically in the figure

Intact fruits of the native species Celtis ehrenbergiana on white filter paper, a exposed and b inside wire cages
Fruit manipulation experiment
To register the ant species assemblage interacting with diaspores, as well as the kind of interaction established with the four plant species selected, in each site we placed two transects with five diaspore depots in each one (depots were 10 m from one another). In one transect, on each diaspore depot we placed 10 intact fruits on white filter paper. In the other transect, in each diaspore depot we placed 10 fruits that were manipulated with forceps to simulate pecking by bird's beak (i.e., imitating fallen fruits beneath trees after manipulation by birds) (Fig. 1b). Transects were also separated at least by 10 m. Diaspores were set at 08:00 a.m. until 20:00 p.m. on sunny days and checked at 15 min intervals over 1 h, allowing us to record ant activity during all day. We registered the species and number of ants at each diaspore depot and their behavior towards the diaspores of selected plant species in three different categories, based and adapted from previous studies: (1) Inspect/collect liquids (IC-L): this include ants that contact the fruits with their mandibles, antennae or legs but do not remove them, and ants that collect liquid from the fleshy diaspore (mainly fruits with the pulp exposed, simulating previous handling by frugivores); (2) Consume/remove fruit pulp (CR-FP): this include ants that consume fruit pulp at the diaspore depot, and ants that remove small pieces of fruit pulp; and (3) Remove diaspores (RD): ants that grab the fruits and carry them away from the diaspore depots (Pizo and Oliveira 1998; Passos Oliveira 2002; 2004; Aranda-Rickert and Fracchia 2011; Pirk and Lopez de Casenave 2017) (Fig. 3). These three types of behaviours towards the diaspores were treated as ant-diaspore interactions in the present study. When we could not identify the ant species in the field, we collected and preserved them in 96% ethanol for further identification. Code numbers assigned to morphospecies apply only to this study. A full collection of voucher specimens was deposited at Instituto Multidisciplinario de Biología Vegetal (UNC-CONICET).

Types of behaviors performed by ants towards the diaspores: a ants performing diaspore inspection or liquid collection on the diaspore depot (i.e. Gnamptogenys striatula on Celtis eherenbergiana), b ants consuming and removing fruit pulp (i.e. Acromyrmex crassispinus on Pyracantha angustifolia), and c ants removing diaspores (i.e., Acromyrmex crassispinus removing a fruit of Lantana camara)
Data analyses
We used the data from the caging experiment to analyze if ants remove fruits from each of the selected plant species. For this analysis, we performed Generalized Mixed Effect Models (GLMM) using ‘MASS’ package (Ripley et al. 2013). We used ‘glmmPQL’ function with quassibinomial distribution as the data were overdispersed. The model was constructed with Site as random effect, and Treatment (open control and vertebrates excluded) and Species origin (native, neonative, IAPS-L and IAPS-P) as explanatory variables. The ant assemblage was visualized through a Venn Diagramm using ‘venn.diagram’ function from ‘VennDiagram’ package (Chen and Boutros 2011). The dataset from the fruit manipulation experiment was used to test the effects of Fruit condition (intact vs. manipulated) and Species origin as well as Fruit condition (intact vs. manipulated) and Type of interaction on the response variable: Number of ants visiting diaspore depots. In both cases, we performed a GLMM using ‘MASS’ package with Poisson distribution. The model was constructed with Site as random effect, and Fruit condition, Type of interaction and Species origin as explanatory variables.
To evaluate if diaspore mass and seed number differed among species, we performed a GLM using the ‘glm’ function with a Gaussian and Poisson distribution, respectively; and then compared with the null model using the ‘anova’ function. For all models performed, we tested for differences between factor levels using ‘lsmeans’ function from ‘emmeans’ package (Lenth et al. 2018). All the analyses and graphics were performed using R version 2020 (R Core Team 2020); except the first diagram performed entirely in Inkscape (2020).
Results
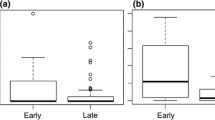
The caging experiments revealed that the quantity of diaspores removed within 24 h in the enclosure and the control is similar for all plant species (χ2 = 0.91; Df = 1; P = 0.33; Table 1; Fig. 4), suggesting that ants are the main dispersers of the diaspores on the Chaco Mountain Forest ground. However, the analysis also showed meaningful differences in the removal rate among the plant species (χ2 = 12.22; Df = 3; P = 0.006; Fig. 4). Diaspores of the neonative species had higher removal rates in both type of conditions (exposed and wired cages) compared with the native species (Estimate = 1.43; Std error = 0.55; t ratio = 2.63; P = 0.04) and IAPS-P (Estimate = 1.72; Std error = 0.55; t ratio = 3.12; P = 0.01).

