
Abstract
Urbanisation drives overall declines in insect pollinators. Although urban green spaces can provide suitable habitat for pollinators much remains to be learned about how urban landscapes either promote or negatively impact pollinators. We investigated how backyard design, local (100 m) and landscape (500 m) scale vegetation cover and human population density were associated with non-eusocial native bee species, eusocial bees (Apis mellifera and Tetragonula spp.), and hoverflies, in residential green spaces of the subtropical city Brisbane, Australia. We found that associations between bee abundance and vegetation cover were moderated by human density, but the direction of this effect differed for non-eusocial and eusocial species. Non-eusocial bee abundance was positively associated with tree cover at local and landscape scales when human densities were low, but negatively so at high human population densities. We suggest this may be because the quality of vegetation for non-eusocial bees deteriorates as human density increases. In contrast, abundance of eusocial bees was negatively associated with increasing local cover of grass and shrubs at low levels of human density, but positively associated at high densities. This affinity to humans could partly be explained by domesticated “kept” hives. We found no effect of urban gradients on bee species richness. Hoverfly abundance was negatively related to human density and positively related to vegetation cover at local and landscape scales. At the backyard scale, both bee species richness and bee and hoverfly abundances were positively associated to flower abundance. Backyards with more vegetation cover had higher densities of non-eusocial bees. Our results thus support the idea that urban greening in densely populated areas at multiple spatial scales can benefit a range of insect pollinators.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Documented global declines of insect pollinators (e.g. Zattara and Aizen 2021) have led to concerns about the consequences for ecosystem functioning and pollination services to wild and crop plant species (Potts et al. 2016). The main drivers behind global pollinator losses are thought to include agricultural intensification, the loss of natural and semi-natural habitats that reduces flower and nesting resources, and an increased use of pesticides with both direct and indirect negative effects on pollinators (Pott et al. 2010, Goulson et al. 2015). While urbanisation is a recognised driver of biodiversity loss (Foley et al. 2005) some urban elements, such as parks, allotments, and ruderal habitats (brown fields), are known to benefit insect pollinators in some landscapes (Baldock et al. 2019, Twerd and Banaszak-Cibicka 2019). Urban landscapes may even act as habitat refuges for some species (Hall et al. 2017). This is especially true at intermediate (suburban) levels of urbanisation (Wenzel et al. 2019), and when urban areas are surrounded by intensively managed agricultural landscapes (Wenzel et al. 2019; Persson et al. 2020). However, the effects of urbanisation on insect pollinators vary depending on region (De Palma et al. 2016), taxon (Theodorou et al. 2020) and functional traits (Wilson and Jamieson 2019). Most studies of urban pollinators to date are from temperate regions in the northern hemisphere and focus on bees (De Palma et al. 2016; Wenzel et al. 2019; Prendergast et al. 2022a). Studies from other biomes, and those including both bee and non-bee pollinator genera, are therefore needed to improve our understanding of how pollinators respond to urbanisation more generally.
Actions to benefit urban pollinator communities will likely require interventions at multiple spatial scales and include consideration of local green space design and management (Aronson et al. 2017; Wenzel et al. 2019; Baldock 2020). For example, bee abundance and diversity in human modified landscapes have been shown to increase with both local (Winfree et al. 2011; Burdine and McCluney 2019), and landscape scale availability of flower resources in agricultural (Persson and Smith 2013; Palma et al. 2015) and urban (Fortel et al. 2014; Threlfall et al. 2015; Buchholz et al. 2020) ecosystems. Yet, in some cases human population density is a stronger predictor of pollinator diversity than direct measures of vegetation cover (Kuussaari et al. 2020; Persson et al. 2020). A possible reason is that vegetation quality for pollinators deteriorate as human density increases (McKinney 2008; Aronson et al. 2017; Banaszak-Cibicka and Żmihorski 2020). This can occur for several reasons. For example, human use and disturbance increase as more people share spatially-reduced green infrastructure (Haaland and van den Bosch 2015), and green space design and management is adapted to withstand increased disturbance and reduce costs of management. In addition, aesthetic ideals negatively affecting habitat quality for pollinators and other biodiversity are pervasive (Hanson et al. 2021), such as promoting extensive lawn cover (Ignatieva and Hedblom 2018), removing wild and dead plant-material and weeds (Smith et al. 2006a; Matteson and Langellotto 2010), using mulch as ground cover (Quistberg et al. 2016), and introducing a large proportion of non-native ornamental plants (Smith et al. 2006b; Baldock et al. 2019), including trees (Kowarik 2008). High habitat quality has indeed been shown to be more important than large habitat areas to support bee community functional diversity (Buchholz et al. 2020).
Ecological and life-history traits of pollinator species will interact with the environment to determine if, and how, urban resources can benefit particular taxa, including exotic ones (Wilson and Jamieson 2019). For example, among bees sociality may be beneficial in heterogeneous highly urbanised landscapes with spatially scattered and diverse floral resources (reviewed by Wenzel et al. 2019, but see Wilson and Jamieson 2019), as is a generalist (polylectic) diet (Wenzel et al. 2019; Buchholz et al. 2020). However, specialist (oligolectic) species may still find habitat in hot spots such as urban gardens and allotments (Martins et al. 2017; Baldock et al. 2019). Small-bodied species have been shown to benefit from urbanisation (Banaszak-Cibicka and Żmihorski 2012; Hinners et al. 2012, Threlfall et al. 2015), and while bee species nesting underground are often common in urbanised areas (Buchholz et al. 2020), they may also be more sensitive to high levels of impervious surfaces compared to cavity nesters (Fortel et al. 2014; Wenzel et al. 2019; Prendergast et al. 2022a). Thus, while we generally expect lower abundances of bees in more highly urbanised areas (e.g., Fortel et al. 2014; Geslin et al. 2016), species turnover along urbanisation gradients (Banaszak-Cibicka and Zmihorski 2012), as well as high α-diversity of urban sites may, at least partly, mask general negative effects of urbanisation on bee species richness and diversity (Banaszak-Cibicka and Zmihorski 2020, Persson et al. 2020). While hoverflies generally respond negatively to urbanisation (e.g., Verboven et al. 2014; Persson et al. 2020), local habitat quality, such as high flower cover and vegetation height in natural or semi-natural habitats, can benefit this group (Dylewski et al. 2019). It is thus important to tease apart how both local and landscape scale components of urbanisation shape pollinator communities, and why some taxa may respond differently.
Cities are largely unique with regard to combinations of, biogeographical, historical and socio-economic variables (Alberti et al. 2003), affecting both local and landscapes scale resources for biodiversity (McKinney 2008; Hahs et al. 2009a, b). Hence, using a simple urban to rural gradient to analyse how parameters of biodiversity respond to urbanisation could lead to an inability to reveal why any detected patterns occur (Alberti et al. 2003). There is thus a value in including variables relating to urban form (Persson et al. 2020) and green space (backyard) design and quality (Aronson et al. 2017) in analyses of urban biodiversity. Residential areas and private green spaces are an important part of urban ecosystems, often covering more than 30% of urban areas (Loram et al. 2007; Goddard et al. 2010). Residents invest ample resources to provide ecosystem services from their backyards, such as space for recreation (Barnes et al. 2020) and gardening of crops that require insect pollination for fruit set (Lin and Egerer 2017). The societal value of residential green spaces suggests great potential in introducing design and management practices that are positive for biodiversity (Goddard et al. 2013). To plan and manage urban areas to promote insect pollinators and pollination services, it is thus necessary to evaluate how pollinators respond to both local landscape design and greening of backyards, and landscape scale interventions such as increasing vegetation cover or building denser residential areas at the neighbourhood scale.
