
Abstract
This study aimed at determining factors influencing response of Sahiwal cows/heifers to fixed time artificial insemination protocol in pastoral systems in Kenya. Available cows/heifers were inspected for conformity to Sahiwal breed characteristics, parity, body condition score, and subsequently rectal palpation to determine pregnancy status, ovarian structures, and estimated ovarian diameter. Consequently, these animals were injected with 100 µg of gonadotrophin-releasing hormone. On days 7 and 9, only responsive cows/heifers were injected with 500 µg of cloprostenol and 100 µg of gonadorelin Acetate, respectively. On day 10, animals were inseminated and separated from bulls for 45 days and pregnancy diagnosis done after 90 days. Analysis of variance was performed to determine the effects of production system, parity, and ovarian structures on ovary diameters pre- and post-hormonal treatment. Logistic regression was used fitting a logit function to account for the binomial distribution of conception. Overall, 56.2%, 23.1%, and 20.7% of the animals had follicles (F), corpus luteum (CL), and corpus albicans (CA), respectively, at day 0, and 16.6%, 68.6%, and 14.8%, respectively, at day 7. Human and environmental factors had no influence on conception. Among the animal factors, only the ovarian structures at day 7 had a significant effect on conception. Ovaries with CL at this time were about 6 times significantly more likely to conceive than those with F. For higher conception rates, animals with ovaries with CL should be recruited into the FTAI program as they are significantly more likely to conceive than those with other ovarian structures.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Pastoral dairy cattle systems provide for almost 80% needs of the milk requirements of the arid and semi-arid areas (ASALs) that comprise over 80% of the Kenyan land mass (Kahi et al., 2004; Ojango et al., 2016). Milk production in these areas are mainly from indigenous cattle (East African Zebu), dual purpose cattle (Sahiwal), and their crossbred genotypes with the Bos taurus (KNBS, 2019). The Sahiwal is preferred by pastoralists to other zebu breeds due to its high milk production, growth performance, good reproductive ability (Ilatsia et al., 2007), and their relative adaptation to the challenging range conditions (Ilatsia et al., 2011). Despite the impressive results from this breed and subsequent high demand for milk and beef due to increase in population, supply of superior bulls and cows from the nucleus herd to the commercial herds remain low (Mwangi & Ilatsia 2021). Improving reproductive efficiency of the breed has been identified as one of measures that may be exploited to alleviate the problem.
Reproductive performance directly impacts on onset of milk production of cows and consequently the production herd life of the cow and the total lifetime milk production (Ojango et al., 2016). It also influences rate of replacement and the selection intensity in the herds since it affects the number of available animals for selection. Primarily, pastoral dairy systems utilize natural mating to serve their cows (Awulachew, 2021). Bulls are allowed to roam within female herds, mating with any female that show signs of Estrus. Consequently, cows calve down throughout the year. This possess management challenges especially for calves born during the dry season. In order to enhance conception, more than one bull are allowed in the herd. In such cases, it is not possible to determine the paternity of the calves born in these herds. This results into incomplete pedigrees that have negative impacts on breed improvement efforts (Wasike et al., 2011; Nilforooshan et al., 2021). In addition, keeping bulls in the herds increases the cost of herd management which could otherwise be saved if use of assisted reproductive technologies (ART) was adopted (Lardner et al., 2020).
Assisted reproductive technologies have been existence since the early twentieth century (Parish and Riley, 2011). Some of these technologies include artificial insemination (AI), oestrus synchronization (ES), semen sexing, multiple ovulation and embryo transfer (MOET), invitro fertilization (IvF), and their combinations (Daly et al., 2020; Lardner et al., 2020). The extent of use of these technologies vary with AI leading in popularity. Though AI is extensively used in specialized commercial dairy systems, its utilization remains low in pastoral systems where indigenous livestock are predominant and livestock play multiple roles (Kosgey et al., 2011; Lardner et al., 2020). The deterrence to utilization of AI in pastoral systems arises from cost, environmental, and infrastructural challenges. As a result, use of ART especially FTAI has been introduced and promoted in pastoral dairy systems to enable use of superior genetic resources especially Sahiwal cattle as a strategy towards breed improvement to enable increase in milk production.
