
Abstract
Polyhydroxyalkanoates (PHA) are a group of biopolymers produced naturally by microorganisms with properties similar to various petroleum-based plastics. However, to date their commercial production has remained uncompetitive due to substrate, sterilization, aeration and processing costs. Purple non-sulfur bacteria (PNSB) are a group of anoxygenic photoheterotrophic bacteria that have the ability to accumulate PHA under unbalanced conditions in anaerobic environments and constant feeding with high conversion ratios. Such characteristics could potentially overcome some of the bottlenecks of conventional chemoheterotrophic PHA production. Yet these organisms have received relatively limited attention. This review explores the factors involved in the PHA accumulation process from PNSB, highlighting the differences to conventional PHA production and the areas yet to be optimized. The roles of fermentation systems, carbon substrate, feeding conditions, nutrients, pH and various aspects of light are reviewed to understand their role in PHA accumulation in PNSB.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Single-use plastics have revolutionized modern life since their commercialization. However, their disposal is challenging as the majority of plastics are non-biodegradable and can remain in the environment more than 100 years before decomposing. While other sources of biodegradable bioplastics are available, polyhydroxyalkanoates (PHA) are one of the most promising bioplastic options due to their similar properties to polyethylene (Sathya et al. 2018) and ability to produce them through microbial conversion from a wide range of organic waste streams. Despite these benefits, PHA produced commercially via single culture aerobic fermentation processes are currently uncompetitive in the market due to their high production costs. Significant costs are associated with high purity feedstocks, sterilization, aeration, and downstream processing. Microbial mixed culture (MMC) systems dominated by purple non-sulfur bacteria (PNSB) could potentially reduce the production costs of PHA, as PNSB are phototrophic and grow under anaerobic conditions, allowing easy enrichment without aeration. Moreover, their phototrophic metabolism allows high carbon to PHA conversion ratios and they are capable of using a wide range of carbon sources to grow. Recent research revealed that PHA accumulation occurs in parallel with cell growth in PNSB, unlike many chemoheterotrophs where PHA accumulation is a means of storing carbon that cannot be directed towards cell growth under concurrent conditions (Higuchi-Takeuchi and Numata 2019). Furthermore, it was established that mixed phototrophic bacteria can grow under continuous substrate availability, avoiding unproductive time associated with famine periods during the common feast-famine feeding approach used to drive PHA storage in aerobic PHA fermentation (Fradinho et al. 2016). Previous reviews examined PHA production in various microorganisms (Verlinden et al. 2007; Serafim et al. 2008; Urtuvia et al. 2014; Dietrich et al. 2017; Sali and Mackey 2021) and explored the various types of PNSB microorganisms (Imhoff et al. 2005; Kompantseva et al. 2010; Imhoff 2017a). However, while biohydrogen production using PNSB has received significant attention (Ismail et al. 2008; Carlozzi and Lambardi 2009; Laurinavichene et al. 2018), this review focuses specifically on PNSB application for PHA production. Given the recent and rapidly growing research in this area (Tanskul et al. 2016; Padovani et al. 2018; Fradinho et al. 2019; Higuchi-Takeuchi and Numata 2019), this manuscript explores the potential and bottlenecks for PHA production using PNSB within the fermentation process. Aspects related to biomass harvesting and PHA extraction, which are less specific to PNSB and well-covered elsewhere (Molina Grima et al. 2003; Christenson and Sims 2011; Anis et al. 2013; Villano et al. 2014; Samorì et al. 2015) are beyond the scope of this review.
2 Polyhydroxyalkanoates
PHA are polyesters produced naturally by various microorganisms. They are stored intracellularly as spherical polymers as a source of energy, carbon or reducing equivalents under unbalanced conditions as a survival strategy (Sathya et al. 2018; Tamang et al. 2019). PHA are both bio-based and biodegradable plastics with a short decomposition time once disposed into various types of environments, including marine environments. The decomposition duration varies from 3 to 18 months depending on the environment conditions, monomer type and bacteria present (Brandl et al. 1990; Verlinden et al. 2007; Voinova et al. 2008; Chee et al. 2010). For these reasons, PHA production has received significant attention in recent years to overcome the negative environmental impacts associated with petroleum-based plastics. Currently, there are more than 300 genera of bacteria with the ability to store PHA naturally (Choi and Lee 1999) with some prokaryote strains reported to accumulate up to 90% of their cell dry weight (CDW) as PHA under specific conditions (Możejko-Ciesielska and Kiewisz 2016). The best PHA storage conditions in traditionally applied aerobic cultures reported are under nutrient limitation, excess of carbon and intermittent feeding (Marang et al. 2018).
2.1 PHA properties
More than 150 hydroxyalkanoic acids can constitute PHA. The most widespread monomer is poly(3-hydroxyburtyrate) (PHB), but other common PHA monomers include poly(3-hydroxyvalerate) (PHV) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV). Other types of monomers can be composed of polyhydroxyvalerate (PHV), 3-hydroxyhexanoate (3HHx) and 3-hydroxyheptanoate (3HHp) (Chee et al. 2010). These PHA monomers qualify as short chain length (SCL), which consist of three to five carbons in a chain (Sharma et al. 2017). Medium chain length (MCL) PHA are composed of six to 14 carbons in a chain. They typically include monomers such as 3-hydroxyoctanoate (3HO), 3-hydroxydecanoate (3HD) and 3-hydroxydodecanoate (3HDD) (Rigouin et al. 2019).
PHA are biodegradable amorphous polymers with versatile properties and wide functionality (Koller and Braunegg 2018), which makes them a suitable candidate for bioplastic production. They have thermoplastic and elastomeric properties comparable to petroleum-based plastics such as polypropylene (PP) and polyethylene (PE) plastics (Bugnicourt et al. 2014). In particular, they have high resistance to ultraviolet light, oxygen permeability and are moisture resistant (Verlinden et al. 2007).
Their mechanical properties vary significantly depending on factors such as the monomer type(s), bacterial strains involved and their environmental conditions (Dietrich et al. 2017), as well as resulting molecular weight of the extracted PHA. SCL-PHA have high crystallinity, are brittle and demonstrate high melting temperatures (Castilho et al. 2009), while MCL-PHA are slightly flexible with a high elongation to break ratio, low melting temperatures and enhanced mechanical properties (Możejko-Ciesielska and Kiewisz 2016). PHB are the most commonly found polymer, but SCL-PHA have displayed 70 to 80% crystallinity causing their low resistance to brittle fractures (Bugnicourt et al. 2014). From a commercial perspective, PHV and MCL-PHA have a higher potential than the more commonly produced PHB.
2.2 Production process
Commercial PHA production is currently estimated to be 66,000 t y−1 (Vandi et al. 2018). The current commercial production processes rely on pure bacterial species like Cupriavidus necator and pure sources of carbon (Vandi et al. 2018; Tamang et al. 2019). While these approaches achieve high PHA storage and productivity, they do not enable PHA to compete against petroleum-based plastics due to high costs of substrate, sterilization and energy associated with aeration (Fradinho et al. 2013a). MMC, on the other hand, could eliminate the need for sterilization and allow use of diverse sources of substrate, which reduces overall production costs (Serafim et al. 2008). Although the economic and environmental benefits of such system are advantageous, MMC have not reached industrialization level yet due to challenges in polymer consistency and lower culture densities. Because the biopolymers are intracellularly stored and undergo extraction processes that destroy the cell, risks of unknown organisms frequently associated with MMC processes are not a significant issue for PHA production.
PHA production is a complex process that requires the optimization of various factors. Strain selection, adequate carbon source, suitable environmental conditions and bioreactor design are key to achieving an enhanced PHA production system. The selection of pure or mixed cultures as well as the purity and composition of carbon substrate can affect the storage abilities of the microorganisms. Predicting the monomer composition can be challenging due to the different metabolic pathways of carbon to PHA conversion. The same bacteria can produce PHA with different composition and properties due to the carbon present in the environment. Three metabolic pathways are identified for PHA production and can result in either SCL-PHA when the precursors are simple sugars via TCA (tricarboxylic acid) cycle or MCL-PHA when the precursors are fatty acids via β-oxidation and/or simple carbons via the de novo fatty acids pathway (Kniewel et al. 2019).
Various substrates have been effectively utilized for PHA production via the microbial route (Jiang et al. 2016). Carbohydrates such as simple sugars are quickly digested by bacteria for PHA production. For instance, whey, a precursor of lactose, has been used as the primary carbon source by both Povolo et al. (2013) and Ahn et al. (2000), and resulted in a mixture of 3HV and 4HB monomers by Hydrogenophaga pseudoflava in the former case and only 3HB monomers by E. coli in the latter. While carbohydrates have shown some promising results, they also represent a major food source for human beings and therefore are considered as competition and non-sustainable carbon sources. Animal fats and plant oils are composed of triacylglycerols, which have demonstrated good SCL-PHA accumulation in the case of plants oils (Akiyama et al. 1992) and MCL-PHA in the case of esters from the long-chain fatty acids present in animal fats (Muhr et al. 2013). Nonetheless, these long-chain fatty acids have major drawbacks such as high melting temperatures required for their fermentation, their immiscible properties and their inhibitory nature at high concentrations (Akiyama et al. 1992; Zhu et al. 2010). On the other hand, short-chain fatty acids such as acetate and propionate were preferred by various PHA-producing bacteria (Montano-Herrera et al. 2017). However, the ratio of even-chain (i.e. acetate) to odd-chain (i.e. propionate) can play a significant role in the monomer composition since acetate and propionate are known precursors to HB and HV monomers, respectively (Akiyama et al. 1992). Studies about hydrocarbons to PHA conversion are still limited, but the published data reports that hydrocarbons’ productivity is low (Brandl et al. 1988; Chayabutra and Ju 2001; Ni et al. 2010; Tufail et al. 2017). Nevertheless, their capability to produce MCL-PHA is better due to their longer carbon chain monomers (Ward et al. 2006) and, therefore, are promising substrates for PHA production with enhanced properties. Additionally, environmental conditions such as feeding regimen, pH and nutrients availability, mixing and light availability (for phototrophs) all play a role in the biomass growth and PHA production. For phototrophic systems in particular, bioreactor dimensions and lighting layout are essential design considerations to maintain suitable light availability.
3 Purple non-sulfur bacteria
PNSB are a subset of anoxygenic phototrophic bacteria that grow by feeding photoheterotrophically on carbon substrates under anoxygenic conditions and have the ability to accumulate PHA, polyphosphate and polysaccharides among many others (Imhoff et al. 2005; Higuchi-Takeuchi et al. 2016a; Lai et al. 2017). Table 1 summarizes some of the main findings for PHA accumulation by PNSB. They are mainly found in aquatic and marine environments that experience some degree of light exposure such as soil and wastewater (Madigan and Jung 2009). PNSB have diverse metabolic capabilities including the ability to conduct three important biological reactions; the conversion of light into chemical energy and the fixation of both CO2 and N2 (Higuchi-Takeuchi and Numata 2019). These reactions make using PNSB an environmentally friendly option for various applications. PNSB can switch between various metabolic modes for growth, with photoheterotrophic growth being the preferred. Most types of PNSB were reported to grow rapidly in environments with high content of organic compounds and show good tolerance to compounds typically considered toxic (García et al. 2019). PNSB have the photochemical reaction center photosystem II (Hanada 2016). This has a maximum absorption in the infrared (IR) region of the spectrum, which provides an advantage for the tissue penetration (Wu and Butt 2017), and has a minimum adsorption in the blue light range (Zhou et al. 2015a). PNSB contain carotenoids, BChls, high protein content, biopolymers and pantothenic acids, all of which have been explored for further applications (Merugu et al. 2012).