With the aim to inform future actions to improve urban areas for insect pollinators, we investigate how different types of urban residential landscapes affect insect pollinators present in private backyards in a subtropical, southern hemisphere, city (Brisbane, Australia). We study three spatial scales relevant to pollinator foraging and movement, and urban planning: the local backyard (< 50 m), 100 and 500 m radii. We (i) investigate the characteristics of a pollinator friendly backyard, block, and neighbourhood, and (ii) determine whether these characteristics differ between pollinator taxa with different life-histories. To characterise local backyards, we perform surveys of backyard features (vegetation, soil, flower cover, etcetera) and use cluster analysis to obtain types of backyard designs for inclusion in further analyses.
We study bees (Apoidea: Anthophila) and hoverflies (Syrphidae). These taxa are important pollinators, visiting a large portion of native plants and crops globally (Ollerton et al. 2011; Rader et al. 2016), but have contrasting ecological and life-history traits. Most wild bee species are non-eusocial, that is, they are solitary, communal or semi-social. Each female forages for her own offspring (solitary and communal) or shares the task with cohabiting sisters (semi-social). Eusocial bees, in contrast, have a single queen, which heads an annual (e.g., bumblebees) or perennial (e.g., honeybees) colony of up to several thousand workers. In Brisbane, two groups of eusocial bees exist: Two species of native Tetragonula spp. (Meliponini—stingless bees) and the exotic Apis mellifera (Apini—honeybees). Both groups occur as managed and wild colonies throughout the study region. The non-eusocial bee species in the region are either polylectic or oligolectic, where the former are generalists that forage from a variety of plant families, while the latter have a specialised diet and typically forage on a few or a single plant family. In contrast, eusocial bee species in this region are highly polylectic and forage from both native and exotic plants (Threlfall et al. 2015; Makinson et al. 2017). With their abundant flowering, eucalypt tree species (Myrtaceae) provide important forage resources for both generalist and specialist bees in the study region (Michener 1965; Threlfall et al. 2015). Bees are central-place foragers and commute to and from a fixed nest when foraging for their offspring. Foraging ranges of bees are typically a few hundred meters for smaller solitary species (Greenleaf et al. 2007) and around 330–800 m for stingless bees (Smith et al. 2017; Forbes Saurels et al. unpublished), while the European honeybee can travel several kilometres (Hagler et al. 2011).
While adult hoverflies feed on pollen and nectar, they lay their eggs on vegetation, dead organic matter or in ephemeral aquatic environments where larvae feed on e.g., aphids, dead wood, or dung (Rotheray and Gilbert 2008). Although hoverflies are not restricted to a fixed nest, they require complementary resources in the form of local flower resources for adults and across landscapes, larval habitats, e.g., within 500—750 m (Meyer et al. 2009; Moquet et al. 2018). Compared to bees, they can thus be expected to be similarly restricted in movement, but more sensitive to urbanisation (Bates et al. 2011; Verboven et al. 2014; Persson et al. 2020), as typical larval habitats are often lacking in urban landscapes due to green space design and management practices (Aronson et al. 2017).
We surveyed bee and hoverfly potential pollinators (referred to as pollinators from here on) in residential backyards along selected gradients of human population density and vegetation cover. We investigated to what extent pollinator abundance and species richness were related to local backyard flower abundance, type of backyard design, and the surrounding vegetation cover and human population density at spatial scales (100 and 500 m radii) relevant to pollinator movement and urban spatial planning. We expect that pollinator abundance and richness will be positively associated with local flower abundance and local and landscape scale vegetation cover, and negatively to local and landscape scale human population density. We also expect that the type of backyard design will affect bees and hoverflies differently, based on their different nesting/larval requirements. For example, yards with scant vegetation cover will support fewer pollinators overall, and yards with few trees and shrubs may especially support fewer native non-eusocial bees as these largely lack both local foraging and nesting resources.
Methods
Study design and site selection
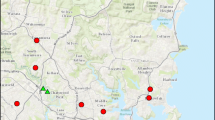
This study was conducted during November 2017 in Brisbane, Australia, in an area of approximately 20 × 30 km of suburban residential neighbourhoods varying in human population density and extent of green spaces (Fig. 1). We identified potential survey sites (private backyards) using social media and the staff newsletter at the University of Queensland. We received approximately 200 applications from Brisbane residents. From these, we selected 45 backyards (of 40 detached houses and 5 multi-family complexes) along gradients of vegetation cover and human population density per sealed surface, as detailed below.

Map of the study area in Brisbane (Queensland), Australia. Black dots mark surveyed backyards. Background map: ©Open street map
We obtained data on human population density and land cover from Brisbane City Council: the 2011 population census mesh blocks and 2014 land cover map. The latter had been processed into a raster of 100 × 100 m grid cell size, containing sealed surfaces, and three vegetation classes based on LiDAR data (Caynes et al. 2016): tree canopy cover (≥ 2 m height), shrub cover (< 2 m), and grass cover. We combined all vegetation classes using R version 3.2.5 (R Core Team 2015) and the Raster package (Hijmans 2016). The polygon shape layer on human population density was rasterized in R into a 100 × 100 m grid raster, aligned with the vegetation raster. The census mesh block polygons vary in size and shape. We therefore divided the human population within a polygon between all 100 × 100 m raster cells that it intersected with, in proportion to the fraction of the polygon area each cell made up. We calculated human population density per sealed (grey) surface area in each grid-cell (Hahs and McDonnell 2006). This measure is more related to urban form and how intensively the built land is used than human population density per se.
To select focal backyards along urban gradients, we used these raster layers to perform moving window analyses to calculate mean vegetation cover and population density in each 1 × 1 km window (10 × 10 pixels) over an area including all candidate backyards. The output was visually inspected and data points (focal pixels) outside the 10th and 90th percentiles on each axis (population and vegetation) were removed to avoid selecting backyards in areas where the two gradients were highly correlated (Pasher et al. 2013). This allowed us to include both vegetation and human density gradients in the same statistical models. We divided the remaining data points into four categories: 1) low vegetation cover and low human density, 2) low vegetation and high human density, 3) high vegetation and low human density, and 4) high vegetation and high human density. The categories were exported as tiff-files and used to draw maps in ArcGIS (Esri). Based on these maps and the position of the candidate backyards, we selected 45 sites to cover a wide variation in vegetation cover and human population density (Fig. 2). Selected backyards were on average situated 1584 ± 1035 m (mean ± std) from the closest other backyard in the study.

Distribution of surveyed backyards along the two urban gradients (500 m radius) used for site selection: total vegetation cover and human population density
To obtain the urban gradients for analyses of pollinator response variables, we used the extract function in the Raster package (Hijmans 2016) to extract vegetation cover, cover of sealed surface, and human population numbers within 100 m and 500 m radii of each selected backyard centre coordinate. Vegetation data was categorised as: tree canopy cover, and grass and shrub cover combined, because of the potentially contrasting effects these categories may have on pollinators (Hall et al. 2019).
Pollinator and backyard surveys
Each backyard was surveyed for pollinators three times in early summer, 3rd to 25th November 2017, between 08:00 – 13.00 on days with favourable weather for flying insects (wind < 5 m/s, temperature between 22 and 34 o C). To avoid bias, the three surveys per backyard were rotated to be either early, mid, or late in the day, within the set sampling window (8:00 – 13:00). Each survey involved slowly walking along two 50 m long 1 m wide transects for a total of 10 min, while collecting pollinators with a standard insect aerial sweep net. In cases where we were unable to capture specimens, we instead noted them to the closest identifiable genus or morpho-group. Eleven small backyards (< 100 m2) only had space for a single 50 m transect, which was surveyed for 5 min. This difference in sampling effort was accounted for in analyses, as detailed below. To index the attractiveness to pollinators of individual transects, we estimated the number of open insect-pollinated flower units in each transect. A flower unit is an easy countable entity that requires a pollinator to fly between them (Szigeti et al. 2016). The flower unit counted varied by taxa, e.g., individual flowers for Rosaceae, inflorescences for Asteraceae, racemes for Fabaceae, umbels for Apiaceae and flowering stalks for small-flowered Lamiaceae.