Fixed time artificial insemination involves hormonal treatment of the cows/heifers to synchronize their oestrus cycles so that they all come on heat at relatively same time. The cows/heifers that manifest oestrus are then artificially inseminated using frozen semen. There are various protocols that have been advanced for FTAI such as 3-day, 7-day, 10-day, and 14-day protocols with varied procedures of hormonal treatment of animals (Lamb et al., 2006; Nowicki et al., 2017). Equally, the resultant conceptions from these protocols in different studies remain inconsistent. Therefore, this study aimed at determining the factors that influence response of Sahiwal cows/heifers to hormonal treatment and the success rates of insemination under FTAI in pastoral dairy systems.
Materials and methods
Study area
The study was carried out in Kajiado and Narok counties from March to August 2021. Kajiado County represented an extensive pastoral production system, while Narok County represented a semi intensive pastoral production system. Kajiado County lies between − 1° 51′ 8.57′ S and 36° 46′ 36.59′ E at altitudes between 500 and 2500 m above sea level. The county covers an area of 2,1292.7 km2. The region has four ecological zones: semi-humid to semi-arid, semi-arid, arid, and very arid. It is occasioned by seasonal rains that fall in two seasons, the long rains season fall in March to May, and the short rains are from October to December. The mean annual rainfall ranges from 300 to 800 mm. Pastoralism is the main source of livelihood for majority of the households. Livestock are grazed in communally managed or open access natural pastures. During dry seasons, livestock are migrated to other areas such as Amboseli and even in the neighboring Tanzania (County Government of Kajiado, 2019).
Narok County lies between 1° 5′ 15.1548′ S, 35° 52′ 37.4304′ E, and the altitude peaks at 3100 m above sea level. The county covers an area of 17,933.1 km2. The county has 4 agroclimatic zones, namely, humid, sub humid, semi humid, to arid and semi-arid (Suke, 2020). Two-thirds of the county are classified as semi-arid. The county has bimodal rainfall with long rains between February and June and short rains between August and November. The mean rainfall ranges from 500 to 2500 mm. Data was collected at Keiyian cooperative farm, which is located in Trans-Mara West Sub County. The altitude ranges between 900 to 1950 m above sea level with a mean annual rainfall between 1000 and 1800 mm. The rainfall is bimodal with long rains from March to May and short rains in August to November and mean temperature ranges from 14.8 to 20.3 °C (Suke, 2020).
Synchronization protocol and data collection
Animals were assembled into five insemination camps in Kajiado county distributed in three Sub-counties, namely, Kajiado Central, Mashuru, and Kajiado North. There were two camps in Kajiado central, one camp in Mashuru and two camps in Kajiado North. Narok had only one camp. The Sahiwal cows/heifers used in the study were inspected for conformity to Sahiwal breed characteristics as described by Muhuyi et al. (1999), parity and body condition score. Cows beyond 3rd parity and body score below 2.5 were excluded and thus exited the camp. The cows/heifers that passed this first stage of examination proceeded to the second stage where they were examined by rectal palpation to determine pregnancy status, ovarian structures, and estimated ovarian diameter. The ovarian structures were used to determine the cyclicity of the selected cows/heifers. Key structures palpated were follicles (F), corpus luteum (CL), and corpus albicans (CA).