Number of fruits removed per plant species (IAPS-L, Ligustrum lucidum; IAPS-P, Pyracantha angustifolia; Native, Celtis ehrenbergiana and Neonative, Lantana camara) considering fruits exposed (black) and inside wire cages (gray) in three sites of Chaco Mountain Forest (Córdoba, central Argentina). Black continuous lines indicate individual values, if there are two identical values, the line is drawn twice as long; the dashed line represents the mean; different letters indicate differences between plant species
From the fruit manipulation experiment, we observed that the ant species assemblage interacting with diaspores was composed of 12 native species belonging to four subfamilies: Formicinae, Myrmicinae, Ectatomminae and Dolichoderinae (Table 2). We found a greater number of ant species interacting with the native species Celtis ehrenbergiana (10 species) compared with the neonative and both IAPS: Lantana camara (6), Ligustrum lucidum (4) and Pyracantha angustifolia (4), respectively (Fig. 5a). Moreover, the native plant species presented the higher number of exclusive ant species interacting with their diaspores (3) (Fig. 5a), while two ant species were observed interacting exclusively with IAPS-P and IAPS-L (Fig. 5a; Table 2). All ant species inspected the fruits or collected liquid from them (Table 2). Seven species consumed fruit pulp on the diaspore depots and/or removed it; including Gnamptogenys striatula (mainly an arthropod predator; Latkke 1990). However, only two ant species removed diaspores, Acromyrmex crassispinus and Pheidole cordiceps (Table 2). While the former removed diaspores of all plant species, P. cordiceps only removed diaspores of Lantana camara (we followed one individual of each species; > 15 m and up to 5 m, respectively).

a Venn diagram showing the ant species assemblage and b barplot of the number of ants in relation to the type of interaction (i.e., consume/remove fruit pulp (CR-FP), inspect/collect liquids (IC-L) and remove diaspores (RD)) and the fruit condition: manipulated (black) and intact (gray) of native (Celtis ehrenbergiana), neonative (Lantana camara), IAPS-L (Ligustrum lucidum) and IAPS-P (Pyracantha angustifolia) in three sites of Chaco Mountain Forest in Córdoba, central Argentina; different letters indicate significant differences
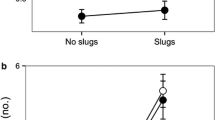
The model performed between fruit condition (i.e., intact or manipulated) and species origin from the fruit manipulation experiment did not show a significant interaction term (χ2 = 5.79; Df = 3; P = 0.12). The number of ants visiting diaspore depots was different among plant species with different origins (χ2 = 31.89; Df = 3; P ≤ 0.0001). IAPS-L had significantly less ants visiting their fruits compared with the other three plant species selected, whereas no significant differences was found among the latter. Moreover, we detected meaningful differences concerning fruit condition (χ2 = 18.3; Df = 1; P ≤ 0.0001), with a higher number of ants visiting diaspore depots with manipulated fruits (3.06 ± 7.45; mean ± SD) compared to diaspore depots with intact fruits (1.53 ± 4.77).
We did not detect an interaction between fruit condition and type of interaction (χ2 = 2.1; Df = 2; P = 0.38). But we detect differences between type of interactions (χ2 = 13.1; Df = 2; P = 0.001); inspect/collect liquid was different from consume/remove fruit pulp (Estimate = -0.93; Std error = 0.27; t value = -3.41; P = 0.002; Fig. 5b). The other type of interactions did not differ between them. Moreover, we ran an analysis to evaluate variation in the type of treatment by ants to each plant species considered (Supplementary Information Appendix 1). For the native plant species, we did not detect differences in the number of ants performing the different type of interactions, but the IC-L seems the most frequent interaction. In the neonative plant species, RD was the interaction most frequently observed, and it differed significantly from CR-FP and IC-L. For IAPS-L and IAPS-P, CR-FP was the most frequent ant behavior toward fruits (Supplementary Information Appendix 1).
Diaspore mass differed significantly among selected plant species (χ2 = 1027.1; Df = 3; P ≤ 0.0001). The neonative plant species presented the lowest diaspore mass compared with the other species (Fig. 6). The native plant species and IAPS-P presented the highest mass but did not differ from one another (Fig. 6). The native plant species presented a higher variability in diaspore mass data compared with the other species. Overall, selected plant species differed significantly in the number of seeds per fruit (χ2 = 415.62; Df = 3; P ≤ 0.0001), but only IAPS-P was meaningful different from the other plant species. The number of seeds per fruit (mean ± SD) was 4.97 ± 0.48 for IAPS-P, followed by 1.38 ± 0.49 for neonative, 1.25 ± 0.17 for IAPS-L, and 1 ± 0 for native species.

Beanplot (the dashed line represents the mean) showing diaspore mass of 100 fruits of the Native (Celtis ehrenbergiana), IAPS-P (Pyracantha angustifolia), IAPS-L (Ligustrum lucidum) and Neonative (Lantana camara) plants selected in this study. Black continuous lines indicate individual values, if there are two identical values, the line is drawn twice as long. Different letters indicate significant differences between diaspore mass
Discussion
Our results highlight the role of ants in the secondary dispersal processes of native, neonative and IAPS with fleshy fruits in seasonally dry forests of central Argentina, as it has been observed in other regions of the world (Rico-Gray and Oliveira 2007). Additionally, our results reveal that ants could also be important in primary dispersal processes (i.e., removing fruits without necessarily being previously manipulated by a primary disperser). Twelve native ant species interacted with the fleshy fruits; the native plant species presented a higher number of ant species interacting with their fruits, followed by the neonative species and the two IAPS. Acromyrmex crassispinus and Pheidole cordyceps were the only two species removing diaspores. Furthermore, the fleshy fruits of selected plant species differed in their mass and in the number of seeds per fruit, two important traits that could influence the likelihood of being removed by the ants. The plant species with fruits with a lesser mass (i.e., neonative) had higher rates of removal. These results highlight the importance that ants and diaspore traits have in these diffuse mutualisms as well as in the invasive processes of plants in subtropical ecosystems.