We mapped backyard features to characterise backyards into design types. We overlaid a 2 × 2 m grid onto aerial photos of backyards obtained from NearMap (freely available to researchers at University of Queensland, 2017). During field visits, we noted the proportion cover of the following features in each grid cell: short cut lawn, tall grass, shrubs, tree-trunks, tree crown-cover, leaf-litter, open soil, vegetable and berry patches, flowerbeds, gravel or stones, and sealed surfaces (e.g., wooden deck, pool, concrete paths) and calculated the proportion of each per backyard. All features, except for tree crown-cover were mutually exclusive and thus summed to one. In addition, for each backyard we estimated its size (m2), total number of open flower units, and counted the number of eucalypt trees as a proxy for the number of native trees. We counted the number of flowering trees within a 50 m radius of backyards as a measure of locally available large flower resources. The most common trees in flower were several species of Myrtaceae, mainly Eucalyptus spp., and the exotic Jacaranda mimosifolia.
Analysis of backyard features
All statistical analyses were performed in R version 3.6.1. To evaluate potential effects of contrasting gardening styles on pollinators we used hierarchical cluster analysis on the mapped garden features, to obtain groups of backyard types. We included: lawn cover, tree crown-cover, shrub cover, backyard size, and flower density. Leaf-litter, tall grass, open soil, vegetable and berry patches, and flowerbeds were combined into “other features”, as each of these were missing or constituted only a small fraction of the majority of backyards. We first calculated a dissimilarity matrix using function daisy() in package Cluster (Maechler et al. 2020), then used hierarchical clustering with function hclust() to obtain groups. We selected three “backyard design type” groups based on visual inspection of the dendrogram and barplot, and interpretability of the resulting groups. The latter was checked by modelling how groups related to each of the backyard features included in the clustering using linear models, assigning each garden feature as the response variable and backyard design type as the fixed effect. We visually checked barplots of model residuals for equal variance among groups. The resulting three (3) groups were later included in pollinator models.
The backyard design types differed significantly in flower density (F2,42 = 6.35; P = 0.0039), tree canopy cover (F2,42 = 6.378; P = 0.0038), proportion lawn (F2,42 = 50.46; P < 0.0001), shrubs (F2,42 = 13.75; P < 0.0001), and “other features” (F2,42 = 14.15; P < 0.0001), with a non-significant tendency for backyard size to differ between types (F2,42 = 2.86; P = 0.069). Type 1 was characterised by lower cover of flowers, lawn and shrubs, and higher cover of “other features” (e.g., open soil, flowerbeds, vegetable patches, leaf litter and tall grass) than types 2 and 3. Type 2 was characterised by mixed vegetation with high cover of both flowers, lawn, trees, and shrubs. Type 3 was characterised by very high cover of lawn, but low cover of flowers, trees, shrubs, and “other features”, with a non-significant tendency to be of larger size than the other types.
We tested whether local backyard features differed along the urban gradients. First, we tested if backyard design types were related to the three gradients at 100 m and 500 m scales, using a non-parametric Kruskal–Wallis test, with design type as the response and gradient as the explanatory variable. Second, we used GLMs to check if the proxies for local native vegetation (number of eucalypt trees in backyards) and large flower resources (number of flowering trees within 50 m), were related to the urban gradients. Because models were over-dispersed, we used a negative binomial distribution with glm.nb from package MASS (Venables and Ripley 2002). We assigned the number of eucalypts, or flowering trees, as the response variable, and one of the urban gradients (low vegetation, tree cover and human population density at 100 or 500 m) as an explanatory variable in each model.
Pollinator data
Collected insect samples were determined to species or morpho-species level by experts at the University of Queensland, the Australian Museum (Sydney) and Queensland Museum (Brisbane). Pollinator observations for the two transects and three survey rounds were aggregated into one observation per backyard. We calculated the mean flower abundance along the surveyed transects per backyard over the three surveys. We calculated the abundance of all observed and collected eusocial bee species (A. mellifera and Tetragonula spp.), non-eusocial bee species, and hoverflies. There were two reasons for analysing eusocial and non-eusocial species separately. First, because the former occurs both as feral wild colonies and managed hived colonies in the study region, their abundance is partly directly governed by humans. Second, because eusocial bees can communicate within the colony to recruit foragers to distant but rewarding foraging patches (Beekman and Ratnieks 2000), they may respond differently to urbanisation (Wenzel et al. 2019).
Estimates of bee species richness were based on all collected specimens and those easily identified to genus in the field without collection, i.e., A. mellifera, Tetragonula spp., Amegilla spp., Thyreus nitidulus, and Xylocopa (Koptortosoma) sp. The 26 specimens observed but not identified were left out of this estimate. Because sampling effort differed between backyards, with either one (50 m) or two (100 m) transects, we predicted the effective number of bee species (Hill number = 0) using the iNEXT() function (Hsieh et al. 2016). We combined eusocial and non-eusocial species to improve sample size, and thus robustness of the estimate. We did not analyse species richness for hoverflies, because only 82 specimens were caught and identified to species, mostly of a single species (Simosyrphus grandicornis, 57 of 82 specimens) (Online resource: Tables 3 and 4). Bee species were categorised in terms of their flower relationship, nesting habitat, body, and tongue length, according to Michener (2007), the PaDIL database, and measurements in the lab (Online resource: Table 4).
We modelled the effects of local and landscape predictors on pollinator response variables using GLMs. Full models were pollinator response variable = garden design type + (log)transect flower abundance + proportion tree cover + proportion grass and shrubs + human population density + (tree cover × human density) + (grass and shrubs × human density). In models of pollinator abundance, we included (log) transect survey area (50 or 100 m2) as an offset to account for unequal sampling efforts. The GLMs of pollinator abundance were over-dispersed, and we therefore used a negative binomial distribution with glm.nb from package MASS (Venables and Ripley 2002). Bee species richness (logged) was modelled assuming a Gaussian distribution as iNEXT predicted species richness are not integers. We ran one model for each combination of pollinator response variables (bee species richness, eusocial bee abundance, non-eusocial bee abundance, hoverfly abundance) and spatial scale (100 m and 500 m), i.e., eight models in total. Non-significant interactions were removed (backward selection), and models re-run to obtain final results for component factors.
All fixed factors were scaled prior to statistical analyses to facilitate interpretation of effect estimates, using function scale to centre values and divide them by their standard deviations. We calculated Variance Inflation Factors (VIF) to assess potential collinearity between fixed factors. We assessed over-dispersion and outliers using the package Dharma, function testResiduals() (Hartig 2020) and residual plots. We checked for spatial autocorrelation among residuals using Moran’s I. All VIFs of included variables were < 1.7, i.e., low enough to allow inclusion in the same model (Zuur et al. 2009). There was no autocorrelation in any model (Moran’s I, all P > 0.14).
Results
We sampled (caught and/or observed) 309 bees from 25 species, and 124 hoverflies from 14 species (Table 1; Online resource: Tables 3 and 4). The bee sample was dominated by Amegilla spp. (N = 109) and Tetragonula spp. (N = 83), and the hoverfly sample by S. grandicornis (N = 57). The vast majority of bees sampled were flower generalists (polylectic), with only 16 individuals from four specialist (oligolectic) species, all from the Megachilidae, (Table 3 Online resource).
Associations between backyards design types, trees, and urban gradients
There were no significant associations between the three backyard design types and the urban gradients at either spatial scale (all P > 0.09). The number of eucalypt trees inside backyards was not related to urban gradients at the 100 m scale (all P > 0.10). However, at the 500 m scale, there was a positive association between the number of eucalypts and tree canopy cover (slope estimate = 0.6522, Z = 2.367, P = 0.0180) and a non-significant tendency for a negative association to human density (est = -0.6475, Z = -1.873, P = 0.0611). The number of flowering trees within 50 m of backyards was positively related to tree canopy cover (est = 0.18806, Z = 0.09023, P = 0.0371), and negatively related to human population density (est = -0.21041, Z = -1.961, P = 0.0499) at the 100 m scale. There were no significant associations between flowering trees and gradients at the 500 m scale (all P > 0.15).