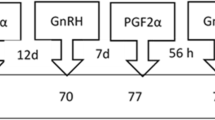
A 10-day OvSynch protocol was used for estrus synchronization and AI. At the start of estrus synchronization protocol (day zero), the selected animals were injected with 100 µg of gonadotrophin releasing hormone (GnRH) analogue gonadorelin Acetate intramuscularly on the neck muscles. The time of injection for the first and last animal injected was then recorded. On day 7, the cows/heifer recruited on day 0 were examined by rectal palpation to confirm their non-pregnancy status and their response to the hormonal treatment based on the changes in ovarian structures (follicles, corpus luteum, and corpus albicans). All non-pregnant and responsive cows/heifers were injected with 500 µg of cloprostenol intramuscularly on the neck same as day 0 injection time. On day 9, all animals that were treated on day 7 were injected with 100 µg of gonadorelin acetate at the same injection time as on day 7. On day 10, animals treated on day nine were inseminated between 14 and 16 h after the last gonadorelin acetate injection. A schematic presentation of the protocol is shown in Fig. 1.

A schematic presentation of 10-day OvSynch protocol
At the time of insemination, the identity of sire used and the inseminator were recorded. The inseminated animals continued to stay separated from the bulls for 45 more days. Pregnancy diagnosis by rectal palpation was done 90 days post-insemination.
Data preparation and analysis
The data was divided into two sets. Data set 1 included 168 animals that were selected and treated with hormones from day 0 to day 7. Data set 2 included 79 animals that went through FTAI and were pregnancy diagnosed after 90 days. Normality tests of the ovarian diameter data using the Shapiro–Wilk test showed normal distribution (p < 0.001). Analysis of variance using the PROC GLM of SAS (SAS v. 9.1, 2002) was performed to determine the effects of production system, parity, and ovarian structures on ovary diameters pre- and post-hormonal treatment. To assess animal response to GnRH, change in ovarian structures pre- (day 0) and post- (day 7) hormonal treatment were determined. Given that ovarian structure data was categorical, this was converted to count data, and thereafter, a chi-square test was used. In both analyses, data set 1 was used.
To determine factors influencing conception, a logistic model presented in Eq. 1 was fitted on data set 2. The factors included animal (parity, ovary diameter at day 0, ovarian structures at day 0, ovary diameter at day 7, and ovarian structures at day 7), environmental (production systems), and human (inseminator and time of insemination) effects. The PROC LOGISTIC function of SAS (SAS v. 9.2, 2008) was used for the analysis, and a logit function fitted to account for the binomial distribution of the response variable.
where \({y}_{ijklmn}\) is the conception status (positive or negative) of animal n; \({P}_{i}\) is the effect of ith parity (i = 0, 1, 2); \({OS}_{j}\) is the effect of jth ovarian structure (j = follicle, corpus luteum, corpus albicans); \({T}_{k}\) is the effect of kth insemination duration (k = 1, 2… 7); \({PS}_{l}\) is the effect of lth production system (l = extensive, semi-intensive); \({I}_{m}\) is the effect of mth inseminator (m = 1,2.0.4); \({\varepsilon }_{ijklmn}\) is the random error term (assumed to be independent, normally distributed with zero mean and constant variance).
Time to insemination for each individual was computed as the difference between time of insemination on day 10 and time of GnRH injection on day 9. This resulted in a range of 13 to 15 h across the study population. Time to insemination (13 to 15 h) was categorized into three groups from time 1 (T1) to time 3 (T3) with each group differing by 40 min.
Results
The number of animals, their ovarian structures, and pregnancy status are presented in Table 1. A total of 169 animals qualified for FTAI out of which 76.3% were in extensive and 23.7% in semi-intensive system. On the overall, 56.2%, 23.1%, and 20.7% of the animals had follicles, corpus luteum, and corpus albicans, respectively, at day 0, and 16.6%, 68.6%, and 14.8%, respectively, at day 7. At day 0, animals with follicles, corpus luteum, and corpus albicans in the extensive system were 60.5%, 23.3%, and 16.3% of, respectively, while in the semi-intensive system, they were 42.5%, 22.5%, and 35.0%, respectively. At day 7, however, 14.7%, 73.6%, and 11.6% of animals had follicles, corpus luteum, and corpus albicans, respectively, in the extensive system, and 22.5%, 52.5%, and 25.0%, respectively, in the semi-intensive system. Whereas only 30.0% of animals in the extensive system were available for post FTAI pregnancy diagnosis, 97.5% were available in the semi-intensive system. Out of the available animals, the conception rates were 60.0% and 59.0% in extensive and semi-intensive systems, respectively.
