
Abstract
The use of halotolerant beneficial plant-growth-promoting (PGP) bacteria is considered as a promising eco-friendly approach to improve the salt tolerance of cash crops. One strategy to enhance the possibility of obtaining stress-alleviating bacteria is to screen salt impacted soils. In this study, amongst the 40 endophytic bacteria isolated from the roots of Sahara-inhabiting halophytes Atriplex halimus L. and Lygeum spartum L., 8 showed interesting NaCl tolerance in vitro. Their evaluation, through different tomato plant trials, permitted the isolate IS26 to be distinguished as the most effective seed inoculum for both plant growth promotion and mitigation of salt stress. On the basis of 16S rRNA gene sequence, the isolate was closely related to Stenotrophomonas rhizophila. It was then screened in vitro for multiple PGP traits and the strain-complete genome was sequenced and analysed to further decipher the genomic basis of the putative mechanisms underlying its osmoprotective and plant growth abilities. A remarkable number of genes putatively involved in mechanisms responsible for rhizosphere colonization, plant association, strong competition for nutrients, and the production of important plant growth regulator compounds, such as AIA and spermidine, were highlighted, as were substances protecting against stress, including different osmolytes like trehalose, glucosylglycerol, proline, and glycine betaine. By having genes related to complementary mechanisms of osmosensing, osmoregulation and osmoprotection, the strain confirmed its great capacity to adapt to highly saline environments. Moreover, the presence of various genes potentially related to multiple enzymatic antioxidant processes, able to reduce salt-induced overproduction of ROS, was also detected.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Salinity is a major abiotic stress affecting cultivable land worldwide, especially in arid and semi-arid regions. It results in billion-dollar losses in crop production annually (Cuin and Shabala 2007; UNU-INWEH Annual Report 2014). One of the regions severely affected, in connection with intense desertification, is situated on the southern shore of the Mediterranean Sea (Marquina 2012). Impacted by global warming, this region is also faced with regular rainfall deficiency, so profitable cultivation requires the use of irrigation systems and intensive fertilization, which make the soil particularly prone to salinization, especially when salty waters and dripping systems are used (UNU-INWEH Annual Report 2014).
Most crops are glycophytes and thus unable to grow where salt concentrations in the soil are high (Paul and Lade 2014). In such plants, salt stress causes an initial water deficit (osmotic effect), accumulation of specific ion toxicity and nutrient imbalance, which inhibit plant growth by disrupting physiological processes (Munns and Tester 2008; Shu et al. 2010; Gerszberg and Hnatuszko-Konka 2017). Tomato (Solanum lycopersicum L.) is one such plant with economic importance that is seriously affected by salinity, which limits yield and fruit quality (Cuartero and Fernández-Muñoz 1998). Increasing NaCl concentration in the nutrient solution has been shown to adversely affect tomato shoots and roots, plant height, K+ concentration, and K+/Na+ ratio (Al-Karaki 2000).
Given the above facts, limiting salt accumulation in arable lands by approaches such as leaching, planting halophytes, and using organic matter conditioner has become one of the key aspects of agriculture development, together with improving crop salt tolerance (UNU-INWEH 2014, Paul and Lade 2014). However, all attempts so far have had limited success or do not seem to be sustainable approaches, considering their high cost and/or difficulty of application (Cuin and Shabala 2007; Fan et al. 2016).
One of the promising eco-friendly alternative approaches consists of using beneficial bacteria associated with cultivated plants that have already shown plant growth enhancement and salt stress mitigation properties (Mayak et al. 2004; Paul and Lade 2014; Zhumakayev et al. 2021; Dif et al. 2021). These plant-growth-promoting bacteria (PGPB) have been identified from various genera, of which Pseudomonas and Bacillus are the most extensively studied (Islam et al. 2014; Kumari et al. 2015). In fact, associated bacteria can directly influence plant growth by producing indole acetic acid (IAA), solubilizing phosphorus, and secreting siderophores (Bhattacharyya and Jha 2012; Jaiswal et al. 2019).
Salt-tolerant bacteria refer to microorganisms able to grow in environments with a wide range of salinities, from 1 to 33% NaCl, and also in the absence of NaCl (Khan et al. 2016).
Interestingly, by exhibiting multiple stress-related traits and mechanisms of adaptation that enable them to live in highly saline environments, halotolerant bacteria may contribute to plant-protection capabilities under cultivation-inhibiting levels of salt (Etesami and Beattie 2018).
An important mechanism is related to the ability to accumulate compatible osmolytes, such as trehalose, proline, and glycine betaine, to maintain intracellular osmotic balance (Slama et al. 2015). Another well documented mechanism of stress alleviation is related to plant metabolic modulation of ethylene signalling and cellular oxidative stress (Etesami and Beattie 2018; Sharma et al. 2016). In fact, plants growing under a wide variety of environmental stresses, including pathogen infections and soil salinity, significantly increase their endogenous ethylene production and initiate programmed cell death through the generation of reactive oxygen species (ROS) (Glick et al. 2007; Trobacher 2009; Baxter et al. 2014). Interestingly, several PGPB strains have been shown to inhibit salt-induced ethylene production via their capacity for enzymatic hydrolysis of the ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC) (Zahir et al. 2009; Siddikee et al. 2011; Shin et al. 2016; Silambarasan et al. 2020).
In addition, beneficial bacteria have been shown to help maintain ROS-homeostasis in plants by stimulating both enzymatic and non-enzymatic antioxidant systems, which prevent plant cells from deleterious effects of stress-induced oxidative bursts (Hamilton et al. 2012; Islam et al. 2015; Alexander et al. 2020). The enzymatic antioxidants involved mainly include superoxide dismutase, catalase and peroxidase, while non-enzymatic antioxidants include ascorbate, glutathione, carotenoids and phenolic compounds (Gill and Tuteja 2010b; Hasanuzzaman et al. 2018).
In response to various environmental stresses, proline synthesis, like ethylene synthesis, is often induced in plants to mediate free radical scavenging and subcellular structure stabilization (Verbruggen and Hermans 2008), and plant inoculation with proline-producing bacteria has already proved to enhance proline content, helping the plant to raise its water status even under stressed conditions (Vardharajula et al. 2011; Metoui Ben Mahmoud et al. 2020).
In the aim of discovering new potential PGPB, the search for isolates living in ecological niches, especially in harsh environmental conditions, has increased considerably (Ramakrishna et al. 2019).
In this context, the present investigation focuses on the study of plants that have successfully adapted to the extremely arid and hot climate conditions of some regions of the Algerian Sahara desert, especially those characterized by poor and salty soils. The success of the natural establishment of Atriplex halimus L. (Chenopodiaceae) and Lygeum spartum L. (Poaceae) in such harsh environments suggests that associated bacteria may help to promote their growth and counteract salt stress effects; thus contributing to the salt-tolerance characteristics of the host plants (Rodríguez-Llorente et al. 2019).
The objective of the research presented here was to explore both the presence and potentialities, in terms of osmoprotection and plant growth enhancement, of bacteria that colonize Sahara-living A. halimus L. and L. spartum L. The work included screening plant-associated bacteria with halotolerant abilities and evaluating their stress-alleviating capacities in the context of establishing tomato plant culture. The plant-growth-promoting traits potentially involved were investigated concurrently. The genome of the most interesting strain was sequenced and analysed for taxonomic characterization and putative identification of mechanisms conferring environmental plasticity and increased metabolic potential.
Materials and methods
Sampling sites and collection procedure
Root samples of the native halophytes Atriplex halimus L. and Lygeum spartum L. were taken from two locations in the arid regions of Djelfa, Algeria, noted S1 (34°56′08.0"N 3°03′19.8"E) and S2 (34°50′03.0"N, 3°05′58.56"E). These sites are characterized by hypersaline rocky soils (EC > 8.3 mS/cm; pH > 7.5) corresponding to "soil SET#7" in the FAO World Reference Base (IUSS Working Group WRB 2015).
Intact roots of the vigorous halophytes were sampled at depths of 10 to 20 cm and transferred to sterile plastic bags, then stored at 4 °C until processing. A total of three root samples per plant and per site were collected for the isolation of endophytic bacteria.
Isolation of endophytic bacteria
The isolation was performed according to the procedure described by Bashan et al. (1993). Briefly, collected root samples were first washed in running tap water and divided into pieces 4 to 5 cm in length. They were subjected to surface sterilization by stepwise washing in 70% ethanol for 5 min and sodium hypochlorite solution (3% available Cl−) for 10 min, followed by several rinses in sterile distilled water.
To release the endogenous microflora, surface sterilized root samples were weighed (10 g) and aseptically ground in a mortar and pestle with 60 ml sterile water. In order to increase the initial biomass yield, aliquots of 100 µl of the resulting turbid suspensions were inoculated into tubes containing 10 ml of nutrient broth and incubated for 24–36 h on a rotary shaker (200 rpm) at 30 °C.
Each resulting sample was plate-spread (100 µl/plate) on Tryptic Soy Agar medium (TSA) containing 1.3% NaCl, then incubated at 30 °C for 24 h. Isolated colonies were purified by repeated culturing, in the same conditions, then stored at − 20 °C in 20% glycerol-saline TSA (1.5% NaCl) broth until use.
To prove that epiphytic microorganisms could not grow after the surface-sterilization of the roots and that all the isolated bacteria were endophytic, a 0.3 ml aliquot of the washing water of the surface-sterilized roots was inoculated on TSA agar at 30 °C for 3 days (Schulz et al. 1993). In addition, plugs (6 mm in diameter) from bacterial cultures were subjected to the same surface-sterilization treatments. Plates were then incubated for 14 days at 30 °C and recorded for microbial growth (Zamoum et al. 2015).
Screening of salt-tolerant isolates
The tolerance of isolated strains to NaCl was evaluated on TSA agar medium supplemented with increasing NaCl concentrations (0, 1, 2, 3, 5 and 7%). The medium was poured into Petri plates and bacterial strains were streaked on each plate, in three replicates, then incubated at 30 °C for 24 h. The influence of NaCl concentrations on bacterial growth was recorded.
In planta evaluation of growth promotion and salt stress mitigation abilities
Plant material
Seeds of tomato (Solanum lycopersicum L.) variety Aicha (fixed characteristics and determined growth), were used in this study since this variety is extensively grown by farmers despite its known high sensitivity to salt.
Preparation of microbial inoculants
The selected bacterial strains were cultivated at 30 °C and 120 rpm in 150 ml of Tryptic Soy Broth (TSB) for 48 h. Cell pellets were then collected by centrifugation at 6000 g for 15 min and re-suspended in physiological saline solution (0.9%). The concentrations of the bacterial suspensions were adjusted to approximately 1–2 × 108 CFU/ml (Landa et al. 2001) using a DO standard curve, after which the bacterial suspensions were distributed at 18 ml per tube and used for seed bacterization.
