
Abstract
The Cerrado, a Neotropical savanna, is the second largest vegetation domain in Brazilian territory and presents a gradient of physiognomies. Among the savanna formations, the cerrado stricto sensu stands out. The responses of woody species of the cerrado related to increased CO2 concentration ([CO2]), as predicted by global climate change, may differ among distinct leaf persistence groups. Elevated [CO2] could enhance the resprouting capacity of woody species, which might intensify encroachment in grasslands, such as the cerrado stricto sensu. We investigated the influence of elevated [CO2] (eCO2) on vegetative growth, root non-structural carbohydrates (NSC), and biomass allocation pattern among roots, stems, and leaves in two deciduous and three evergreen cerrado woody species. We carried out the experiment in open-top chambers with five young individuals of cerrado woody species under ambient [CO2] (aCO2) and eCO2 of 430 and 700 ppm, respectively. Measurements occurred in young plants with 294, 379, and 466 days old, i.e., after 104, 189, and 276 days of eCO2 exposition. While the species under aCO2 decreased the NSC in roots on the 189th day, the plants under eCO2 did not show a decrease over time. The deciduous species under eCO2 showed higher RGR at the beginning of the experiment (104 days) than the evergreens species under eCO2. Also, the eCO2 promoted fast leaf development in four out of five studied species (except Anadenanthera peregrina). The characteristics analyzed among the species could demonstrate that the deciduous grown under eCO2 presented higher relative growth rate and root/stem ratio than the species with evergreen habit.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Human activities have increased CO2 concentration [CO2] in the atmosphere, intensifying global climate change (IPCC 2019). Rising [CO2] and atmospheric warming would change species distribution and abundance (Siqueira and Peterson 2003; Van der Putten et al. 2010; Bellard et al. 2012; Peterson et al. 2019; Pecl et al. 2017; Nunez et al. 2019), and regional variations in productivity patterns (Nowak et al. 2004; Newingham et al. 2013). In C3 woody species, it is expected that one of the main effects of increase in [CO2] occurs close to the light compensation point of photosynthesis, making plants more efficient at low light intensity (Ainsworth and Long 2005). Increases are expected in the light-driven photochemical reactions in C3 species growing under elevated (eCO2), when photosynthesis is limited by the amount of active Rubisco (Li et al. 2009). In addition, in C3 species growing under eCO2 an increase in water use efficiency, decreased transpiration, and stomatal conductance is expected (Ainsworth and Rogers 2007). Under appropriate water and nutrient availability in the soil, the increase in [CO2] promotes the growth of woody species (Ainsworth and Long 2005; Souza et al. 2016). In a general context, woody species growing under eCO2 would show increased root growth, a higher concentration of non-structural carbohydrates in the roots, and improved ability to resprout after disturbances (Hoffman et al. 2000; Kgope et al. 2010; Manea and Leishman 2019) than plants growing under ambient [CO2] (aCO2). Therefore, it is essential to investigate the effects caused by climate change, mainly the increase in [CO2], on the resprouting potential of cerrado woody species.
Resprout capacity is an essential functional trait determining plant responses and vegetation dynamics after disturbances, such as fire, which shape most savannas (Clarke et al. 2016). The capacity to resprout after the fire is related to the accumulation of carbohydrates and nutrient reserves that need recharging between burn events (Hoffman et al. 2000; Franco et al. 2014). In response to the disturbance, plants can die because they do not have the capacity or resources to replace the foliage through protected meristems, or they can resprout using underground reserves in the roots, such as the non-structural carbohydrates and nitrogen (Clarke et al. 2013). The atmospheric [CO2] is known to have high effects on plant carbohydrate status (Korner and Miglietta 1994; Bassirirad et al. 1996; Read and Morgan 1996; Cruz et al. 1997). Furthermore, the increased [CO2] in the atmosphere would change plant resource allocation patterns and resprouting capacity (Clarke et al. 2016; Manea and Leishman 2019). Kgpoe et al. (2010) in a study with two Acacia species found an increased C allocation to non-structural carbohydrates storage along a [CO2] gradient. In addition, the resprouting ability and root non-structural carbohydrates of a tropical savanna tree, Kielmeyera coriacea Mart & Zucc., were significantly greater under eCO2 and nutrient enrichment (Hoffmann et al. 2000). Manea and Leishman (2019) analyzed the effect of increasing [CO2] on the resprout capacity of 16 woody and grass species from temperate regions. Woody and grass species grown under eCO2 showed an increase in biomass production. However, only woody species showed a higher accumulation of non-structural carbohydrates in roots under eCO2, and increased root biomass (Manea and Leishman 2019), indicating improved resprout capacity.
Savanna ecosystems cover about 20% of the terrestrial land surface (Kgope et al. 2010). The Cerrado, a Neotropical savanna, is the second largest vegetation domain in Brazilian territory and presents a gradient of physiognomies, ranging from grasslands to tree forests rich in endemic species (Coutinho 2006; Overbeck et al. 2015; Durigan and Ratter 2016). Among the savanna formations found in this vegetational domain (Batalha 2011), the cerrado stricto sensu stands out, being characterized by the abundance of woody species, with different degrees of leaf deciduity and by a herbaceous stratum rich in different growth forms (Eiten 1972). In Cerrado, the influence of climate change on vegetation dynamics is more dramatic because half of the original vegetation area has been converted by anthropic activities (Myers et al. 2000; Klink and Machado 2005; Franco et al. 2014; Lapola et al. 2014; Hughes 2017). The global climate change in the Cerrado could modify interspecific competition for limited resources (such as water and nutrients), changing the vegetation structure (Melo et al. 2018). Cerrado plant species are expected to lose more than half of their distribution range under the pessimistic scenario (RCP8.5) of climate change and lande use (Velazco et al. 2019). Understanding the response of Cerrado vegetation to climate change implies investigating the effect of eCO2 on the growth, development, and resprouting of woody species (Souza et al. 2016; Melo et al. 2018; Maia et al. 2019). These responses could be different between species grouped by leaf functional traits (Souza et al. 2009a, b, 2011; Borges and Prado 2018).