PNSB belong to the Proteobacteria phylum. Currently, more than 20 genera and 95 species of PNSB are recognized and classified as either Alpha-proteobacteria or Beta-proteobacteria (Imhoff 2017b). They come in different shapes such as rods, spirilla, vibrios and cocci as well as various colors including purple, red, orange, pink and brown, which are results of blends of various photo-pigments (Madigan and Jung 2009). These photopigments in PNSB are a mixture of carotenoids and BChls. Their main role is light harvesting and conversion to chemical energy. The presence of oxygen can halt their photoheterotrophic metabolism but varies significantly depending on species (Madigan and Jung 2009). While there exists a group of anoxygenic phototrophic bacteria (APB) that can grow in the presence of oxygen, commonly known as aerobic APB, they are beyond the scope of this review.
PNSB absorbance spectrum covers all of the visible range as well as the IR and UV ranges (Adessi and De Philippis 2014). Carotenoids absorb light in the range of 400-595 nm, while BChl a absorb light ranging from 805 to 930 nm and BChl b absorb lights beyond 1000 nm (Hülsen et al. 2014; Zhou et al. 2015a). PNSB can therefore utilize various wavelengths of light to produce chemical energy, with the primary advantage being their ability to utilize IR light, even with very narrow bandwidth such as with laser diodes (Bertling et al. 2006).
3.1 Metabolism
PNSB have a versatile metabolism and switch between various metabolic growth modes according to the environment’s conditions and demands. Higher growth is often observed under photoheterotrophic and photoautotrophic conditions (Pfennig et al. 1997), but PNSB can also grow under chemoheterotrophic mode (Kantachote et al. 2005). However, their activities reduce under such conditions as they do not utilize light, but rather rely on organic and inorganic carbon sources as electron donors and acceptors, respectively (Lai et al. 2017; García et al. 2019). PNSB cannot utilize water as an electron donor and rather rely on organic compounds and sulfide (Imhoff et al. 2005; Hanada 2016). Under a photoautotrophic mode, PNSB use light as an energy source and sulfide or hydrogen as electron donor and CO2 as an electron acceptor. Under a photoheterotrophic mode, PNSB utilize organic compounds like organic acids as electron donors, as well as light as energy source (Hülsen et al. 2014). This is their preferred mode of growth. Photoheterotrophic production of PHA is the predominant route in PNSB, but photoelectroautotroph and photoferroautotrophic routes have been reported with Rhodopseudomonas palustris TIE-1. Under these two growth modes, PHA production was more efficient using photoelectroautotrophy but had higher rates with photoferroautotrophy (Ranaivoarisoa et al. 2019).
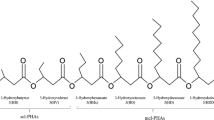
PHA accumulation routes via PNSB are complex and vary based on various parameters including strain of bacteria, carbon substrate and metabolic pathway. Three biosynthetic enzymes are involved in the accumulation of 3HB, the most commonly synthesized and extensively studied monomers. Two acetyl-CoA molecules are merged by 3-ketothiolase (PhaA) enzyme to produce acetoacetyl-CoA. The acetoacetyl-CoA reductase (PhaB) enzyme reduces acetoacetyl-CoA to 3-hydroxybutyryl-CoA. Ultimately, 3-hydroxybutyryl-CoA is polymerized into PHB via PHB synthase (PhaC) enzyme (Verlinden et al. 2007). Yet, the conversion of organic substrates to acetyl-CoA, an important precursor to PHB accumulation, can follow different biochemical pathways. Pure PNSB strains can produce PHA through different pathways under the same carbon substrate. For instance, acetate assimilation for PHA production in Rhodobacter sphaeroides has been reported via the ethylmalonyl-CoA pathway, Rhodobacter capsulatis via the TCA cycle and Rhodopseudomonas palustris via the glyoxylate cycle (Kars and Gündüz 2010). While acetate typically results in the production of 3HB monomers (Özsoy Demiriz et al. 2019), their ratio can be impacted by the acetate assimilation pathways explained in Fig. 1 (Montiel-Corona et al. 2015). The PNSB preference for acetate as a substrate for the production of PHA has been reported repeatedly. A study by Higuchi-Takeuchi et al. (2019) revealed that anaerobic conditions were essential for the high production of PHA in marine PNSB as these conditions enhanced the expression levels of IDH (isocitrate dehydrogenase), which is a key enzyme in the catalyzation of isocitrate to β-ketoglutarate in the TCA cycle. Other PNSB such as Rhodobacter sphaeroides and Rhodospirillum rubrum are incapable of isocitrate lyase, but they still assimilate acetate via anaplerotic pathways (Petushkova et al. 2019). The ethylmalonyl-CoA assimilation pathway is triggered by the presence of malate synthase in bacteria that do not contain isocitrate lyase to produce C5-acids (Erb et al. 2007). Substrates such as lactate and malate can easily be oxidized to pyruvate, which is then transformed to acetyl-CoA. Other substrates need to go through one of the anaplerotic pathways to enter the TCA cycle (Montiel-Corona and Buitrón 2021).

Acetate assimilation pathways from various PNSB
3.2 Ecology
Most PNSB thrive in environments with mesophilic temperatures and neutral pH. However, a number of PNSB genera have the ability to withstand environments with extreme temperatures, varying pH and elevated salinity, provided that the environment is anoxygenic or microaerobic (Tanskul et al. 2016). For instance, PNSB have been isolated from thermal springs and alkaline lakes at temperatures beyond 50 °C (Favinger et al. 1989; Imhoff et al. 2005; Kumar et al. 2013). Rhodopseudomonas palustris, Rhodomicrobium vannielii and Rubrivivax gelatinosus were all recovered from a Russian hot spring with temperatures of 72–74 °C and pH beyond 9 (Namsaraev et al. 2003), although further testing revealed that these conditions were not optimal for growth.
PNSB can inhabit a wide range of salinity environments. Microorganisms such as Roseospira, Rhodovibrio, and Rhodovulum are classified as slightly halophilic PNSB (Imhoff 2017b). Rhodovibrio sodomensis, recovered from the Dead Sea, has shown optimal growth at 12% salinity, but adapted to environments with 6–21% salinity (Mack et al. 1993). Similarly, Rhodothalassium salexigens, reached an optimal growth in an environment with 7% salinity, but withstands environments with 20% salinity (Kanekar et al. 2012; Imhoff 2017b). The photosynthesis ability of PNSB tends to be limited in environments with salinity beyond 32% (García et al. 2019) but the ability to withstand saline conditions provides a promising additional selective pressure for these organisms in mixed culture heterotrophic conditions.
Although it is reported that PNSB thrive in a neutral or slightly acidic environments, some types of PNSB have the ability to grow in strongly alkaline or acidic environments. The Yellowstone National Park encloses warm springs with pH ranging from 3.5 to 4.9, where Rhodopila globiformisi was recovered (Pfennig 1974). Rhodomicrobium vannielii and Rhodoblastus acidophilus were recovered from various hot acidic lakes in the United States and Germany with pH varying between 4.8 and 7.2 (Pfennig 1969). Alkaline soda lakes are also extreme environments with pH ranging from 8.1 to 10.6, where growth of Rhodovulum and Rhodobaca strains was reported (Kompantseva et al. 2010).
4 Influencing Factors of PNSB PHA Storage
4.1 Carbon substrate
PNSB feed on various types of organic acids, fatty acids and sugars (Madigan and Jung 2009). They also consume short-chain alcohols and inorganic carbon sources (Madigan and Jung 2009; Talaiekhozani and Rezania 2017). The consumption of carbon substrate varies according to the PNSB species. For instance, although belonging to same genus, the strain Rhodospirillum rubrum is known for not taking up glycerol and mannitol, yet Rhodospirillum photometricum consumes these substrates (Imhoff et al. 2005). Regardless, it is agreed upon that excess of carbon substrate is a key enabler for PHA accumulation.
Various experiments explored PNSB strains and their preference towards specific carbon compounds for final by-products generation. For instance, the strain Rhodopseudomonas palustris WP3-5 was proven to take up lactate for biohydrogen production rather than for PHA storage (Wu et al. 2012). Further investigations revealed that PNSB consume substrates such as malate, pyruvate and succinate for biohydrogen production, while butyrate and propionate are converted to PHA (Cardeña et al. 2017). Additionally, most PNSB strains were reported to prefer pure acetate as substrate for PHA accumulation due to its easy assimilation into the metabolic pathway for Acetyl-CoA synthesis, which is a precursor to PHA (Brandl et al. 1991; Liebergesell et al. 1991; Khatipov et al. 1998b; Fradinho et al. 2014). Other studies have revealed that PHA accumulation under a pure butyrate substrate was higher than acetate under the same tested conditions (Brandl et al. 1989; Chen et al. 2012; Carlozzi et al. 2019a). The work of Khatipov et al. (1998a) revealed that Rhodobacter sphaeroides accumulated higher PHA under the presence of acetate, yet when subjected to a lactate or pyruvate medium, the strain showed a higher cell growth but slowed its PHA production. This may provide opportunities for dual treatment of different wastewaters in separate growth and accumulation reactors. In the work conducted by Demiriz et al. (2019), 65 mM of acetate was found to give the highest biomass content and PHB storage, but beyond this reduced PHB content was observed, due light limitations caused by higher biomass concentrations. Additionally, when n-alkanoic acids were present in the medium, PHB accumulation was doubled compared to an acetate only medium (Mukhopadhyay et al. 2005). Fradinho et al. (2014, 2019) explored the substrate preference of a mixed APB system and their impact on PHA accumulation. The presence of acetate in propionate and butyrate environments accelerated the assimilation of VFA and the culture showed a preference towards acetate. While acetate, butyrate and propionate were successful in enabling PHA accumulation, environments with malate, citrate and lactate showed no improvement in PHA content (Fradinho et al. 2014). This may be because some of these substrates are shown to be preferential for biohydrogen production, which competes with PHA production for reducing equivalents from photosynthesis (Ghosh et al. 2017). Similar trends were reported by Brandl et al. (1991) where PNSB growth was higher under the presence of acetate compared to malate and crotonate. Yet, mixed VFAs that contained acetate resulted in comparable PHA accumulations to pure acetate tests (Brandl et al. 1991; Fradinho et al. 2014). Finally, organic concentrations play a crucial role in PHA accumulation. Fradinho et al. (2019) observed that APB grown under high acetate content exhibited inhibition compared to APB grown with low initial acetate content. Similarly, Ali Hassan et al. (1996) demonstrated the presence of formic acid (FAc) may cause a reduction in PHA yield and accumulation from 0.5 g g−1 and 67% CDW (0 g-FAc L−1) to 0.21 g g−1 and 18% CDW (3.3 g-FAc L−1) with Rhodobacter sphaeroides (IFO 12203) using synthetic fermented palm oil mill effluent at pH 7.
In general, PNSB utilize various types of carbon substrate for their growth and excess of carbon substrates is a trigger for PHA accumulation. However, the final by-product varies depending on the substrate available. It was observed that VFA are consumed first from the environment by PNSB compared to other carbon substrates. A preference for acetate is recorded across most PNSB strains and is directed towards PHA storage. Similarly, butyrate and propionate are also assimilated for PHA production. However, while organic acids such as lactate and succinate consumed by PNSB, their contribution towards PHA production is low and does not exceed 12% CDW and 8.5% CDW for lactate and succinate, respectively (Khatipov et al. 1998a; Kim et al. 2012). Finally, the presence of precursors such as butyrate and valerate increases the production of diverse PHA (Mukhopadhyay et al. 2005).