Petri-dish assay
The selected salt-tolerant bacteria were evaluated for their ability to promote tomato seedling growth and establishment under stress-free and acute salt stress conditions. Prior to use, tomato seeds were surface-sterilized by sequential washing in 70% ethanol for 1 min and 3% NaClO for 10 min, followed by five rinses in sterile distilled water and aseptic drying (Landa et al. 2001); then treated by soaking in each endophytic strain suspension (108 CFU/ml; 120 rpm; 2 h in darkness) and air-drying under laminar flow (Lee et al. 2010). Seeds were then placed in 9 cm Petri dishes (20 seeds per dish) containing a filter paper watered with 5 ml of sterile NaCl solution at 7 dS/m (equivalent to 0.5% NaCl) or non-saline distilled water. The plates were placed in a phytotronic growth chamber (14 h photoperiod, 20–25 °C night–day, 70% relative humidity) for 10 days. At the end of this period, emergence (germination %), fresh weight, shoot length, and root length, were evaluated. In order to limit measuring errors, shoot and root lengths were estimated by digital image analysis using ImageJ software (https://imagej.nih.gov/ij/; v.1.52a; Rueden et al. 2017). Images were obtained by placing seedlings on a Hewlett-Packard Scanjet 5300C flatbed scanner with a resolution of 400 dpi. A vigour index (VI) was also calculated as defined by Abdul Baki and Anderson (1973) as follows: VI = (RL + SL) × GP, where RL is main root length (mm), SL is shoot length (mm) and GP is germination percentage.
Non-bacterized seeds subjected to the same saline and non-saline growth conditions acted as controls. The experiment was conducted in a randomized design and repeated three times with at least 20 replicates per treatment.
Greenhouse assay
Plant growth-promoting and saline-stress-alleviating abilities of selected isolates were evaluated by trials in controlled saline soil conditions.
A pot experiment was carried out under plastic film in a greenhouse (natural illumination with a 14 h photoperiod, 25 °C and 60% relative humidity). Plastic pots (8 cm in diameter × 12 cm high) were filled with arable soil (loamy sand texture with slightly basic pH (7.6), non-salt (EC = 0.2 mS/cm) and containing a low amount of organic carbon (less than 0.4%)), sterilized (autoclaved three times for 60 min at 120 °C at 24 h intervals) and sown (2 cm deep) with surface-bacterized tomato seeds (five seeds per pot) prepared as described earlier (Landa et al. 2001). The plants were supplied daily with 10 ml of sterile NaCl solutions variably adjusted in such a manner as to maintain a salt concentration in the soil corresponding approximately to 0, 5 and 7 dS/m (equivalent to 0, 3.25 and 5% NaCl, respectively) and representing non-saline, moderate and acute saline conditions, respectively.
Pots were inspected daily and the evaluation was performed 21 days post-sowing by determining the seedlings’ emergence, fresh weight, shoot length, root length, and vigour index.
Non-bacterized seeds subjected to the same saline and non-saline growth conditions acted as positive and negative controls, respectively. The experiment was conducted in a randomized design and repeated three times with at least 15 replicates per treatment.
16S rRNA gene-based identification of the selected isolate
The bacterial isolate that exhibited the best results in planta was subjected to molecular characterization via the sequencing of its 16S rRNA gene as previously described (Springer et al. 1995). Briefly, after rapid DNA extraction (Liu et al. 2000), the 16S rRNA gene was amplified by PCR (using the forward 10-30F, 5′-GAGTTTGATCCTGGCTCA-3′, and reverse 1500R, 5'-AGAAAGGTGGAGATCCAGCC-3' primers) as described by Rainey et al.
(1996), then purified and sequenced by the Macrogen company (Amsterdam, Netherlands). The gene sequences obtained were compared for similarity with type strains in the EzTaxon-e database (http://eztaxon-e.ezbiocloud.net/; Kim et al. 2012). The neighbour-joining method with 1000 bootstrap replications was used to infer the phylogenetic tree in the MEGA X package (Kumar et al. 2018).
Whole genome sequencing of selected strain, assembly and annotation
For genomic analysis of the selected strain, total genomic DNA obtained from the bacterial culture was extracted by the NucleoSpin® Tissue kit using the support protocol targeting the bacterial DNA according to the manufacturer´s instructions (Macherey–Nagel, Germany). DNA concentration of the sample was estimated with a Modulus™ Single Tube Multimode Reader fluorometer (Turner BioSystems, CA, USA). We used 1 ng of DNA as input for the library construction with the Nextera XT DNA library prep kit (Illumina, CA, USA). The final genomic library was measured with a Bioanalyzer 2100 (Agilent Technologies, CA, USA) using the Bioanalyzer High Sensitivity DNA Kit, Modulus Single Tube Multimode Reader (Turner Biosystems, CA, USA), the Quant-iT™ dsDNA Assay Kit (Thermo Fisher Scientific, PE, USA) and, finally, the MCNext™ SYBR® Fast qPCR Library Quantification Kit (MCLAB, CA, USA) used with the Rotor-Gene 3000 (Corbett Research, Australia). The pooled library of samples was constructed in the laboratory of Mendeleum (Mendel University in Brno, Czech Republic). Sequencing was carried out with the MiniSeq Mid Output Kit (Illumina, CA, USA) on the MiniSeq instrument (Illumina). The quality of raw reads was examined by FastQC 0.52 (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/).
De novo assembly of the raw reads (cleaned using Trimmomatic 0.36.5) (Bolger et al. 2014) was achieved using Unicycler 0.4.7.0 (Wick et al. 2017). The assembly quality was assessed by Quast 5.0.2 (Gurevich et al. 2013). Initial raw reads were mapped against the assembled genome using Bowtie2 2.3.4.3 (Langmead and Salzberg 2012). The mapping quality was evaluated using Qualimap 2.2.1 (García-Alcalde et al. 2012) to estimate mean coverage.
In order to detect genes involved in salt stress tolerance and PGPB traits, the draft genome was annotated using Prokka 1.13 (http://www.ncbi.nlm.nih.gov/genome/annotation_prok; Seemann 2014).
Clusters of genes coding for secondary metabolites were analysed using AntiSMASH 4.1 (Weber et al. 2015) (detection strictness: relaxed, with all extra features on) with a focus on clusters involved in salt stress tolerance.
Comparative genomics of S. rhizophila IS26 and its close-related strains
Core and accessory genes in S. rhizophila IS26 and its closely related strains were identified using Roary ver. 3.11.2 (Page et al. 2015) with default settings (minimum percentage identity for blastp: 95%, percentage of isolates a gene must be in to be core: 99%), based on the GFF3 files of all the selected genomes, generated by the annotation (Seemann 2014).The indicative neighbour joining phylogenetic tree generated by ROARY was visualized using the Interactive Tree of Life (iToL) (Letunic and Bork 2019). Phandango (Hadfield et al. 2018) was used to visualize the matrix showing the presence or absence of each core-gene in each strain. The “summary_statistics” file generated by Roary was then used to visualize the proportions of the pan-genome.
In vitro detection of plant-growth-promoting and salt-stress-mitigation traits
The bacterial isolate that exhibited the best results in planta was explored in connection with its stress-mitigation and plant growth-promoting abilities.
ACC production
Production of 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase was evaluated using the nitrogen-free Dworkin–Foster’s salt minimal agar medium (Dworkin and Foster 1958). The medium was supplemented with ACC (3 mM) as a sole nitrogen source. The strain was streak-inoculated on the medium and incubated at 30 °C for 7 days. Bacterial growth indicated a positive result for ACC-deaminase production (Tamreihao et al. 2016).
IAA production
One millilitre aliquots of the bacterial suspensions (≈106 CFU/ml) were inoculated into 250 ml Erlenmeyer flasks containing 50 ml of yeast extract-tryptone broth (Khamna et al. 2010) amended with L-tryptophan (5 mg/ml). Flasks were cultured for 5 days on a rotary shaker (200 rpm, 30 °C). The liquid cultures were then centrifuged at 4000 g for 30 min and the production of IAA was revealed by mixing 2 ml of the supernatant with 4 ml of Salkowski reagent (1 ml of 0.5 M FeCl3 solution in 49 ml of 35% w/v HClO4 solution). IAA production was highlighted by the appearance of a pink colour after 30 min in a dark room. Optical density was measured by spectrophotometry at 530 nm and the quantity of IAA was calculated using a pure IAA standard curve (Passari et al. 2015).
Catalase production
A small amount of bacterial colony was transferred to the surface of a clean, dry, glass slide using a loop, then mixed with a drop of 3% H2O2. A positive result corresponded to a rapid evolution of oxygen (within 5–10 s.) shown by the formation of bubbles; while the absence of bubble formation indicated a negative result (Reiner 2010).
Siderophore production
The production of siderophore was evaluated on chrome azurol S agar plates (Hu and Xu 2011). Six-millimetre plugs from bacterial cultures were inoculated on the medium and incubated for 1 week at 30 °C. The production of siderophores was revealed by the presence of orange halos surrounding the bacterial colonies.
Phosphate solubilization
The experiments were conducted in 500 ml Erlenmeyer flasks containing 100 ml of Pikovskaya broth (Pikovskaya 1948) amended with an insoluble phosphate source: Ca3(PO4)2, AlPO4 or FePO4, at a concentration of 5 g/l. Flasks were inoculated by 1 ml aliquot of the bacterial suspensions (≈106 CFU/ml) and cultured for 1 week on a rotary shaker (200 rpm, 30 °C). The supernatant cultures were harvested by centrifugation (10 000 g, 10 min) then used to evaluate the soluble phosphate. The rate of dissolved phosphate was determined by the spectrophotometric molybdenum-blue method (Liu et al. 2014). A non-inoculated flask acted as control.
Hydrogen cyanide production
HCN production was highlighted by growing bacteria in Bennett medium supplemented with glycine (4.4 g/l). A Whatman filter paper was flooded with 0.5% picric acid in 2% sodium carbonate for 1 min and stuck underneath the Petri dish lids. The plates were sealed with parafilm and incubated for 1 week at 30 °C. The presence of orange to red colour on the Whatman paper margins indicated a positive result for HCN production (Passari et al. 2015).
All the in vitro assays were performed in triplicate.
Statistical analysis
All the quantitative data generated in this study were subjected to an analysis of variance (ANOVA) and principal component analysis (PCA) using Minitab 17 software. Mean separation was accomplished by a Tukey's HSD post-hoc test, considering significance at P ≤ 0.05.
Results
Isolation and screening of endophytic salt-tolerant bacteria
As shown in Table 1, a total of 40 strains were isolated from the roots of A. halimus (13 isolates) and L. spartum L. (27 isolates).
The endophytic nature of the isolated bacteria was confirmed by (1) representative cultures that were subjected to the surface sterilization protocol exhibited no growth on TSA medium and (2), the wash water samples from all surface-sterilized roots showed no microbial growth in the same medium. This indicates that the epiphytic bacteria were effectively eliminated with the procedure used and confirms that the bacterial strains obtained were indeed endophytic. As shown in Table 1, all the isolates grew well at 0% NaCl, indicating that salt was not required for bacterial development. From the total number of isolates, eight (noted IS5, IS8, IS10, IS12, IS14, IS26, IS34 and IS38) were able to grow in 3% to 7% NaCl and were thus selected for evaluation of their salt stress mitigation and plant growth potentials.