Although in central Argentina we lack large ponerines that are well-known for their role as secondary dispersers in the Brazilian Atlantic rainforest and Cerrado savanna (e.g., Odontomachus, Pachycondyla and Dinoponera; Pizo and Oliveira 1998; 2000; Passos and Oliveira 2002; 2004; Christianini et al. 2012; Magalhães et al. 2018), our results indicate that ants in Argentine seasonally dry forests can remove diaspores from different plant species. The results obtained in the cage experiments (i.e., similar removal rates in caged and exposed fruits) indicate that ants are the main diaspore dispersers of the selected plant species on the ground of Chaco Mountain Forest. This result is consistent with previous studies in Neotropical forests and savannas, where ants have been reported as the main secondary dispersers of fleshy diaspores primarily dispersed by vertebrates (Oliveira et al. 2017, and included references). Indeed, our removal rates in cage vs. exposed treatments are similar to those found for primarily bird-dispersed plants in Cerrado savanna such as Ocotea pulchella (Nees & Mart.) Mez and Erythroxylum pelleterianum A. St.-Hil. (Christianini et al. 2007). Furthermore, a previous study performed with Erythroxylum ambiguum Peyr. in Brazil, showed that ants and birds had the same quantitative importance for seed dispersal effectiveness, highlighting the importance of diplochory for several non-myrmechocorus species (Camargo et al. 2016).
In accordance with other studies, we registered a higher number of interactions and visitation by ants to plant species whose fruits were treated with the forceps simulating a previous bird manipulation. Bieber et al. (2013) found a higher frequency of ant visits when fruits were mandibulated or embedded in bird faces, and Guerra et al. (2018) showed that partially defleshed fruits were more removed than intact fruits. Mandibulated fruits are more accessible to ants because previous handling by frugivores increases exposure of the fleshy pulp (Bieber et al. 2013). In our study, the native plant species had a higher number of native ants interacting with its diaspores, suggesting that the ant species are more acquainted with these fruits compared to other diaspores that are relatively new in the ecosystems when considering a temporal scale (i.e., neonative and both IAPS; Thompson 1999). Some species interacted on the diaspore depot with the fruits, only inspecting them or performing liquid collection (e.g., Brachymyrmex sp., Crematogaster sp., Linephitema gallardoi, Wasmannia auropunctata, Camponotus rufipes). All these are small ants, except for C. rufipes, that has been reported to feed mainly on liquid rewards (Ronque et al. 2018). Other ant species consumed and removed fruit pulp, although the seeds remained on the diaspore depot. This kind of behavior could not only reduce the fungal attack but also facilitate seed germination, as previously reported for other plant species (Oliveira et al. 1995; Leal and Oliveira 1998). Gnamptogenys striatula was one of the ant species observed performing this task, and in Brazil this species was seen removing fruits of the shrub Erythroxylum ambiguum to their nests (Camargo et al. 2016). Only two ant species dispersed diaspores in our study: Acromyrmex crassispinus and Pheidole cordiceps. The small P. cordiceps only removed diaspores of the neonative species (i.e., with less diaspore mass). Indeed, P. cordiceps recruits nestmates to carry fruits as a group, as observed in other ant species (i.e., Pheidole bergi Mayr, 1887 in Monte Desert; Pirk et al. 2009), this constitutes a flexible mechanism for simulating a larger-bodied forager (Traniello 1989) that could broaden the range of food size that ants can carry. On the other hand, Acromyrmex crassispinus removed diaspores of all the plant species selected for this study (see also Calheiros et al. 2019, on A. subterraneus (Forel 1893) in Cerrado savanna). We also saw some seedlings of the selected plant species (as well as other species with fleshy fruits) growing in refuse dumps of A. crassispinus. Similar effects on plant recruitment by Atta and Acromyrmex were previously documented by Farji-Brener and Silva (1996) and Farji-Brener et al. (2017) in southern Argentina, and Dalling and Wirth (1998) in Panamanian rainforest. In grasslands of Argentina, Amodeo et al. (2017) found a higher germination rate by invasive alien Prunus mahaleb L. after their seeds had their pulp removed by Acromyrmex lundii (Guérin-Méneville 1838) and were deposited near the nest. This behavior was also observed in A. crassispinus in the present study, indicating that further investigation is needed with germination tests after ant manipulation of the seeds (see Leal and Oliveira 1998; Christianini and Oliveira 2009).