Associations between pollinators, local and landscape scale resources
Species richness of bees was positively related to local transect flower abundance in models at both spatial scales (Table 2; Fig. 3). Neither backyard design types, nor any of the urban gradients at either spatial scale, showed significant associations with bee species richness.

Association between bee species richness and flower abundance in survey transects, as predicted from a GLM at the neighbourhood 500 m scale. Grey dots show observations and shaded area show 95% confidence interval
Abundance of non-eusocial bees was significantly related to an interaction between human population density and tree canopy cover at the 100 m scale (Table 2), such that the association was positive at low levels of human population density but turned negative at high human densities (Fig. 4a). The proportion of grass and shrub cover at the 100 m scale was not significantly associated with bee abundance (Table 2). Non-eusocial bee abundance was positively associated with flower abundance in surveyed transects (Table 2; Fig. 4b). The three backyard design types differed significantly (Table 2), such that mixed vegetation (Tukey post hoc: Z = 2.781, P = 0.0149; mean = 4.38, 95% CI: 3.64–5.27) and lawn dominated backyards (Z = 3.118, P = 0.0051; mean = 4.78, CI: 3.90–5.86) had higher bee abundance than yards with an overall low vegetation cover and dominated by “other features” (mean = 2.91, CI: 2.34–3.62).

a Model (GLM) predicted association between non-eusocial bee abundance and the interaction of tree cover and human population density at the 100 m scale, illustrated by plotting bee abundance in relation to tree cover at three population levels: low (156 inhabitants in 100 m radius), medium (230 inhabitants) and high (349 inhabitants), b the association between non-eusocial bee abundance and the local flower abundance in surveyed transects, and c grass and shrub cover at the 500 m scale. Grey dots show observations and shaded area show 95% confidence intervals
At the 500 m scale, the abundance of non-eusocial bee species was negatively related to cover of grass and shrubs (Fig. 4c), while a significant interaction showed that the effect of tree cover again was moderated by human population density (Table 2). As for the 100 m scale, bee abundnace was positively associated to tree cover at low population levels, while the association turned negative at high levels of human density. Again, flower abundance in transects was postively related to bee abundance, and the effect of backyard design groups was significant (Table 2; Fig. 5), with more bees observed in mixed vegetation (Tukey post hoc: Z = 2.625, P = 0.0235; mean = 4.60, 95% CI: 3.71–5.71) and lawn dominated (Z = 2.837, P = 0.0127; mean = 4.62, CI: 3.73–5.72) backyards than the “other features” type (mean = 2.82, CI: 2.27–3.50).
Abundance of eusocial bees was significantly related to an interaction between human population density and grass and shrub cover at the 100 m scale, (Table 2). Eusocial bee abundance was negatively related to grass and shrub cover at low population levels, but the association turned positive at high population levels, (Fig. 6a). The abundance of eusocial bee species was negatively associated with tree cover within 100 m (Fig. 6b). Bee abundance was strongly positively associated with flower abundance in the surveyed transect, while the backyard design types did not differ in eusocial bee abundance (Table 2).

Boxplot of observed abundance (number of individuals) of non-eusocial bees in the three backyard design types. The design types were obtained through a cluster analysis of vegetation, soil and design features surveyed in the field (see Methods). The type “other features” was characterised by low cover of flowers, lawn and shrubs, and high cover of e.g., open soil, flowerbeds, vegetable patches, leaf litter and tall grass. Type “mixed vegetation” was characterised by high cover of both flowers, lawn, trees, and shrubs. Type “lawn-dominated “ was characterised by very high cover of lawn, and low cover of all other elements

Model (GLM) predicted association between eusocial bee abundance and a the interaction between tree cover and human density at the 100 m scale, illustrated by plotting bee abundance in relation to tree cover at three population levels: low (156 inhabitants in 100 m radius), medium (230 inhabitants) and high (349 inhabitants), b the proportion tree cover at the 100 m scale, c human population density at the 500 m scale, and d proportion grass and shrub cover at the 500 m scale. Grey dots show observations and shaded area show 95% confidence intervals
At the 500 m scale, the abundance of eusocial bees was positively related to human population density, and negatively related to proportion grass and shrub cover (Table 2; Fig. 6c, d). Again, flower abundance in transects had a strong positive effect on bee abundance, while backyard types did not differ (Table 2).
The abundance of hoverflies was positively related to local cover of both grass and shrubs, and tree canopy, and negatively related to human population density at both spatial scales (Table 2; Fig. 7). Hoverfly abundance was positively associated with flower abundance in the surveyed transect, while backyard types did not differ significantly (Table 2).

Model (GLM) predicted association between hoverfly abundance and a proportion grass and shrub cover at the 500 m scale, b tree cover at the 500 m scale, and c human population density at the 500 m scale. Grey dots show observations and shaded area show 95% confidence intervals
Discussion
Contrary to expectations, our results show that the association between bee abundance and urban vegetation cover shifted along the observed human population density gradient. Moreover, the direction of associations differed between eusocial and non-eusocial bees. Such differences in responses between bees with different functional traits, in combination with shifts in the direction of association to vegetation along population gradients, may explain why meta-analyses (e.g. De Palma et al. 2016) show mixed responses of bees to urbanisation. Analyses using a single measure of urbanisation (e.g., human density, vegetation, impervious surface, or an amalgamated urbanisation gradient) will not detect such shifts. Our results therefore point to the need for measures that better capture vegetation quality, especially to be able to inform urban planning and greening. Although vegetation cover and human population density can be considered two different components of urbanisation, they are often combined into an urbanisation gradient in urban biodiversity studies (Wenzel et al. 2019). However, from an urban planning perspective, it is possible to build neighbourhoods that are both dense and green if homes are smaller and/or buildings taller while interstitial or surrounding green space is extensive (Lin and Fuller 2013). We designed this study so that these two factors were sufficiently independent to allow evaluation of both their separate and combined (interactive) effects on insect pollinators.
Bee species richness positively associated to local flower abundance
The positive association found between bee species richness and availability of flowers in surveyed transects is in line with previous research, e.g., reviewed by Winfree et al. (2011) and Wenzel et al. (2019). Thus, habitat improvements in individual backyards may benefit urban bees, at least the generalist species that dominated our sample. Contrary to expectations, we found no effects of urban gradients on bee species richness, which has been shown by a study in Perth (Western Australia), where species richness in backyards and remnant bushland decreased with proportion of built space and distance to nearest bushland (Prendergast et al. 2022b). This may be a result of our relatively low sample size, and consequently that only a small proportion of the species pool was sampled (see below). There can be a high species turnover of bees between different type of urban green spaces, such as backyards, parks, and semi-natural sites (Martins et al. 2017; Banaszak-Cibicka and Żmihorski 2020; Prendergast et al. 2022b). As we only sampled backyard habitats, we may subsequently have missed the part of the bee community dependent on natural or semi-natural habitat. Alternatively, our proxies for habitat quality (the urban gradients) did not capture true habitat quality for the larger bee community. However, the fact that we found significant relationships between urban gradients and pollinator abundance suggests that we used relevant proxies for bee habitat quality.