Plant-growth-promotion and salt-stress-mitigation assays
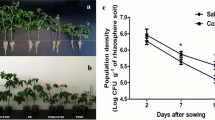
The Petri-dish experiment indicated that the eight selected strains varied notably in their efficiency for enhancing the germination rate, fresh weight, shoot length, root length, and vigour of tomato seedlings. As shown in Fig. 1a, strains IS26 and IS38 exhibited the most significant vigour index, with increases of about 1/2 to 4 times compared to non-inoculated controls under salt-stress and salt-stress-free conditions, respectively.

Evaluation of salt-tolerant bacterial strain inocula on tomato seedlings. a Assessment of plant vigour index in non-saline (0 S/m) and acute saline (7 dS/m) conditions. Seeds were inoculated with one of the following screened salt-tolerant bacteria: IS5, IS8, IS10, IS12, IS14, IS26, IS34 or IS38 in a Petri-dish trial. Vigour indexes [germination (%) × plant length (mm)] were evaluated 10 days after seed planting (variety Aicha). Bars indicate standard error of the mean; columns with the same letters are not significantly different at P = 0.05. b Principal component (PC) analysis plot (PC1 vs. PC2) obtained from data on the Petri-dish experiment of screened salt-tolerant bacterial strains. Numbers in square brackets indicate the saline exposure (dS/m). Controls correspond to non-bacterized seeds. Groups are indicated by grey background
To obtain a general view of the plant growth promotion and salt stress mitigation efficiency of the tested strains, a multivariate principal component (PC) analysis was conducted on all data recorded in the Petri-dish experiment (Fig. 1b). Together, the first axis (PC1) and the second axis (PC2) explained 94% of the differences between treatments (83% for PC1 and 11% for PC2). PC analysis clearly split the treatments into three groups (I, II and III), within each of which the plant growth capacity was equivalent. Group I, comprising treatments IS5, IS8, IS10, IS12, IS14 and IS34 under a salinity of 7 dS/m, showed low plant growth potential similar to that of the saline control. The treatments that clustered in group II, with strains exposed to non-saline conditions, exhibited plant growth performance equivalent to that of the non-saline control. Group III, composed of strains IS26 and IS38 under a salinity of 7 dS/m, exhibited PC values intermediate between groups I and II. These results highlighted the potential of isolates IS26 and IS38 for salt stress mitigation and enhancement of plant growth.
Based on these findings, the isolates IS26 and IS38 were selected for subsequent greenhouse experiments. As shown in Fig. 2a, the vigour index was significantly influenced by salt concentrations and by the bacterial treatment. As expected, increasing salt concentration caused significant, progressive reductions in the vigour index.

Effects of selected bacterial treatments on tomato plant development. a Assessment of plant vigour index in non-saline (0 dS/m) and saline (5 and 7 dS/m) conditions. Seeds (variety Aicha) were inoculated with one of the selected salt-stress alleviating bacteria, IS26 or IS38, in a pot trial and vigour indexes [germination (%) × plant length (mm)] were evaluated 21 days after planting in greenhouse conditions. Bars indicate standard error of the mean; columns with the same letters are not significantly different at P = 0.05. b Principal component (PC) analysis plot (PC1 vs PC2) obtained from data of pot experiment using selected salt-tolerant bacterial strains. Numbers in square brackets indicate the salinity to which the plants were exposed (dS/m). Controls correspond to non-bacterized seeds. Groups are indicated by grey background
Under non-saline conditions, strains IS26 and IS38 showed highly significant improvement of plant vigour compared to the non-inoculated control, indicating their effective ability to favour plant growth. This ability was notably maintained under moderate and acute salt concentrations (5 and 7 dS/m, respectively), especially with IS26 treatment.
The PC analysis (Fig. 2b) conducted with data recorded in the greenhouse experiment (germination rate, fresh weight, and shoot and root length) demonstrated global plant growth promotion and salt stress mitigation efficiency for strains IS26 and IS38 in both saline (5 and 7 dS/m) and non-saline conditions. Together, the first axis (PC1) and the second axis (PC2) explained 95.83% % of the variation between treatments (80.33% for PC1 and 15.49% for PC2).
PC analysis grouped saline-exposed-IS38 treatments and the corresponding saline (5 and 7 dS/m) controls, which exhibited the lowest plant growth, in the same cluster (I). However, IS26 treatment under both saline and non-saline conditions (cluster II), expressed more interesting PC values than the non-saline uninoculated control, thus confirming the potential expressed by this strain for both PGP and salt stress alleviation. Accordingly, isolate IS26 was subjected to further characterization.
Molecular identification and phylogenetic analysis of the selected isolate IS26
To identify the strain IS26 at the molecular level, a 1.462 kb amplicon of 16S rDNA gene was obtained by PCR. The sequence of the resulting amplicon was submitted to the NCBI Genbank database under the accession number MT755732. Based on the BLAST results, the strain was related to the Stenotrophomonas genus. The phylogenic relationship between strain IS26 and the other Stenotrophomonas type species can be seen in the neighbour-joining dendrogram (Fig. 3). The similarity level was 100% with Stenotrophomonas rhizophila DSM 14,405 (Wolf et al. 2002).

Neighbour-joining tree based on 16S rDNA sequences showing the relations between strains IS26 and type-species of the Stenotrophomonas genus. The numbers at the nodes indicate the levels of bootstrap support based on neighbour-joining analyses of 1000 resampled data sets. Bar, 0.050 nt substitution per nt position. Xanthomonas cassavae CFBP 4642 T was used as outgroup
General genome features of the selected strain IS26
The draft genome assembly genome of strain IS26 consists of a single circular chromosome of 4,485,597 bp and an average G + C content of 66.2% (Fig. 4a). No plasmid was found in the genome. The genome contained 4532 predicted coding sequences (CDSs). A total of three rRNAs, comprising a single 5S rRNA, a single 16S rRNA and a single 23S rRNA, together with a single tmRNA and 54 tRNA genes representing 24 amino acids were identified in the IS26 genome. Among CDSs, 2934 genes were associated with Clusters of Orthologous Groups of proteins (COGs) composed of 25 functional categories (Fig. 4b).

Strain IS26 genome overview. a Circular genome map of strain chromosome. From the outer circle to the inner circle: scale marks, CDS on the forward (brown) and reverse (green) strand, rRNA (crimson), tRNA (blue), GC content (black/pink) and GC skew (red/purple). Figure generated by DNA Plotter. b Genome COGs functional categories and associated frequencies
This whole-genome shotgun project has been deposited in DDBJ/ENA/GenBank under accession no. WELC00000000.1. The version described in this paper is version WELC00000000.1.
Biosynthetic gene clusters and secondary metabolites
With a total of 30 gene-clusters predicted by antiSMASH (Fig. 5a), the strain IS26 genome harbours genes for many potential secondary metabolites. Interestingly, the analysis highlights a single cluster encoding for the potential antioxidant bacterial pigment “aryl polyene” (Fig. 5b, c), that could be involved in the reduction of oxidative salt stress-induced damage to plant cells (Schöner et al. 2016). Furthermore, three clusters encoding non-ribosomal peptides (NRPS), three clusters of polysaccharide coding genes, a single cluster of genes encoding a bacteriocin, and three clusters involved in fatty acid biosynthesis were also revealed. The remaining clusters are putative ones. Thus, antiSMASH results exhibited the potential of the strain for the biosynthesis of antimicrobial compounds.

Strain IS26 genome-antiSMASH analysis Overview. a Predicted biosynthetic clusters. b Cluster 10: arylpolyene biosynthetic gene cluster. c Closest biosynthetic gene clusters of arylpolyene from other bacteria
Analysis of beneficial genes and functions putatively related to strain IS26 bioperformance
To further decipher the genomic basis of the putative mechanisms contributing to the osmoprotective and plant growth abilities of S. rhizophila IS26 in a salt-stressed environment, an extended strain-specific genome mining study was conducted in connection with bacterial invasion and plant-association behaviour, ability to compete with other organisms for rhizospheric/endophytic growth, production of plant growth-stimulating compounds, and potential involvement of mechanisms related to salt stress alleviation.
The annotation of the strain IS26 genome showed the presence of several genes/ signalling pathways potentially related to chemotaxis, cell motility, cell attachment, biofilm formation, quorum sensing, production of lytic enzymes and siderophores, and resistances to different group of antibiotics (Table 2). The genome analysis also identified genes/ clusters putatively involved in phosphate solubilization, production of phytohormones, enzymatic mechanisms triggered to reduce reactive oxygen species (ROS), and processes related to salt sensing and regulation (Table 3).
Comparative genomics of S. rhizophila IS26 and its closely related strains
A comparative genomic study through the pan-genome/core genome analysis was performed using the 12 genomes of the closest strains to S. rhizophila IS26 (Table 4), all recovered from the NCBI's Genome resource (https://www.ncbi.nlm.nih.gov/genome/). The neighbour-joining phylogenetic tree based on the core genes showed that the closest strain from a genomic point of view was S. rhizophila USBA GBX 843 (Fig. 6a), as they clustered in the same clade. In each cluster, strains show similar patterns in their gene absence/presence profiles (Fig. 6b).

Pan-genome analysis of S. rhizophila IS26 and its closest strains. a Neighbour joining (NJ) indicative phylogenetic tree generated from the core genome (Roary output). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. b Distribution of genes across the pan-genome of S. rhizophila IS26 and its closest strains (from Roary). Pan-genome content is displayed as presence/absence (blue/white) blocks according to their order of appearance in the genome annotation panel (top ruler). S. rhizophila IS26 lane is dash framed in purple. c Genes distribution graph. The blue line shows the percentage of strains (y axis) carrying a gene at each position (x axis), the purple line shows S. rhizophila IS26 profile for each position in the pan-genome. Under the graph, core genes; accessory genes; shell genes and cloud genes are located in the pan-genome according to graph. d Pan-genome statistics. The pan-genome matrix and graph were visualized using Phandango (https://jameshadfield.github.io/phandango/#/main)
Although the NJ tree remains indicative, core genome phylogenomy could be a good strategy to highlight genomic similarities between closely related strains of the S. rhizophila species.
For the thirteen strains including S. rhizophila IS26, the pan-genome contains 18,773 genes, of which the core genome represents only 2.55% (480), the shell genome represents 47.45% (8909), and the cloud genome represents a staggering 49.98% (9384), constituting the largest part of the gene pool in this group (Fig. 6c, d).
While the core genome constitutes only 2.5% of the total pan-genome of the compared S. rhizophila strains, the cloud genome represented almost 50% of the total genes. Thus, the richness of the cloud genome confirms the diversity and variety of beneficial traits of the strains in this group. Among the 4,532 genes detected through annotation in S. rhizophila IS26 genome, 480 genes are part of the core genome (10.59%), 3790 genes constitute the shell genome (83.62%), and 262 genes (5.78%) are considered as part of its cloud genome. Of these, 165 genes encode non-hypothetical proteins, highlighting its interesting genomic richness for novel traits and new encoded products.