While employing pure carbon substrates such as acetate or glucose is a costly approach, wastewater tends to contain a mixture of various organic acids, sugars and occasionally alcohols that PNSB can utilize. It also constitutes a cheap and readily available substrate source. Wastewaters from dairy and cheese production industries (Alloul et al. 2019; Fradinho et al. 2019; Carlozzi et al. 2019b), winery industry (Policastro et al. 2020), food waste (Montiel-Corona et al. 2015; Ghimire et al. 2016; Montiel Corona et al. 2017), and municipal wastewater (Luongo et al. 2017) were all explored for the successful culturing and growth of PNSB for PHA accumulation with values varying between 12 and 83% CDW. A majority of these studies had to go through a dark-fermentation process in order to increase VFA present in the effluent, which increases the costs further, but is also common to non-PNSB fermentations for PHA production. Additional studies are warranted to find sustainable and suitable carbon substrates for PHA production by PNSB without impacting the overall production costs.
4.2 Nutrients
In chemoheterotrophic systems, limiting nutrient availability has frequently been used to halt protein production and cell growth forcing bacteria to store carbon as PHA for later use. Similar limitation of nutrients such as nitrogen or phosphorus is recommended to achieve high PHA accumulation for PNSB. Photoheterotrophic systems require higher N and P ratios than chemoheterotrophic systems as their catabolic energy source is emerging from light, which directs a higher proportion of carbon to anabolism. N and P assimilation is proportionate to the amount of COD, more specifically the amount of VFA consumed (Hülsen et al. 2014), thus ratios of COD:N and COD:P are crucial for PNSB growth and their main activities. However, there is little study on the optimum ratio of COD:N and COD:P among researchers for PNSB growth and PHA accumulation.
Nitrogen limitation and depletion are often linked to increased 3HB accumulation in the system. (Melnicki et al. 2009; Montiel Corona et al. 2017; Luongo et al. 2017; Carlozzi et al. 2019a). Carlozzi et al. (2018) reported an increase in 3HB storage by Rhodopseudomonas to 377 mg-PHA L−1 after 144 h of cultivation under ammonia limitation which amounts to a 32% increase from nitrogen sufficient conditions. Under different settings, the same group identified that ammonia availability was linked to biomass growth due to nitrogenase inhibition, while the presence of sufficient glutamate as nitrogen source enabled PHA accumulation in the environment (Carlozzi et al. 2019a). Ali Hassan et al. (1996) also showed PHA yield on a mixed acetate and propionate source was more than three times higher under ammonia limited conditions. Khatipov et al. (1998a) confirmed the link between increased PHA accumulation and nitrogen deprived conditions with lactate, pyruvate, glucose and succinate. However, both nitrogen availability and depletion did not have any impact on PHA accumulation with acetate as the sole carbon source in their tests, based on similarly high PHA content (Khatipov et al. 1998b). In contrast to the above studies, Vincenzini et al. (1997) investigated the COD:N limits to achieve a simultaneous PNSB growth and PHA accumulation in a system. The experiment revealed that Rhodopseudomonas palustris cultured in a malate environment with an initial COD:N of 100:6 accumulated 25 mg-PHB L−1 d−1, while a slightly lower COD:N of 100:12 resulted in higher PHB accumulation rates overall in the system at 40 mg-PHB L−1 d−1. While nitrogen limitation did enhance PHB accumulation, the overall biomass productivity of PHB decreased only slightly from 0.0657 g-PHA.g-X−1 to 0.0558 g-PHA.g-X−1 under nitrogen limitation. Such apparent discrepancies may well be due to the competition between PHA production and biohydrogen production, which is also promoted under nitrogen limited conditions which initiate biohydrogen production via nitrogenase (Melnicki et al. 2009; Ghimire et al. 2016; Ghosh et al. 2017; Montiel Corona et al. 2017; Luongo et al. 2017; Carlozzi et al. 2019a). It is therefore critical to apply other conditions that will prevent biohydrogen production if nitrogen limitation will be used to enhance PHA production such as increasing pH (Kim et al. 2012) or foregoing pH control (Kim et al. 2011), using acetate as the sole substrate source (Hustede et al. 1993; Khatipov et al. 1998a), and increasing the frequency of light-dark cycling (Montiel Corona et al. 2017).
Limitation of other nutrients such as phosphorous and sulfur could also lead to PHA accumulation in PNSB (Vincenzini et al. 1997; Melnicki et al. 2009; Mukhopadhyay et al. 2013). Since biohydrogen and PHA production from PNSB are competitive processes, the work conducted by Carlozzi et al. (2019a) and Vincenzini et al. (1997) provides an indication about the role of P and S nutrient limitations on the production of biohydrogen. Vincenzini et al. (1997) observed that phosphate depletion conditions resulted in an increase of PHA accumulation in Rhodopseudomonas palustris and halted the production of biohydrogen. In a study with Rhodospirillum rubrum, sulfur deprivation not only halted biohydrogen production but resulted in rapid storage of PHB to more than 50% CDW (Melnicki et al. 2009). Many have labeled sulfur as the switch between PHA accumulation and biohydrogen production in PNSB (Melnicki et al. 2009; Chen et al. 2017). However, there are contradicting reports about the role of P limitation on PHA storage. While Fradinho et al. (2016) concluded that P availability and limitation had very little to no effect on PHA production on a mixed PNSB system, Carlozzi et al. (2019a) observed high PHA storage under P and S limitations with Rhodopseudomonas in a period of 96 h. Various research groups also confirmed that P limitations resulted in high PHA content in pure PNSB strains (Brandl et al. 1989; Mukhopadhyay et al. 2005). Thus, the difference in the reported results is likely due to the pure and mixed cultures utilized. Finally, Higuchi-Takeuchi et al. (2019) revealed that nutrient limitation does not improve PHA accumulation in marine PNSB, possibly due to their natural propensity for growth in low-nutrient conditions.
The literature body therefore confirms the involvement of nutrients such as ammonia, phosphorous and sulfur, in driving PHA accumulation. For nitrogen, the source and concentration are important as well as the carbon sources linked with it to achieve a high PHA storage (Khatipov et al. 1998a). Sulfur-free environments were also linked to high PHA content (Mukhopadhyay et al. 2005). However, further research may be required to confirm the role of phosphorous limitation on PHA, where differences in literature may be associated with different PNSB strains used.
4.3 pH
The environment’s pH value influences the cells’ physiological activities and PHA accumulation (Hustede et al. 1993). An environment with controlled pH results in higher PHA production with reports revealing that optimal microbial growth and PHA synthesis is observed in neutral to slightly alkaline conditions (Amulya et al. 2016; Cheah et al. 2019). It was reported that a controlled pH ranging from 5 to 8 does not affect the long-term PNSB growth, but an initial pH of 4 is not suitable for growth (Wang et al. 2016). A pH optimization study of Rhodocyclus gelatinous revealed that the optimal growth conditions were at pH 7 (Prasertsan et al. 1993). Other types of PNSB were also reported to grow optimally at pH 7 (Kapdan et al. 2009; Pattanamanee et al. 2012; Kars and Ceylan 2013; Zagrodnik and Laniecki 2015; Ghosh et al. 2017; Laurinavichene et al. 2018). Certain strains of Rhodobaca genus, on the other hand, have shown optimal growth at pH 9 (Imhoff et al. 2005).
There are reports that slightly alkaline environments enhance PHA production due to inhibited biohydrogen production (Cardeña et al. 2017). Khatipov et al. (1998b) observed that Rhodobacter sphaeroides were able to reach a PHA content of 40% CDW at a pH of 10 with pyruvate, shifting from biohydrogen production at more neutral pH to PHA production at more alkali pH. Similar observations were made by Hustede et al. (1993) who noticed decreased biohydrogen production and increased PHA production in poorly buffered solutions. Suzuki et al. (1995) also found that a pH of 8.0-8.5 results in higher PHA accumulation in the same strain. Brandl et al. (1989) did not directly assess pH, but the highest PHA accumulation observed in their study was consistent with a high final pH during the experiments. Montiel-Corona et al. (2015) found complimentary findings when investigating headspace flushing, where PHB production was highest for argon gas flushing, followed by a low vacuum treatment and lastly carbon dioxide flushing. The pH resulting from these conditions followed a similar trend of high to low. While these results might suggest that a neutral pH is more suitable for PNSB growth and a slightly alkaline pH is beneficial for PHA growth, a detailed study under controlled pH conditions and an optimal pH range is yet to be determined.
4.4 Light
4.4.1 Light intensity and penetration
Light is the energy driving force for cellular reactions in PNSB, but can also be inhibitory if at too greater intensity. Zhou et al. (2014) found a light intensity of 2000 lx (16 W m−2) to be optimal for organic removal and PNSB growth while lighting intensity beyond 5000 lx (40 W m−2) resulted in decreased COD removal and biomass growth rates. The group also reported that the lower (optimal) light intensity resulted in the lowest production of BChl and carotenoid pigments. Fradinho et al. confirmed the link between high light intensity and PHA accumulation in APB, where specific light intensity per unit biomass was the critical parameter. At values of less than 2 W g−1, very low PHA accumulation was observed (around 3–5% CDW). During a period of biomass washout, light intensity increased to 11.4 W g−1 and coincided with an increase in PHA to 70% CDW. In a follow up set of tests, a light intensity of 1.5–3.5 W g−1 gave a final PHA content of around 30%, while a light intensity of 5.6–6.7 W g−1 gave a PHA content of 60%. However, light intensity did not seem to impact the specific carbon removal rate (Fradinho et al. 2016). Similar results were found by Montiel-Corona et al. (2015), who observed a production of 0.19 g g−1 in outdoor cultures (natural light-dark cycle) with intensity of approximately 60 kLux, compared with 0.07 g g−1 under indoor continuous light at 3 kLux.
Light penetration controls the amount of light reaching the biomass. Higher cell growth reduces the light intensity reaching the cells and the overall productivity (Carlozzi et al. 2018). Fradinho et al. (2016) demonstrated that as cell growth increases, the light penetration decreases, which results in lower PHA accumulation, and is an important consideration for bioreactor design in an accumulation phase, given that in contrast, high cell densities are desired for subsequent cell processing. In fact, the group reported the highest biomass yield of 0.11 Cmol X Cmol−1 acetate when moderate illumination of 127 W m−2 was provided as well as 35% CDW. However, the cell yield decreased significantly to 0.11 Cmol X Cmol−1 acetate under higher light presence of 227 W m−2, but resulted in enhanced PHA accumulation reaching 60% CDW PHA, one of the highest accumulations reported so far. Under very high light presence or insufficient light presence, however, the photosynthesis is interrupted, halting the activities of PNSB (Serôdio et al. 2008). The optimal light intensity value that results in high PHA accumulation as well as biomass yield is yet to be identified.
4.4.2 Light cycling and frequency
Sunlight availability and intensity vary widely from laboratory conditions, which represents a challenge for scale up. Some light conditions can lead to photo-inhibition, while others promote other cellular activities than PHA accumulation. In one experiment using a Rhodopseudomonas enriched inoculum at Renmin University of China the highest APB biomass growth was achieved under a 24 h/24 h dark/light cycle compared to shorter dark/light tested cycles of equal length, while the highest cell protein content was achieved at short 3 h/3 h cycles (Zhi et al. 2019). In an earlier study by the same group, varying light dark ratios of 2.5 h/0.5 h through to 1 h/2 h, and a constant light control were assessed (Zhou et al. 2015b). It was observed that the highest biomass growth and COD removal were achieved at a light/dark cycle of 2 h/1 h for a Rhodopseudomonas strain. The results of these studies are in contrast and may be due to a different dominant strain. In accordance with the previous study, Montiel Corona et al. (2017) found shorter dark/light cycles of 30 min/30min were more beneficial for PHA production, although the shortest photoperiod of 15 min/15min provided slightly lower production for Rhodobacter capsulatis. Biomass growth was not greatly influenced by photoperiod in this study, with the unknown Rhodopseduomonas strain growing slightly more with constant illumination and Rhodobacter capsulitis at 15 min/15 min. Fradinho et al. observed that under a 4 h/4 h light/dark cycle, APB doubled their net PHA accumulation rate and increased their overall PHA accumulation to 30% PHA/VSS compared to continuous light availability Fradinho et al. 2013a, b). However, in this study the higher PHA accumulation was due to the selection of APB over microalgae under the light cycling, as later studies with continuous feed and lighting achieved higher accumulation and net production rates (Fradinho et al. 2016).