Tree canopy cover is positively associated with non-eusocial bees, but only in low-density blocks and neighbourhoods
We found a positive association between non-eusocial bee abundance and tree cover at low levels of human population density, but a negative one at high human densities. This effect was consistent across spatial scales. Our sample of non-eusocial bees was dominated by generalist species, mainly Amegilla spp., but did include a few specialists, as well as a mix of cavity and ground nesting species. Foraging and reproduction of generalist solitary bee species have been shown to respond to local availability of single flowering species (Persson et al. 2018). A recent study found that the number of provisioned brood cells of non-eusocial bees in urban parks increased with the proportion of native plants locally (Sexton et al. 2021), and it has been suggested that flowering phenology of managed and non-native plants may not match the activity period of native pollinators (Harrison and Winfree 2015). Assuming that more ornamental and intensively managed vegetation is found in more densely populated areas (Kowarik 2008; Aronson et al. 2017), it is therefore possible that the present result is caused by a reduction in quality of trees as foraging resources, and possibly also as nesting habitat, for non-eusocial bees along the human density gradient. Although not directly tested here, this is supported by the negative relation found between human density at the block scale and the number of (native and non-native) flowering trees in and near backyards, as well as the tendency for lower numbers of (native) eucalypt trees in backyards in more densely populated neighbourhoods. The negative associations between non-eusocial bee abundance and grass and shrub cover at the neighbourhood (500 m) scale further suggests that non-eusocial bee species in the Brisbane region of Australia do not prefer open urban habitats. In addition, the presence of pesticides, both by direct application (Larson et al. 2013) and through purchased ornamentals (Lentola et al. 2017), may reduce habitat quality in backyards for bees, and non-eusocial species may be particularly negatively affected (Rundlöf et al. 2015).
Eusocial generalist bees benefit from human modified habitats
In contrast to non-eusocial species, eusocial bees (A. mellifera and Tetragonula spp.) were negatively associated with tree cover at the local block scale. In addition, we found that grass and shrub cover had a negative association with bee abundance at low human population density, but a positive one at high population density. At the larger neighbourhood scale, there was a positive association of bee abundance with human density, and a negative association with grass and shrub cover. These results indicate that, at local spatial scales, eusocial generalist species can capitalise on highly human modified green spaces, which include exotic ornamental shrubs and herbaceous flowers (Threlfall et al. 2015; Kaluza et al. 2016). In fact, urbanisation may even favour exotic bee species (Makinson et al. 2017; Wilson and Jamieson 2019), in our case the European honeybee. The affinity to humans among eusocial bees could partly be related to domesticated hives being kept in backyards. Urban beekeeping, using both honeybees and stingless bees, is indeed an increasingly common hobby in Australia (Heard 2016).
However, the negative effect of grass and shrub cover at larger spatial scales suggests that residential neighbourhoods dominated by such vegetation do not provide enough resources for eusocial bees, possibly because of a lack of habitat variation and complementation (Dunning et al. 1992) over the extended activity period of these perennial species. In addition to flower resources, stingless bees require resins collected from wounded trees (Kaluza et al. 2016), which is more likely found in mature trees (Roulston and Goodell 2010). The larger scale used here (500 m) corresponds well to stingless bee foraging ranges (Kaluza et al. 2016; Smith et al. 2017), making lack of habitat complementarity a possible explanation.
All green is good for generalist hoverflies
As expected, hoverfly abundance was positively associated with vegetation cover (both of trees, grass, and shrubs) and negatively associated to human population density, at both examined spatial scales. This suggests that increased vegetation cover partly can offset negative effects of dense urban areas on hoverflies. Only the adult life stage of hoverflies forage from flowers, while larvae are, for example, predators on aphids, decomposers, or live in aquatic environments. Most gardens likely lack a variety of these features due to management, including pesticide use, design, and plant choice. The dominant species, S. grandicornis, is a habitat generalist whose larvae feed on aphids and soft caterpillars and is one of Australia’s most common hoverflies (Soleyman-Nezhadiyan and Laughlin 1998). The abundance of tree dwelling invertebrates, including aphids and caterpillars, can be reduced in urban non-native (compared to native) vegetation (Jensen et al. 2022), possibly due to lack of co-evolution (Ehrlich and Raven 1964), with potential negative bottom-up effects on predatory insects. Combined, this could explain the overall low hoverfly sample size. As hoverflies can benefit from remnant natural or semi-natural habitats in both urban (Dylewski et al. 2019) and rural (Verboven et al. 2014) areas, the general positive effect of vegetation cover may thus result from higher availability of such resources at landscape scales, and subsequent spill-over of individuals into backyards. Interestingly, the results indicate that not even generalist aphidofagous hoverflies are common in densely populated blocks and neighbourhoods. Our results thus corroborate studies showing that hoverflies are more sensitive to urbanisation than bees (e.g. Bates et al. 2011; Verboven et al. 2014; Persson et al. 2020). As hoverflies are important pollinators (Rader et al. 2016) and contribute to other ecosystem functions (pest insect control, nutrient cycling), loss of hoverflies may lead to shifts in several ecosystem services in urbanised landscapes.
Limitations to the study
The sampling throughout November covered part of the peak season of insect pollinators in the region. Despite three survey rounds, we obtained a small sample compared to equal efforts in temperate urban study systems (pers. obs.). A likely reason is that bees mainly foraged from flowering trees and were therefore less attracted to the comparably low flower density (of mainly exotic species) in surveyed ground transects, resulting in low bee densities. Similar results were found in Sydney, Australia, where ground level visual surveys were unsuccessful in natural habitats dominated by flowering trees (Makinson et al. 2017). Our results therefore mainly give an indication on the availability of insect pollinators to herbaceous backyard plants and crops, and we cannot rule out that there are other patterns in bee diversity across Brisbane that we did not capture. Obtaining a larger sample with more species represented, for example through traps placed in flowering trees, may have allowed us to detect effects of urban gradients on species richness of both bees and hoverflies, and to look closer into which pollinator traits that may drive such relationships. However, we believe our main results (positive effects of tree cover for native non-eusocial bees, and high affinity to human habitats for eusocial generalist bees) would likely persist even if a larger part of the species pool were to be included in analyses. Including more pollinator taxa, such as Lepidopterans which are known to be sensitive to urbanisation (e.g., Chong et al. 2014; Theodorou et al. 2020), may also have rendered detectable effects on species richness.
Suggestions for local and landscape scale urban greening to benefit insect pollinators
The abundance of all pollinator taxa was positively associated with local flower abundances in surveyed transects. However, only non-eusocial bees differed in abundance between backyard design types, with higher abundances in the two types with more vegetation cover (both mixed vegetation and dominated by lawn). Non-eusocial bees, containing many small bodied species are more restricted in foraging range than the eusocial species (especially A. mellifera) (Greenleaf et al. 2007), and require nesting and foraging habitats at close distance. In contrast, eusocial bees are more efficient in using large but ephemeral flower resources because of recruitment of foragers into the same patch (Beekman and Ratnieks 2000), compared to foraging by a single female of non-eusocial species. Thus, backyard designs with high vegetation cover, and flowering trees and shrubs, can particularly benefit non-eusocial bees. If, on the other hand, the objective of a backyard owner is to attract large numbers of bees to pollinate garden crops, our results indicate that addition of flowering herbaceous ornamentals and reduced tree cover can increase the abundance of honeybees and stingless bees locally. Interestingly, the backyard design types were not related to any of the urban gradients, potentially because individual life-style identity is more important in shaping gardening preferences than surrounding neighbourhood features (Grove et al. 2006). This may open up for information campaigns, or so called “nudging”, to promote pollinator friendly design and management.