It also appears that all the plant beneficial genes detected in the strain S. rhizophila IS26 are shared with the rest of the compared strains (part of the core genome) (Tables 2 and 3).
Numerous interesting genes were found solely in S. rhizophila IS26 (Supplementary Table 1). Some genes linked to mobile elements and genomic plasticity were found in the cloud part of the S. rhizophila IS26 genome, such as traG (Conjugal transfer protein TraG), soj_1(Chromosome-partitioning ATPase Soj), spo0C (Chromosome-partitioning protein Spo0J), intA (Prophage integrase IntA), helD_1 (DNA helicase IV), intS_2 and intS_2 (Prophage integrase IntS), traI (Multifunctional conjugation protein TraI), rayT_2 (REP-associated tyrosine transposase), and hindIIIM (Modification methylase HindIII).
Other unique genes linked to the metabolism of heavy metals and toxic compounds were detected as a particularity in S. rhizophila IS26 when compared to its closest strains: sugE_1 (Quaternary ammonium compound-resistance protein SugE), ftsH_1 (ATP-dependent zinc metalloprotease FtsH), and czcA_3 (Cobalt-zinc-cadmium resistance protein CzcA).
Moreover, interesting genes related to motility, chemotaxis and biofilm formation were found only in S. rhizophila, such as: swrC_1 (Swarming motility protein SwrC), fliC_1 and fliC_2 (A-type flagellin), bmaC_1 (Adhesin BmaC autotransporter), pilE_2 (Fimbrial protein), ehaG (Autotransporter adhesin EhaG), and cfaE (CFA/I fimbrial subunit E).
In vitro detection of some IS26-related plant-growth-promoting and salt-mitigation traits
In view of the positive PGP and salt-stress-mitigation results exhibited by the strain IS26 in planta and the significant number of potential benefit-related genes highlighted, an additional in vitro assay was undertaken to explore the presence of some putative mechanisms. Production of IAA, hydrogen cyanide and siderophores were evaluated, along with phosphate solubilization, catalase and ACC deaminase activities. The results showed that strain IS26 grew well and dissolved phosphorus in PVK media containing Ca3(PO4)2 as sole phosphorus source. The amount of dissolved phosphorus was 364 ± 11 mg ml−1 from Ca3 (PO4)2. However, the strain IS26 failed to grow and to dissolve phosphorus in the PVK medium containing FePO4 and AlPO4. Data also showed that strain IS26 was positive for IAA production (80.2 ± 5 μg.ml−1) and catalase. Furthermore, it was qualified as able to produce ACC-deaminase and siderophores but not HCN.
Discussion
The inoculation of PGPB in cultivated plants is currently regarded as a promising practice to promote the development of agriculture in adverse conditions, such as in saline soils (Gamalero et al. 2020). Tomato cultivation is one of the farming practices that are drastically limited by saline environments (Magán et al. 2008). Several studies have reported the role of halotolerant bacteria in managing salt stress in cultivated plants, such as mung beans (Vigna radiata L.) (Islam et al. 2015), wheat (Singh and Jha 2017; Afridi et al. 2019) and tomato (Mayak et al., 2004; Fan et al. 2016; Dif et al. 2021).
In the present study, amongst 40 endophytic bacterial isolates from the roots of native halophytes, 8 showed interesting tolerance to high NaCl concentrations. It has previously been reported that bacteria isolated from saline soils are more likely to withstand saline conditions (Damodaran et al. 2013; Piernik et al. 2017).
Since germination and the early stages of plant growth are the most sensitive stages for plant establishment, especially in hostile soil conditions (Abo-Kassem 2007; Bojović et al. 2010), the 8 isolates showing notable salt-tolerance were inoculated into tomato seeds and consecutively plant-evaluated in Petri-dish and greenhouse assays.
Globally, treatments with halotolerant bacteria exhibited notable PGP effects and permitted differently appreciable limitation of the impact of salt stress on the tomato plant, as previously reported (Kim et al. 2014; Kumar et al. 2017; Cordero et al. 2018).
Through the different plant trials undertaken under moderate (5 dS/m) and acute (7 dS/m) salt stress, the halotolerant endophytic strain IS26 exhibited the best performance for both plant growth enhancement and salt stress mitigation. Under such stress conditions, plant maintenance and development levels were closer to those expressed in non-saline conditions.
Based on these findings, the strain IS26 was considered as the most promising bacterium and thus required further characterization. Based on the complete 16S rDNA sequence and phylogenetic analysis, the isolate IS26 was found to be related to Stenotrophomonas rhizophila DSM 14405 T (Wolf et al. 2002). The Stenotrophomonas genus, phylogenetically placed in the γ-subclass of the Proteobacteria (Moore et al. 1997), is considered of biotechnological interest due to its great metabolic versatility and intraspecific heterogeneity (Ryan et al. 2009; Berg and Martinez 2015). Members of this genus are ubiquitously present in the environment, with soil and plants as their main reservoirs (Ryan et al. 2009).
The phylogenetically and ecologically closely related species S. maltophilia and S. rhizophila are documented as initiating endophytic lifestyle with plants, triggering beneficial antagonism towards soil-borne fungal diseases and promoting plant growth in connection with the production of both antifungal compounds and phytohormones (Ryan et al. 2009; Berg et al. 2010; Schmidt et al. 2012).
By using comparative genomics to analyse the difference between S. maltophilia and S. rhizophila, Alavi et al. (2014) noted the absence of several factors in S. rhizophilia that are crucial for human pathogenic traits, such as virulence factors and heat shock proteins, despite the presence of shared factors related to host invasion and antibiotic resistance. Moreover, the inability of S. rhizophila to grow at 37 °C, which is a critical temperature for strains to colonize the human body, was identified as a very simple criterion for differentiating between pathogenic and non-pathogenic Stenotrophomonas isolates (Alavi et al. 2014).
The plant-associated S. rhizophila, which is thus safe to use, was also observed to produce an osmoprotective substance, glucosylglycerol, which confers remarkable resistance to highly saline soils (Egamberdieva et al. 2016; Schmidt et al. 2012; Alavi et al. 2013). Stenotrophomonas rhizophila was found to exert beneficial effects on the growth of cash crops such as cotton, wheat, tomato, sweet pepper and soybean cultivated under salt stress (Schmidt et al. 2012; Egamberdieva et al. 2016; Afridi et al. 2019).
In this context, to further decipher the genomic basis of the putative mechanisms contributing to S. rhizophila IS26 osmoprotective and plant growth abilities in a salt-stressed environment, an extended strain-specific genome mining study was conducted.
A comparative genomic analysis of S. rhizophila IS26 and its 12 closest strains was also performed to highlight the commonalities and particularities of potential genes and functions.
As a whole, the genome annotation of the strain IS26 highlighted a remarkable number of encoded functionalities potentially involved in plant association and persistence, as well as plant growth performance and salt stress mitigation. Moreover, through comparative genomics, it appeared that all the plant-beneficial genes detected in the strain S. rhizophila IS26 are shared with the rest of the strains compared (as a part of the core genome), which leads this group of species to be considered as a valuable reservoir for other plant-beneficial strains yet to be discovered. Interestingly, unique genes linked to the metabolism of heavy metals and toxic compounds were detected in S. rhizophila IS26 genome. This result suggests future investigations in the context of evaluating bioremediation potentialities.
Root colonization is regarded as an essential step in the promotion of plant growth by PGP rhizobacteria (Lugtenberg and Kamilova 2009). Several genes related to root colonization, such as genes involved in chemotaxis, cell motility and biofilm formation, were found in the strain IS26. Chemotaxis, motility and adhesion are determinant characteristics for bacterial plant-association, allowing bacteria to move through the soil matrix, to mediate the adhesion to plant roots through the production of a variety of exopolysaccharides, and to systematically spread within the plant (Hardoim et al. 2015; Römling and Galperin 2015). In fact, genes related to chemotaxis pathways have been shown to regulate bacterial motility in presence of extracellular attractants or repellents (Erbse and Falke 2009). Flagella- and fimbria-driven motility play a key role in biofilm formation and host plant colonization by bacteria (Shen et al. 2017). The presence of these determining characteristics for bacteria-plant association confirms the endophytic lifestyle of the strain (Hardoim et al. 2015; Römling and Galperin 2015; Egamberdieva et al. 2016).
The potential of stain IS26 to move through the soil matrix and inside the plant was exhibited by the presence of a large number of genes involved in the biosynthesis and assembly of flagella as previously documented (Khanra et al. 2016).
Genes related to fimbriae biosynthesis were also detected as a component putatively implicated both in adhesion to root surfaces and in the formation of complex biofilms (de Oliveira-Garcia et al. 2003). Biofilm formation ability is crucial for bacterial survival, adaptation, and dissemination in a versatile environment (Hall-Stoodley et al. 2004). Alginate metabolism genes were detected in IS26 (algA, algL). Alginate is involved in the development and architecture of biofilms that protect bacteria from antibiotics and other harmful environmental factors (Stapper et al. 2004; MacDonald and Berger 2014). The algL gene highlights a potential production of an alginate lyase that is known to be implied in bacterial invasion or microbial biofilm formation often associated with enhanced drug resistance (MacDonald and Berger 2014).
This colonization process seemed potentially related to quorum sensing communication, through a DSF system for quorum sensing regulation (Ryan et al. 2015; Huedo et al. 2018). Quorum sensing genes RpfC and RpfG responsible for DSF perception and signal transduction, respectively (Ryan et al. 2015; Huedo et al. 2018), are likely to be involved in strain IS26 communication with the plant root during the colonization process, since this strain has genes related to the rpf/DSF (regulation of pathogenicity factors/diffusible signaling factor) system for quorum sensing and cell–cell communication. In this system, the synthesis and perception of the DSF molecule, a quorum sensing regulatory compound, is controlled by a pathogenicity factor gene cluster rpf (Ryan et al. 2015; Huedo et al. 2018). Furthermore, genes encoding for adhesin (an outer membrane-located protein that contributes to host cell surface attachment) were also identified (Willis et al. 2013). LPS-related genes detected in IS26, essential components of the cell surface of Gram-negative bacteria, are also involved in attachment to a plant surface and subsequent potential endophytic colonization behaviour (Zhang et al. 2013; Balsanelli et al. 2013; Hicks and Jia 2018).
Colonization of a plant also requires identified mechanisms to gain access to the plant and to develop. These mechanisms mainly consist of the ability to hydrolyse plant cell wall-bound complex polysaccharide and the capacity to metabolize plant-derived complex carbohydrates (Hallmann et al. 1997; Wolf et al. 2002; Andrés-Barrao et al. 2017), such as the cellulases, hemicellulases and xylases encoded by the S. rhizophila IS26 genome. The degradation ability of this significant root constituent suggests the potential of the strain to subsist and develop in a plant-associated environment. The genome analysis also suggested the presence of genes related to the production of other lytic enzymes, including glucanases and chitinases. As a result, through their ability to degrade glucan and chitin, the main constituents of fungal and oomycete cell walls, these enzymes may have plant pathogen protective functions, suggesting putative strain biocontrol attributes (Veliz et al. 2017).