Interactions among photosynthesis, photorespiration, and respiration processes would likely define how much carbon is available for plant growth and resprouting after drought or fire events (Franco et al. 2014). Some studies have found that CO2 stimulates resprouting following defoliation (Fajer et al. 1991; Pearson and Brooks 1996; Wilsey et al. 1997; Maia et al. 2019). Oliveira et al. (2021), found a positive response to eCO2 in plants of Chrysolaena obovata (Less.) M. Dematt.,a C3 species native from cerrado. The plants grown under eCO2 showed higher growth, photosynthetic rate, aerial biomass production, underground rhizophore biomass, and consequently higher carbohydrate productivity than plants under aCO2. Souza et al. (2016) found higher biomass production, growth, leaf area ratio, and net photosynthesis in Hymenaea stigonocarpa Mart. ex Hayne, Solanum lycocarpum A. St.-Hil., and Tabebuia aurea (Silva Manso) Benth. & Hook. f. ex S. Moore, growing under eCO2. However, all these studies did not analyze the possible differential responses among species with distinct leaf persistence in cerrado vegetation due to climate change. Species with different leaf persistence could show different responses to eCO2 due to distinct ecological strategies. Deciduous woody species showed a more acquisitive strategy, maximizing uptake and use of resources (Ouédraogo et al. 2013; Souza et al. 2020). On the other hand, evergreens show a more conservative strategy to resource use (Ouédraogo et al. 2013; Souza et al. 2020). Thus, rising [CO2] could result in different ecophysiological responses among species with distinct leaf persistence.
The objective of the present study was to evaluate the influence of eCO2 on vegetative growth, biomass allocation, and resprouting potential in deciduous and evergreen woody species from Cerrado. We predicted that eCO2 would increase plant biomass, vegetative growth, and non-structural carbohydrates in the roots of all species. The higher non-structural carbohydrates allocation to the root system could increase the potential for resprouting of cerrado woody species, which would benefit them mainly after disturbance (Souza et al. 2016; Manea and Leishman 2019). These changes would help the establishment of cerrado woody species in native areas under the new climatic conditions. We also predicted a higher investment in the aerial growth of deciduous species grown under eCO2 as these species show faster shoot growth (synchronous leaf production) at the end of the dry season (Damascos et al. 2005; Damascos 2008; Souza et al. 2009a, b) to maximize growth in the next rainy season. However, evergreens could show few changes, as these species have an immediate consumption of the current resource throughout the year (Damascos et al. 2005; Damascos 2008; Souza et al. 2009a, b), being more conservative in the growth.
Materials and methods
Study area and open-top chambers
The experiment was carried out in open-top chambers (OTC, Figure S1) located at the Federal University of Viçosa, campus Florestal, Minas Gerais, Brazil (19º52´29´´ S and 44º25´12´´ W).We use OTC built with rectangular modules with side doors facilitating data collection (Silva 2010; Souza et al. 2016). The air intake in the chambers was controlled through an air injection system composed of a centrifugal fan, with a 20 cm diameter propeller and 50 mm diameter perforated PVC tubes. In this system, the air flow from the fan was directed into the chambers through PVC tubes. To enrich the atmosphere of the chambers with CO2 during the experiment, CO2 storage cylinders with a capacity of 25 kg each were used. The primary flow of CO2 into the chambers was regulated by a special pressure gauge (White Martins—R201C). The flow was regulated and directed to the chambers through high pressure hoses (500 psi). Plants were exposed to elevated [CO2] during the day (6:00 am to 6:00 pm) and monitored with a CO2 meter (Testo 535). Inside OTCs, fans worked during the day, avoiding excess moisture and temperature rise inside OTCs. The OTCs were monitored periodically by measuring air temperature (thermo-hygrometer, model INCOTERM 7666.02.0.00).
Plant material and soil properties
The five native species studied were classified according to their corresponding leaf persistence group (Souza et al. 2009a, b; Souza et al. 2011; Borges and Prado 2014): deciduous (DE) Anadenanthera peregrina var. falcata (Benth.) Altschul and Tabebuia aurea (Silva Manso) Benth & Hook. f. ex. Moore, and evergreen (EV) Hymenaea stigonocarpa Mart. Ex Hayne, Stryphnodendron adstringens Mart., and Stryphnodendron polyphyllum Mart.
Anadenanthera peregrina var. falcata (Benth.) Altschul, popularly known as angico-do-cerrado, belongs to the Fabaceae family and is a tree species that can reach 15 m in height (Silva et al. 2020). Tabebuia aurea (Manso) Benth. & Hook f. ex S. Moore, species native to Brazil belonging to the Bignoniaceae Family, occurs in the Amazon Region, Northeast, Midwest, and Southeast, in several vegetation formations, being popularly known as paratudo, caraíba, ipê-do-cerrado (Lorenzi 2002; Santos et al. 2019). Hymenaea stigonocarpa Mart.ex Hayne, known as jatobá-do-cerrado, jutaí, jatobá-capo, jatobá-de-cascafina, jitaí or jutaicica, being common in the open formations of the cerrado and campo cerrado (Lorenzi 1992). Stryphnodendron adstringens (Mart.), known as barbatimão, has a wide geographic distribution, occurring from Pará, through the central plateau, to Minas Gerais and São Paulo (Felfili et al. 1999). Stryphnodendron polyphyllum Mart. is a plant with 4–6 m high, deciduous, heliophyte, pioneer and characteristic of the cerrado, whose distribution extends from the cerrado of Central Brazil to Paraná and Mato Grosso do Sul (Lorenzi 2000).
The soil used for the species growth was typical of cerrado areas in the State of Minas Gerais (Haridasan et al. 2000). The chemical characteristics were: pH in H2O = 6.1; H + Al = 3.54 cmol.charge/dm3; Al3+ = 1.83 cmol.charge/dm3; Ca2+ = 0.33 cmol.charge/dm3; Mg2+ = 0.07 cmol.charge/dm3; P = 0.8 mg/dm3; K = 8 mg/dm3; Sum of bases = 0.42 cmol.charge/dm3; Cation exchange capacity = 3.96 cmol.charge/dm3; effective cation exchange capacity = 2.25 cmol.charge/dm3; Aluminum saturation index = 81.20%, and base saturation index = 10.68%. Soil analysis was carried out in the Agricultural Chemistry Laboratory of the Minas Gerais Institute of Agriculture (IMA).
Pre-germination treatment and experimental design
Seeds of A. peregrina, T. aurea, H. stigonocarpa, S. adstringens, and S. polyphyllum were treated with 2% sodium hypochlorite, for five minutes (De Lemos Filho et al. 1997; Botelho et al. 2000; Cabral et al. 2003; Miranda et al. 2012) and then washed with water. The following species were mechanically scarified to break the physical dormancy of seeds: H. stigonocarpa, S. adstringens, and S. polyphyllum. Seeds of all species germinated on filter paper, in a germination chamber Solab (SL.225). Seeds of H. stigonocarpa, S. adstringens, and T. aurea remained at alternating temperatures of 25–30 °C (night and day, respectively) and 12 h of photoperiod (Botelho et al. 2000; Cabral et al. 2003) and the seeds of S. polyphyllum and A. peregrina at constant light and temperatures of 30 °C (De Lemos Filho et al. 1997; Miranda et al. 2012). After germination, the seedlings were transferred to 8.0 L plastic pots filled with soil. The treatment with eCO2 started when the seedlings were 190 days old.