At the block and neighbourhood scales, an increased tree cover, including flowering species, will improve urban habitats both for non-eusocial bees and for hoverflies, and provide landscape scale habitat complementation for generalist eusocial bees. Because a large part of the Australian bee community will benefit from native flowering plants, especially trees within the Myrtaceae (Michener 1965; Threlfall et al. 2015), retaining or increasing such vegetation may be particularly important to promote bee diversity. Retaining remnant natural vegetation and green spaces with less intensive management (e.g., leaving dead plant material) may further benefit hoverflies by providing larval habitats, and subsequently facilitate spill-over into backyards. Urbanisation in Australia has occurred recently, compared to e.g., Europe, and remnant natural areas in and surrounding cities can harbour a high diversity of specialised and rare species (Hahs et al. 2009a, b; Ives et al. 2016). This makes urban conservation efforts particularly valuable. Urban areas are growing increasingly dense, with smaller backyards as a consequence (Sushinsky et al. 2017). It is therefore pivotal to both provide ample tree cover from public green spaces (Threlfall et al. 2017) and to promote biodiversity friendly gardening (Wenzel et al. 2019), in order to support urban biodiversity and important ecosystem service providers, such as bees and hoverflies.
Availability of data and material
Pinned insect specimen are kept at the Biological Museum, Lund University. We intend to archive our data at Dryad.org upon acceptance of the manuscript.
Code availability
Pre-existing functions in R were used and are referred to in Methods.
References
Alberti M, Marzluff JM, Shulenberger E, Bradley G, Ryan C, Zumbrunnen C (2003) Integrating humans into ecology: Opportunities and challenges for studying urban ecosystems. Bioscience 53:1169–1179. https://doi.org/10.1007/978-0-387-73412-5_9
Aronson MFJ, Lepczyk CA, Evans KL, Goddard MA, Lerman SB, MacIvor JS et al (2017) Biodiversity in the city: key challenges for urban green space management. Front Ecol Environ 15:189–196. https://doi.org/10.1002/fee.1480
Baldock KCR, Goddard MA, Hicks DM, Kunin WE, Mitschunas N, Morse H et al (2019) A systems approach reveals urban pollinator hotspots and conservation opportunities. Nat Ecol Evol 3:363–373. https://doi.org/10.1038/s41559-018-0769-y
Baldock KCR (2020) Opportunities and threats for pollinator conservation in global towns and cities. Curr Opin Insect Sci 38:63–71. https://doi.org/10.1016/j.cois.2020.01.006
Banaszak-Cibicka W, Żmihorski M (2012) Wild bees along an urban gradient: winners and losers. J Insect Conserv 16:331–343. https://doi.org/10.1007/s10841-011-9419-2
Banaszak-Cibicka W, Żmihorski M (2020) Are cities hotspots for bees? Local and regional diversity patterns lead to different conclusions. Urban Ecosyst 23:713–722. https://doi.org/10.1007/s11252-020-00972-w
Barnes MR, Nelson KC, Dahmus ME (2020) What’s in a yardscape? A case study of emergent ecosystem services and disservices within resident yardscape discourses in Minnesota. Urban Ecosyst 23:1167–1179. https://doi.org/10.1007/s11252-020-01005-2
Bates AJ, Sadler JP, Fairbrass AJ, Falk SJ, Hale JD, Matthews TJ (2011) Changing bee and hoverfly pollinator assemblages along an urban-rural gradient. PLoS ONE 6:e23459. https://doi.org/10.1371/journal.pone.0023459
Beekman M, Ratnieks FLW (2000) Long-range foraging by the honey-bee, Apis mellifera L. Funct Ecol 14:490–496. https://doi.org/10.1046/j.1365-2435.2000.00443.x
Buchholz S, Gathof AK, Grossmann AJ, Kowarik I, Fischer LK (2020) Wild bees in urban grasslands: Urbanisation, functional diversity and species traits. Landsc Urban Plan 196:103731. https://doi.org/10.1016/j.landurbplan.2019.103731
Burdine JD, McCluney KE (2019) Interactive effects of urbanization and local habitat characteristics influence bee communities and flower visitation rates 190:715–723. https://doi.org/10.1007/s00442-019-04416-x
Caynes RJC, Mitchell MGE, Wu D, Johansen K, Rhodes JR (2016) Using high-resolution LiDAR data to quantify the three-dimensional structure of vegetation in urban green space. Urban Ecosyst 19:1749–1765. https://doi.org/10.1007/s11252-016-0571-z
Chong KY, Teo S, Kurukulasuriya B, Chung YF, Rajathurai S, Tan HTW (2014) Not all green is as good: Different effects of the natural and cultivated components of urban vegetation on bird and butterfly diversity. Biol Cons 171:299–309. https://doi.org/10.1016/j.biocon.2014.01.037
De Palma A, Abrahamczyk S, Aizen MA, Albrecht M, Basset Y, Bates A et al (2016) Predicting bee community responses to land-use changes: Effects of geographic and taxonomic biases. Sci Rep 6:31153. https://doi.org/10.1038/srep31153
Dunning JB, Danielson BJ, Pulliam HR (1992) Ecological processes that affect populations in complex landscapes. Oikos 65:169–175. https://doi.org/10.2307/3544901
Dylewski Ł, Maćkowiak Ł, Banaszak-Cibicka W (2019) Are all urban green spaces a favourable habitat for pollinator communities? Bees, butterflies and hoverflies in different urban green areas. Ecol Entomol 44:678–689. https://doi.org/10.1111/een.12744
Ehrlich PR, Raven PH (1964) Butterflies and plants: a study in coevolution. Evolution 18(4):586–608. https://doi.org/10.1111/j.1558-5646.1964.tb01674.x
Foley JA, Defries R, Asner GP, Barford C, Bonan G, Carpenter SR et al (2005) Global consequences of land use. Science 309:570–574. https://doi.org/10.1126/science.1111772
Fortel L, Henry M, Guilbaud L, Guirao AL, Kuhlmann M, Mouret H et al (2014) Decreasing abundance, increasing diversity and changing structure of the wild bee community (Hymenoptera: Anthophila) along an urbanization gradient. PLoS ONE 9:e104679. https://doi.org/10.1371/journal.pone.0104679
Geslin B, Le Féon V, Folschweiller M, Flacher F, Carmignac D, Motard E, Perret S, Dajoz I (2016) The proportion of impervious surfaces at the landscape scale structures wild bee assemblages in a densely populated region. Ecol Evol 6(18):6599–6615. https://doi.org/10.1002/ece3.2374
Goddard MA, Dougill AJ, Benton TG (2010) Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol Evol 25:90–98. https://doi.org/10.1016/j.tree.2009.07.016
Goddard MA, Dougill AJ, Benton TG (2013) Why garden for wildlife? Social and ecological drivers, motivations and barriers for biodiversity management in residential landscapes. Ecol Econ 86:258–273. https://doi.org/10.1016/j.ecolecon.2012.07.016
Goulson D, Nicholls E, Botias C, Rotheray EL (2015) Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science. https://doi.org/10.1016/10.1126/science.1255957
Greenleaf S, Williams NM, Winfree R, Kremen C (2007) Bee foraging ranges and their relationship to body size. Oecologia 153:589–596. https://doi.org/10.1007/s00442-007-0752-9
Grove JM, Troy AR, O’Neil-Dunne JPM, Burch WR, Cadenasso ML, Pickett STA (2006) Characterization of households and its implications for the vegetation of urban ecosystems. Ecosystems 9:578–597. https://doi.org/10.1007/s10021-006-0116-z
Haaland C, van den Bosch CK (2015) Challenges and strategies for urban green-space planning in cities undergoing densification: A review. Urban For Urban Green 14:760–771. https://doi.org/10.1016/j.ufug.2015.07.009