From the point of view of the invasive process, our results confirm the relevance of native ant species as dispersal agents of the IAPS selected in the present study. As previously mentioned, former studies revealed that these plant species are dispersed by birds (i.e., Pyracantha angustifolia, Ligustrum lucidum, see Ferreras et al. 2008; Vergara-Tabares et al. 2016). Although dispersal mutualisms are important to ensure the establishment of some IAPS, diaspore dispersal is most relevant during the spread phase (Theodarides and Dukes 2007; Traveset and Richardson 2014). The removal of fallen fleshy diaspores by ants can benefit plants in different ways. By removing the diaspores to their nests, ants could provide favorable microsites for seed germination and seedling establishment (Passos and Oliveira 2002; 2004; Christianini et al. 2007; Christianini and Oliveira 2009). If the diaspores do not reach the nest, the seeds can get lost on the way and be dropped in favorable microsites for seedling recruitment (Horvitz 1981; Vander Wall and Longland 2005). Additionally, ants could diminish seed predation and seedling competition by moving seeds away from parent trees (Pizo 1997; Pizo and Oliveira 1998; Vander Wall and Longland 2005). Furthermore, the ant-induced movements can rearrange the seed rain generated primarily by vertebrate dispersers (Robert and Heithaus 1986; Horvitz and Le Corff 1993; Pizo and Oliveira 1999; Pizo et al. 2005). Just as fruit removal has several positive effects on plants, it can also benefit ant colonies. Indeed, there is evidence that consumption of lipid-rich aril improves larval development in predominantly carnivorous ponerines such as Odontomachus chelifer (Latreille, 1802) (Bottcher and Oliveira 2014). Given that some of the IAPS in our study have a wider fruiting period than native species (Tecco et al. 2006), increased availability of fallen fleshy diaspores in periods of food scarcity in this subtropical forest may enhance dispersal by ants. Furthermore, we observed differences in the number of ants visiting the diaspore depots, being IAPS-L the plant species with less visits; this could be related to differences in the chemical attributes of the fruits, that would be interesting to study. For example, previous evidence has shown that ants prefer lipid rich diaspores (Pizo and Oliveira 2001).
Finally, it is well known that the dispersal process depends on fruit size relative to the body size of the frugivorous animal (Jordano 2000). Ants are not the exception, and it has been shown that small-sized diaspores are removed in greater numbers and to increased distances than medium and large-sized diaspores (Pizo and Oliveira 2001; Anjos et al. 2020). In this way, the differences in diaspore mass between species that we observed could be related to the higher removal rate that we found in the neonative plant species compared with the other plant species (in both conditions: treatment and control). The neonative plant species presented the lowest diaspore mass. Furthermore, the high variability in diaspore mass observed in native compared to non-native species, could allow a broader range of dispersal agents from different sizes (Jordano 2000). Despite the lower removal rate of IAPS-P, the diaspores of this plant species have more seeds than the other selected species, likely conferring an advantage to this invasive alien since seed number can be relevant for seed dispersal effectiveness (Schupp et al. 2010). These results highlight the importance of considering diaspore traits when plant–seed dispersal interactions are assessed.
Our results suggest that ants are relevant removal agents of native and invasive alien plant species in seasonally subtropical forests of the southernmost part of South America. This study is the first to describe the ant species assemblage attending diaspores, and to report secondary dispersal by ants in central Argentina. Our field account includes native plant species and plant species of different origins, highlighting the relevance of these ant-plant mutualisms for the invasive process (Richardson et al. 2000a, b). The Chaco Mountain Forest has a rich community of fleshy-fruited plants (i.e., native, neonative, exotic and invasive species) that could benefit from the litter-dwelling ant assemblage. Further investigation should focus on post dispersal processes resulting from ant-diaspore interactions, including experiments to find out if manipulation by ants enhances seed germination, as well as if ant-induced seed displacement improves seedling establishment in suitable sites (Horvitz 1981; Passos and Oliveira 2002). In this way, it would be possible to determine if ants contribute to the establishment of new seedlings in native communities and acquire a whole picture of the role of ants in the invasive process. Our study shows that ants can be considered relevant agents increasing the invasion success of alien fleshy-fruited plants in central Argentina. The relative contribution of ants versus birds in the spread stage of these plants remains to be assessed.
Data availability
Not applicable.
Code availability
Not applicable.