Young plants were assigned in two treatments with different [CO2]: 25 plants of each species divided in four OTC growing under elevated CO2 (eCO2, ~ 700 ± 5.10 ppm) and 25 plants of each species divided in four OTC growing under ambient CO2 (aCO2, ~ 430 ± 7.20 ppm, without CO2 enrichment).The plants in both CO2 treatments were daily watered.
Leaf area, and leaf, stem, and root dry mass
The morphological measurements were taken in four individuals of each species in each CO2 treatment in each measurement date (n = 4 individuals per species in each treatment). These measurements were carried out when the young plants were 294, 379, and 466 days old and growing under eCO2 for 104, 189, and 276 days. All leaves were separated from the stem and petioles and then scanned in black and white with the help of a table scanner to determine leaf area. Subsequently, the leaf area was calculated using the Image-Pro 5.0 software (Media Cybernetics, Inc., Silver Spring, MD, USA). The root length (measurement of the largest root) was determined with a 30 cm millimeter plastic ruler. The dry mass of each stem was determined after removing all buds and leaves when the leaf mass was also determined (the same leaves detached from the stems to leaf area determination). Leaves, stems, and roots were dried in an oven (Tecnal TE-394/3, Piracicaba, São Paulo, BR) at 60 °C for 72 h (Pérez-Harguindeguy et al. 2013) and had their masses measured using an analytical balance (Shimadzu- BL Series—20H, Tokyo, Japan). The total dry mass was the sum of the dry masses of root, stem, and leaves in the same individual.
Growth indices and vegetative morphometry
After determining the morphological traits, the following growth indices were calculated: relative growth rate (RGR, g g−1 days−1), leaf area index (LAI, cm2 days−1), leaf area ratio (LAR, total leaf area/total dry mass, cm2 g−1), specific leaf area (SLA, total leaf area/total leaf dry mass, cm2 g−1), and root/stem ratio (RSR, root dry mass/stem dry mass, g g −1) in four individuals of each species in each treatment and measurement date.
Relative growth rates (RGR, g g−1 days−1) were calculated using the following equation:
Leaf area index (LAI, cm2 days−1) were calculated using the following equation:
The leaf number (LN), stem length (SL, cm), and stem diameter (SD, cm) were monitored weekly throughout the experiment, up to 283 days after the beginning of the experiment (DBE) in 25 individuals of each species in each [CO2]. We analyzed the leaf development in five leaves from five individuals (n = 5 plants per species in each treatment) to determine the leaf expansion interval (LEI, days) and leaf expansion rate (LER, cm days−1). The length and width of each marked leaf (or leaflet) were monitored weekly. When the leaf stabilized the growth, the LEI was determined. Using the leaf expansion interval, we estimated LER by dividing the stabilized leaf area by the leaf expansion interval (LER = LEA/LEI).
Non-structural carbohydrates
The same individuals used for growth measures were used to quantify non-structural carbohydrates (NSC) in the roots using the sulfuric phenol method (Dubois et al. 1956). 0.5 g of root dry biomass was ground with liquid nitrogen, then mixed in ethanol 80% (5 ml) and incubated in a water bath at 75 °C for 30 min. Afterward, the sample was quickly placed on ice. Subsequently, the material was centrifuged at 3000×g for 15 min then the supernatant was collected. This procedure was performed twice. We collected 1 ml of the supernatant from the alcoholic extraction and incubated it in a water bath at 75 °C. The evaporates were hydrolyzed in 1 ml of distilled water, added 1 ml of hydrochloric acid (HCL 2.5N), and placed in a water bath at 100 °C for 60 min. After 60 min, this material was placed on ice for 10 min and centrifuged at 10000 rpm for 5 min. We collected 0.1 ml of extraction solution with new tubes with more 0.9 ml of deionized water and added 0.5 ml of 5% phenol solution and 2.5 ml of 96% sulfuric acid in each tube. After 10 min, the tube content was incubated in a water bath at 25–30 °C for 20 min. The absorbance reading was performed on a spectrophotometer (Shimadzu, Model UV-1800 120 V Tokyo, Japan) with a wavelength of 490 nm. We calculate the amount of total carbohydrate present in the sample solution using the standard graph.
Non-structural carbohydrates were calculated using the following equation:
Data analysis
Principal component analysis (PCA) was performed with all variables: total leaf area (TLA), root length (RL), leaf dry mass (LDM), stem dry mass (SDM), root dry mass (RDM), total dry mass (TDM), relative growth rate (RGR), leaf area index (LAI), leaf area ratio (LAR), specific leaf area (SLA), root/stem ratio (RSR), non-structural carbohydrates (NSC), leaf number (LN), stem length (SL), stem diameter (SD), leaf expansion interval (LEI), and leaf expansion rate (LER). For PCA analysis, we utilized the average values of each variable in every species and CO2 treatment.
We calculated the mean and standard deviation for all physiological, morphological, and morphometrical parameters. The Shapiro–Wilk test was used to evaluate the normal distribution in each dataset. The data which did not show normal distribution (RGR, NSC, LER) were log-transformed. Afterward, the Levene test was used to verify variance homogeneity. For TDM, RGR, LAI, SLA, RSR, and NSC, we used Mixed Design Analysis of Variance (MIXED ANOVA) with two factors and repeated measures, considering species and [CO2] between-subjects factors and time as within-subject factor. We used a posteriori Tukey’s posthoc test to investigate significant differences (p < 0.05) between the means of the species, [CO2], and time interval (days after the beginning of the experiment, DBE). For LER, we used Analysis of variance (two-way ANOVA), and a posteriori Tukey’s test was also applied to investigate significant differences (p < 0.05) between the means of the species and [CO2]. All analyses were performed in the R program, version 3.0.0 (R Core Team 2019).
Results
Principal component analysis
The ordination diagram resulting from the principal component analysis (PCA) summarized 77% total variation of the data in the first two dimensions (Dim1 and Dim2), with 45% and 32% variation explained by Dim1 and Dim2, respectively (Figure 1). EV H. stigonocarpa independent of [CO2] had a positive relationship with the characteristics: LDM, SDM, RDM, TDM, LEA, RGR, LAI, SLA, RSR, LAR, LN, SL, SD, and LEI and were distributed along Dim1 (Figure 1). The distribution of EV species S. polyphyllum and S. adstringens were more influenced by RL and LER (Figure 1). The PCA ordination diagram indicated that species were separated by growth characteristics in relation to CO2 concentration (~ 430 and ~ 700 ppm).