Hahs AK, McDonnell MJ, Mccarthy MA, Vesk PA, Corlett RT, Norton BA et al (2009a) A global synthesis of plant extinction rates in urban areas. Ecol Lett 12(11):1165–1173. https://doi.org/10.1111/j.1461-0248.2009.01372.x
Hanson HI, Eckberg E, Widenberg M, Olsson JA (2021) Gardens’ contribution to people and urban green space. Urban For Urban Green 127198. https://doi.org/10.1016/j.ufug.2021.127198
Hagler JR, Mueller S, Teuber LR, Machtley SA, Van Deynze A (2011) Foraging range of honey bees, Apis mellifera, in alfalfa seed production fields. J Insect Sci 11:1–12. https://doi.org/10.1673/031.011.14401
Hahs AK, McDonnell MJ (2006) Selecting independent measures to quantify Melbourne’s urban–rural gradient. Landsc Urban Plan 78:435–448. https://doi.org/10.1016/j.landurbplan.2005.12.005
Hahs AK, McDonnell MJ, Mccarthy MA, Vesk PA, Corlett RT, Norton BA et al (2009b) A global synthesis of plant extinction rates in urban areas. Ecol Lett 12:1165–1173. https://doi.org/10.1111/j.1461-0248.2009.01372.x
Hall DM, Camilo GR, Tonietto RK, Ollerton J, Ahrné K, Arduser M et al (2017) The city as a refuge for insect pollinators. Conserv Biol 31:24–29. https://doi.org/10.1111/cobi.12840
Hall MA, Nimmo DG, Cunningham SA, Walker K, Bennett AF (2019) The response of wild bees to tree cover and rural land use is mediated by species’ traits. Biol Cons 231:1–12. https://doi.org/10.1016/j.biocon.2018.12.032
Harrison T, Winfree R (2015) Urban drivers of plant-pollinator interactions. Funct Ecol 29:879–888. https://doi.org/10.1111/1365-2435.12486
Hartig F (2020) DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. http://florianhartig.github.io/DHARMa/
Heard T (2016) The Australian Native Bee Book, keeping stingless bee hives for pets, pollination and sugarbag honey. Sugarbag Bees, Brisbane, Qld, Australia
Hijmans RJ (2016) raster: Geographic Data Analysis and Modeling. R package version 2.5–8. Available at: https://CRAN.R-project.org/package=raster
Hinners SJ, Kearns CA, Wessman CA (2012) Roles of scale, matrix, and native habitat in supporting a diverse suburban pollinator assemblage. Ecol Appl 22:1923–1935. https://doi.org/10.1890/11-1590.1
Hsieh TC, Ma KH, Chao A (2016) iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol Evol 7:1451–1456. https://doi.org/10.1111/2041-210X.12613
Ignatieva M, Hedblom M (2018). An alternative urban green carpet. Science, 362,148–149. https://www.science.org/doi/https://doi.org/10.1126/science.aau6974
Ives CD, Lentini PE, Threlfall CG, Ikin K, Shanahan DF, Garrard GE et al (2016) Cities are hotspots for threatened species. Glob Ecol Biogeogr 25:117–126. https://doi.org/10.1111/geb.12404
Kaluza BF, Wallace H, Heard TA, Klein AM, Leonhardt SD (2016) Urban gardens promote bee foraging over natural habitats and plantations. Ecol Evol 6:1304–1316. https://doi.org/10.1002/ece3.1941
Jensen JK, Jayousi S, von Post M, Isaksson C, Persson AS (2022) Contrasting effects of tree origin and urbanization on invertebrate abundance and tree phenology. Ecol Appl 32(29):e2491. https://doi.org/10.1002/eap.2491
Kowarik I (2008) On the role of alien species in urban flora and vegetation. Pages 321–338 in Marzluff JM, Shulenberger E, Endlicher W, Alberti M, Bradley G, Ryan C, Simon U, ZumBrunnen C, editors. Urban Ecology: An International Perspective on the Interaction Between Humans and Nature. Springer US, Boston, MA. https://doi.org/10.1007/978-0-387-73412-5_20
Kuussaari M, Toivonen M, Heliölä J, Pöyry J, Mellado J, Ekroos J et al (2020) Butterfly species’ responses to urbanization: differing effects of human population density and built-up area. Urban Ecosyst 24:515–527. https://doi.org/10.1007/s11252-020-01055-6
Larson JL, Redmond CT, Potter DA (2013) Assessing insecticide hazard to bumble bees foraging on flowering weeds in treated lawns. PLoS ONE 8:e66375. https://doi.org/10.1371/journal.pone.0066375
Lentola A, David A, Abdul-Sada A, Tapparo A, Goulson D, Hill EM (2017) Ornamental plants on sale to the public are a significant source of pesticide residues with implications for the health of pollinating insects. Environ Pollut 228:297–304. https://doi.org/10.1016/j.envpol.2017.03.084
Lin BB, Egerer MH (2017) Urban Agriculture - An opportunity for biodiversity and food provision in urban landscapes. In Ossola A, Niemelä J (eds). Urban Biodiversity - From Research to Practice. Routledge London, UK, p. 274. https://doi.org/10.9774/GLEAF.9781315402581_6
Lin BB, Fuller RA (2013) FORUM: Sharing or sparing? How should we grow the world’s cities? J Appl Ecol 50:1161–1168. https://doi.org/10.1111/1365-2664.12118
Loram A, Tratalos J, Warren PH, Gaston KJ (2007) Urban domestic gardens (X): The extent & structure of the resource in five major cities. Landscape Ecol 22:601–615. https://doi.org/10.1007/s10980-006-9051-9
Maechler M, Rousseeuw P, Struyf A, Hubert M, Hornik K, Studer M et al (2020) "Finding Groups in Data": Cluster Analysis Extended Rousseeuw et al. https://svn.r-project.org/R-packages/trunk/cluster/
Makinson JC, Threlfall CG, Latty T (2017) Bee-friendly community gardens: Impact of environmental variables on the richness and abundance of exotic and native bees. Urban Ecosyst 20:463–476. https://doi.org/10.1007/s11252-016-0607-4
Martins KT, Gonzalez A, Lechowicz MJ (2017) Patterns of pollinator turnover and increasing diversity associated with urban habitats. Urban Ecosyst 20:1359–1371. https://doi.org/10.1007/s11252-017-0688-8
Matteson KC, Langellotto GA (2010) Determinates of inner city butterfly and bee species richness. Urban Ecosyst 13:333–347. https://doi.org/10.1007/s11252-010-0122-y
McKinney ML (2008) Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst 11:161–176. https://doi.org/10.1007/s11252-007-0045-4
Meyer B, Jauker F, Steffan-Dewenter I (2009) Contrasting resource-dependent responses of hoverfly richness and density to landscape structure. Basic Appl Ecol 10:178–186. https://doi.org/10.1016/j.baae.2008.01.001
Michener CD (1965) A classification of the bees of the Autralian and South Pacific regions. Bull Am Mus Nat Hist 130:1–394
Michener CD (2007) Bees of the world, 2nd Edition, 2nd edn. John Hopkins University Press, Baltimore
Moquet L, Laurent E, Bacchetta R, Jacquemart A-L (2018) Conservation of hoverflies (Diptera, Syrphidae) requires complementary resources at the landscape and local scales. 11:72–87. https://doi.org/10.1111/icad.12245
Ollerton J, Winfree R, Tarrant S (2011) How many flowering plants are pollinated by animals? Oikos 120:321–326. https://doi.org/10.1111/j.1600-0706.2010.18644.x
PaDIL, website. https://www.padil.gov.au/. Accessed Dec 2020
Palma AD, Kuhlmann M, Roberts SPM, Potts SG, Börger L, Hudson LN et al (2015) Ecological traits affect the sensitivity of bees to land-use pressures in European agricultural landscapes. J Appl Ecol 52:1567–1577. https://doi.org/10.1111/1365-2664.12524
Pasher J, Mitchell SW, King DJ, Fahrig L, Smith AC, Lindsay KE (2013) Optimizing landscape selection for estimating relative effects of landscape variables on ecological responses. J Landsc Ecol 28:371–383. https://doi.org/10.1007/s10980-013-9852-6
Persson AS, Ekroos J, Olsson P, Smith HG (2020) Wild bees and hoverflies respond differently to urbanisation, human population density and urban form. Landsc Urban Plan 204:103901. https://doi.org/10.1016/j.landurbplan.2020.103901