References
Amodeo MR, Vázquez MB, Zalba SM (2017) Generalist dispersers promote germination of an alien fleshy-fruited tree invading natural grasslands. PLoS ONE 12:e0172423
Anjos DV, Leal LC, Jordano P, Del-Claro K (2020) Ants as diaspore removers of non-myrmecochorous plants: a meta-analysis. Oikos. https://doi.org/10.1111/oik.06940
Aranda-Rickert A, Fracchia S (2011) Pogonomyrmex cunicularius as the keystone disperser of elaiosome-bearing Jatropha excisa seeds in semi-arid Argentina. Entomol Exp Appl 139:91–102
Berg CC, Dahlberg SV (2001) A revision of Celtis subg. Mertensia (Ulmaceae). Brittonia 53:66–81
Bieber AGD, Silva PS, Oliveira PS (2013) Attractiveness of fallen fleshy fruits to ants depends on previous handling by frugivores. Ecoscience 20:85–89
Blackburn TM, Pyšek P, Bacher S et al (2011) A proposed unified framework for biological invasions. Trends Ecol Evol 26:333–339
Bottcher C, Oliveira PS (2014) Consumption of lipid-rich seed arils improves larval development in a Neotropical primarily carnivorous ant, Odontomachus chelifer (Ponerinae). J Trop Ecol 30:621–624. https://doi.org/10.1017/S0266467414000479
Byrne MM, Levey DJ (1993) Removal of seeds from frugivore defecations by ants in a Costa Rican rain forest. Vegetatio 107:363–374
Cabido M, Zeballos SR, Zak M, Carranza ML, Giorgis MA, Cantero JJ, Acosta AT (2018) Native woody vegetation in central Argentina: classification of Chaco and Espinal forests. Appl Veg Sci 21:298–311
Cabrera AL (1976) Regiones fitogeográficas argentinas. In: Kugler WF (ed) Enciclopedia argentina de agricultura y jardineira, Tomo 2, vol 2. Acme, Buenos Aires, pp 1–8
Calheiros AC, Ronque MUV, JrH S, Oliveira PS (2019) Field biology and foraging ecology of the leaf-cutter ant Acromyrmex subterraneus (Hymenoptera: Formicidae) in Neotropical Cerrado savanna. Environ Entomol 48:1434–1441
Camargo PH, Martins MM, Feitosa RM, Christianini AV (2016) Bird and ant synergy increases the seed dispersal effectiveness of an ornithochoric shrub. Oecologia 181:507–518
Chari LD, Martin GD, Steenhuisen SL, Adams LD, Clark VR (2020) Biology of invasive plants 1. Pyracantha angustifolia (Franch.) CK Schneid. Invasive Plant Sci Manag 13:120–142
Chen H, Boutros PC (2011) VennDiagram: a package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform 12:1–7
Christianini AV, Oliveira PS (2009) The relevance of ants as seed rescuers of a primarily bird-dispersed tree in the Neotropical cerrado savanna. Oecologia 160:735–774
Christianini AV, Oliveira PS (2010) Birds and ants provide complementary seed dispersal in a neotropical savanna. J Ecol 98:573–582
Christianini AV, Mayhé-Nunes AJ, Oliveira PS (2007) The role of ants in the removal of non-myrmecochorous diaspores and seed germination in a neotropical savanna. J Trop Ecol 23:343–351. https://doi.org/10.1017/S0266467407004087
Christianini AV, Mayhé-Nunes AJ, Oliveira PS (2012) Exploitation of fallen diaspores by ants: are there ant–plant partner choices? Biotropica 44:360–367
Corlett RT (2015) The anthropocene concept in ecology and conservation. Trends Ecol Evol 30:36–41
Cronk QCB, Fuller JL (1995) Plant invaders. The threat to natural ecosystems. Chapman & Hall, London
Dalling JW, Wirth R (1998) Dispersal of Miconia argentea seeds by the leaf-cutting ant Atta colombica. J Trop Ecol 14:705–771
Delucchi G (1991) Especies adventicias y naturalizadas nuevas o críticas para Argentina. B Soc Argent Bot 27:179–181
Diagne D, Leroy B, Vaissière AC et al (2021) High and rising economic costs of biological invasions worldwide. Nature 592:571–576. https://doi.org/10.1038/s41586-021-03405-6
Diamond JM, Ordunio D (1999) Guns, germs, and steel. Books on Tape, New York
Díaz Vélez MC, Ferreras AE, Silva WR, Galetto L (2017) Does avian gut passage favour seed germination of woody species of the Chaco Serrano Woodland in Argentina? Botany 95:493–501
Dietz H, Edwards PJ (2006) Recognition that causal processes change during plant invasion helps explain conflicts in evidence. Ecology 87:1359–1367
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, London
Essl F, DullingerbS GP et al (2019) A conceptual framework for range-expanding species that track human-induced environmental change. Bioscience 69:908–919
Farji-Brener AG, Silva JF (1996) Leaf cutter ants (Atta laevigata) aid to establishment success of Tapirira velutinifolia (Anacardiaceae) seedlings in a parkland savanna. J Trop Ecol 12:163–168
Farji-Brener AG, Tadey M, Lescano MN (2017) Leaf-cutting ants in Patagonia: how human disturbances affect their role as ecosystem engineers on soil fertility, plant fitness and trophic cascades. In: Oliveira PS, Koptur S (eds) Ant-plant interactions: impacts of humans on terrestrial ecosystems. Cambridge University Press, Cambridge, pp 377–390