Principal component analysis (PCA) showing the distribution of all features according to the five Cerrado woody species growing under ambient (430 ppm) and elevated (700 ppm) CO2 concentrations in open-top chambers. LDM Leaf dry mass, SDM stem dry mass, RDM root dry mass, TDM total dry mass, TLA total leaf area, RL root length, RGR relative growth rate, LAI leaf area index, LAR leaf area ratio, SLA specific leaf area, RSR root/stem ratio, NSC non-structural carbohydrates, LN leaf number, SL stem length, SD stem diameter, LEI leaf expansion interval, and LER leaf expansion rate. 1 = A. peregrina 700 ppm, 2 = A. peregrina 430 ppm, 3 = H. stigonocapra 700 ppm, 4 = H. stigonocapra 430 ppm, 5 = S. adstringens 700 ppm, 6 = S. adstringens 430 ppm, 7 = S. polyphyllum 700 ppm, 8 = S. polyphyllum 430 ppm, 9 = T. aurea 700 ppm, 10 = T. aurea 430 ppm. Dimensions 1 and 2 are responsible for 45.4% and 31.8% of the total data variation, respectively
Leaf responses to eCO2
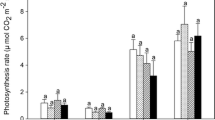
Deciduous A. peregrina and evergreen H. stigonocarpa independent of [CO2], showed a significant increase in the specific leaf area (SLA) at the middle of the experiment (189th day, Fig. 2a, F-test = 7.62, p < 0.05) than other species. Leaf area index (LAI) peaked at the middle of the experiment (189th day) under eCO2 compared to the beginning (104th day) and at the end of the experiment (276th day, Fig. 3b, F-test = 3.70, p < 0.05 interaction between CO2 and DBE). The species did not show significant differences in LAI between [CO2] in the same DBE (Fig. 3b). S. polyphyllum and S. adstringens plants in eCO2 showed higher leaf expansion rate (LER) than their plants in aCO2 (Table 1, F-test = 8.90, p < 0.05). Also, S. polyphyllum and S. adstringens in eCO2 showed higher LER than plants of other species in eCO2 (Table 1, F-test = 8.90, p < 0.05).

a Specific leaf area (SLA) in five Cerrado woody species growing under ambient (430 ppm) and elevated (700 ppm) CO2 concentrations in open-top chambers. Symbols represent averages (n = 4), and bars indicate standard deviation. Capital letters compare significant differences within the same species in different DBE. Lowercase letters compare significant differences between species in the same DBE (p < 0.05, Tukey’s test)

a NSC = non-structural carbohydrates and b LAI = leaf area index in five Cerrado woody species growing under ambient (430 ppm) and elevated (700 ppm) CO2 concentrations in open-top chambers. Symbols represent averages (n = 4), and bars indicate standard deviation. Capital letters compare significant differences within the same [CO2] in different DBE. Lowercase letters compare significant differences between [CO2] in the same DBE (p < 0.05, Tukey’s test)
Root responses to eCO2
There was a decrease in non-structural carbohydrates (NSC) at the middle of the experiment (189th day) under aCO2 (Fig. 2e, F-test < 0.0001, p < 0.05 interaction between CO2 and DBE). However, under eCO2 there were no significant differences in NSC across species over time (Fig. 3a). However, at the middle of the experiment (189th day) the species under eCO2 showed higher NSC (Fig. 3a, F-test < 0.0001, p < 0.05 interaction between CO2 and DBE) than under aCO2.
Individual-level responses to eCO2
All species showed greater total dry mass (TDM) at the middle of the experiment (189th day) remained higher at the end of the experiment (276th day, Fig. 4a, F-test = 23.06, p < 0.05), regardless of [CO2].

a TDM = total dry mass in five Cerrado woody species growing under ambient (430 ppm) and elevated (700 ppm) CO2 concentrations in open-top chambers. Symbols represent averages (n = 4), and bars indicate standard deviation. Capital letters compare significant differences between days after the beginning of the experiment (DBE) (p < 0.05, Tukey’s test)
The individuals of deciduous species, A. peregrina and T. aurea, under eCO2 at beginning of the experiment (104th day) showed higher relative growth rate (RGR, Fig. 5a) than their plants under eCO2 at the middle of the experiment (189th day). Furthermore, A. peregrina and T. aurea plants (Fig. 5a, F-test = 3.01, p < 0.05 interaction among CO2, species, and DBE) under eCO2 at beginning of the experiment (104th day) showed higher RGR than all species under eCO2 at beginning of the experiment (104th day). Also, A. peregrina and T. aurea plants (Fig. 5a, F-test = 3.01, p < 0.05 interaction among CO2, species, and DBE) under eCO2 at the beginning of the experiment (104th day) showed higher RGR than their plants under aCO2 at beginning of the experiment (104th day).

a RGR = relative growth rate (RGR) and b RSR = root/stem ratio in five Cerrado tree species growing under ambient (430 ppm) and elevated (700 ppm) CO2 concentrations of CO [CO2] in open-top chambers. Symbols represent averages (n = 4), and bars indicate standard deviation. Capital letters compare differences within the same [CO2] and species in different DBE. Lowercase letters compare differences within the same [CO2] and DBE in different species. Asterisks indicate differences within the same species and DBE at different [CO2] (p < 0.05, Tukey’s test)
T. aurea plants at the middle of the experiment (189th day) under aCO2 showed a higher root/stem ratio (RSR) than their plants under aCO2 at beginning (104th day) and at the end of the experiment (276th day) Fig. 5b, F-test < 0.0001, p < 0.05 interaction among CO2, species, and DBE). At end of the experiment (276th day), T. aurea plants under eCO2 showed higher RSR than plants of the other species under eCO2 (Fig. 5b, F-test < 0.0001, p < 0.05 interaction among CO2, species, and DBE). Also, T. aurea plants under aCO2 at the middle of the experiment (189th day) showed a higher RSR than plants of the other species under aCO2 (Fig. 5b, F-test < 0.0001, p < 0.05 interaction among CO2, species, and DBE). At the middle of the experiment (189th day) T. aurea plants under aCO2 showed a higher RSR than their plants under eCO2 (Fig. 5b, F-test < 0.0001, p < 0.05 interaction among CO2, species, and DBE).
All significances and interactions between species and DBE, and correlation matrix of principal component analysis were presented in the supplementary material (Table S1, S2, and S3).