4.4.3 Light wavelength
While natural light is the main target to reduce overall PNSB production costs, it is important to understand PNSB’s reactions to the varied synthetic wavelengths that could be used to assist production, especially since light source and wavelength are known to affect the uptake of carbon substrate from the environment (Zhou et al. 2015a). Uyar et al. (2007) observed a slower PNSB growth when a wider range of wavelengths was available. However, an enhanced growth was detected when only IR wavelengths were supplied. The study of Zhou et al. (2015a) is an outlier, indicating that red LED light (650 nm) resulted in the highest biomass growth and COD removal compared to blue, yellow and white LED lights under the same conditions. However, the group did not test infrared lights due to the high associated costs. While pigment production was higher with a yellow light source, the highest carotenoid to BChl ratio was observed under red light. Qi et al. (2017) confirmed that changes in light wavelength affected COD and nutrients removal on Rhodopseudomonas sphaeroides using monochromatic LEDs of 750, 850 and 940 nm wavelengths at various intensities in a Box-Behnken experimental design that also included intensity and photoperiod. The concluded 850 nm was optimal for COD and ammonia removal, as well as biomass growth, while changes in light intensity and light exposure duration had the least impact on removal of COD and nutrients.
While the light intensity was different for each of these studies, varying between 500 and 4000 lx, the outcome was similar. The 850 nm wavelength and/or presence of IR was more beneficial for the growth of PNSB. Additionally, the main concern lays in the maximum absorption capacity of BChl a, which was previously reported between 771 and 801 nm (Ohashi et al. 2010; Olson 2013), Further investigations have proven that it is still possible to obtain good BChl a absorption results at 850 nm wavelength alone for PNSB (Qi et al. 2017). In fact, it was reported that the presence of IR along with the absence of UV wavelengths results in the selective growth of PNSB even under non-sterile conditions (Capson-Tojo et al. 2020). Therefore, the use of monochromatic IR also provides a suitable low-energy method to reduce microalgae competition.
4.5 Fermentation system and feeding strategy
Various fermentation systems are used for PNSB culturing. The one stage process is a single fermenter where PNSB culturing and PHA production take place simultaneously. Frequently, a combination of carbon excess, for biomass growth, and nutrient limitation, for PHA accumulation, is employed in a single stage system (Hustede et al. 1993; Chen et al. 2012; Özsoy Demiriz et al. 2019). Recently, high substrate availability was coupled with high light intensity in a single system and resulted in the accumulation of 60% PHA (Fradinho et al. 2016). The two-stage fermentation consists of two consecutive stages operating under different conditions. The most popular two-stage setup is an initial growth stage with nutrient availability for PHA-producing cells selection, followed by a second stage that consists of excess of carbon and nutrient limitation where PHA accumulation takes place (Pagliano et al. 2017). Two-stage systems were successfully employed at lab-scale for PHA production either with a first stage that consists of sufficient nutrients to increase biomass followed by a second stage with nutrient limitation to trigger PHA accumulation (Mukhopadhyay et al. 2005; Corneli et al. 2016) as described above; a biphasic growth with a different medium at each stage (Mukhopadhyay et al. 2005, 2013); or a fermentation first stage and a simultaneous growth and accumulation second stage (Ali Hassan et al. 1997). In the latter system dark fermentation using acidogenic fermentative bacteria is often used in the initial stage rather than PNSB (Ghimire et al. 2015, 2016; Carlozzi et al. 2019b).
Three-stage fermentation is a common process for mixed cultures, where the first stage targets the production of organic acids and volatile fatty acids (VFA) via fermentative bacteria, followed by a second stage of PNSB enrichment where PHA-producing biomass is promoted, and a final stage that consists of PHA accumulation under high organic substances (Ghimire et al. 2015, 2016). Recently, research is shifting towards the development of a three-stage fermentation process operated with PNSB in all stages. Carlozzi et al. (2019a) developed a three-stage system where the first stage consisted of nutrient sufficient conditions, a second stage of nitrogen limitation and final stage of sulfur limitation. While the N-sufficient stage followed by a double nutrient N and S deficient stage did result in a moderate PHA accumulation of 25.9% CDW, the final PHA accumulation of the three-stage process reached 34.4% CDW. This is explained by the effect of S-depletion on the system, which hindered the nitrogenase activities leading to a halt in production of biohydrogen and stopped biomass growth due to the inactivity of de novo synthesis of amino acids. The main purpose of employing a three-stage system was the separation of the competing processes of biohydrogen production and PHA accumulation. Fradinho et al. (2019) explored a three-stage process with an initial stage of low substrate concentration to avoid growth inhibition, a second stage of moderately high light intensity in the range of 10.6 W L−1, and a final stage of N residual removal from the media to halt biomass growth. The first and second stages have consistently increased PHB production in the system, with the third stage having a negative effect on PHB accumulation that caused a decreased from 30 to 20% CDW. However, this research indicates the first two stages could be used as a second and third stage for growth and accumulation and be preceded by a dark fermentation stage to prepare the optimum substrate. In this process high light intensity is the primary driver for PHA accumulation in the final stage. It is therefore evident that three-stage systems can provide optimized systems but that their full potential and various configuration possibilities remain relatively unexplored. Figure 2 summarizes the fermentation setups that are used for PHA accumulation.
Each fermentation system provides benefits and disadvantages, with simpler systems providing lower capital costs and less complexity, but reduced overall productivity and process control. However, given PNSB’s desirable ability to both grow and accumulate PHA simultaneously, they provide the opportunity to rely on single or two-stage systems, where in both cases one reactor is a combined growth-accumulation reactor. Nonetheless, a separate growth and accumulation phase may still be warranted due to the high light requirements to maximize accumulation (Fradinho et al. 2019). So far, outdoor PHA production systems are still rare and need to be investigated further. However, a successful one-stage outdoor prototype has been investigated using high initial substrate and no nutrient limitation (Carlozzi et al. 2018). Nonetheless, the highest PHB accumulation of 377 mg-PHB L−1 was achieved under N-depleted conditions, confirming once again the involvement of nutrient limitation in PHA production.

Fermentation systems for PHA accumulation
While the two-stage system is preferred in lab-scale setups, the bioreactor operation also plays a role in the selection of PHA-producing biomass and their ability to accumulate PHA. Continuous, sequencing-batch reactor (SBR) and fed-batch reactor systems have all been employed for PNSB enrichment as they control the feeding strategy of the system. Continuous processes allow an exponential growth of bacteria through continuous addition of substrate and nutrients without interrupting the fermentation process and providing a continuous supply of carbon substrate. While this process is the most desirable in an industrial setup, most lab-scale research employs a semi-continuous bioprocess by replenishing the substrate and nutrients before they are depleted to simulate a continuous system. In a two-stage system, the SBR is often employed for the culture selection and enrichment step, while a fed-batch process is more suitable for PHA accumulation (Albuquerque et al. 2010a).
Carlozzi et al. (2018) compared the semi-continuous and fed-batch processes with Rhodopseudomonas species. The experiment revealed that the fed-batch process was more effective at accumulating higher PHB content of 377 mg-PHB L−1 under a shorter period of time of 144 h as opposed to the semi-continuous process that accumulated 255 mg-PHB L−1 in 336 h. For PHA accumulation in chemoheterotrophic (non-PNSB) systems, transient feeding is typically used to provide unbalanced conditions that promote VFA assimilation (Basak et al. 2011; Basset et al. 2016). Fradinho et al. (2013a) explored various feeding strategies coupled with various light conditions and their impacts on APB. Under a feast-famine regime and continuous light, PHA accumulation reached 20% CDW in MMC containing both APB and algae. The group, however, observed that coupling feeding with the beginning of the dark phase resulted in the out-competition of algae and, surprisingly, increased the assimilation of acetate as well as PHA accumulation, which reached 30% CDW (Fradinho et al. 2013b). Later experiments investigated the impacts of a semi-continuous feeding regime and continuous light, achieving an increase in PHA accumulation in APB, reaching a value of 60% CDW (Fradinho et al. 2016). Under this process, it was hypothesized that high PHA accumulation was due the combined enrichment of the APB over algae and their ability to store PHA concomitantly with growth under the continuous feeding. These results contrast Carlozzi and may be due to additional effects from lighting or due to a possible difference in species present in the MMC system.
Bioreactor designs could also affect the final output of PHA as light distribution and penetration can be key factors for the cultivation system. A large surface area or surface-to-volume ratio is often preferred in the case of PNSB as it allows higher light distribution into the media. One method to control this factor is by changing the geometry of the bioreactor. The most studied shapes include stirred tank, flat-panel and tubular bioreactors (Adessi and De Philippis 2014). Low surface-to-volume ratio is obtained by a stirred tank bioreactor, which can affect the light exposure. However, it provides easy scale-up and better biomass mixing to ensure cells receive uniform exposure to light (Cerruti et al. 2020). On the other hand, a flat-sheet reactor offers a high surface-to-volume ratio and light exposure as it has a small thickness (Akkerman et al. 2002). This provides a challenge for scaling-up due to capital cost and potential mixing challenges or energy requirements. Tubular systems are tubes that be arranged under different orientations such as vertical, horizontal, or serpentine. They provide high light penetration and avoid short-circuiting risks of flat-panel reactors, but have high energy requirements for pumping. They are frequently reported at pilot-scale in outdoor environments (Gebicki et al. 2009; Boran et al. 2010; Carlozzi et al. 2018). Available information on photobioreactor design for phototrophic microorganisms mainly focuses on microalgae, while those reported for PNSB are mainly for biohydrogen production. Reports on PNSB photobioreactors designed specifically for PHA production are scant and require further research to select the best bioreactor design for high yield and productivity. Nevertheless, due to a large similarity between the competing PHA and biohydrogen production processes by PNSB, bioreactor designs for biohydrogen provide key information about the best conditions for biomass growth and can be found in the in-depth review prepared by Adessi and De Philippis (2014).
4.6 PHA monomer
Strain purity and substrate consistency are considered the major causes for the consistency of PHA monomer. It is well established that carbon substrates influence the type of monomers produced (Bugnicourt et al. 2014). Many experiments showed that when using acetate as the sole carbon source, it results in a majority of 3HB monomers (Fradinho et al. 2013a). Pure butyrate or a mixture of butyrate and acetate were reported to produce a large proportion of 3HB regardless of the culture (Carlozzi et al. 2019a). Lactate is also a precursor to PHB polymer production, but it tends to result in lower accumulation than values reported with acetate and butyrate (Carlozzi et al. 2019a). On the other hand, valerate promotes the production of PHV in the environment. The addition of valerate in any environment increases the accumulation of PHV and lowers that of PHB (Sheu et al. 2009). While there are various reports on the effects of different alkyl groups on the production of MCL-PHA in non-phototrophic cultures (Tan et al. 2014), there is only one report on the ability of genetically-modified Rhodospirillum rubrum to produce MCL-PHA (Heinrich et al. 2016).
Higuchi-Takeuchi et al. (2016b) investigated the impact of acetate and/or sodium bicarbonate medium on the production of 3HB and 3HV monomers in various pure strains of PNSB. While the medium concentration and growth conditions were identical, it was revealed that the strain is also responsible for the production of different PHA monomers. Strains such as Rhodovulum sulfidophilum and Rhodovulum imhoffii resulted in higher concentration of 3HB monomers, 94.9:5.1 and 100:0 HB:HV, respectively. Yet under the same conditions, other strains such as Rhodovulum tesquicola and Roseospira marina, resulted in higher 3HV content accumulating 63.3:36.7 and 49.9:50.1 HB:HV, respectively.