Persson AS, Mazier F, Smith HG (2018) When beggars are choosers—How nesting of a solitary bee is affected by temporal dynamics of pollen plants in the landscape. Ecol Evol 8:5777–5791. https://doi.org/10.1002/ece3.4116
Persson AS, Smith HG (2013) Seasonal persistence of bumblebee populations is affected by landscape context. Agric Ecosyst Environ 165:201–209. https://doi.org/10.1016/j.agee.2012.12.008
Pott SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE (2010) Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 25:345–353. https://doi.org/10.1016/j.tree.2010.01.007
Potts SG, Imperatriz-Fonseca V, Ngo HT, Aizen MA, Biesmeijer JC, Breeze TD et al (2016) Safeguarding pollinators and their values to human well-being. Nature (London) 540:220–229. https://doi.org/10.1038/nature20588
Prendergast KS, Dixon KW, Bateman PW (2022a) A global review of determinants of native bee assemblages in urbanised landscapes. Insect Conserv Divers 1–22. https://doi.org/10.1111/icad.12569
Prendergast KS, Tomlinson S, Dixon KW, Bateman PW, Menz MHM (2022b) Urban native vegetation remnants support more diverse native bee communities than residential gardens in Australia’s southwest biodiversity hotspot. Biol Cons 265:109408. https://doi.org/10.1016/j.biocon.2021.109408
Quistberg RD, Bichier P, Philpott SM (2016) landscape and local correlates of bee abundance and species richness in urban gardens. Environ Entomol 45:592–601. https://doi.org/10.1093/ee/nvw025
Rader RA, Bartomeus IB, Garibaldi LA, Garratt MPD, Howlett BG, Winfree RG et al (2016) Non-bee insects are important contributors to global crop pollination. Proc Natl Acad Sci USA 113:146–151. https://doi.org/10.1073/pnas.1517092112
Rotheray G, Gilbert F (2008) Phylogeny of Palaearctic Syrphidae (Diptera): evidence from larval stages. Zool J Linn Soc 127:1–112. https://doi.org/10.1111/j.1096-3642.1999.tb01305.x
Roulston TH, Goodell K (2010) The Role of Resources and Risks in Regulating Wild Bee Populations. Annu Rev Entomol 56:293–312. https://doi.org/10.1146/annurev-ento-120709-144802
Rundlöf M, Andersson GKS, Bommarco R, Fries I, Hederström V, Herbertsson L et al (2015) Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 521:77–80. https://doi.org/10.1038/nature14420
Sexton AN, Benton S, Browning AC, Emery SM (2021) Reproductive patterns of solitary cavity-nesting bees responsive to both local and landscape factors. Urban Ecosyst. In press. https://doi.org/10.1007/s11252-021-01116-4
Smith JP, Heard TA, Beekman M, Gloag R (2017) Flight range of the Australian stingless bee Tetragonula carbonaria (Hymenoptera: Apidae). Austral Entomol 56:50–53. https://doi.org/10.1111/aen.12206
Smith RM, Warren PH, Thompson K, Gaston KJ (2006a) Urban domestic gardens (VI): environmental correlates of invertebrate species richness. Biodivers Conserv 15:2415–2438. https://doi.org/10.1007/s10531-004-5014-0
Smith RM, Thompson K, Hodgson JG, Warren PH, Gaston KJ (2006b) Urban domestic gardens (IX): Composition and richness of the vascular plant flora, and implications for native biodiversity. Biol Cons 129:312–322. https://doi.org/10.1016/j.biocon.2005.10.045
Soleyman-Nezhadiyan E, Laughlin R (1998) Voracity of larvae, rate of development in eggs, larvae and pupae, and flight seasons of adults of the hoverflies Melangyna viridiceps Macquart and Symosyrphus grandicornis Macquart (Diptera: Syrphidae). Aust J Entomol 37:243–248. https://doi.org/10.1111/j.1440-6055.1998.tb01578.x
Sushinsky JR, Rhodes JR, Shanahan DF, Possingham HP, Fuller RA (2017) Maintaining experiences of nature as a city grows. Ecol Soc 22:22. https://doi.org/10.5751/ES-09454-220322
Szigeti V, Kőrösi Á, Harnos A, Nagy J, Kis J (2016) Measuring floral resource availability for insect pollinators in temperate grasslands – a review. Ecol Entomol 41:231–240. https://doi.org/10.1111/een.12298
R Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Theodorou P, Radzevičiūtė R, Lentendu G, Kahnt B, Husemann M, Bleidorn C et al (2020) Urban areas as hotspots for bees and pollination but not a panacea for all insects. Nat Commun 11:576. https://doi.org/10.1038/s41467-020-14496-6
Threlfall CG, Mata L, Mackie JA, Hahs AK, Stork NE, Williams NSG et al (2017) Increasing biodiversity in urban green spaces through simple vegetation interventions. J Appl Ecol 54:1874–1883. https://doi.org/10.1111/1365-2664.12876
Threlfall CG, Walker K, Williams NSG, Hahs AK, Mata L, Stork N et al (2015) The conservation value of urban green space habitats for Australian native bee communities. Biol Cons 187:240–248. https://doi.org/10.1016/j.biocon.2015.05.003
Twerd L, Banaszak-Cibicka W (2019) Wastelands: their attractiveness and importance for preserving the diversity of wild bees in urban areas. J Insect Conserv 23(3):573–588. https://doi.org/10.1007/s10841-019-00148-8
Venables WN, Ripley BD (2002) Modern Applied Statistics with S, 4th edn. Springer, New York
Verboven HAF, Uyttenbroeck R, Brys R, Hermy M (2014) Different responses of bees and hoverflies to land use in an urban-rural gradient show the importance of the nature of the rural land use. Landsc Urban Plan 126:31–41. https://doi.org/10.1016/j.landurbplan.2014.02.017
Wenzel A, Grass I, Belavadi VV, Tscharntke T (2019) How urbanization is driving pollinator diversity and pollination – A systematic review. Biol Cons 108321. https://doi.org/10.1016/j.biocon.2019.108321
Wilson CJ, Jamieson MA (2019) The effects of urbanization on bee communities depends on floral resource availability and bee functional traits. PLoS ONE 14:e0225852. https://doi.org/10.1371/journal.pone.0225852
Winfree R, Bartomeus I, Cariveau DP (2011) Native Pollinators in Anthropogenic Habitats. Annu Rev Ecol Evol Syst 42(42):1–22. https://doi.org/10.1146/annurev-ecolsys-102710-145042
Zattara EE, Aizen MA (2021) Worldwide occurrence records suggest a global decline in bee species richness. One Earth 4:114–123. https://doi.org/10.1016/j.oneear.2020.12.005
Zuur A, Leno EN, Walker NJ, Saveliey AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
Fieldwork: Albin Andersson, Sofía Lopez, Michelle Gibson, and Michaela Plein. Species identification: Michael Batley, Australian Museum (Sydney) and Susan Wright, Queensland Museum (Brisbane). Liam Kendall and Romain Carrié provided valuable comments on statistical methods. We are very grateful to the homeowners who participated in the study.
Funding
Open access funding provided by Lund University. Swedish research council Formas, grant no. 2014–01313 to ASP.
Author information
Authors and Affiliations
Contributions
ASP conceived the idea and designed the study with input from HGS, RF and PO. ASP and AW carried out field sampling. AW and TJS assisted in species identification and compiled the bee trait table. PO compiled land use data. ASP carried out statistical analyses with input from MMM. All authors contributed critically to the writing of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest/competing interests
The authors declare no conflict of interest.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Persson, A.S., Westman, A., Smith, T.J. et al. Backyard buzz: human population density modifies the value of vegetation cover for insect pollinators in a subtropical city. Urban Ecosyst 25, 1875–1890 (2022). https://doi.org/10.1007/s11252-022-01277-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-022-01277-w