The factors influencing ovarian diameter at days 0 and 7 are presented in Table 2. The overall mean diameter at day 0 and 7 were 1.6 ± 0.07 cm and 1.7 ± 0.08 cm, respectively. Production system and parity had no influence on ovary diameter at both days. At day 0, ovaries with follicles (F) and corpus luteum (CL) had similar diameter that was significantly higher than for ovaries with corpus albicans (CA). At day 7, ovaries with CL had significantly higher diameter than both ovaries with F and CA.
The effect of GnRH treatment on ovarian structures pre- and post-treatment is presented in Table 3. There was a significant (P < 0.05) response in the proportion of ovarian structures between day 0 and day 7. The proportion of follicles (F) reduced by about 40%, corpus luteum (CL) increased by about 46%, while corpus albicans reduced by about 6%.
Animal, environmental, and human factors influencing conception are presented in Table 4. Human (inseminator and insemination time) and environmental (production system) factors had no influence on conception. Among the animal factors (parity, ovarian structures at day 0 and ovarian structures at day 7), only the ovarian structures at day 7 had a significant effect on conception. Ovaries with CL at this period were about 6 times significantly more likely to conceive than those with F.
Discussion
Artificial insemination (AI) is a widely used reproductive technology for dissemination of superior genetics in the high rainfall dairy zones of Kenya (Ojango et al., 2016). In the semi-arid pastoral areas, however, adoption of the technology is relatively low due to several challenges that include accessibility, affordability, estrus detection efficiency, and availability of skilled technician (Mwai et al., 2020). The application of estrus synchronization (ES) and fixed time artificial insemination (FTAI) technology is possible to increase adoption of AI in such areas. The fact that more than 169 cows/heifers were willingly availed for the study may be an indicator of the awareness and willingness of the pastoralists to adopt AI. Indeed, the demand for Sahiwal genetics in the study area has been reported to exceed the annual availability of breeding bulls from the national Sahiwal stud (Mwangi and Ilatsia, 2021).
On the overall and in both production systems, the follicle (F) was the predominant ovarian structure at day 0. Although an on-line literature search did not yield any similar study for comparative purposes, the finding in the current study was in close agreement with the results of Mahzabin et al. (2020). In that study, 62.5% of ovaries collected from an abattoir were in the follicular stage and had no CL, while the rest (37.5%) had CL. The estrus cycle is hormonally controlled and divided into estrus, metestrus, diestrus, and proestrus phases (Noseir, 2003). The proestrus and estrus are termed as the follicular phase. At the proestrus phase, pre-existing primordial follicles develop in wave-like patterns into ovarian follicles, with generally one follicle becoming dominant (Webb et al., 2016). At estrus, the dominant follicle is ovulated in readiness for fertilization. Concurrently and in preparation for fertilization, metestrus and diestrus (known as the luteal phase) are initiated. Metestrus, the early stage of CL formation is characterized by presence of corpus hemorrhagicum (CH), while diestrus is characterized by a prominent functional CL (Maina et al., 2008). If fertilization fails after ovulation, the CL regresses into a corpus albican (CA). The presence of these structures in the current study therefore indicate that the animals were cycling and that most were in the follicular phase. The ratios of the structures were not significantly different between production systems (figure not shown) indicating that the conditions in both systems were similar.
Fewer animals were available for pregnancy diagnosis (30%) in the extensive system than in the semi-intensive system (97.5%). This was due to livestock migration in search of pastures due to onset of drought. Although the Maasai community are sedentary with permanent settlements, seasonal livestock migration occurs from permanent settlements to temporal grazing areas. A study by Butt et al. (2009) suggested that household relocation reduces the stress faced by pastoralists and their cattle during the drought. Nevertheless, the conception rates observed in this study (60.0% and 59.0% in extensive and semi-intensive systems, respectively) were within the range reported for Bos indicus beef breeds in Argentina (Bó & Baruselli, 2014), irrespective of the protocol used. The results are also consistent with conception rate of 58% in a Bos taurus breed reported by Lardner et al. (2020). It may therefore be inferred that the protocol used led to acceptable conception rates.