In general, Stenotrophomonas species are known to show multiple antibiotic resistances (Ryan et al. 2009). In the context of both intense microbiological activity and nutrient-limited environments in the soil, multiple antibiotic resistances could help bacteria to compete in the rhizosphere (Hibbing et al. 2010). As documented for other Stenotrophomonas species (Alavi et al. 2014), the strain IS26 genome was shown to contain numerous genes for antibiotic resistance, including several multi-drug efflux systems, while others provided resistance against particular classes of antibiotics, such as penicillin and first-generation cephalosporins.
Concurrently, the genome analysis showed notable potential of strain IS26 to compete for iron in nutrient-limited environments. Bacterioferritin (bfr) and bacterioferritin-associated ferredoxin bfd and bfrD siderophore genes were detected. Iron, the bioavailability of which is often limited, is an essential metal for almost all microbial growth. Some microorganisms have developed specific iron uptake mechanisms to compete for iron. These mechanisms include production of siderophores, involvement of specific uptake transporters, and synthesis of receptors able to scavenge siderophores that are produced by other microorganisms (Andrés-Barrao et al. 2017). Many of these genes have also been found in Stenotrophomonas maltophilia (Alavi et al. 2013, 2014).
As determinant traits for plant development, strain IS26 was shown to possess genes related to direct and indirect mechanisms for PGP and enhanced tolerance to salt stress. Most of these important putative properties were consecutively confirmed by detection in vitro.
The potential direct mechanisms highlighted were related to phosphate solubilization, and synthesis of plant growth regulator substances, as already mentioned for other Stenotrophomonas strains (Wu et al. 2015; Reinhardt et al. 2008).
As phosphate is an essential macronutrient, limitation of its presence in the soil has a major negative effect on plant growth (Eida et al. 2017). The genome annotation of the strain IS26 highlighted different genes related to a phosphate transport system and inorganic phosphorus solubilization (pap, alpl, ppi, ygiF, cheZ, gdh, gltA, and lldD).
IAA is an important auxin phytohormone related to physiological growth processes such as cell division and tissue differentiation (Duca et al. 2014). The strain IS26 was shown to produce appreciable amounts of IAA phytohormone in vitro, as previously documented for several S. rhizophila strains (Kumar et al. 2019; Silambarasan et al. 2020). Different genes coding for enzymes involved in the IAA biosynthesis pathways from tryptophan were detected in S. rhizophila IS26. In fact, two potential biosynthesis pathways, namely indole-3-acetonitrile (IAN) and indole-3-pyruvate (IPyA), were identified. In the IAN pathway, IAN is first converted to indole-3-acetamide (IAM) by nitrile hydratase, then to IAA by amidase. In the IPyA pathway, IPyA is converted to indole-3-acetaldehyde (IAAld) by indole-pyruvate decarboxylase and then to IAA by aldehyde dehydrogenase (Liu et al. 2014).
Furthermore, another phytohormone gene was detected: the fpp gene, which is involved in the production of abscisic acid (ABA) (Finkelstein 2013). Interestingly, accumulation of ABA is known to inhibit the production of excess of ethylene in plant during exposure to stress such as a salty environment (Yao et al. 2010).
Key genes involved in the biosynthesis of the polyamines spermine and spermidine, such as speE and sped (Alavi et al. 2013), have been detected. These polyamines act as plant growth regulators and have also been found to protect against stress by reducing ethylene level in the plant (Gill and Tuteja 2010a; Alavi et al. 2013; Gevrekci 2017). In the genome of S. rhizophila strain IS26, the genes encode for S-adenosylmethionine synthase, S-adenosyl-methionine decarboxylase, and spermidine synthase may lead to the transformation of methionine to polyamines (Jang et al. 2012). These findings are in agreement with results reported by Pinski et al. (2020), who detected several genes coding for polyamine biosynthesis in the genome of S. rhizophila.
Other genes encoding key molecules related to general stress (such as ROS metabolism: superoxide dismutase, catalase, polyphenol oxidase, glutathione S-transferase, stress response kinase A, peroxidases and reductases) were also detected, suggesting involvement of indirect PGP mechanisms mostly related to antioxidant defence and osmotic balance recovery. The presence of these genes indicates the strain’s significant potential to reduce the accumulation of harmful ROS, which are known to be generated by the plant in response to an induced stress, especially osmotic stress (Sharma et al. 2012). In fact, accumulation of these ROS, namely H2O2, O−2 and OH−, is known to damage the DNA/RNA, oxidize proteins and lipids, and destroy plant chlorophyll and meristem activity (Foreman et al. 2003; Habib et al. 2016). Bacteria that exhibit effective enzymatic antioxidant systems have already been proven to play an important protective role against the effects of stress-induced ROS (Vurukonda et al. 2016). Afridi et al. (2019) reported involvement of bacterial antioxidant enzymes that conferred a significant salt-stress resistance on wheat after inoculation with a fuel biodegrader and salt-tolerant PGP Kocuria rhizophila strain. However, reports on PGPB-induced salinity tolerance of tomato plants in connection with changes in ROS-scavenging enzymes status remain scarce. Through biosynthetic gene cluster analysis by antiSMASH, the strain IS26 was predicted to produce potential free radical scavengers such as aryl polyenes (Deng et al. 2018). These pigments function as carotenoids able to protect from ROS, thus reducing potential oxidative stress-related cell damage (Schöner et al. 2016), and may play significant roles in plant growth-promotion by enhancing tolerance to environmental stresses such as salt (Lee et al. 2019).
The antiSMASH analysis also predicts production of various antimicrobial compounds that merit chemical investigation as potentially interesting biomolecules. The strain’s significant potential for the biosynthesis of antimicrobial compounds suggests an ability to compete with intense microbiological activity in the soil, thereby enhancing strain maintenance and conferring putative plant protection against pathogens (Hibbing et al. 2010).
The analysis of strain genomic data revealed the presence of numerous genes potentially involved in salt stress sensing and mitigation. Multiple salt-induced-stress sensing genes were identified, such as osmolarity sensor protein EnvZ, stress response protein NhaX, sensor histidine kinase, as well as the previously reported osmolyte biosynthesis genes such as trehalose, glucosylglycerol, proline and glycine betaine (Wolf et al. 2002; Roder et al. 2005). These osmoprotectant low-mass organic compounds have been shown to mediate osmotic adjustment, free radical scavenging and subcellular structure stabilization (Slama et al. 2015).
Amongst the Stenotrophomonas species studied, S. rhizophila is regarded as possessing a specific and remarkable mechanism for bacterial stress protection through its production and excretion of glucosylglycerol (Alavi et al. 2013). Furthermore, the key genes betA and betB for glycine-betaine synthesis, which respectively encode choline dehydrogenase and betaine aldehyde dehydrogenase, are considered to be among the most effective genes responsible for PGP-conferred salt tolerance in plants (Alavi et al. 2014).
Conclusions
In this study, Sahara-inhabiting halophytes Atriplex halimus and Lygeum spartum have been used as an effective source of associated halotolerant PGPB that can be applied as crop inoculants because of the potentialities they exhibit in terms of osmoprotection and plant growth enhancement. In the context of enhancing technical progress for sustainable agriculture, this approach would help us to consider profitable eco-friendly cultivation of non-halophytic plants, such as tomatoes, in saline soil environments.
The successive evaluation of the bacterial isolates for their halotolerance and PGP-induced performances in tomato seedlings grown under salt stress conditions, pointed to the isolate IS26 as the most promising (Fig. 7). Based on its 16S rRNA gene sequence, the strain was related to Stenotrophomonas rhizophila.

Effect of strain IS26 inoculation on the growth of tomato plant under non-stressed and NaCl-stressed conditions. Photographs were taken 15 days after planting
The strain genome analysis revealed the presence of numerous genes involved in mechanisms responsible for rhizosphere colonization, plant association, and strong competition for nutrients and niches, as well as the production of important plant growth regulating substances, some of which also act as protectors against stress. By possessing genes related to complementary mechanisms such as osmosensing, osmoregulation and osmoprotection, the strain definitely confirmed its remarkable skills for adapting to and tolerating highly saline conditions. The strain’s putative ability to produce osmolytes and the potential involvement of multiple enzymatic antioxidant systems in case of deleterious stress-induced overproduction of ROS were considered among the most effective salt mitigation processes.
With regard to the interesting potentialities exhibited in planta, brought out by specific genome analysis, the strain Stenotrophomonas rhizophila IS26 would merit extended evaluation at field level in diverse salt-stressed contexts to confirm its beneficial potential as a PGP and stress protecting agent in the perspective of a bioformulation study.
Data availability
All generated data are available.
Code availability
Not applicable.
References
Abdul-Baki AA, Anderson JD (1973) Vigor determination in soybean seed by multiple criteria 1. Crop Sci 13(6):630–633. https://doi.org/10.2135/cropsci1973.0011183x001300060013x
Abo-Kassem EE (2007) Effects of salinity: Calcium interaction on growth and nucleic acid metabolism in five species of Chenopodiaceae. Turk J Bot 31(2):125–134
Afridi MS, Mahmood T, Salam A, Mukhtar T, Mehmood S, Ali J, Bibi M, Javed MT, Sultan T, Chaudhary HJ (2019) Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: Involvement of ACC deaminase and antioxidant enzymes. Plant Physiol Biochem 139:569–577. https://doi.org/10.1016/j.plaphy.2019.03.041
Alavi P, Starcher MR, Zachow C, Müller H, Berg G (2013) Root-microbe systems: the effect and mode of interaction of Stress Protecting Agent (SPA) Stenotrophomonas rhizophila DSM14405(T.). Front Plant Sci 4:141. https://doi.org/10.3389/fpls.2013.00141
Alavi P, Starcher MR, Thallinger GG, Zachow C, Müller H, Berg G (2014) Stenotrophomonas comparative genomics reveals genes and functions that differentiate beneficial and pathogenic bacteria. BMC Genom 15:482. https://doi.org/10.1186/1471-2164-15-482
Alexander A, Singh VK, Mishra A (2020) Halotolerant PGPR Stenotrophomonas maltophilia BJ01 induces salt tolerance by modulating physiology and biochemical activities of Arachis hypogaea. Front Microbiol 11:2530. https://doi.org/10.3389/fmicb.2020.568289
Al-Karaki G (2000) Growth of mycorrhizal tomato and mineral acquisition under salt stress. Mycorrhiza 10(2):51–54. https://doi.org/10.1007/s005720000055
Andrés-Barrao C, Lafi FF, Alam I, de Zélicourt A, Eida AA, Bokhari A, Alzubaidy H, Bajic VB, Hirt H, Saad MM (2017) Complete genome sequence analysis of Enterobacter sp. SA187, a plant multi-stress tolerance promoting endophytic bacterium. Front Microbiol 8:2023. https://doi.org/10.3389/fmicb.2017.02023
Arp HPH, Knutsen H (2020) Could we spare a moment of the spotlight for persistent, water-soluble polymers? Environ Sci Technol 54:3–5. https://doi.org/10.1021/acs.est.9b07089
Balsanelli E, Tuleski TR, De Baura VA, Yates MG, Chubatsu LS, De Oliveira PF, De Souza EM, Monteiro RA (2013) Maize root lectins mediate the interaction with Herbaspirillum seropedicae via N-acetyl Glucosamine residues of Lipopolysaccharides. PLoS ONE. https://doi.org/10.1371/journal.pone.0077001
Bashan Y, Holguin G, Lifshitz R (1993) Isolation and characterization of plant growth-promoting rhizobacteria. In: Glick BR, Thompson JE (eds) Methods in Plant Molecular Biology and Biotechnology. CRC Press, Boca Raton, FL, USA, pp 331–345
Baxter A, Mittler R, Suzuki N (2014) ROS as key players in plant stress signalling. J Exp Bot 65(5):1229–1240. https://doi.org/10.1093/jxb/ert375
Berg G, Martinez JL (2015) Friends or foes: can we make a distinction between beneficial and harmful strains of the Stenotrophomonas maltophilia complex? Front Microbiol 6:241. https://doi.org/10.3389/fmicb.2015.0024
Berg G, Alavi M, Schmidt CS, Zachow C, Egamberdieva D, Kamilova F, Lugtenberg BJ (2013) Biocontrol and Osmoprotection for plants under Salinated conditions. Mol Microbial Ecol Rhizosphere. https://doi.org/10.1002/9781118297674.ch55
Berg G, Egamberdieva D, Lugtenberg B, Hagemann M (2010) Symbiotic Plant–Microbe Interactions: Stress Protection, Plant Growth Promotion, and Biocontrol by Stenotrophomonas. Cellular Origin, Life in Extreme Habitats and Astrobiology 17: 445–460.