Ferreras AE, Torres C, Galetto L (2008) Fruit removal of an invasive exotic species (Ligustrum lucidum) in a fragmented landscape. J Arid Environ 72:1573–1580
Ferreras AE, Giorgis MA, Tecco PA, Cabido MR, Funes G (2015) Impact of Ligustrum lucidum on the soil seed bank in invaded subtropical seasonally dry woodlands (Córdoba, Argentina). Biol Invasions 17:3547–3561
Ferreras AE, Whitworth-Hulse JI, Tecco PA, Marcora PI, Funes G (2019) Environmental constraints to native woody species recruitment in invaded mountain woodlands of central Argentina. For Ecol Manag 440:189–201
Flörchinger M, Braun J, Böhning-Gaese K, Schaefer HM (2010) Fruit size, crop mass, and plant height explain differential fruit choice of primates and birds. Oecologia 164:51–161
Giorgis M, Tecco PA (2014) Arboles y arbustos invasores de la provincia de Córdoba (Argentina): una contribución a la sistematización de bases de datos globales. B Soc Argent Bot 49:681–603
Giorgis MA, Cingolani AM, Chiarini F et al (2011) Composición florística del Bosque Chaqueño Serrano de la provincia de Córdoba, Argentina. Kurtziana 36:9–43
Giorgis MA, Cingolani AM, Gurvich DE, Tecco PA, Chiapella J, Chiarini F, Cabido M (2017) Changes in floristic composition and physiognomy are decoupled along elevation gradients in central Argentina. Appl Veg Sci 20:558–571
Gorb E, Gorb S (2003) Seed dispersal by ants in a deciduous forest ecosystem: mechanisms, strategies, adaptations. Springer, New York
Grilli G, Galetto L (2009) Remoción de frutos de una especie invasora (Lantana camara L.) en el Bosque Chaqueño de Córdoba (Argentina). Ecol Austral 19:2
Guerra TJ, Messeder JV, Arruda AJ, Fuzessy LF, Dayrell RL, Neves FS, Silveira FA (2018) Handling by avian frugivores affects diaspore secondary removal. PLoS ONE 13:e0202435. https://doi.org/10.1371/journal.pone.0202435
Gurvich DE, Tecco PA, Díaz S (2005) Plant invasions in undisturbed ecosystems: the triggering attribute approach. J Veg Sci 6:723–728
Holmes PM (1990a) Dispersal and predation in alien Acacia. Oecologia 83:288–290
Holmes PM (1990b) Dispersal and predation of alien Acacia seeds: effects of season and invading stand density. S Afr J Bot 56:428–434
Horvitz CC (1981) Analysis of how ant behaviours affect germination in a tropical myrmecochore Calathea microcephala (P. & E.) Koernicke (Marantaceae): microsite selection and aril removal by neotropical ants, Odontomachus, Pachycondyla, and Solenopsis (Formicidae). Oecologia 51:47–52
Horvitz CC, Le Corff J (1993) Spatial scale and dispersion pattern of ant- and bird-dispersed herbs in two tropical lowland rain forests. Vegetatio 107:351–362
Hoyos LE, Gavier-Pizarro GI, Kuemmerle T, Bucher EH, Radeloff VC, Tecco PA (2010) Invasion of glossy privet (Ligustrum lucidum) and native forest loss in the Sierras Chicas of Córdoba, Argentina. Biol Invasions 12:3261–3275
Inkscape Project (2020) Inkscape. Retrieved from https://inkscape.org
Janzen DH (1985) The natural history of mutualisms. In: Janzen DH (ed) The biology of mutualisms. Oxford University Press, Oxford, pp 40–99
Jordano P (2000) Fruits and frugivory. In: Fenner M (ed) Seeds: the ecology of regeneration in plant communities, 2nd edn. CABI Publishing, Wallingford, pp 125–166
Leal IR, Oliveira PS (1998) Interactions between fungus-growing ants (Attini), fruits and seeds in cerrado vegetation in southeast Brazil. Biotropica 30:170–178
Lengyel S, Gove AD, Latimer AM, Majer JD, Dunn RR (2010) Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey. Perspect Plant Ecol Evol Syst 12:43–55
Lenth R, Singmann H, Love J, Buerkner P, Herve M (2018) Emmeans: estimated malrginal means, aka least-squares means. R Package Version 1:3
Luti, R, Bertrán de Solís MA, Galera MF, Muller de Ferreira N, Berzal M, Nores M, Herrera MA, Barrera JC (1979) Vegetación. In: Vázquez, Miatello y Roque (eds) geografía física de la provincia de Córdoba. Boldt, Córdoba, pp 1–464
Magalhaes VB, Espírito Santo NB, Salles LFP, JrH S, Oliveira PS (2018) Secondary seed dispersal by ants in Neotropical cerrado savanna: Species-specific effects on seeds and seedlings of Siparuna guianensis (Siparunaceae). Ecol Entomol 43:665–674
Moglia G, Gimenez AM (1998) Rasgos Anatómicos característicos del hidrosistema de las principales especies arbóreas de la región Chaqueña Argentina. For Syst 7:53–71
Moyano J, Rodriguez-Cabal MA, Nuñez MA (2020) Highly invasive tree species are more dependent on mutualisms. Ecology 101:e02997
Novara L (1993) Rosaceae. Aportes botánicos de salta-serie. Flora 2:1–53
Ohkawara K, Akino T (2005) Seed cleaning behavior by tropical ants and its anti-fungal effect. J Ethol 23:93–98