Discussion
Our results confirmed contrasting strategies for resource allocation and growth rate in woody species of distinct leaf persistence groups growing under eCO2. Under eCO2, the higher relative growth rate of the deciduous species (T. aurea and A. peregrina) in the first harvest, with a decrease in the next harvest, was probably due to the rapid accumulation of leaf mass at the beginning of exposure to eCO2, followed by a decline over time. There is an expectation that the increase in [CO2] will stimulate the initial growth of woody species. Poorter and Navas (2003) observed that fast-growing species responded better to the increase in [CO2] than slow-growing species. However, Lloyd and Farquhar (1996) presented an alternative hypothesis, suggesting that slow-growing species tend to consume a pronounced proportion of their daily CO2 assimilation in maintenance respiration. In our results we found more acquisitive strategies in deciduous trees, surpassing evergreen ones with more conservative strategies under eCO2 in the short term, supporting Poorter and Navas (2003). Thus, evergreens could be more responsive to eCO2 in the long-term because of the more conservative strategy of acquisition and resource use (Niinemets et al. 2011; Krishna and Garkoti 2022). However, deciduous cerrado species could show advantages over evergreens under eCO2 due to their fast growth and more acquisitive resource use strategy (Krishna and Garkoti 2022).
The faster leaf expansion found out of five species under eCO2 allowed the leaves to mature quickly. This ecological strategy of rapidly expanding the leaves developed under eCO2 maximizes carbon gain to the whole plant in less time, so the entire photosynthetic apparatus is ready and mature to carry out photosynthesis at high levels during the leaf life span (Reich et al. 1991; Edwards et al. 2014). Only the evergreen species S. polyphyllum and S. adstringesns showed a higher leaf expansion rate under eCO2 than under aCO2. This shift in the amount of leaf area produced by day could accelerate the leaf development and becoming these leaves mature faster over time. However, this faster leaf development could desynchronize leaf production to the increase in rain and temperature over the growing season.
Deciduous and evergreen Cerrado species studied in our experiment growing under eCO2 in 189 DBE showed an increase in stem and leaf dry mass, total leaf area, and non-structural carbohydrates than deciduous and evergreen species growing under aCO2 at 189 DBE. This result is contrary to the results found by Curtis and Wang (1998), Reich and Hobbie (2014), Reich et al. (2014), and Terrer et al. (2018), that at eCO2, growth in poor soil (as the soil used in our experiment) reduces biomass gain in woody species due to low soil nutrient availability. However, in a previous study, Souza et al. (2016) investigated the ecophysiological responses of three woody species from the Cerrado to the eCO2 and found increased biomass production. This response indicates the ability to uptake resources in this deciduous and evergreen species, even growing in poor soil and a relatively short time of exposure to greater CO2 availability. Furthermore, according to Reich et al. (2006), in the early developmental stages, even in poor soils, the high stimulus of biomass production may be more a characteristic of seedlings than of adult plants. In addition, Fox (1978), defined that a plant is efficient in the absorption and use of phosphorus when it produces a large amount of dry matter per unit of time and area, even when the environment (e.g., soil used in our study) has less phosphorus available for maximum productivity as found in our study.
The change in root/stem ratio only observed in the deciduous T. aurea, with a decrease in the biomass accumulation in roots at the expense of stems under eCO2, could impair the competitive capacity of this species for water and nutrients in the soil with other woody and herbaceous species (Melo et al. 2018). This change in biomass allocation in favor of stems seems more frequent in cerrado woody plants (Souza et al. 2016; Melo et al. 2018) compared gramineous plants. The higher biomass allocation favoring the shoots could benefit the species in light capture among dense gramineous strata in Cerrado areas (Melo et al. 2018). However, high efficiency of resource uptake from the soil is of great importance in cerrado areas because the seedlings and young woody plants need to capture water and nutrients in competition with efficient root systems of grass species (Oliveira et al. 2005). In the Cerrado, which is a domain marked by seasonal rainfall, it is commonly proposed that trees develop deep root systems that provide access to deeper, more humid layers of the soil (Rossato et al. 2013). Furthermore, Cerrado trees are capable of adjusting their hydraulic architecture and water characteristics (Bucci et al. 2005, 2008) to minimize seasonal imbalances. Therefore, the greater investment in growth of the aerial part than the belowground part in plants growing under eCO2 indicates a change in the biomass allocation pattern (Melo et al. 2018) which can make it difficult for savanna trees to resist the increase in seasonal droughts resulting from climate change (IPCC 2019).
All species studied showed similar responses in non-structural carbohydrates in roots. Although the roots did not increase the NSC under eCO2, it guaranteed the maintenance of the levels during the initial growth of all species compared to the lower values at 189 DBE in aCO2 plants. Even without an increase over time, maintaining a stock of non-structural carbohydrates in the roots could benefit these species after disturbances such as fire events or loss of aerial parts by herbivory. However, according to Manea and Leishman (2019), eCO2 could increase the woody species encroachment in grassland environments. According to Bond and Midgley (2000), the enhanced potential to resprout in woody plant species under eCO2 may also contribute to the invasion of these plants in pastures after events such as fire and herbivory. This potential change could favor the invasion of native (and alien) species over field formation, modifying the floristic and functional diversity of the Cerrado.
In the future, the establishment of woody seedlings in cerrado physiognomies would be strongly associated with CO2 plant responses and the subsequent competitive effect of grass species in these systems (Manea and Leishman 2015; Melo et al. 2018). Also, it is necessary to take into account that these species have a long life cycle, and therefore, the time required for exposure to eCO2 to produce significant effects on carbohydrate accumulation may be longer than the one observed in our work. However, it is important to investigate the interactions between increased [CO2], regrowth capacity, and vegetative growth in woody species from the Cerrado, as these factors may have an impact on the global carbon cycle and on responses to climate change (Bradley and Pregitzer 2007). Therefore, long-term studies are needed using, for example, open-air CO2 enrichment technologies (FACE), and experiments in open-top chambers (OTC) with trees cultivated with interaction of stresses and in small places with an element of competition or forests stands around natural sources of CO2 to increase the knowledge base on ecosystem responses to elevated atmospheric CO2. Also, understanding the regrowth capacity associated with carbon allocation and storage in woody plants growing under eCO2 is fundamental for the development of dynamic models for Cerrado vegetation in the face of climate change.
Data availability
The data set is being prepared for upload in the Plant Trait Database - TRY (https://www.try-db.org/TryWeb/Home.php).