The overall estimated ovarian diameter at day 0 and day 7 (1.6 and 1.7 cm, respectively) observed in this study are within the range of 1.5 to 1.9 cm reported by various authors in studies employing actual measurements of ovaries collected from slaughter houses (Kunbhar et al., 2003; Kouamo et al., 2014; Islam et al., 2018; Ali et al., 2021). The results by Mahzabin et al. (2020), however, reported lower diameter (1.1 and 1.2 cm) than in the current study. This may be attributed to the cyclicity status of the cows/heifers. Various studies have also shown that the nutritional status, the breed, the presence of follicles, and CL influence the size of the ovary (Cushman et al., 2014; Peralta-Torres et al., 2017). Lack of significant differences in ovarian diameter at days 0 and 7 between production systems and parities is a confirmation that the cows/heifers in the extensive and semi-intensive systems as well as in the different parities were in a similar state of normal estrus cycle (Sharma et al., 2018). The significantly lower diameter of ovaries having CA as the dominant structure than for ovaries with both F and CL as the dominant structures at both day 0 and 7 was in agreement with 1.1 and 1.2 cm reported by Mahzabin et al. (2020). These authors explained that the ovaries were obtained from animals that had been culled due to their low reproductive performance. The scenario in the current study could be associated with animals most likely being at the proestrus phase of the estrus cycle. The significantly wider ovary diameter containing CL as the dominant structure at day 7 agrees with Kouamo et al. (2014) and Ali et al. (2021) who reported significantly wider CL present ovaries than those without.
The significant increase in ovarian diameter at day 7 of the ovaries that had CL than both F and CA and the significant increase in the number of animals (46%) that had CL at day 7 (Table 3) is a clear indication of response to GnRH treatment that was carried out at day 0. Administration of GnRH for animals with F induced release of luteinizing hormone (LH) leading to ovulation of the dominant follicle and emergence of a new follicular wave about 1.5 to 2 days later (Bó & Baruselli, 2014). Additionally, luteinizing hormone (LH) and follicle-stimulating hormone (FSH) surge leads to ovarian function, such as follicular growth and maturation, ovulation, corpus luteum formation, and maintenance (Kim, 2018). Hence, some cows/heifers that had CL at day 0 persisted to day 7 after being treated with GnRH while those animals that had CA, the GnRH caused a CL to be formed. This resulted in numbers of animals that had CL to increase by about 46%, while those with F and CA reduced by about 40 and 6%, respectively.
Prostaglandin F2α and its analogues are luteolytic in cattle and are effective on CL from days 5 to 17 of the estrous cycle. Injection of cloprostenol on day 7 leads to the regression of the CL, and hence, the progesterone effect on the reproductive system is reduced leading to production of LH. Treatment with GnRH on day 9 induces release of LH and ovulation of the dominant follicle (Bó & Baruselli, 2014). In this study, conception rate was 60% indicating that 40% of animals that had F and CL at day 7 did not conceive. This is because PGF2α hormone administered on day 7 sustains the follicle, and hence, primordial follicles would endure after administration of GnRH on day 9. Additionally, primordial follicles (non-growing follicles) and growing (primary, secondary, and tertiary) follicles have been found in the ovary undergoing either development or degeneration as a wave-like pattern (Bihon and Assefa, 2021). This implies that follicles are continuously in processes of recruitment, dominance, selection, and atresia where some follicles are entering from the non-growth phase to a growing population. As a result, these animals with primordial follicles on day 9 would not conceive during insemination because of their follicles not being mature. This means that only animals that had follicles in growth phase conceived. This is in agreement with a study by de Lima et al. (2020) that showed that the pregnancy rate was higher in animals with low antral follicle count than in those with a high antral follicle count.