Oliveira PS, Galetti M, Pedroni F, Morellato LPC (1995) Seed cleaning by Mycocepurus goeldii ants (Attini) facilitates germination in Hymenaea courbaril (Caesalpiniaceae). Biotropica 27:518–522
Oliveira PS, Christianini AV, Bieber AGD, Pizo MA (2017) Anthropogenic disturbances affect the interactions between ants and fleshy fruits in two Neotropical biodiversity hotspots. In: Oliveira PS, Koptur S (eds) Ant-plant interactions: impacts of humans on terrestrial ecosystems. Cambridge University Press, Cambridge, pp 133–156
Ortiz DP, Elizalde L, Pirk GI (2021) Role of ants as dispersers of native and exotic seeds in an understudied dryland. Ecol Entomol. https://doi.org/10.1111/een.13010
Osunkoya OO, Perrett C (2011) Lantana camara L. (Verbenaceae) invasion effects on soil physicochemical properties. Biol Fert Soils 47:349–355
Oyarzabal M, Clavijo J, Oakley L et al (2018) Vegetation units of argentina. Ecol Austral 28:040–063
Palacio FX, Siepielski AM, Lacoretz MV, Ordano M (2020) Selection on fruit traits is mediated by the interplay between frugivorous birds, fruit flies, parasitoid wasps and seed-dispersing ants. J Evol Biol 33:874–886
Passos L, Oliveira PS (2002) Ants affect the distribution and performance of seedlings of Clusia criuva, a primarily bird-dispersed rain forest tree. J Ecol 90:517–528
Passos L, Oliveira PS (2004) Interaction between ants and fruits of Guapira opposita (Nyctaginaceae) in a Brazilian sandy plain rainforest: ant effects on seeds and seedlings. Oecologia 139:376–382
Pikart TG, Souza GK, Serrao JE, Zanuncio JC (2011) Leafcutter ants: a small dispersal agent of the invasive plant Murraya paniculata. Weed Res 51:548–551
Pirk GI, De Casenave JL (2017) Ant interactions with native and exotic seeds in the Patagonian steppe: influence of seed traits, disturbance levels and ant assemblage. Plant Ecol 218:1255–1268
Pirk GI, Di Pasquo F, de Casenave JL (2009) Diet of two sympatric Pheidole spp. ants in the central Monte desert: implications for seed–granivore interactions. Insect Soc 56:277–283
Pizo MA (1997) Seed dispersal and predation in two populations of Cabralea canjerana (Meliaceae) in the Atlantic Forest of southeastern Brazil. J Trop Ecol 13:559–577
Pizo MA, Oliveira PS (1998) Interaction between ants and seeds of a nonmyrmecochorous neotropical tree, Cabralea canjerana (Meliaceae), in the Atlantic forest of Southeast Brazil. Am J Bot 85:669–674
Pizo MA, Oliveira PS (1999) Removal of seeds from vertebrate faeces by ants: effects of seed species and deposition site. Can J Zool 77:1595–1602
Pizo MA, Oliveira PS (2000) The Use of Fruits and seeds by ants in the Atlantic forest of southeast Brazil. Biotropica 32:851–861
Pizo MA, Oliveira PS (2001) Size and lipid content of nonmyrmecochorous diaspores: effects on the interaction with litter-foraging ants in the Atlantic rain forest of Brazil. Plant Ecol 157:37–52
Pizo MA, Guimarães Jr PR, Oliveira PS (2005) Seed removal by ants from faeces produced by different vertebrate species. Ecoscience 12:136–140
Ponce AM, Grilli G, Galetto L (2012) Frugivoría y remoción de frutos ornitócoros en fragmentos del bosque chaqueño de Córdoba (Argentina). Bosque 33:33–41
Prior KM, Saxena K, Frederickson ME (2014) Seed handling behaviours of native and invasive seed-dispersing ants differentially influence seedling emergence in an introduced plant. Ecol Entomol 39:66–74
R Core Team (2020) R: a language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. URL https://www.R-project.org/
Rejmánek M, Richardson DM (2013) Trees and shrubs as invasive alien species–2013 update of the global database. Divers Distrib 19:1093–1094
Richardson DM, Pyšek P (2001) Plant invasions. Encycl Biodivers 4:677–688
Richardson DM, Allsopp N, D’Antonio CM, Milton SJ, Rejmánek M (2000a) Plant invasions–the role of mutualisms. Biol Rev 75:65–93
Richardson DM, Pyšek P, Rejmánek M, Barbour MG, Panetta FD, West CJ (2000b) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107
Rico-Gray V, Oliveira PS, Oliveira PS (2007) The ecology and evolution of ant-plant interactions. University of Chicago Press, Chicago
Ripley B, Venables B, Bates DM, Hornik K, Gebhardt A, Firth D, Ripley MB (2013) Package ‘mass.’ Cran r 538:113–120
Roberts JT, Heithaus ER (1986) Ants rearrange the vertebrate-generated seed shadow of a neotropical fig tree. Ecology 67:1046–1051
Ronque MU, Fourcassié V, Oliveira PS (2018) Ecology and field biology of two dominant Camponotus ants (Hymenoptera: Formicidae) in the Brazilian savannah. J Nat His 52:237–252
Sala OE, Chapin FS, Armesto JJ et al (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Sanhueza C, Arcila A, Rodríguez D, Zalba S (2007) El papel de las hormigas en la dispersión de dos leguminosas exóticas invasoras. https://doi.org/10.13140/RG.2.2.26578.81603