Bhattacharyya PN, Jha DK (2012) Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J Microb Biot 28(4):1327–1350. https://doi.org/10.1007/s11274-011-0979-9
Bojović B, Delić G, Topuzović M (2010) Effects of NaCl on seed germination in some species from families Brassicaceae and Solanaceae. Kragujevac J Sci 32:83–87
Bolger AM, Lohse M, Usadel B (2014) Trimmomatic: A flexible Trimmer for Illumina sequence data. Bioinformatics 30(15):2114–2120. https://doi.org/10.1093/bioinformatics/btu170
Brooke JS (2012) Stenotrophomonas maltophilia: an emerging global opportunistic pathogen. Clin Microbiol Rev 25(1):2–41. https://doi.org/10.1128/CMR.00019-11
Cordero I, Balaguer L, Rincón A, Pueyo JJ (2018) Inoculation of tomato plants with selected PGPR represents a feasible alternative to chemical fertilization under salt stress. JPNSS 181(5):694–703. https://doi.org/10.1002/jpln.201700480
Cuartero J, Fernández-Muñoz R (1998) Tomato and Salinity. Sci Hortic 78(1–4):83–125. https://doi.org/10.1016/S0304-4238(98)00191-5
Cuin TA, Shabala S (2007) Amino acids regulate salinity-induced potassium efflux in barley root epidermis. Planta 225(3):753–761. https://doi.org/10.1007/s00425-006-0386-x
Damodaran T, Sah V, Rai RB, Sharma DK, Mishra VK, Jha SK, Kannan R (2013) Isolation of salt tolerant endophytic and rhizospheric bacteria by natural selection and screening for promising plant growth-promoting rhizobacteria (PGPR) and growth vigour in tomato under sodic environment. Afr J Microbiol Res 7(44): 5082–5089.
Deng K, Yang Z, Luo J, Wu K (2018) Synthesis of aryl and heterocyclic Polyenes and their activity in free radical scavenging. J Chem Res 42(3):129
Dif G, Belaouni HA, Goudjal Y, Yekkour A, Djemouai N, Zitouni A (2021) Potential for plant growth promotion of Kocuria arsenatis Strain ST19 on tomato under salt stress conditions. S Afr J Bot 138:94–104. https://doi.org/10.1016/j.sajb.2020.12.014
Duca D, Lorv J, Patten CL, Rose D, Glick BR (2014) Indole-3-acetic acid in plant-microbe interactions. Anton Leeuw 106:85–125. https://doi.org/10.1007/s10482-013-0095-y
Dworkin M, Foster JW (1958) Experiments with some microorganisms which utilize ethane and hydrogen1. J Bacteriol 75(5):592–603. https://doi.org/10.1128/jb.75.5.592-603.1958
Egamberdieva D, Jabborova D, Berg G (2016) Synergistic interactions between Bradyrhizobium japonicum and the endophyte Stenotrophomonas rhizophila and their effects on growth, and nodulation of soybean under salt stress. Plant Soil 405:35–45. https://doi.org/10.1007/s11104-015-2661-8
Eida AA, Hirt H, Saad MM (2017) Challenges faced in field application of phosphate-solubilizing bacteria. Rhizotrophs: plant growth promotion to bioremediation 125–143. doi: https://doi.org/10.1007/978-981-10-4862-3_6
Erbse AH, Falke JJ (2009) The core signaling proteins of bacterial chemotaxis assemble to form an ultrastable complex. Biochemistry 48(29):6975–6987. https://doi.org/10.1021/bi900641c
Etesami H, Beattie GA (2018) Mining halophytes for plant growth-promoting halotolerant bacteria to enhance the salinity tolerance of non-halophytic crops. Front Microbiol 9:148. https://doi.org/10.3389/fmicb.2018.00148
Etesami H, Maheshwari DK (2018) Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol Environ Saf 156:225–246. https://doi.org/10.1016/j.ecoenv.2018.03.013
Fan P, Chen D, He Y, Zhou Q, Tian Y, Gao L (2016) Alleviating salt stress in tomato seedlings using Arthrobacter and Bacillus megaterium isolated from the rhizosphere of wild plants grown on saline-alkaline lands. Int J Phytoremediation 18(11):1113–1121. https://doi.org/10.1080/15226514.2016.1183583
Finkelstein R (2013) Abscisic Acid synthesis and response. The Arabidopsis Book. https://doi.org/10.1199/tab.0166
Foreman J, Demidchik V, Bothwell JHF, Mylona P, Miedema H, Torres MA, Linstead P, Costa S, Brownlee C, Jones JD, Davies JM, Dolan L (2003) Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 422(6930):442–446. https://doi.org/10.1038/nature01485
Gamalero E, Bona E, Todeschini V, Lingua G (2020) Saline and arid soils: impact on bacteria, plants, and their interaction. Biology 9(6):116. https://doi.org/10.3390/biology9060116
García-Alcalde F, Okonechnikov K, Carbonell J, Cruz LM, Götz S, Tarazona S, Dopazo J, Meyer TF, Conesa A (2012) Qualimap: Evaluating next-generation sequencing alignment data. Bioinformatics 28(20):2678–2679. https://doi.org/10.1093/bioinformatics/bts503
Gerszberg A, Hnatuszko-Konka K (2017) Tomato tolerance to abiotic stress: a review of most often engineered target sequences. Plant Growth Regul 83:175–198. https://doi.org/10.1007/s10725-017-0251-x
Gevrekci AÖ (2017) The roles of polyamines in microorganisms. World J Microbiol Biotechnol 33(11):204. https://doi.org/10.1007/s11274-017-2370-y
Gill SS, Tuteja N (2010a) Polyamines and abiotic stress tolerance in plants. Plant Signal Behav 5(1):26–33. https://doi.org/10.4161/psb.5.1.10291
Gill SS, Tuteja N (2010b) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48(12):909–930. https://doi.org/10.1016/j.plaphy.2010.08.016
Glick BR, Todorovic B, Czarny J, Cheng Z, Duan J, McConkey B (2007) Promotion of plant growth by bacterial ACC deaminase. Crit Rev Plant Sci 26(5–6):227–242. https://doi.org/10.1080/07352680701572966
Gurevich A, Saveliev V, Vyahhi N, Tesler G (2013) QUAST: Quality assessment tool for genome assemblies. Bioinformatics 29(8):1072–1075. https://doi.org/10.1093/bioinformatics/btt086
Habib SH, Kausar H, Saud HM (2016) Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes. BioMed Res Int. https://doi.org/10.1155/2016/6284547
Hadfield J, Croucher NJ, Goater RJ, Abudahab K, Aanensen DM, Harris SR (2018) Phandango: an interactive viewer for bacterial population genomics. Bioinformatics 34(2):292–293. https://doi.org/10.1093/bioinformatics/btx610
Hallmann J, Quadt-Hallmann A, Mahaffee WF, Kloepper JW (1997) Bacterial endophytes in agricultural crops. Can J Microbiol 43:895–914. https://doi.org/10.1139/m97-131
Hall-Stoodley L, Costerton JW, Stoodley P (2004) Bacterial biofilms: from the Natural environment to infectious diseases. Nat Rev Microbiol 2(2):95–108. https://doi.org/10.1038/nrmicro821
Hamilton CE, Gundel PE, Helander M, Saikkonen K (2012) Endophytic mediation of reactive oxygen species and antioxidant activity in plants: a review. Fungal Diver 54(1):1–10. https://doi.org/10.1007/s13225-012-0158-9
Hardoim PR, Van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, Döring M, Sessitsch A (2015) The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol R 79(3):293–320. https://doi.org/10.1128/mmbr.00050-14
Hasanuzzaman M, Mahmud JA, Anee TI, Nahar K, Islam MT (2018) Drought stress tolerance in wheat: Omics approaches in understanding and enhancing antioxidant defense. Abiotic Stress-Mediated Sensing and Signalling in Plants: An Omics Perspective 267–307.