Successful synchronization of ovulation relies on the stage of follicular development at the time of initial GnRH treatment at day 0. For higher conception rates animals with ovaries with CL should be recruited into the FTAI program as they are significantly more likely to conceive than those with other ovarian structures.
Data availability
The datasets generated during and/or analyzed during the current study are not publicly available due to the lead institution (KALRO) policies but are available from the corresponding author on reasonable request.
References
Ali, S., Degefa, T. and Lemma, A., 2021. Morphometrical study of the reproductive tract in Boran (Bos indicus) heifers in Ethiopia. Ethiopian Veterinary Journal, 25(2), pp.1-13.
Awulachew, M.T., 2021. Assessment on cattle breeding practice and production system associated with their environmental implication in Ethiopia. Animal and Veterinary Sciences, 9(4), p.99.
Bihon, A. and Assefa, A., 2021. Prostaglandin based estrus synchronization in cattle: A review. Cogent Food & Agriculture, 7(1), p.1932051.
Bó, G.A. and Baruselli, P.S., 2014. Synchronization of ovulation and fixed-time artificial insemination in beef cattle. Animal, 8(s1), pp.144-150.
Butt, B., Shortridge, A. and WinklerPrins, A.M., 2009. Pastoral herd management, drought coping strategies, and cattle mobility in southern Kenya. Annals of the Association of American Geographers, 99(2), pp. 309-334.
County Government of Kajiado, 2019. County Integrated Development Plan 2018–2022. https://www.cog.go.ke/downloads/category/106-county-integrated-development-plan-2018-2022, Retrieved on 25th January 2023
Cushman, R.A., McNeel, A.K. and Freetly, H.C., 2014. The impact of cow nutrient status during the second and third trimesters on age at puberty, antral follicle count, and fertility of daughters. Livestock science, 162, pp.252-258.
Daly, J., Smith, H., McGrice, H.A., Kind, K.L. and van Wettere, W.H., 2020. Towards improving the outcomes of assisted reproductive technologies of cattle and sheep, with particular focus on recipient management. Animals, 10(2), p.293.
de Lima, M.A., Morotti, F., Bayeux, B.M., de Rezende, R.G., Botigelli, R.C., De Bem, T.H.C., Fontes, P.K., Nogueira, M.F.G., Meirelles, F.V., Baruselli, P.S. and da Silveira, J.C., 2020. Ovarian follicular dynamics, progesterone concentrations, pregnancy rates and transcriptional patterns in Bos indicus females with a high or low antral follicle count. Scientific reports, 10(1), pp.1-13.
Ilatsia, E.D., Muasya, T.K., Muhuyi, W.B. and Kahi, A.K., 2007. Genetic and phenotypic parameters and annual trends for milk production and fertility traits of the Sahiwal cattle in semi-arid Kenya. Tropical Animal Health and Production, 39(1), pp.37-48.
Ilatsia, E.D., Migose, S.A., Muhuyi, W.B. and Kahi, A.K., 2011. Sahiwal cattle in semi-arid Kenya: Genetic aspects of growth and survival traits and their relationship to milk production and fertility. Tropical Animal Health and Production, 43(8), pp.1575-1582.
Islam, R., Khaton, R., Islam, H., Ullah, N., Nishi, S.A., Paul, B.R., and Sarder, J.U., 2018. Biometry of ovary in different ruminant animals. Bangladesh Livestock Journal, 1, pp. 44- 48.
Kahi, A.K., Nitter, G. and Gall, C.F., 2004. Developing breeding schemes for pasture based dairy production systems in Kenya: II. Evaluation of alternative objectives and schemes using a two-tier open nucleus and young bull system. Livestock Production Science, 88(1-2), pp.179-192.