References
Ainsworth EA, Long SP (2005) What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol 165:351–372. https://doi.org/10.1111/j.1469-8137.2004.01224.x
Ainsworth EA, Rogers A (2007) The response of photosynthesis and stomatal conductance to rising [CO2]: mechanisms and environmental interactions. Plant, Cell Environ 30:258–270
Bassirirad H et al (1996) Response of Eriophorum vaginatum to CO2 enrichment at different soil temperatures: effects on growth, root respiration and PO43− uptake kinetics. New Phytol 133:423–430. https://doi.org/10.1111/j.1469-8137.1996.tb01909.x
Batalha MA (2011) O cerrado não é um bioma. Biota Neotrop 11:21–24. https://doi.org/10.1590/S1676-06032011000100001
Bellard C, Bertelsmeier C, Leadley P, Thuiller W, Courchamp F (2012) Impacts of climate change on the future of biodiversity. Ecol Lett 15:365–377. https://doi.org/10.1111/j.1461-0248.2011.01736.x
Bond WJ, Midgley GF (2000) A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas. Glob Change Biol 6:865–869. https://doi.org/10.1046/j.1365-2486.2000.00365.x
Borges MP, Prado CHBA (2014) Relationships between leaf deciduousness and flowering traits of woody species in the Brazilian neotropical savanna. Flora-Morphol Distrib Funct Ecol Plant 209:73–80. https://doi.org/10.1016/j.flora.2013.10.004
Borges MP, Prado CHBA (2018) The significance of crown traits in cerrado vegetation. In: Price R (ed) Savannas: Exploration, Threats and Management Strategies. NOVA Science Publishers, NY, pp 1–29
Botelho SA, Ferreira RA, Malavasi MM, Davide AC (2000) Aspectos morfológicos de frutos, sementes, plântulas e mudas de jatobá-do-cerrado (Hymenaea stigonocarpa Mart. ex Hayne)-Fabaceae. Rev Bras Sementes 22:144–152
Bradley KL, Pregitzer KS (2007) Ecosystem assembly and terrestrial carbon balance under elevated CO2. Trends in Ecology & Evolution 22:538–547. https://doi.org/10.1016/j.tree.2007.08.005
Bucci SJ, Goldstein G, Meinzer FC, Franco AC, Campanello P, Scholz FG (2005) Mechanisms contributing to seasonal homeostasis of minimum leaf water potential and predawn disequilibrium between soil and plant water potential in Neotropical savanna trees. Trees 19:296–304. https://doi.org/10.1007/s00468-004-0391-2
Bucci SJ, Scholz FG, Goldstein G, Hoffmann WA, Meinzer FC, Franco AC, Giambelluca T, Miralles-Wilhelm F (2008) Controls on stand transpiration and soil water utilization along a tree density gradient in a Neotropical savanna. Agric for Meteorol 148:839–849. https://doi.org/10.1016/j.agrformet.2007.11.013
Cabral EL, Barbosa DDA, Simabukuro EA (2003) Armazenamento e germinação de sementes de Tabebuia aurea (Manso) Benth. & Hook. f. ex. S Moore Acta Botanica Brasilica 17:609–617. https://doi.org/10.1590/S0102-33062003000400013
Clarke PJ, Lawes MJ, Midgley JJ et al (2013) Resprouting as a key functional trait: how buds, protection and resources drive persistence after fire. New Phytol 197:19–35. https://doi.org/10.1111/nph.12001
Clarke PJ, Manea A, Leishman MR (2016) Are fire resprouters more carbon limited than non-resprouters? Effects of elevated CO2 on biomass, storage and allocation of woody species. Plant Ecol 217:763–771
Coutinho LM (2006) The biome concept. Acta Botanica Brasilica 20:13–23. https://doi.org/10.1590/S0102-33062006000100002
Cruz C, Lips SH, Martins-Louçzão MA (1997) Changes in the morphology of roots and leaves of carob seedlings induced by nitrogen source and atmospheric carbon dioxide. Ann Bot 80:817–823. https://doi.org/10.1006/anbo.1997.0524
Curtis PS, Wang X (1998) A meta-analysis of elevated CO2 effects on woody plant mass, form, and physiology. Oecologia 113:299–313. https://doi.org/10.1007/s00442005038
Damascos MA, Prado CHBA, Ronquim CC (2005) Bud composition, branching patterns and leaf phenology in cerrado woody species. Ann Bot 96:1075–1084. https://doi.org/10.1093/aob/mci258
Damascos M (2008) Conteúdo das gemas, momento da brotação e padrão de produção de folhas em espécies lenhosas do cerrado. Fisiologia Vegetal: práticas em relações hídricas, fotossíntese e nutrição mineral. –Barueri: editora Manole. https://www.researchgate.net/publication/316862707_Conteudo_das_gemas_momento_da_brotacao_e_padrao_de_producao_de_folhas_em_especies_lenhosas_do_cerrado. Acessed 01 June 2019.
De Lemos Filho JP, Guerra STM, Lovato MB, Scotti MRML (1997) Germinação de sementes de Senna macranthera, Senna multijuga e Stryphnodendron polyphyllum. Pesq Agrop Brasileira 32:357–361
DuBois M, Gilles KA, Hamilton JK, Rebers PT, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28:350–356. https://doi.org/10.1021/ac60111a017
Durigan G, Ratter JA (2016) The need for a consistent fire policy for Cerrado conservation. J Appl Ecol 53:11–15. https://doi.org/10.1111/1365-2664.12559
Edwards EJ, Chatelet DS, Sack L, Donoghue MJ (2014) Leaf life span and the leaf economic spectrum in the context of whole plant architecture. J Ecol 102:328–336. https://doi.org/10.1111/1365-2745.12209
Eeiten G (1972) The cerrado vegetation of Brazil. Bot Rev 38:201–341. https://doi.org/10.1007/BF02859158
Fajer ED, Bowers MD, Bazzaz FA (1991) Performance and allocation patterns of the perennial herb, Plantago lanceolata, in response to simulated herbivory and elevated CO2 environments. Oecologia 87:37–42. https://doi.org/10.1007/BF00323777
Felfili J, Silva Júnior MCD, Dias BJ, Rezende AV (1999) Estudo fenológico de Stryphnodendron adstringens (Mart.) Coville no cerrado sensu stricto da fazenda Água Limpa no Distrito Federal. Brasil Brazil J Bot 22:83–90. https://doi.org/10.1590/S0100-84041999000100011
Fox RH (1978) Selection for phosphorus efficiency in corn. Commun Soil Sci Plant Anal 9:13–37
Franco AC, Rossatto DR, Silva LDCR, da Silva FC (2014) Cerrado vegetation and global change: the role of functional types, resource availability and disturbance in regulating plant community responses to rising CO2 levels and climate warming. Theor Exp Plant Physiol 26:19–38. https://doi.org/10.1007/s40626-014-0002-6
Haridasan M (2000) Nutrição mineral de plantas nativas do cerrado. Rev Bras Fisiol Veg 12:54–64
Hoffmann WA, Bazzaz FA, Chatterton NJ, Harrison PA, Jackson RB (2000) Elevated CO2 enhances resprouting of a tropical savanna tree. Oecologia 123:312–317. https://doi.org/10.1007/s004420051017
Hughes CE (2017) Are there many different routes to becoming a global biodiversity hotspot? Proc Natl Acad Sci 11(4275–4277):2017. https://doi.org/10.1073/pnas.1703798114
Ipcc (2019) Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems In: P.R. Shukla, J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H.-O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi, J. Malley, (eds). (In press).