Schupp EW (1995) Seed-seedling conflicts, habitat choice, and patterns of plant recruitment. Am J Bot 82:399–409
Schupp EW, Jordano P, Gómez JM (2010) Seed dispersal effectiveness revisited: a conceptual review. New Phytol 188:333–353
Seebens H, Blackburn TM, Dyer EE et al (2017) No saturation in the accumulation of alien species worldwide. Nat Commun 8:1–9
Tecco PA, Gurvich DE, Díaz S, Pérez-Harguindeguy N, Cabido M (2006) Positive interaction between invasive plants: the influence of Pyracantha angustifolia on the recruitment of native and exotic woody species. Austral Ecol 31:293–300
Tecco PA, Urcelay C, Díaz S, Cabido M, Pérez-Harguindeguy N (2013) Contrasting functional trait syndromes underlay woody alien success in the same ecosystem. Austral Ecol 38:443–451
Theoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phytol 176:256–273
Thompson JN (1999) The evolution of species interactions. Science 284:2116–2118
Torres CC, Galetto L (2014) Nectar sugar composition and flower visitors for the naturalized exotic Lantana camara (Verbenaceae) at Central Argentina. Int J Plant Reprod Biol 6:174–180
Traniello JF (1989) Foraging strategies of ants. Annu Rev Entomol 34:191–210
Traveset A, Richardson DM (2014) Mutualistic interactions and biological invasions. Annu Rev Ecol Evol S 45:89–113
Turbelin AJ, Malamud BD, Francis RA (2017) Mapping the global state of invasive alien species: patterns of invasion and policy responses. Global Ecol Biogeogr 26:78–92
Van der Pijl L (1982) Principles of dispersal in higher plants. Springer, Berlin
Van Kleunen M, Dawson W, Schlaepfer D, Jeschke JM, Fischer M (2010) Are invaders different? A conceptual framework of comparative approaches for assessing determinants of invasiveness. Ecol Lett 13:947–958
Vander Wall SB, Longland WS (2004) Diplochory: Are two seed dispersers better than one? Trends Ecol Evol 19:155–161
Vander Wall SB, Longland WS (2005) Diplochory and the evolution of seed dispersal. In: Forget PM, Lambert JE, Hulme PE, Vander Wall SB (eds) Seed fate: predation, dispersal and seedling establishment. CABI publishing, Wallingford, pp 297–314
Vergara-Tabares DL, Badini J, Peluc SI (2016) Fruiting phenology as a “triggering attribute” of invasion process: Do invasive species take advantage of seed dispersal service provided by native birds? Biol Invasions 18:677–687
Vilà M, Espinar JL, Hejda M et al (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14:702–708
Whitworth-Hulse JI, Magliano PN, Zeballos SR, Gurvich DE, Spalazzi F, Kowaljow E (2020) Advantages of rainfall partitioning by the global invader Ligustrum lucidum over the dominant native Lithraea molleoides in a dry forest. Agr for Meteorol. https://doi.org/10.1016/j.agrformet.2020.108013
Acknowledgements
The authors would like to acknowledge the financial support and assistance of the following institutions: the National Scientific & Technological Research Council (CONICET), the National University of Córdoba (SECyT – UNC) and FONCyT (PICT 2015-0538) from the Ministry of Science and Technology (MINCyT). This study was performed when MP was a post-doctoral fellow of CONICET. We would like to thank the owners of all the field sites selected for the possibility of performing the study in their properties and also Juan Ignacio Whitworth-Hulse for showing us the field sites. Furthermore, the authors would like to thank Gabriela Pirk and two anonymous reviewers for reading and performing very helpful comments on a previous version of the manuscript. PSO was supported by CAPES (Finance Code 001), and by research grants from the CNPq (302219/2017-0, 303730/2021-8) and FAPESP (BIOTA Program, 2014/23141-1, 2017/16645-1).
Funding
This research was funded by the National Scientific & Technological Research Council (CONICET), the National University of Córdoba (SECyT – UNC) and FONCyT (PICT 2015-0538) from the Ministry of Science and Technology (MINCyT).
Author information
Authors and Affiliations
Contributions
Conceptualization, M.P., S.R.Z., L.G. and P.S.O.; methodology, M.P. and S.R.Z.; formal analysis, M.P. and S.R.Z.; investigation, M.P. and S.R.Z.; data curation, M.P.; writing—original draft preparation, M.P.; writing—review and editing, M.P., S.R.Z., L.G. and P.S.O., funding acquisition, L.G. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Ethical approval
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Pereyra, M., Zeballos, S.R., Galetto, L. et al. Influence of secondary dispersal by ants on invasive processes of exotic species with fleshy fruits. Biol Invasions 24, 3275–3289 (2022). https://doi.org/10.1007/s10530-022-02845-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-022-02845-z