A pure bacterial strain is more likely to produce PHA with stable monomers, whereas MMCs are known to produce various types of PHA monomers and chain lengths (Sharma et al. 2017). In an experiment where various types of PHA producing (non-APB) bacteria were tested under the same conditions, it was revealed that both strain of bacteria and carbon substrate played a significant role in the production of SCL-PHA and MCL-PHA. For instance, Pseudomonas putida fed with different substrates always resulted in MCL-PHA with the exception of pentanoic acid that resulted in about 35% SCL-PHA. On the other hand, Bacillus megaterium resulted in far more SCL-PHA monomers when fed with the same substrates with the exception of octanoic acid that resulted in only MCL-PHA (Shahid et al. 2013).
While the bacterial strain purity and carbon substrate are important in the PHA monomer production, other factors are also involved in the process. Chen et al. (2015) observed that pH is involved in the monomer PHA composition, while Albuquerque et al. (2010b) confirmed that feeding regimes manipulate types of monomer accumulated. Hence, controlling PHA monomer stability is a complex process that involves various factors. Nevertheless, monomer stability in PNSB systems is yet to be investigated and most studies reporting PHA in PNSB are related with biohydrogen production, where frequently only PHB is quantified which could also give misleading results on PHA accumulation.
4.7 Productivity rates
PHA productivity rates of PNSB are fairly low compared to the reported values of aerobic bacteria. For instance, the highest reported microbial PHA productivity is with Alcaligenes latus which achieved 5.13 g-PHA L−1 h−1 (Wang and Lee 1997). However, the majority of aerobic prokaryotes were reported to synthesize between 1 and 2 g-PHA L−1 h−1 (Blunt et al. 2018). Meanwhile, the average reported values in PNSB are in the range of 0.005–0.059 g-PHA L−1 h−1 for pure strains (Ali Hassan et al. 1997; Cardeña et al. 2017; Carlozzi et al. 2019a; Fradinho et al. 2019) which is 2–3 orders of magnitude lower compared to aerobic bacteria productivity. Rhodobacter sphaeroides was reported to have the highest single culture productivity of PNSB at 0.056–0.059 g-PHA L−1 h−1 (Ali Hassan et al. 1997; Khatipov et al. 1998a, b) though reported productivities were only found for this species and Rhodopseudomonas. While pure strains are generally known to produce PHA at higher rates, recently a phototrophic MMC has achieved a PHA production rate at 0.111 g-PHA L−1 h−1 in the accumulation phase (Almeida et al. 2021), which is just over 1 order of magnitude lower than chemoheterotrophic systems by comparison. The same group achieved 0.091 g-PHA L−1 h−1 in an earlier study with mixed PNSB (Fradinho et al. 2019). These differences in productivity are a significant challenge for PNSB based PHA production but should not be seen as a critical hurdle, given the extensive investment and research in chemoheterotrophic systems compared to PNSB systems. Moreover, PNSB provide opportunities for recovery of various other biomolecules that can add significant value to the overall process.
5 Summary and Future challenges
Producing PHA from pure strains has been widely studied and has reached industrial scale, yet its commercial growth has been limited due to high production costs. PNSB have been proposed as an alternative as they provide potential environmental and energy-saving advantages. Their growth does not require aeration as it necessitates an anaerobic environment. PNSB feed on a wide range of organic compounds and can accumulate PHA under both nutrient sufficient and deficient conditions. Furthermore, PNSB extract their catabolic energy from light sources such as sunlight resulting in high carbon to PHA yields and can be easily enriched in a mixed culture, which eliminates the frequent sterilization requirement and the costs associated with keeping a pure culture.
For PNSB to compete economically, the design of an optimized system is crucial that will select for optimal strains and promote high growth and accumulation. However, research to date has been limited, focusing primarily on a few organisms, primarily belonging to the genera Rhodopseudomonas and Rhodobacter, and often as a by-product of hydrogen production. Further investigation of other PNSB genera, as well as mixed culture behavior, for PHA production under various environmental controls is warranted. Light is one of the key design parameters in a photoheterotrophic PHA production system. Light/dark cycling and sunlight intensity vary significantly based on the location, season and time of the day, which affect growth rates, pigment expression and PHA accumulation. Their further investigation, particularly with respect to culture densities and light transmittance will be critical for efficient commercial scale systems. Additionally, continuous or external light sources provide an opportunity to increase overall PHA accumulation in PNSB, but require additional energy and cost. Variations exist currently in results from light spectra and cycling studies on PNSB growth and are currently absent for PHA production. This information is necessary to adequately assess the economics of providing auxiliary lighting, since Capson-Tojo et al. (2020) have shown it may cost as much as 1.9 $ kg-biomass−1 if LED lighting is used. Other incandescent light options such as tungsten lamps are cheaper, but less efficient in terms of both PNSB biomass yield and electrical efficiency.
Other major costs for PHA production include substrate, biomass harvesting and PHA extraction costs. Evaluation of various industrial wastes and wastewater sources with suitable clarity and high organic load are areas of potential research, as well as the appropriate pre-treatment fermentation steps to maximize productivity. The harvesting of low cell densities associated with phototrophic systems is still a challenge, as evident from the algal biofuel industry, which raises the question whether new separation techniques need to be developed or whether there should be an increased focus on biofilm-based systems (Podola et al. 2017). The subsequent PHA extraction process is a costly step that needs to be optimized to reach high recovery values at low costs, particularly for MMC where substrate and disinfection are no longer significant costs. It is unclear whether PHA extraction conditions optimal for chemoheterotrophic organisms will also result in the best extraction from PNSB, given the different cellular composition and potentially different PHA polymers produced. Finally, there is lack of published data on outdoor bioreactors systems for PHA production from PNSB. Therefore, there is a need to design an optimal bioreactor for large scale PHA production from MMC PNSB with optimized substrate and nutrients, pH, temperature, and light factors.
Despite these challenges PNSB’s straightforward enrichment, avoidance of aeration, large pool of carbon substrates and concomitant growth and PHA accumulation make PNSB a viable alternative worthy of further research to develop bioprocesses suitable for sustainable PHA bioplastics production. The findings presented here are a practical reference point to develop a PHA production system coupled with wastewater treatment that could potentially put PNSB at a competitive advantage and help develop a large-scale production system.
References
Adessi A, De Philippis R (2014) Photobioreactor design and illumination systems for H2 production with anoxygenic photosynthetic bacteria: a review. Int J Hydrog Energy 39:3127–3141. https://doi.org/10.1016/j.ijhydene.2013.12.084
Ahn WS, Park SJ, Lee SY (2000) Production of poly(3-hydroxybutyrate) by fed-batch culture of recombinant Escherichia coli with a highly concentrated whey solution. Appl Environ Microbiol 66:3624–3627. https://doi.org/10.1128/AEM.66.8.3624-3627.2000
Akiyama M, Taima Y, Doi Y (1992) Production of poly(3-hydroxyalkanoates) by a bacterium of the genus Alcaligenes utilizing long-chain fatty acids. Appl Microbiol Biotechnol 37:698–701. https://doi.org/10.1007/BF00174830
Akkerman I, Janssen M, Rocha J, Wijffels RH (2002) Photobiological hydrogen production: photochemical efficiency and bioreactor design. BIOHYDROGEN 2002 27:1195–1208. https://doi.org/10.1016/S0360-3199(02)00071-X
Albuquerque MGE, Concas S, Bengtsson S, Reis MAM (2010) Mixed culture polyhydroxyalkanoates production from sugar molasses: the use of a 2-stage CSTR system for culture selection. Bioresour Technol 101:7112–7122. https://doi.org/10.1016/j.biortech.2010.04.019
Albuquerque MGE, Torres CAV, Reis MAM (2010) Polyhydroxyalkanoate (PHA) production by a mixed microbial culture using sugar molasses: effect of the influent substrate concentration on culture selection. Water Res 44:3419–3433. https://doi.org/10.1016/j.watres.2010.03.021
Ali Hassan M, Shirai Y, Kusubayashi N et al (1996) Effect of organic acid profiles during anaerobic treatment of palm oil mill effluent on the production of polyhydroxyalkanoates by Rhodobacter sphaeroides. J Ferment Bioeng 82:151–156. https://doi.org/10.1016/0922-338X(96)85038-0
Ali Hassan M, Shirai Y, Kusubayashi N et al (1997) The production of polyhydroxyalkanoate from anaerobically treated palm oil mill effluent by Rhodobacter sphaeroides. J Ferment Bioeng 83:485–488. https://doi.org/10.1016/S0922-338X(97)83007-3
Allegue LD, Puyol D, Melero JA (2020) Food waste valorization by purple phototrophic bacteria and anaerobic digestion after thermal hydrolysis. Biomass Bioenergy 142:105803. https://doi.org/10.1016/j.biombioe.2020.105803
Alloul A, Wuyts S, Lebeer S, Vlaeminck SE (2019) Volatile fatty acids impacting phototrophic growth kinetics of purple bacteria: Paving the way for protein production on fermented wastewater. Water Res 152:138–147. https://doi.org/10.1016/j.watres.2018.12.025
Almeida JR, Serrano E, Fernandez M et al (2021) Polyhydroxyalkanoates production from fermented domestic wastewater using phototrophic mixed cultures. Water Res 197:117101. https://doi.org/10.1016/j.watres.2021.117101
Amulya K, Reddy MV, Rohit MV, Mohan SV (2016) Wastewater as renewable feedstock for bioplastics production: understanding the role of reactor microenvironment and system pH. J Clean Prod 112:4618–4627. https://doi.org/10.1016/j.jclepro.2015.08.009
Anis SNS, Md Iqbal N, Kumar S, Amirul A-A (2013) Effect of different recovery strategies of P(3HB-co-3HHx) copolymer from Cupriavidus necator recombinant harboring the PHA synthase of Chromobacterium sp. USM2. Sep Purif Technol 102:111–117. https://doi.org/10.1016/j.seppur.2012.09.036