Kenya National Bureau of Statistics (KNBS) (2019). Gross County product report, Kenya National Bureau of Statistics, Nairobi, Kenya, Pp 49. https://www.knbs.or.ke/download/gross-county-product-2019/, Retrieved on 25 January 2023
Kim, S.J., 2018. Patterns of ovarian changes associated with surge mode secretion of gonadotropin in dairy cows with cyclic estrous cycle. Journal of Embryo Transfer, 33(4), pp. 297-304.
Kosgey, I.S., Mbuku, S.M., Okeyo, A.M., Amimo, J., Philipsson, J. and Ojango, J.M., 2011. Institutional and organizational frameworks for dairy and beef cattle recording in Kenya: a review and opportunities for improvement. Animal Genetic Resources/Resources génétiques animales/Recursos genéticos animales, 48, pp.1-11.
Kouamo, J., Dawaye, S.M., Zoli, A.P., and Bah, G.S., 2014. Evaluation of bovine (Bos indicus) ovarian potential for in vitro embryo production in the Adamawa plateau (Cameroon). Open Veterinary Journal, 4(2), pp. 128-136.
Kunbhar, H.K., Samo, M.U., Memon, A., and Solangi, A.A., 2003. Biometrical studies of reproductive organs of Thari cow. Pakistan Journal of Biological Sciences, 6(4), pp. 322-324.
Lamb, G.C., Larson, J.E., Geary, T.W., Stevenson, J.S., Johnson, S.K., Day, M.L., Ansotegui, R.P., Kesler, D.J., DeJarnette, J.M. and Landblom, D.G., 2006. Synchronization of estrus and artificial insemination in replacement beef heifers using gonadotropin-releasing hormone, prostaglandin F2α, and progesterone. Journal of animal science, 84(11), pp. 3000-3009.
Lardner, H., Damiran, D., and Larson, K., 2020. Comparison of FTAI and natural service breeding programs on beef cow reproductive performance, program cost and partial budget evaluation. Journal of Agricultural Science, Vol. 12, No. 9. https://doi.org/10.5539/jas.v12n9p1
Mahzabin, R., Khandoker, M., Sakhawat, H.S., Islam, M., Jahan, S.S., Habib, M., and Vargas-Bello-Perez, E., 2020. Evaluation of cattle ovaries and follicles by histological analysis for potential in vitro production of embryos in tropical conditions. Tropical and Subtropical Agroecosystems, 23(3). https://www.revista.ccba.uady.mx/ojs/index.php/TSA/article/view/3356, Retrieved on 25th January 2023
Maina, V.A., Muktar, A., and Sabo, Y.G., 2008. Effects of body condition score on ovarian activity of Bos indicus (ZEBU) Cows. Asian Journal of Scientific Research, 1(4), pp. 421- 428, https://doi.org/10.3923/ajsr.2008.421.428
Muhuyi, W.B., Lokwaleput, I. and Sinkeet, S.O., 1999. Conservation and utilisation of the Sahiwal cattle in Kenya. Animal Genetic Resources/Resources génétiques animales/Recursos genéticos animales, 26, pp.35-44.
Mwai, O.A., Tsuma, V., Owino, J.O., Ochieng., L., and Rao, E.J. O., 2020. Accelerating genetic improvement in emerging smallholder dairy systems through fixed-time and conventional artificial insemination technologies: organizational and operational experiences from Kenya. ILRI Project Report. ILRI, Nairobi, Kenya. pp 28. https://cgspace.cgiar.org/handle/10568/111149, Retrieved on 23 January 2023
Mwangi S.I., and Ilatsia E.D., 2021. Sahiwal breed in pastoral production systems in Kenya: Future roles and existing gaps in knowledge and supportive technical logistics; A Review. In: Proceedings of the XXIV International Grassland Congress / XI International Rangeland Congress, 25 – 29 October, 2021, Nairobi, Kenya. https://uknowledge.uky.edu/igc/24/3-2/18/, Retrieved on 25 January 2023
Nilforooshan, M.A., Garrick, D. and Harris, B., 2021. Alternative ways of computing the numerator relationship matrix. Frontiers in Genetics, 12:655638. https://doi.org/10.3389/fgene.2021.655638
Noseir, W., 2003. Ovarian follicular activity and hormonal profile during estrous cycle in cows: the development of 2 versus 3 waves. Reproductive Biology and Endocrinology, 1(1), pp. 1–6.