Kgope BS, Bond WJ, Midgley GF (2010) Growth responses of African savanna trees implicate atmospheric [CO2] as a driver of past and current changes in savanna tree cover. Austral Ecol 35:451–463. https://doi.org/10.1111/j.1442-9993.2009.02046.x
Klink CA, Machado RB (2005) A Conservação Do Cerrado Brasileiro. Megadiversidade 1:147–155
Körner C, Miglietta F (1994) Long term effects of naturally elevated CO2 on Mediterranean grassland and forest trees. Oecologia 99:343–351. https://doi.org/10.1007/BF00627748
Krishna M, Garkoti SC (2022) Evergreen and deciduous tree species show distinct strategies to synchronize with seasonality in mid-elevational forests of central Himalaya. For Ecol Manage 526:120567. https://doi.org/10.1016/j.foreco.2022.120567
Lapola DM, Martinelli LA, Peres CA, Ometto JP, Ferreira ME, Nobre CA, Joly CA (2014) Pervasive transition of the Brazilian land-use system. Nat Clim Change 4:27–35. https://doi.org/10.1016/j.foreco.2022.120567
Li Y, Gao Y, Xu X, Shen Q, Guo S (2009) Light-saturated photosynthetic rate in high-nitrogen rice (Oryza sativa L.) leaves is related to chloroplastic CO2 concentration. J Exp Bot 60:2351–2360. https://doi.org/10.1093/jxb/erp127
Lloyd J, Farquahar GD (1996) The CO2 dependence of photosynthesis, plant growth responses to elevated atmospheric CO2 concentrations and their interaction with soil nutrient status I. General principles and forest ecosystems. Funct Ecol. https://doi.org/10.2307/2390258
Lorenzi H (1992) Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Plantarum, Nova Odessa
Lorenzi H (2000) Árvores Brasileiras. Plantarum, Nova Odessa
Lorenzi H (2002) Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Plantarum, Nova Odessa
Maia RA, Fernandes GW, Silva AI, Souza JP (2019) Improvement in light utilization and shoot growth in Hymenaea stigonocarpa under high CO2 concentration attenuates simulated leaf herbivory effects. Acta Botanica Brasilica 33:558–571. https://doi.org/10.1590/0102-33062019abb0149
Manea A, Leishman MR (2015) Competitive interactions between established grasses and woody plant seedlings under elevated CO2 levels are mediated by soil water availability. Oecologia 177:499–506. https://doi.org/10.1007/s00442-014-3143-z
Manea A, Leishman MR (2019) The resprouting response of co-occurring temperate woody plant and grass species to elevated [CO2]: An insight into woody plant encroachment of grasslands. Austral Ecol 44:917–926. https://doi.org/10.1111/aec.12760
Melo NM, Rayete SER, Pereira EG, Souza JP (2018) Rising [CO2] changes competition relationships between native woody and alien herbaceous Cerrado species. Funct Plant Biol 45:854–864. https://doi.org/10.1071/FP17333
Miranda CDC, Souza DMDS, Manhone PR, Oliveira PCD, Breier TB (2012) Germinação de sementes de Anadenanthera peregrina (L.) Speg. com diferentes substratos em condições laboratoriais. Floresta e Ambiente 19:26–31. https://doi.org/10.4322/floram.2012.004
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GA, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. https://doi.org/10.1038/35002501
Newingham BA, Vanier CH, Charlet TN, Ogle K, Smith SD, Nowak RS (2013) No cumulative effect of 10 years of elevated [CO2] on perennial plant biomass components in the Mojave Desert. Glob Change Biol 19:2168–2181. https://doi.org/10.1111/gcb.12177
Niinemets Ü, Flexas J, Peñuelas J (2011) Evergreens favored by higher responsiveness to increased CO2. Trends Ecol Evol 26:136–142. https://doi.org/10.1016/j.tree.2010.12.012
Nowak RS, Ellsworth DS, Smith SD (2004) Functional responses of plants to elevated atmospheric CO2–do photosynthetic and productivity data from FACE experiments support early predictions? New Phytol 162:253–280
Nunez S, Arets E, Alkemade R, Verwer C, Leemans R (2019) Assessing the impacts of climate change on biodiversity: is below 2° C enough? Climatic Change 154:351–365. https://doi.org/10.1007/s10584-019-02420-x
Oliveira RS, Bezerra L, Davidson EA, Pinto F, Klink CA, Nepstad DC, Moreira A (2005) Deep root function in soil water dynamics in cerrado savannas of central Brazil. Funct Ecol 19:574–581. https://doi.org/10.1111/j.1365-2435.2005.01003.x
Oliveira ACG, Rios PM, Pereira EG, Souza JP (2021) Growth and competition between a native leguminous forb and an alien grass from the Cerrado under elevated CO2. Austral Ecol 46:750–761. https://doi.org/10.1111/aec.13006
Ouédraogo DY, Mortier F, Gourlet-Fleury S, Freycon V, Picard N (2013) Slow-growing species cope best with drought: evidence from long-term measurements in a tropical semi-deciduous moist forest of C entral A frica. J Ecol 101:1459–1470. https://doi.org/10.1111/1365-2745.12165
Overbeck GE et al (2015) (2015) Conservation in Brazil needs to include non-forest ecosystems. Divers Distrib 21:1455–1460. https://doi.org/10.1111/ddi.12380
Pearson M, Brooks GL (1996) The effect of elevated CO and grazing by Gastrophysa viridula on the physiology and regrowth of Rumex obtusifolius. New phytologist 133:605–616. https://doi.org/10.1111/j.1469-8137.1996.tb01929.x
Pecl GT, Araújo MB, Bell JD, Blanchard J, Bonebrake TC, Chen IC, Williams SE (2017) Biodiversity redistribution under climate change: impacts on ecosystems and human well-being. Science 355:6332. https://doi.org/10.1126/science.aai9214
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte MS, Cornwell WK, Craine JM, Gurvich DE, Urcelay C, Veneklaas EJ, Reich PB, Poorter L, Wright IJ, Ray P, Enrico L, Pausas JG, de Vos AC, Buchmann N, Funes G, Quétier F, Hodgson JG, Thompson K, Morgan HD, ter Steeger H, van der Heijden A, Sack L, Blonder B, Poschlod P, Vaieretti MV, Conti G, Staver AC, Aquino S, Cornelissen JHC (2013) New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot 61:167–234
Peterson ML, Doak DF, Morris WF (2019) Incorporating local adaptation into forecasts of species’ distribution and abundance under climate change. Glob Change Biol 25:775–793. https://doi.org/10.1111/gcb.14562
Poorter H, Navas ML (2003) Plant growth and competition at elevated CO2: on winners, losers and functional groups. New Phytol 157:175–198. https://doi.org/10.1046/j.1469-8137.2003.00680.x
R Core Team (2019) ‘A language and environment for statistical computing’. (Vienna, Austria: R Foundation for Statistical Computing) Available at: http://www.R-project.org/ Accessed 01 Jan 2019.