Basak B, Ince O, Artan N et al (2011) Effect of nitrogen limitation on enrichment of activated sludge for PHA production. Bioprocess Biosyst Eng 34:1007–1016. https://doi.org/10.1007/s00449-011-0551-x
Basset N, Katsou E, Frison N et al (2016) Integrating the selection of PHA storing biomass and nitrogen removal via nitrite in the main wastewater treatment line. Bioresour Technol 200:820–829. https://doi.org/10.1016/j.biortech.2015.10.063
Bertling K, Hurse TJ, Kappler U, Rakić AD (2006) Lasers—an effective artificial source of radiation for the cultivation of anoxygenic photosynthetic bacteria. Biotechnol Bioeng 94:337–345. https://doi.org/10.1002/bit.20881
Blunt W, Levin DB, Cicek N (2018) Bioreactor operating strategies for improved polyhydroxyalkanoate (PHA) productivity. Polymers 10:1197. https://doi.org/10.3390/polym10111197
Boran E, Özgür E, van der Burg J et al (2010) Biological hydrogen production by Rhodobacter capsulatus in solar tubular photo bioreactor. J Clean Prod 18:S29–S35. https://doi.org/10.1016/j.jclepro.2010.03.018
Brandl H, Gross RA, Lenz RW, Fuller RC (1988) Pseudomonas oleovorans as a source of poly(beta-Hydroxyalkanoates) for potential applications as biodegradable polyesters. Appl Environ Microbiol 54:1977–1982. https://doi.org/10.1128/aem.54.8.1977-1982.1988
Brandl H, Knee EJ, Fuller RC et al (1989) Ability of the phototrophic bacterium Rhodospirillum rubrum to produce various poly (β-hydroxyalkanoates): potential sources for biodegradable polyesters. Int J Biol Macromol 11:49–55. https://doi.org/10.1016/0141-8130(89)90040-8
Brandl H, Gross RA, Lenz RW, Fuller RC (1990) Plastics from bacteria and for bacteria: poly(β-hydroxyalkanoates) as natural, biocompatible, and biodegradable polyesters. Adv Biochem Eng Biotechnol 41:77–93. https://doi.org/10.1007/BFb0010232
Brandl H, Gross RA, Lenz RW et al (1991) The accumulation of poly(3-hydroxyalkanoates) in Rhodobacter sphaeroides. Arch Microbiol 155:337–340. https://doi.org/10.1007/BF00243452
Bugnicourt E, Cinelli P, Lazzeri A, Alvarez V (2014) Polyhydroxyalkanoate (PHA): review of synthesis, characteristics, processing and potential applications in packaging. Express Polym Lett 8:791–808. https://doi.org/10.3144/expresspolymlett.2014.82
Capson-Tojo G, Batstone DJ, Grassino M et al (2020) Purple phototrophic bacteria for resource recovery: challenges and opportunities. Biotechnol Adv 43:107567. https://doi.org/10.1016/j.biotechadv.2020.107567
Cardeña R, Valdez-Vazquez I, Buitrón G (2017) Effect of volatile fatty acids mixtures on the simultaneous photofermentative production of hydrogen and polyhydroxybutyrate. Bioprocess Biosyst Eng 40:231–239. https://doi.org/10.1007/s00449-016-1691-9
Carlozzi P, Sacchi A (2001) Biomass production and studies on Rhodopseudomonas palustris grown in an outdoor, temperature controlled, underwater tubular photobioreactor. J Biotechnol 88:239–249. https://doi.org/10.1016/S0168-1656(01)00280-2
Carlozzi P, Lambardi M (2009) Fed-batch operation for bio-H2 production by Rhodopseudomonas palustris (strain 42OL). Renew Energy 34:2577–2584. https://doi.org/10.1016/j.renene.2009.04.016
Carlozzi P, Seggiani M, Cinelli P et al (2018) Photofermentative poly-3-hydroxybutyrate production by Rhodopseudomonas sp. S16-VOGS3 in a novel outdoor 70-L photobioreactor. Sustainability 10:3133. https://doi.org/10.3390/su10093133
Carlozzi P, Giovannelli A, Traversi ML et al (2019a) Poly-3-hydroxybutyrate and H2 production by Rhodopseudomonas sp. S16-VOGS3 grown in a new generation photobioreactor under single or combined nutrient deficiency. Int J Biol Macromol 135:821–828. https://doi.org/10.1016/j.ijbiomac.2019.05.220
Carlozzi P, Touloupakis E, Di Lorenzo T et al (2019b) Whey and molasses as inexpensive raw materials for parallel production of biohydrogen and polyesters via a two-stage bioprocess: new routes towards a circular bioeconomy. J Biotechnol 303:37–45. https://doi.org/10.1016/j.jbiotec.2019.07.008
Castilho LR, Mitchell DA, Freire DMG (2009) Production of polyhydroxyalkanoates (PHAs) from waste materials and by-products by submerged and solid-state fermentation. Bioresour Technol 100:5996–6009. https://doi.org/10.1016/j.biortech.2009.03.088
Cerruti M, Stevens B, Ebrahimi S et al (2020) Enrichment and aggregation of purple non-sulfur bacteria in a mixed-culture sequencing-batch photobioreactor for biological nutrient removal from wastewater. Front Bioeng Biotechnol 8:1432. https://doi.org/10.3389/fbioe.2020.557234
Chayabutra C, Ju LK (2001) Polyhydroxyalkanoic acids and rhamnolipids are synthesized sequentially in hexadecane fermentation by Pseudomonas aeruginosa ATCC 10145. Biotechnol Prog 17:419–423. https://doi.org/10.1021/bp010036a
Cheah Y-K, Vidal-Antich C, Dosta J, Mata-Álvarez J (2019) Volatile fatty acid production from mesophilic acidogenic fermentation of organic fraction of municipal solid waste and food waste under acidic and alkaline pH. Environ Sci Pollut Res 26:35509–35522. https://doi.org/10.1007/s11356-019-05394-6
Chee J, Yoga S, Lau N et al (2010) Bacterially produced polyhydroxyalkanoates (PHA): Converting renewable resources into bioplastics. In: Mendez-Vilas A (ed) Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology. Formatex Research Center, Spain, pp 1395–1404
Chen Y-T, Wu S-C, Lee C-M (2012) Relationship between cell growth, hydrogen production and poly-β-hydroxybutyrate (PHB) accumulation by Rhodopseudomonas palustris WP3-5. ICCE-2011 37:13887–13894. https://doi.org/10.1016/j.ijhydene.2012.06.024
Chen Z, Huang L, Wen Q, Guo Z (2015) Efficient polyhydroxyalkanoate (PHA) accumulation by a new continuous feeding mode in three-stage mixed microbial culture (MMC) PHA production process. J Biotechnol 209:68–75. https://doi.org/10.1016/j.jbiotec.2015.06.382
Chen Y, Jiang X, Xiao K et al (2017) Enhanced volatile fatty acids (VFAs) production in a thermophilic fermenter with stepwise pH increase – investigation on dissolved organic matter transformation and microbial community shift. Water Res 112:261–268. https://doi.org/10.1016/j.watres.2017.01.067
Choi J, Lee SY (1999) Factors affecting the economics of polyhydroxyalkanoate production by bacterial fermentation. Appl Microbiol Biotechnol 51:13–21. https://doi.org/10.1007/s002530051357
Christenson L, Sims R (2011) Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol Adv 29:686–702. https://doi.org/10.1016/j.biotechadv.2011.05.015
Corneli E, Adessi A, Dragoni F et al (2016) Agroindustrial residues and energy crops for the production of hydrogen and poly-β-hydroxybutyrate via photofermentation. Bioresour Technol 216:941–947. https://doi.org/10.1016/j.biortech.2016.06.046
De Philippis R, Ena A, Guastini M et al (1992) Factors affecting poly-β-hydroxybutyrate accumulation in cyanobacteria and in purple non-sulfur bacteria. FEMS Microbiol Rev 9:187–194. https://doi.org/10.1111/j.1574-6968.1992.tb05837.x
Dietrich K, Dumont M-J, Del Rio LF, Orsat V (2017) Producing PHAs in the bioeconomy—towards a sustainable bioplastic. Sustain Prod Consump 9:58–70. https://doi.org/10.1016/j.spc.2016.09.001
Erb TJ, Berg IA, Brecht V et al (2007) Synthesis of C5-dicarboxylic acids from C2-units involving crotonyl-CoA carboxylase/reductase: the ethylmalonyl-CoA pathway. Proc Natl Acad Sci 104:10631. https://doi.org/10.1073/pnas.0702791104
Favinger J, Stadtwald R, Gest H (1989) Rhodospirillum centenum, sp. nov., a thermotolerant cyst-forming anoxygenic photosynthetic bacterium. Antonie Van Leeuwenhoek 55:291–296. https://doi.org/10.1007/BF00393857
Fradinho JC, Domingos JMB, Carvalho G et al (2013a) Polyhydroxyalkanoates production by a mixed photosynthetic consortium of bacteria and algae. Bioresour Technol 132:146–153. https://doi.org/10.1016/j.biortech.2013.01.050
Fradinho JC, Oehmen A, Reis MAM (2013b) Effect of dark/light periods on the polyhydroxyalkanoate production of a photosynthetic mixed culture. Bioresour Technol 148:474–479. https://doi.org/10.1016/j.biortech.2013.09.010
Fradinho JC, Oehmen A, Reis MAM (2014) Photosynthetic mixed culture polyhydroxyalkanoate (PHA) production from individual and mixed volatile fatty acids (VFAs): substrate preferences and co-substrate uptake. J Biotechnol 185:19–27. https://doi.org/10.1016/j.jbiotec.2014.05.035
Fradinho JC, Reis MAM, Oehmen A (2016) Beyond feast and famine: selecting a PHA accumulating photosynthetic mixed culture in a permanent feast regime. Water Res 105:421–428. https://doi.org/10.1016/j.watres.2016.09.022
Fradinho JC, Oehmen A, Reis MAM (2019) Improving polyhydroxyalkanoates production in phototrophic mixed cultures by optimizing accumulator reactor operating conditions. Int J Biol Macromol 126:1085–1092. https://doi.org/10.1016/j.ijbiomac.2018.12.270
García D, de Godos I, Domínguez C et al (2019) A systematic comparison of the potential of microalgae-bacteria and purple phototrophic bacteria consortia for the treatment of piggery wastewater. Bioresour Technol 276:18–27. https://doi.org/10.1016/j.biortech.2018.12.095
Gebicki J, Modigell M, Schumacher M et al (2009) Development of photobbioreactors for anoxygenic production of hydrogen by purple bacteria. Chem Eng Trans 18:363–368. https://doi.org/10.3303/CET0918058
Ghimire A, Valentino S, Frunzo L et al (2015) Biohydrogen production from food waste by coupling semi-continuous dark-photofermentation and residue post-treatment to anaerobic digestion: a synergy for energy recovery. Int J Hydrog Energy 40:16045–16055. https://doi.org/10.1016/j.ijhydene.2015.09.117
Ghimire A, Valentino S, Frunzo L et al (2016) Concomitant biohydrogen and poly-β-hydroxybutyrate production from dark fermentation effluents by adapted Rhodobacter sphaeroides and mixed photofermentative cultures. Spec Issue Bioenergy Bioprod Environ Sustain 217:157–164. https://doi.org/10.1016/j.biortech.2016.03.017
Ghosh S, Dairkee UK, Chowdhury R, Bhattacharya P (2017) Hydrogen from food processing wastes via photofermentation using Purple Non-sulfur Bacteria (PNSB)–a review. Energy Convers Manag 141:299–314. https://doi.org/10.1016/j.enconman.2016.09.001
Guerra-Blanco P, Cortes O, Poznyak T et al (2018) Polyhydroxyalkanoates (PHA) production by photoheterotrophic microbial consortia: effect of culture conditions over microbial population and biopolymer yield and composition. Eur Polym J 98:94–104. https://doi.org/10.1016/j.eurpolymj.2017.11.007
Hanada S (2016) Anoxygenic photosynthesis - a photochemical reaction that does not contribute to oxygen reproduction. Microbes Env 31:1–3. https://doi.org/10.1264/jsme2.ME3101rh
Heinrich D, Raberg M, Fricke P et al (2016) Synthesis gas (Syngas)-derived medium-chain-length polyhydroxyalkanoate synthesis in engineered Rhodospirillum rubrum. Appl Environ Microbiol 82:6132–6140. https://doi.org/10.1128/AEM.01744-16