Hibbing ME, Fuqua C, Parsek MR, Peterson SB (2010) Bacterial competition: surviving and thriving in the microbial jungle. Nat Rev Microbiol 8(1):15–25. https://doi.org/10.1038/nrmicro2259
Hicks G, Jia Z (2018) Structural basis for the lipopolysaccharide export activity of the bacterial lipopolysaccharide transport system. Int J Mol Sci 19(9):2680. https://doi.org/10.3390/ijms19092680
Hu QP, Xu JG (2011) A simple double-layered chrome azurol S agar (SD-CASA) plate assay to optimize the production of siderophores by a potential biocontrol agent Bacillus. Afr J Microbiol Res 5(25):4321–4327. https://doi.org/10.5897/AJMR11.238
Huedo P, Coves X, Daura X, Gibert I, Yero D (2018) Quorum sensing signaling and quenching in the multidrug-resistant pathogen Stenotrophomonas maltophilia. Front Cell Infect Microbiol 8:122. https://doi.org/10.3389/fcimb.2018.00122
Islam F, Yasmeen T, Ali Q, Ali S, Arif MS, Hussain S, Rizvi H (2014) Influence of Pseudomonas aeruginosa as PGPR on oxidative stress tolerance in wheat under Zn stress. Ecotox Environ Safe 104:285–293. https://doi.org/10.1016/j.ecoenv.2014.03.008
Islam F, Yasmeen T, Arif MS, Ali S, Ali B, Hameed S, Zhou W (2015) Plant growth promoting bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul 80(1):23–36. https://doi.org/10.1007/s10725-015-0142y
IUSS Working Group WRB (2015) World Reference Base for soil resources 2014. International soil classification system for naming soils and creating legends for soil maps. Update 2015. World Soil Resources Report 106. FAO, Rome, p 188
Jaiswal DK, Verma JP, Krishna R, Gaurav AK, Yadav J (2019) Molecular characterization of monocrotophos and chlorpyrifos tolerant bacterial strain for enhancing seed germination of vegetable crops. Chemosphere 223:636–650. https://doi.org/10.1016/j.chemosphere.2019.02.053
Jang SJ, Wi SJ, Choi YJ, An G, Park KY (2012) Increased polyamine biosynthesis enhances stress tolerance by preventing the accumulation of reactive oxygen species: T-DNA mutational analysis of Oryza sativa lysine decarboxylase-like protein 1. Mol Cells 34(3):251–262. https://doi.org/10.1007/s10059-012-0067-5
Khamna S, Yokota A, Peberdy JF, Lumyong S (2010) Indole-3-acetic acid production by Streptomyces Sp. isolated from some Thai medicinal plant rhizosphere soils. Eurasia J Biosci. https://doi.org/10.5053/ejobios.2010.4.0.4
Khan, M. A., Boër, B., Ȫzturk, M., Clüsener-Godt, M., Gul, B., & Breckle, S. W. (Eds.). (2016). Sabkha Ecosystems: Volume V: The Americas (Vol. 48). Springer.
Khanra N, Rossi P, Economou A, Kalodimos CG (2016) Recognition and targeting mechanisms by chaperones in flagellum assembly and operation. PNAS 113(35):9798–9803. https://doi.org/10.1073/pnas.1607845113
Kim O, Cho Y, Lee K, Yoon S, Kim M, Na H, Park S, Jeon YS, Lee J, Yi H, Won S, Chun J (2012) Introducing EzTaxon-E: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int J Syst Evol Micr 62:716–721. https://doi.org/10.1099/ijs.0.038075-0
Kim K, Jang YJ, Lee SM, Oh BT, Chae JC, Lee KJ (2014) Alleviation of salt stress by Enterobacter sp. EJ01 in tomato and Arabidopsis is accompanied by up-regulation of conserved salinity responsive factors in plants. Mol Cells 37(2):109–117. https://doi.org/10.14348/molcells.2014.2239
Kumar PA, Reddy NN, Lakshmi NJ (2017) Screening tomato genotypes for salt tolerance. Int J Curr Microbiol App Sci 6(11):1037–1049. https://doi.org/10.20546/ijcmas.2017.611.121
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35(6):1547–1549. https://doi.org/10.1093/molbev/msy096
Kumar A, Soni R, Kanwar SS, Pabbi S (2019) Stenotrophomonas: A versatile diazotrophic bacteria from the rhizospheric soils of Western Himalayas and development of its liquid biofertilizer formulation. Vegetos 32(1):103–109
Kumari S, Vaishnav A, Jain S, Varma A, Choudhary DK (2015) Bacterial-mediated induction of systemic tolerance to salinity with expression of stress alleviating enzymes in soybean (Glycine max L. Merrill). J Plant Growth Regul 34(3):558–573. https://doi.org/10.1007/s00344-015-9490-0
Landa BB, Navas-Cortés JA, Hervás A, Jiménez-Díaz RM (2001) Influence of temperature and inoculum density of Fusarium oxysporum f. sp. ciceris on suppression of Fusarium wilt of chickpea by rhizosphere bacteria. Phytopathology 91(8):807–816. https://doi.org/10.1094/PHYTO.2001.91.8.807
Langmead B, Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2. Nat Methods 9(4):357–359. https://doi.org/10.1038/nmeth.1923
Lee J, Nagano Y, Taylor JP, Lim KL, Yao T (2010) Disease-causing mutations in Parkin impair mitochondrial ubiquitination, aggregation, and HDAC6-dependent mitophagy. J Cell Biol 189(4):671–679. https://doi.org/10.1083/jcb.201001039
Lee SA, Kanth BK, Kim HS, Kim TW, Sang MK, Song J, Weon HY (2019) Complete genome sequence of the plant growth-promoting endophytic bacterium Rhodanobacter glycinis T01E–68 isolated from tomato (Solanum lycopersicum L.) plant roots. The Microbiological Society of Korea 55(4):422–424. https://doi.org/10.7845/kjm.2019.9115
Letunic I, Bork P (2019) Interactive Tree Of Life (iTOL) v4: recent updates and new developments. Nucleic Acids Res 47(W1):W256–W259. https://doi.org/10.1093/nar/gkz239
Liu D, Coloe S, Baird R, Pederson J (2000) Rapid mini-preparation of fungal DNA for PCR. J Clin Microbiol 38(1):471–471
Liu F, Liu H, Zhou H, Dong Z, Bai X, Bai P, Qiao J (2014) Isolation and characterization of phosphate-solubilizing bacteria from betel nut (Areca catechu) and their effects on plant growth and phosphorus mobilization in tropical soils. Biol Fert Soils 50(6):927–937. https://doi.org/10.1007/s00374-014-0913-z
Lugtenberg B, Kamilova F (2009) Plant-growth-promoting rhizobacteria. Annu Rev Microbiol 63:541–556. https://doi.org/10.1146/annurev.micro.62.081307.162918
MacDonald LC, Berger BW (2014) A polysaccharide lyase from Stenotrophomonas maltophilia with a unique, pH-regulated substrate specificity. J Biol Chem 289(1):312–325. https://doi.org/10.1074/jbc.M113.489195
Magán J, Gallardo M, Thompson R, Lorenzo P (2008) Effects of salinity on fruit yield and quality of tomato grown in soil-less culture in greenhouses in Mediterranean climatic conditions. Agric Water Manag 95(9):1041–1055. https://doi.org/10.1016/j.agwat.2008.03.011
Marquina A (2012) Environmental Challenges in the Mediterranean 2000–2050: Proceedings of the NATO Advanced Research Workshop on Environmental Challenges in the Mediterranean 2000–2050 Madrid, Spain 2–5 October 2002 (Vol. 37). Springer Science & Business Media
Mayak S, Tirosh T, Glick BR (2004) Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol Biochem 42(6):565–572. https://doi.org/10.1016/j.plaphy.2004.05.009
Metoui Ben Mahmoud O, Hidri R, Talbi-Zribi O, Taamalli W, Abdelly C, Djébali N (2020) Auxin and proline producing rhizobacteria mitigate salt-induced growth inhibition of Barley plants by enhancing water and nutrient status. SAJB 128:209–217. https://doi.org/10.1016/j.sajb.2019.10.023
Moore ER, Krüger AS, Hauben L, Seal SE, Baere RD, Wachter RD, Timmis KN, Swings J (1997) 16S rRNA gene sequence analyses and inter- and intrageneric relationships of Xanthomonas species and Stenotrophomonas maltophilia. FEMS Microbiol Lett 151(2):145–153. https://doi.org/10.1111/j.1574-6968.1997.tb12563.x
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
De Oliveira-Garcia D, Dall’Agnol M, Rosales M, Azzuz AC, Alcántara N, Martinez MB, Girón JA (2003) Fimbriae and adherence of Stenotrophomonas maltophilia to epithelial cells and to abiotic surfaces. Cell Microbiol 5(9): 625–636.
Page AJ, Cummins CA, Hunt M, Wong VK, Reuter S, Holden MT, Fookes M, Falush D, Keane JA, Parkhill J (2015) Roary: rapid large-scale prokaryote pan-genome analysis. Bioinformatics 31(22):3691–3693. https://doi.org/10.1093/bioinformatics/btv421
Passari AK, Mishra VK, Gupta VK, Yadav MK, Saikia R, Singh BP (2015) In vitro and in vivo plant growth promoting activities and DNA fingerprinting of antagonistic endophytic actinomycetes associates with medicinal plants. PLoS ONE. https://doi.org/10.1371/journal.pone.0139468
Paul D, Lade H (2014) Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron Sustain Dev 34(4):737–752. https://doi.org/10.1007/s13593-014-0233-6
Piernik A, Hrynkiewicz K, Wojciechowska A, Szymańska S, Lis MI, Muscolo A (2017) Effect of halotolerant endophytic bacteria isolated from Salicornia europaea L. on the growth of fodder beet (Beta vulgaris L.) under salt stress. Arch Agron Soil Sci 63(10):1404–1418. https://doi.org/10.1080/03650340.2017.1286329
Pikovskaya RI (1948) Mobilization of phosphorus in soil connection with the vital activity of some microbial species. Microbiology 17:362–370
Pinski A, Zur J, Hasterok R, Hupert-Kocurek K (2020) Comparative genomics of Stenotrophomonas maltophilia and Stenotrophomonas rhizophila revealed characteristic features of both species. Int J Mol Sci 21(14):4922. https://doi.org/10.3390/ijms21144922
Rainey FA, Ward-Rainey N, Kroppenstedt RM, Stackebrandt E (1996) The genus Nocardiopsis represents a phylogenetically coherent taxon and a distinct actinomycete lineage: proposal of Nocardiopsaceae fam. nov. Int J Syst Bacteriol 46:1088–1092
Ramakrishna W, Yadav R, Li K (2019) Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl Soil Ecol 138:10–18. https://doi.org/10.1016/j.apsoil.2019.02.019
Reiner K (2010) Catalase test protocol. ASM Microbe Library. https://www.asmscience.org/content/education/protocol/protocol.3226.