Read JJ, Morgan JA (1996) Growth and partitioning inpascopyrum smithii (C3) andBouteloua gracilis (C4) as influenced by carbon dioxide and temperature. Ann Bot 77:487–496. https://doi.org/10.1006/anbo.1996.0059
Reich PB, Uhl C, Walters MB, Ellsworth DS (1991) Leaf lifespan as a determinant of leaf structure and function among 23 Amazonian tree species. Oecologia 86:16–24. https://doi.org/10.1007/BF00317383
Reich PB, Hobbie SE, Lee T, Ellsworth DS, West JB, Tilman D, Trost J (2006) Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 440:922–925. https://doi.org/10.1038/nature04486
Reich PB, Hobbie SE, Lee TD (2014) Plant growth enhancement by elevated CO2 eliminated by joint water and nitrogen limitation. Nat Geosci 7:920–924. https://doi.org/10.1038/ngeo2284
Rossatto DR, Silveira Lobo Sternberg L, Franco AC (2013) The partitioning of water uptake between growth forms in a Neotropical savanna: do herbs exploit a third water source niche? Plant Biol 15:84–92. https://doi.org/10.1111/j.1438-8677.2012.00618.x
Santos BRVD, Benedito CP, Torres SB, Leal CCP, Alves TRC (2019) Physiological maturity of Tabebuia aurea (Silva Manso) Benth. & Hook. f. ex S. Moore seeds. Journal of Seed Science 41:498–505. https://doi.org/10.1590/2317-1545v42n4222528
Silva OB, Goelzer A, de Oliveira Carnevali T, dos Santos FHM, Zárate NAH, Carmo Vieira M (2020) Produção e qualidade de mudas de Anadenanthera peregrina var. falcata (Benth.) Altschul sob substratos e bioestimulador. Revista Principia-Divulgação Científica e Tecnológica Do IFPB 52:169–178
Silva JBL. Respostas das culturas de feijão submetido ao estresse hídrico e de milho em atmosfera enriquecida com CO2 2010. PhD Thesis, Federal University of Viçosa, Minas Gerais.
Siqueira MF, Peterson AT (2003) Consequences of global climate change for geographic distributions of cerrado tree species. Biota Neotrop 3:1–14. https://doi.org/10.1590/S1676-06032003000200005
Souza JP, Prado CH, Damascos MA, Albino ALS (2009a) Influence of shoot inclination on irradiance and morphophysiological leaf traits along shoots in cerrado trees with distinct leaf deciduousness. Braz J Plant Physiol 21:281–289. https://doi.org/10.1590/S1677-04202009000400004
Souza JP, Prado CH, Albino ALS, Damascos MA (2009b) Shoot-foliage relationships in deciduous, semideciduous, and evergreen cerrado tree species. Braz J Plant Physiol 21:76–86. https://doi.org/10.1590/S1677-04202009000100009
Souza JP, Prado CHB, Albino ALS, Damascos MA, Souza GM (2011) Network analysis of tree crowns distinguishes functional groups of Cerrado species. Plant Ecol 212:11–19. https://doi.org/10.1007/s11258-010-9797-7
Souza JP, Melo NMJ, Pereira EG, Halfeld AD, Gomes IN, Prado CHB (2016) Responses of woody Cerrado species to rising atmospheric CO2 concentration and water stress: gains and losses. Funct Plant Biol 43:1183–1193. https://doi.org/10.1071/FP16138
Souza BC et al (2020) Drought response strategies of deciduous and evergreen woody species in a seasonally dry neotropical forest. Oecologia 194:221–236. https://doi.org/10.1007/s00442-020-04760-3
Terrer C, Vicca S, Stocker BD, Hungate BA, Phillips RP, Reich PB, Prentice IC (2018) Ecosystem responses to elevated CO2 governed by plant–soil interactions and the cost of nitrogen acquisition. New Phytol 217:507–522. https://doi.org/10.1111/nph.14872
Van der Putten WH, Macel M, Visser ME (2010) Predicting species distribution and abundance responses to climate change: why it is essential to include biotic interactions across trophic levels. Philos Trans Royal Soc B: Biol Sci 365:2025–2034. https://doi.org/10.1098/rstb.2010.0037
Velazco SJE, Villalobos F, Galvão F, De Marco JP (2019) A dark scenario for Cerrado plant species: Effects of future climate, land use and protected areas ineffectiveness. Divers Distrib 25:660–673. https://doi.org/10.1111/ddi.12886
Wilsey BJ, Mcnaughton SJ, Coleman JS (1994) Will increases in atmospheric CO2 affect regrowth following grazing in C4 grasses from tropical grasslands? A test with Sporobolus kentrophyllus. Oecologia 99:141–144
Wilsey BJ, Coleman JS, Mcnaughton SJ (1997) Effects of elevated CO2 and defoliation on grasses: a comparative ecosystem approach. Ecol Appl 7:844–853. https://doi.org/10.1890/1051-0761(1997)007[0844:EOECAD]2.0.CO;2
Acknowledgements
The authors thank the CNPq for the Ph.D. studentship awarded to the first author and the FAPEMIG for the financial support (FORTIS-TCT-10254/2014 and CRA-APQ-02935-16). The authors thank Dr. Carlos Henrique Britto de Assis Prado at the Federal University of São Carlos for his support in carrying out the work. EGP received a research productivity grant from the National Council for Scientific and Technological Development (CNPq, Grant No. 312427/2020-4)
Funding
Conselho Nacional de Desenvolvimento Científico e Tecnológico,141003/2016-2, 312427/2020-4, Fundação de Amparo à Pesquisa do Estado de Minas Gerais,FORTIS-TCT-10254/2014.
Author information
Authors and Affiliations
Contributions
Melo, Nayara Magry Jesus Conceptualization Data curation Formal analysis Investigation Methodology Writing - original draft Writing review & editing Gonçalves, Marina Efigenia Methodology Maia, Renata Aparecida Methodology Data curation Pereira, Eduardo Gusmão Methodology Writing review & editing Souza, João Paulo Conceptualization Data curation Formal analysis Investigation Methodology Project administration Supervision Writing - original draft Writing review & editing
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Additional information
Communicated by Lauchlan Fraser.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Melo, N.M.J., Maia, R.A., Gonçalves, M.E. et al. Deciduous trees could benefit more from the rise in [CO2] than evergreens in the Brazilian Cerrado. Plant Ecol 225, 479–490 (2024). https://doi.org/10.1007/s11258-024-01398-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-024-01398-6