Higuchi-Takeuchi M, Numata K (2019) Acetate-inducing metabolic states enhance polyhydroxyalkanoate production in marine purple non-sulfur bacteria under aerobic conditions. Front Bioeng Biotechnol 7:118. https://doi.org/10.3389/fbioe.2019.00118
Higuchi-Takeuchi M, Morisaki K, Numata K (2016a) A screening method for the isolation of polyhydroxyalkanoate-producing purple non-sulfur photosynthetic bacteria from natural seawater. Front Microbiol 7:1509. https://doi.org/10.3389/fmicb.2016.01509
Higuchi-Takeuchi M, Morisaki K, Toyooka K, Numata K (2016b) Synthesis of high-molecular-weight polyhydroxyalkanoates by marine photosynthetic purple bacteria. Plos One 11:e0160981. https://doi.org/10.1371/journal.pone.0160981
Hülsen T, Batstone DJ, Keller J (2014) Phototrophic bacteria for nutrient recovery from domestic wastewater. Water Res 50:18–26. https://doi.org/10.1016/j.watres.2013.10.051
Hustede E, Steinbüchel A, Schlegel HG (1993) Relationship between the photoproduction of hydrogen and the accumulation of PHB in non-sulphur purple bacteria. Appl Microbiol Biotechnol 39:87–93. https://doi.org/10.1007/BF00166854
Imhoff JF (2017) Anoxygenic phototrophic bacteria from extreme environments. In: Hallenbeck PC (ed) Modern topics in the phototrophic prokaryotes: environmental and applied aspects. Springer International Publishing, Switzerland, pp 427–480
Imhoff JF (2017) Diversity of anaerobic anoxygenic phototrophic purple bacteria. In: Hallenbeck PC (ed) Modern topics in the phototrophic prokaryotes: environmental and applied aspects. Springer International Publishing, Switzerland, pp 47–85
Imhoff JF, Hiraishi A, Suling J (2005) Anoxygenic phototrophic purple bacteria. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds) Bergey’s manual of systematic bacteriology. Springer, Boston, pp 119–132
Ismail KSK, Najafpour G, Younesi H et al (2008) Biological hydrogen production from CO: bioreactor performance. Biochem Eng J 39:468–477. https://doi.org/10.1016/j.bej.2007.11.003
Jiang G, Hill DJ, Kowalczuk M et al (2016) Carbon sources for polyhydroxyalkanoates and an integrated biorefinery. Int J Mol Sci 17:1157. doi:https://doi.org/10.3390/ijms17071157. https://doi.org/10.3390/ijms17071157
Kanekar PP, Kanekar SP, Kelkar AS, Dhakephalkar PK (2012) Halophiles – Taxonomy, diversity, physiology and applications. In: Satyanarayana T, Johri B, Anil P (eds) Microorganisms in environmental management. Springer, Dordrecht, pp 1–34
Kantachote D, Torpee S, Umsakul K (2005) The potential use of anoxygenic phototrophic bacteria for treating latex rubber sheet wastewater. Electron J Biotechnol. https://doi.org/10.4067/S0717-34582005000300012
Kapdan IK, Kargi F, Oztekin R, Argun H (2009) Bio-hydrogen production from acid hydrolyzed wheat starch by photo-fermentation using different Rhodobacter sp. Int J Hydrog Energy 34:2201–2207. https://doi.org/10.1016/j.ijhydene.2009.01.017
Kars G, Ceylan A (2013) Biohydrogen and 5-aminolevulinic acid production from waste barley by Rhodobacter sphaeroides O.U.001 in a biorefinery concept. Int J Hydrog Energy 38:5573–5579. https://doi.org/10.1016/j.ijhydene.2013.03.013
Kars G, Gündüz U (2010) Towards a super H2 producer: Improvements in photofermentative biohydrogen production by genetic manipulations. ISMF-09 35:6646–6656. https://doi.org/10.1016/j.ijhydene.2010.04.037
Khatipov E, Miyake M, Miyake J, Asada Y (1998a) Polyhydroxybutyrate accumulation and hydrogen evolution by Rhodobacter sphaeroides as a function of nitrogen availability. In: Zaborsky OR, Benemann JR, Matsunaga T et al (eds) BioHydrogen. Springer, Boston, pp 157–161
Khatipov E, Miyake M, Miyake J, Asada Y (1998) Accumulation of poly-β-hydroxybutyrate by Rhodobacter sphaeroides on various carbon and nitrogen substrates. FEMS Microbiol Lett 162:39–45. https://doi.org/10.1111/j.1574-6968.1998.tb12976.x
Kim M-S, Kim D-H, Son H-N et al (2011) Enhancing photo-fermentative hydrogen production by Rhodobacter sphaeroides KD131 and its PHB synthase deleted-mutant from acetate and butyrate. 2010 AsianAPEC BioH2 36:13964–13971. https://doi.org/10.1016/j.ijhydene.2011.03.099
Kim M-S, Kim D-H, Cha J, Lee JK (2012) Effect of carbon and nitrogen sources on photo-fermentative H2 production associated with nitrogenase, uptake hydrogenase activity, and PHB accumulation in Rhodobacter sphaeroides KD131. Bioresour Technol 116:179–183. https://doi.org/10.1016/j.biortech.2012.04.011
Kniewel R, Lopez OR, Prieto MA (2019) Biogenesis of medium-chain-length polyhydroxyalkanoates. In: Geiger O (ed) Biogenesis of fatty acids, lipids and membranes. Springer International Publishing, Cham, pp 457–481
Koller M, Braunegg G (2018) Advanced approaches to produce polyhydroxyalkanoate (PHA) biopolyesters in a sustainable and economic fashion. Eurobiotech J 2:89–103. https://doi.org/10.2478/ebtj-2018-0013
Kompantseva EI, Komova AV, Sorokin DY (2010) Communities of anoxygenic phototrophic bacteria in the saline soda lakes of the Kulunda steppe (Altai Krai). Microbiol 79:89–95. https://doi.org/10.1134/S0026261710010121
Kumar BV, Ramprasad EVV, Sasikala C, Ramana CV (2013) Rhodopseudomonas pentothenatexigens sp. nov. and Rhodopseudomonas thermotolerans sp. nov., isolated from paddy soils. Int J Syst Evol Microbiol 63:200–207. https://doi.org/10.1099/ijs.0.038620-0
Lai Y-C, Liang C-M, Hsu S-C et al (2017) Polyphosphate metabolism by purple non-sulfur bacteria and its possible application on photo-microbial fuel cell. J Biosci Bioeng 123:722–730. https://doi.org/10.1016/j.jbiosc.2017.01.012
Laurinavichene T, Tekucheva D, Laurinavichius K, Tsygankov A (2018) Utilization of distillery wastewater for hydrogen production in one-stage and two-stage processes involving photofermentation. Enzyme Microb Technol 110:1–7. https://doi.org/10.1016/j.enzmictec.2017.11.009
Liebergesell M, Hustede E, Timm A et al (1991) Formation of poly(3-hydroxyalkanoates) by phototrophic and chemolithotrophic bacteria. Arch Microbiol 155:415–421. https://doi.org/10.1007/BF00244955
Luongo V, Ghimire A, Frunzo L et al (2017) Photofermentative production of hydrogen and poly-β-hydroxybutyrate from dark fermentation products. Bioresour Technol 228:171–175. https://doi.org/10.1016/j.biortech.2016.12.079
Mack EE, Mandelco L, Woese CR, Madigan MT (1993) Rhodospirillum sodomense, sp. nov., a Dead Sea rhodospirillum species. Arch Microbiol 160:363–371. https://doi.org/10.1007/BF00252222
Madigan MT, Jung DO (2009) An overview of purple bacteria: systematics, physiology, and habitats. In: Hunter CN, Daldal F, Thurnauer MC, Beatty JT (eds) The purple phototrophic bacteria. Advances in photosynthesis and respiration, vol 28. Springer, Dordrecht
Marang L, van Loosdrecht MCM, Kleerebezem R (2018) Enrichment of PHA-producing bacteria under continuous substrate supply. N Biotechnol 41:55–61. https://doi.org/10.1016/j.nbt.2017.12.001
Melnicki MR, Eroglu E, Melis A (2009) Changes in hydrogen production and polymer accumulation upon sulfur-deprivation in purple photosynthetic bacteria. Int J Hydrog Energy 34:6157–6170. https://doi.org/10.1016/j.ijhydene.2009.05.115
Merugu R, Rudra M, Girisham S, Reddy SM (2012) Biotechnological applications of purple non sulphur phototrophic bacteria: a minireview. Int J App Biol Pharm 3:376–384
Molina Grima E, Belarbi E-H, Acién Fernández FG et al (2003) Recovery of microalgal biomass and metabolites: process options and economics. Biotechnol Adv 20:491–515. https://doi.org/10.1016/S0734-9750(02)00050-2
Montano-Herrera L, Laycock B, Werker A, Pratt S (2017) The evolution of polymer composition during PHA accumulation: the significance of reducing equivalents. Bioengineering 4:20. https://doi.org/10.3390/bioengineering4010020
Montiel-Corona V, Buitrón G (2021) Polyhydroxyalkanoates from organic waste streams using purple non-sulfur bacteria. Bioresour Technol 323:124610. https://doi.org/10.1016/j.biortech.2020.124610
Montiel-Corona V, Revah S, Morales M (2015) Hydrogen production by an enriched photoheterotrophic culture using dark fermentation effluent as substrate: effect of flushing method, bicarbonate addition, and outdoor–indoor conditions. Int J Hydrog Energy 40:9096–9105. https://doi.org/10.1016/j.ijhydene.2015.05.067
Montiel Corona V, Le Borgne S, Revah S, Morales M (2017) Effect of light-dark cycles on hydrogen and poly-β-hydroxybutyrate production by a photoheterotrophic culture and Rhodobacter capsulatus using a dark fermentation effluent as substrate. Bioresour Technol 226:238–246. https://doi.org/10.1016/j.biortech.2016.12.021
Możejko-Ciesielska J, Kiewisz R (2016) Bacterial polyhydroxyalkanoates: still fabulous? Microbiol Res 192:271–282. https://doi.org/10.1016/j.micres.2016.07.010
Muhr A, Rechberger EM, Salerno A et al (2013) Biodegradable latexes from animal-derived waste: biosynthesis and characterization of mcl-PHA accumulated by Ps. citronellolis. Renew Polym Multifunct Mater Prop Process Appl 73:1391–1398. https://doi.org/10.1016/j.reactfunctpolym.2012.12.009
Mukhopadhyay M, Patra A, Paul AK (2013) Phototrophic growth and accumulation of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by purple nonsulfur bacterium Rhodopseudomonas palustris SP5212. J Polym 2013:523941. https://doi.org/10.1155/2013/523941
Mukhopadhyay M, Patra A, Paul AK (2005) Production of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by Rhodopseudomonas palustris SP5212. World J Microbiol Biotechnol 21:765–769. https://doi.org/10.1007/s11274-004-5565-y
Namsaraev ZB, Gorlenko VM, Namsaraev BB et al (2003) The structure and biogeochemical activity of the phototrophic communities from the Bol’sherechenskii alkaline hot spring. Mikrobiologiia 72:193–203. https://doi.org/10.1023/A:1023272131859
Ni Y-Y, Kim DY, Chung MG et al (2010) Biosynthesis of medium-chain-length poly(3-hydroxyalkanoates) by volatile aromatic hydrocarbons-degrading Pseudomonas fulva TY16. Bioresour Technol 101:8485–8488. https://doi.org/10.1016/j.biortech.2010.06.033
Ohashi S, Iemura T, Okada N et al (2010) An overview on chlorophylls and quinones in the photosystem I-type reaction centers. Photosynth Res 104:305–319. https://doi.org/10.1007/s11120-010-9530-3
Olson JM (2013) Green bacteria: the light-harvesting chlorosome. In: Lennarz WJ, Lane MD (eds) Encyclopedia of biological chemistry (Second Edition). Academic Press, Waltham, pp 513–516
Özsoy Demiriz B, Kars G, Yücel M et al (2019) Hydrogen and poly-β-hydroxybutyric acid production at various acetate concentrations using Rhodobacter capsulatus DSM 1710. Int J Hydrog Energy 44:17269–17277. https://doi.org/10.1016/j.ijhydene.2019.02.036