Reinhardt EL, Ramos PL, Manfio GP, Barbosa HR, Pavan C, Moreira-Filho CA (2008) Molecular characterization of nitrogen-fixing bacteria isolated from brazilian agricultural plants at São Paulo state. Braz J Microbiol 39(3):414–422. https://doi.org/10.1590/S1517-83822008000300002
Roder A, Hoffmann E, Hagemann M, Berg G (2005) Synthesis of the compatible solutes glucosylglycerol and trehalose by salt-stressed cells of Stenotrophomonas strains. FEMS Microbiol Lett 243(1):219–226. https://doi.org/10.1016/j.femsle.2004.12.005
Rodríguez-Llorente ID, Pajuelo E, Navarro-Torre S, Mesa-Marín J, Caviedes MA (2019) Bacterial endophytes from halophytes: how do they help plants to alleviate salt stress? In: Kumar M, Etesami H, Kumar V (eds) Saline Soil-based Agriculture by Halotolerant Microorganisms. Springer, Singapore, pp 147–160
Römling U, Galperin MY (2015) Bacterial cellulose biosynthesis: Diversity of operons, subunits, products, and functions. Trends Microbiol 23(9):545–557. https://doi.org/10.1016/j.tim.2015.05.005
Rueden CT, Schindelin J, Hiner MC, DeZonia BE, Walter AE, Arena ET, Eliceiri KW (2017) Image J2: ImageJ for the next generation of scientific image data. BMC Bioinform 18(1):529. https://doi.org/10.1186/s12859-017-1934-z
Ryan RP, Monchy S, Cardinale M, Taghavi S, Crossman L, Avison MB, Berg G, Van der Lelie D, Dow JM (2009) The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat Rev Microbiol 7:514–525. https://doi.org/10.1038/nrmicro2163
Ryan RP, An SQ, Allan JH, McCarthy Y, Dow JM (2015) The DSF family of cell-cell signals: An expanding class of bacterial virulence regulators. PLoS Pathog. https://doi.org/10.1371/journal.ppat.1004986
Schmidt CS, Alavi M, Cardinale M, Müller H, Berg G (2012) Stenotrophomonas rhizophila DSM14405T promotes plant growth probably by altering fungal communities in the rhizosphere. Biol Fertil Soils 48:947–960. https://doi.org/10.1007/s00374-012-0688-z
Schöner TA, Gassel S, Osawa A, Tobias NJ, Okuno Y, Sakakibara Y, Shindo K, Sandmann G, Bode HB (2016) Aryl polyenes, a highly abundant class of bacterial natural products, are functionally related to antioxidative carotenoids. Chem Bio Chem 17(3):247–253. https://doi.org/10.1002/cbic.201500474
Schulz B, Wanke U, Draeger S, Aust H (1993) Endophytes from herbaceous plants and shrubs: Effectiveness of surface sterilization methods. Mycol Res 97(12):1447–1450. https://doi.org/10.1016/s0953-7562(09)80215-3
Seemann T (2014) Prokka: Rapid prokaryotic genome annotation. Bioinformatics 30(14):2068–2069. https://doi.org/10.1093/bioinformatics/btu153
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and Antioxidative defense mechanism in plants under stressful conditions. J Bot 2012:1–26. https://doi.org/10.1155/2012/217037
Sharma S, Kulkarni J, Jha B (2016) Halotolerant rhizobacteria promote growth and enhance salinity tolerance in peanut. Front Microbiol 7:1600. https://doi.org/10.3389/fmicb.2016.01600
Shen Y, Chen L, Wang M, Lin D, Liang Z, Song P, Yuan Q, Tang H, Li W, Duan K, Liu B, Zhao G, Wang Y (2017) Flagellar hooks and hook protein FlgE participate in host microbe interactions at immunological level. Sci Rep. https://doi.org/10.1038/s41598-017-01619-1
Shin W, Siddikee MA, Joe MM, Benson A, Kim K, Selvakumar G, Kang Y, Jeon S, Samaddar S, Chatterjee P, Walitang D, Chanratana M, Sa T (2016) Halotolerant plant growth promoting bacteria mediated salinity stress amelioration in plants. Korean J Soil Sci Fertil 49(4):355–367. https://doi.org/10.7745/KJSSF.2016.49.4.355
Shu S, Sun J, Guo S, Li J, Liu C, Wang C (2010) Effects of exogenous putrescine on PS II photochemistry and ion distribution of cucumber seedlings under salt stress. Acta Hortic Sinica 37(7):1065–1072
Siddikee MA, Glick BR, Chauhan PS, Wj Y, Sa T (2011) Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing 1-aminocyclopropane-1-carboxylic acid deaminase activity. Plant Physiol Biochem 49(4):427–434. https://doi.org/10.1016/j.plaphy.2011.01.015
Silambarasan S, Logeswari P, Ruiz A, Cornejo P, Kannan VR (2020) Influence of plant beneficial Stenotrophomonas rhizophila strain CASB3 on the degradation of diuron-contaminated saline soil and improvement of Lactuca sativa growth. Environ Sci Pollut 27(28):35195–35207. https://doi.org/10.1007/s11356-020-09722-z
Singh RP, Jha PN (2017) The PGPR Stenotrophomonas maltophilia SBP-9 augments resistance against biotic and abiotic stress in wheat plants. Front Microbiol 8:1945. https://doi.org/10.3389/fmicb.2017.01945
Slama I, Abdelly C, Bouchereau A, Flowers T, Savouré A (2015) Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann Bot 115:433–447. https://doi.org/10.1093/aob/mcu239
Springer B, Böttger EC, Kirschner P, Wallace RJ (1995) Phylogeny of the Mycobacterium chelonae-like organism based on partial sequencing of the 16S rRNA gene and proposal of Mycobacterium mucogenicum sp. nov. Int J Syst Evol Micr 45(2):262–267. https://doi.org/10.1099/00207713-45-2-262
Stapper AP, Narasimhan G, Ohman DE, Barakat J, Hentzer M, Molin S, Kharazmi A, Hoiby N, Mathee K (2004) Alginate production affects Pseudomonas aeruginosa biofilm development and architecture, but is not essential for biofilm formation. J Med Microbiol 53(Pt 7):679–690. https://doi.org/10.1099/jmm.0.45539-0
Tamreihao K, Ningthoujam DS, Nimaichand S, Singh ES, Reena P, Singh SH, Nongthomba U (2016) Biocontrol and plant growth promoting activities of a Streptomyces corchorusii strain UCR3–16 and preparation of powder formulation for application as biofertilizer agents for rice plant. Microbiol Res 192:260–270. https://doi.org/10.1016/j.micres.2016.08.005
Trobacher CP (2009) Ethylene and programmed cell death in plants. Botany 87(8):757–769
UNU-INWEH Annual Report (2014) United Nations University, Institute for Water, Environment and Health. http://inweh.unu.edu/unu-inwehs-2014-annual-report/.
Vardharajula S, Zulfikar Ali S, Grover M, Reddy G, Bandi V (2011) Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J Plant Interact 6(1):1–14. https://doi.org/10.1080/17429145.2010.535178
Veliz EA, Martínez-Hidalgo P, Hirsch AM (2017) Chitinase-producing bacteria and their role in biocontrol. AIMS Microbiol 3(3):689–705. https://doi.org/10.3934/microbiol.2017.3.689
Verbruggen N, Hermans C (2008) Proline accumulation in plants: A review. Amino Acids 35(4):753–759. https://doi.org/10.1007/s00726-008-0061-6
Vurukonda SS, Vardharajula S, Shrivastava M, SkZ A (2016) Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol Res 184:13–24. https://doi.org/10.1016/j.micres.2015.12.003
Weber T, Blin K, Duddela S, Krug D, Kim HU, Bruccoleri R, Lee SY, Fischbach MA, Müller R, Wohlleben W, Breitling R, Takano E, Medema MH (2015) AntiSMASH 3.0-a comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res 43(W1):W237–W243. https://doi.org/10.1093/nar/gkv437
Wick RR, Judd LM, Gorrie CL, Holt KE (2017) Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol. https://doi.org/10.1371/journal.pcbi.1005595
Willis LM, Stupak J, Richards MR, Lowary TL, Li J, Whitfield C (2013) Conserved glycolipid Termini in capsular polysaccharides synthesized by ATP-binding cassette transporter-dependent pathways in Gram-negative pathogens. PNAS 110(19):7868–7873. https://doi.org/10.1073/pnas.1222317110
Wolf A, Fritze A, Hagemann M, Berg G (2002) Stenotrophomonas rhizophila sp. nov., a novel plant-associated bacterium with antifungal properties. Int J Syst Evol Microbiol 52(Pt 6):1937–1944. https://doi.org/10.1099/00207713-52-6-1937
Wu Y, Wang Y, Li J, Hu J, Chen K, Wei Y, Bazhanov DP, Bazhanova AA, Yang H (2015) Draft Genome Sequence of Stenotrophomonas maltophilia Strain B418, a Promising Agent for Biocontrol of Plant Pathogens and Root-Knot Nematode. Genome Announc. https://doi.org/10.1128/genomeA.00015-15
Yao L, Wu Z, Zheng Y, Kaleem I, Li C (2010) Growth promotion and protection against salt stress by Pseudomonas putida Rs-198 on cotton. Eur J Soil Biol 46(1):49–54. https://doi.org/10.1016/j.ejsobi.2009.11.002
Zahir ZA, Ghani U, Naveed M, Nadeem SM, Asghar HN (2009) Comparative effectiveness of Pseudomonas and Serratia sp. containing ACC-deaminase for improving growth and yield of wheat (Triticum aestivum L.) under salt-stressed conditions. Arch Microbiol 191(5):415–424. https://doi.org/10.1007/s00203-009-0466-y
Zamoum M, Goudjal Y, Sabaou N, Barakate M, Mathieu F, Zitouni A (2015) Biocontrol capacities and plant growth-promoting traits of endophytic Actinobacteria isolated from native plants of Algerian Sahara. JPDP 122(5–6):215–223. https://doi.org/10.1007/bf03356555
Zhang G, Meredith TC, Kahne D (2013) On the essentiality of lipopolysaccharide to Gram-negative bacteria. Curr Opin Microbiol 16(6):779–785. https://doi.org/10.1016/j.mib.2013.09.007
Zhumakayev AR, Vörös M, Szekeres A, Rakk D, Vágvölgyi C, Szűcs A, László Kredics L, Škrbić BD, Hatvani L (2021) Comprehensive characterization of stress tolerant bacteria with plant growth-promoting potential isolated from glyphosate-treated environment. World J Microbiol Biotechnol 37(6):1–17. https://doi.org/10.1007/s11274-021-03065-8
Acknowledgements
We are grateful to the late Prof. N. Sabaou (LBSM, Ecole Normale Supérieure de Kouba, Algiers, Algeria) for his valuable advice and continuous assistance.
Funding
This work was also supported by the Direction Générale de la Recherche Scientifique et du Développement Technologique (DGRSDT) of the Ministère de l’Enseignement Supérieur et de la Recherche Scientifique (MESRS) of Algeria and the MŠMT project CZ.02.1.01/0.0/0.0/16_017/0002334 and TAČR TJ02000096, Czech Republic.
Author information
Authors and Affiliations
Contributions
G. Dif and H. A. Belaouni: conducting the experiments, genome analysis/mining and comparative genomics, discussion of the results and preparation of the manuscript; A. Yekkour: supervision of the experiments, management of results, data analysis, discussion of the results and preparation of the manuscript; Y. Goudjal and N. Djemouai: assistance in endophytic bacteria isolation and 16 s DNA-based identification; E. Peňázová, J. Pečenka and A. Eichmeier: assistance in strain-complete genome sequencing and preliminary analysis with provision of the facilities as well as revision of the manuscript; A. Berraf-Tebbal: plant trial supervision, technical advice for bacterial PGP traits characterization, and revision of the manuscript; A. Zitouni: design of experiments, supervision and revision of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
Not applicable.
Consent to participate
All co-authors consented to participate in this work.
Consent for publication
All co-authors consent for publication.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Dif, G., Belaouni, H.A., Yekkour, A. et al. Performance of halotolerant bacteria associated with Sahara-inhabiting halophytes Atriplex halimus L. and Lygeum spartum L. ameliorate tomato plant growth and tolerance to saline stress: from selective isolation to genomic analysis of potential determinants. World J Microbiol Biotechnol 38, 16 (2022). https://doi.org/10.1007/s11274-021-03203-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11274-021-03203-2