Nowicki, A., Barański, W., Baryczka, A. and Janowski, T., 2017. OvSynch protocol and its modifications in the reproduction management of dairy cattle herds–an update. Journal of veterinary research, 61(3), p.329.
Ojango, J.M. K., Wasike, C.B., Enahoro, D.K., and Okeyo, A.M., 2016. Dairy production systems and the adoption of genetic and breeding technologies in Tanzania, Kenya, India and Nicaragua. Animal Genetic Resources, 59, pp. 81–95. https://doi.org/10.1017/S2078633616000096
Parish, J.A., and Riley, J.M., 2011. Economic comparisons of artificial insemination vs. natural mating for beef cattle herds (Publication 2486). Mississippi State University Extension Service. https://extension.msstate.edu/sites/default/files/publications/publications/p2486_1.pdf, Retrieved on January 25, 2023
Peralta-Torres, J.A., Aké-López, J.R., Centurión-Castro, F.G., and Segura-Correa, J.C., 2017. Effect of season and breed group on the follicular population and cyclicity of heifers under tropical conditions. Tropical animal health and production, 49(1), pp. 207–211.
SAS Institute Inc. 2008. SAS OnlineDoc® 9.2. Cary, NC: SAS Institute Inc. SAS OnlineDoc® 9.2. http://support.sas.com/documentation/cdl/en/etlug/60948/HTML/default/images/copyrite.htm. Retrieved on 25 January 2023
Sharma, A.K, Singh M.A, Sharma A, and Kumar, P., 2018. Effect of BCS and parity on uterine involution, ovarian rebound and various fertility parameters in postpartum dairy cows. Indian Journal of Animal Sciences, 88(5), pp. 526–529.
Suke, Y.N., 2020. Factors associated with nutritional status in elderly persons living in Kilgoris, Kenya. PhD Thesis, University of Nairobi, Nairobi, pp 83. http://erepository.uonbi.ac.ke/handle/11295/153186, Retrieved on 25 January 2023
Wasike, C.B., Magothe, T.M., Kahi, A.K. and Peters, K.J., 2011. Factors that influence the efficiency of beef and dairy cattle recording system in Kenya: A SWOT–AHP analysis. Tropical animal health and production, 43(1), pp.141–152.
Webb, R., Buratini, J., Hernandez-Medrano, J.H., Gutierrez, C.G., and Campbell, B.K., 2016. Follicle development and selection: Past, present and future. Animal Reproduction, 13(3), pp. 234–249. https://doi.org/10.21451/1984-3143-AR883
Acknowledgements
Special thanks to Kenya Agricultural and Livestock Research Organization, Naivasha, for technical and computational assistance.
Funding
This work was supported by The Kenya Climate Smart Agriculture Project (KCSAP) jointly supported by Government of Kenya and World Bank (reference number (KCSAP CGS/CRGs-AD-2019).
Author information
Authors and Affiliations
Contributions
All authors contributed to conception and design of this study. Material preparation, data collection, and analysis were performed by Mwaura Magothe, Daniel Mwangi, Bwire Wasike, Ruth Waineina, Sophie Miyumo, Samson Mwangi, and Evans Ilatsia. The first draft of the manuscript was joint effort by all authors who also commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethical approval
The study was exempted from requiring ethical approval since the data used was from a community engagement project.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Magothe, T.M., Mwangi, D.K., Wasike, C.B. et al. Response to hormonal treatment and conception rates of Sahiwal cows subjected to fixed time artificial insemination in pastoral dairy systems. Trop Anim Health Prod 55, 49 (2023). https://doi.org/10.1007/s11250-023-03471-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11250-023-03471-0