Padovani G, Carlozzi P, Seggiani M et al (2016) PHB-rich biomass and BioH2 production by means of photosynthetic microorganisms. Chem Eng Trans 49:55–60. https://doi.org/10.3303/CET1649010
Padovani G, Emiliani G, Giovanelli A et al (2018) Assessment of glycerol usage by five different purple non-sulfur bacterial strains for bioplastic production. J Env Chem Eng 6:616–622. https://doi.org/10.1016/j.jece.2017.12.050
Pagliano G, Ventorino V, Panico A, Pepe O (2017) Integrated systems for biopolymers and bioenergy production from organic waste and by-products: a review of microbial processes. Biotechnol Biofuels 10:113. https://doi.org/10.1186/s13068-017-0802-4
Pattanamanee W, Choorit W, Deesan C et al (2012) Photofermentive production of biohydrogen from oil palm waste hydrolysate. Int J Hydrog Energy 37:4077–4087. https://doi.org/10.1016/j.ijhydene.2011.12.002
Petushkova E, Iuzhakov S, Tsygankov A (2019) Differences in possible TCA cycle replenishing pathways in purple non-sulfur bacteria possessing glyoxylate pathway. Photosynth Res 139:523–537. https://doi.org/10.1007/s11120-018-0581-1
Pfennig N (1969) Rhodopseudomonas acidophila, sp. n., a new species of the budding purple nonsulfur bacteria. J Bacteriol 99:597–602
Pfennig N (1974) Rhodopseudomonas globiformis, sp. n., a new species of the Rhodospirillaceae. Arch Microbiol 100:197–206. https://doi.org/10.1007/BF00446317
Pfennig N, Lünsdorf H, Süling J, Imhoff JF (1997) Rhodospira trueperi gen. nov., spec. nov., a new phototrophic Proteobacterium of the alpha group. Arch Microbiol 168:39–45. https://doi.org/10.1007/s002030050467
Podola B, Li T, Melkonian M (2017) Porous substrate bioreactors: a paradigm shift in microalgal biotechnology? Trends Biotechnol 35:121–132. https://doi.org/10.1016/j.tibtech.2016.06.004
Policastro G, Luongo V, Fabbricino M (2020) Biohydrogen and poly-β-hydroxybutyrate production by winery wastewater photofermentation: effect of substrate concentration and nitrogen source. J Environ Manag 271:111006. https://doi.org/10.1016/j.jenvman.2020.111006
Povolo S, Romanelli MG, Basaglia M et al (2013) Polyhydroxyalkanoate biosynthesis by Hydrogenophaga pseudoflava DSM1034 from structurally unrelated carbon sources. Biotechnol Bio Green Econ 30:629–634. https://doi.org/10.1016/j.nbt.2012.11.019
Prasertsan P, Choorit W, Suwanno S (1993) Optimization for growth of Rhodocyclus gelatinosus in seafood processing effluents. World J Microbiol Biotechnol 9:593–596. https://doi.org/10.1007/BF00386302
Qi X, Ren Y, Tian E, Wang X (2017) The exploration of monochromatic near-infrared LED improved anoxygenic photosynthetic bacteria Rhodopseudomonas sp. for wastewater treatment. Bioresour Technol 241:620–626. https://doi.org/10.1016/j.biortech.2017.05.202
Ramana CV, Sasikala C, Arunasri K et al (2006) Rubrivivax benzoatilyticus sp. nov., an aromatic, hydrocarbon-degrading purple betaproteobacterium. Int J Syst Evol Microbiol 56:2157–2164. https://doi.org/10.1099/ijs.0.64209-0
Ranaivoarisoa TO, Singh R, Rengasamy K et al (2019) Towards sustainable bioplastic production using the photoautotrophic bacterium Rhodopseudomonas palustris TIE-1. J Ind Microbiol Biotechnol 46:1401–1417. https://doi.org/10.1007/s10295-019-02165-7
Rigouin C, Lajus S, Ocando C et al (2019) Production and characterization of two medium-chain-length polydroxyalkanoates by engineered strains of Yarrowia lipolytica. Microb Cell 18:99. https://doi.org/10.1186/s12934-019-1140-y
Sali S, Mackey HR (2021) Integration of polyhydroxyalkanoates production with industrial wastewater treatment. In: Gothandam KM, Ranjan S, Dasgupta N, Lichtfouse E (eds) Environmental biotechnology, vol 3. Springer, Switzerland
Samorì C, Abbondanzi F, Galletti P et al (2015) Extraction of polyhydroxyalkanoates from mixed microbial cultures: impact on polymer quality and recovery. Bioresour Technol 189:195–202. https://doi.org/10.1016/j.biortech.2015.03.062
Sathya A, Velmurugan S, Arockiasamy S et al (2018) Production of polyhydroxyalkanoates from renewable sources using bacteria. J Polym Environ 26:3995–4012. https://doi.org/10.1007/s10924-018-1259-7
Serafim LS, Lemos PC, Albuquerque MGE, Reis MAM (2008) Strategies for PHA production by mixed cultures and renewable waste materials. Appl Microbiol Biotechnol 81:615–628. https://doi.org/10.1007/s00253-008-1757-y
Serôdio J, Vieira S, Cruz S (2008) Photosynthetic activity, photoprotection and photoinhibition in intertidal microphytobenthos as studied in situ using variable chlorophyll fluorescence. Cont Shelf Res 28:1363–1375. https://doi.org/10.1016/j.csr.2008.03.019
Shahid S, Mosrati R, Ledauphin J et al (2013) Impact of carbon source and variable nitrogen conditions on bacterial biosynthesis of polyhydroxyalkanoates: Evidence of an atypical metabolism in Bacillus megaterium DSM 509. J Biosci Bioeng 116:302–308. https://doi.org/10.1016/j.jbiosc.2013.02.017
Sharma PK, Munir RI, Blunt W et al (2017) Synthesis and physical properties of polyhydroxyalkanoate polymers with different monomer compositions by recombinant Pseudomonas putida LS46 expressing a novel PHA SYNTHASE (PhaC116) enzyme. Appl Sci 7:242. https://doi.org/10.3390/app7030242
Sheu D-S, Chen W-M, Yang J-Y, Chang R-C (2009) Thermophilic bacterium Caldimonas taiwanensis produces poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from starch and valerate as carbon sources. Enzyme Microb Technol 44:289–294. https://doi.org/10.1016/j.enzmictec.2009.01.004
Suk AW, Jae PS, Yup LS (2000) Production of poly(3-hydroxybutyrate) by fed-batch culture of recombinant Escherichia coli with a highly concentrated whey solution. Appl Environ Microbiol 66:3624–3627. https://doi.org/10.1128/AEM.66.8.3624-3627.2000
Suzuki T, Tsygankov AA, Miyake J et al (1995) Accumulation of poly-(hydroxybutyrate) by a non-sulfur photosynthetic bacterium, Rhodobacter sphaeroides RV at different pH. Biotechnol Lett 17:395–400. https://doi.org/10.1007/BF00130796
Talaiekhozani A, Rezania S (2017) Application of photosynthetic bacteria for removal of heavy metals, macro-pollutants and dye from wastewater: a review. J Water Process Eng 19:312–321. https://doi.org/10.1016/j.jwpe.2017.09.004
Tamang P, Banerjee R, Köster S, Nogueira R (2019) Comparative study of polyhydroxyalkanoates production from acidified and anaerobically treated brewery wastewater using enriched mixed microbial culture. J Environ Sci 78:137–146. https://doi.org/10.1016/j.jes.2018.09.001
Tan G-YA, Chen C-L, Li L et al (2014) Start a research on biopolymer polyhydroxyalkanoate (PHA): a review. Polymers. https://doi.org/10.3390/polym6030706
Tanskul S, Srisai S, Nualla-Ong A (2016) A purple non-sulfur bacterium producing polyhydroxybutyrate and the conserved region of PHA synthase gene. Biosci J 32:1341–1351. https://doi.org/10.14393/BJ-v32n1a2016-33801
Tufail S, Munir S, Jamil N (2017) Variation analysis of bacterial polyhydroxyalkanoates production using saturated and unsaturated hydrocarbons. Braz J Microbiol 48:629–636. https://doi.org/10.1016/j.bjm.2017.02.008
Urtuvia V, Villegas P, González M, Seeger M (2014) Bacterial production of the biodegradable plastics polyhydroxyalkanoates. Int J Biol Macromol 70:208–213. https://doi.org/10.1016/j.ijbiomac.2014.06.001
Uyar B, Eroglu I, Yücel M et al (2007) Effect of light intensity, wavelength and illumination protocol on hydrogen production in photobioreactors. Int J Hydrog Energy 32:4670–4677. https://doi.org/10.1016/j.ijhydene.2007.07.002
Vandi LJ, Chan CM, Werker A et al (2018) Wood-PHA composites: Mapping opportunities. Polymers 10:751. https://doi.org/10.3390/polym10070751
Verlinden RAJ, Hill DJ, Kenward MA et al (2007) Bacterial synthesis of biodegradable polyhydroxyalkanoates. J Appl Microbiol 102:1437–1449. https://doi.org/10.1111/j.1365-2672.2007.03335.x
Villano M, Valentino F, Barbetta A et al (2014) Polyhydroxyalkanoates production with mixed microbial cultures: from culture selection to polymer recovery in a high-rate continuous process. N Biotechnol 31:289–296. https://doi.org/10.1016/j.nbt.2013.08.001
Vincenzini M, Marchini A, Ena A, De Philippis R (1997) H2 and poly-β-hydroxybutyrate, two alternative chemicals from purple non sulfur bacteria. Biotechnol Lett 19:759–762. https://doi.org/10.1023/A:1018336209252
Voinova O, Gladyshev M, Volova TG (2008) Comparative study of PHA degradation in natural reservoirs having various types of ecosystems. Macromol Symp 269:34–37. https://doi.org/10.1002/masy.200850906
Wang F, Lee SY (1997) Poly(3-hydroxybutyrate) production with high productivity and high polymer content by a fed-batch culture of Alcaligenes latus under nitrogen limitation. Appl Env Microbiol 63:3703–3706. https://doi.org/10.1128/aem.63.9.3703-3706.1997
Wang H, Zhang G, Peng M et al (2016) Synthetic white spirit wastewater treatment and biomass recovery by photosynthetic bacteria: Feasibility and process influence factors. Int Biodeterior Biodegrad 113:134–138. https://doi.org/10.1016/j.ibiod.2016.01.001
Ward PG, Goff M, Donner M et al (2006) A two step chemo-biotechnological conversion of polystyrene to a biodegradable thermoplastic. Environ Sci Technol 40:2433–2437. https://doi.org/10.1021/es0517668
Wu S, Butt HJ (2017) Near-infrared photochemistry at interfaces based on upconverting nanoparticles. Phys Chem Chem Phys PCCP 19:23585–23596. https://doi.org/10.1039/c7cp01838j
Wu SC, Liou SZ, Lee CM (2012) Correlation between bio-hydrogen production and polyhydroxybutyrate (PHB) synthesis by Rhodopseudomonas palustris WP3-5. Bioresour Technol 113:44–50. https://doi.org/10.1016/j.biortech.2012.01.090
Zagrodnik R, Laniecki M (2015) The role of pH control on biohydrogen production by single stage hybrid dark- and photo-fermentation. Bioresour Technol 194:187–195. https://doi.org/10.1016/j.biortech.2015.07.028
Zhi R, Yang A, Zhang G et al (2019) Effects of light-dark cycles on photosynthetic bacteria wastewater treatment and valuable substances production. Bioresour Technol 274:496–501. https://doi.org/10.1016/j.biortech.2018.12.021
Zhou Q, Zhang P, Zhang G (2014) Biomass and carotenoid production in photosynthetic bacteria wastewater treatment: Effects of light intensity. Bioresour Technol 171:330–335. https://doi.org/10.1016/j.biortech.2014.08.088
Zhou Q, Zhang P, Zhang G (2015a) Biomass and pigments production in photosynthetic bacteria wastewater treatment: Effects of light sources. Bioresour Technol 179:505–509. https://doi.org/10.1016/j.biortech.2014.12.077
Zhou Q, Zhang P, Zhang G, Peng M (2015b) Biomass and pigments production in photosynthetic bacteria wastewater treatment: Effects of photoperiod. Bioresour Technol 190:196–200. https://doi.org/10.1016/j.biortech.2015.04.092
Zhu C, Nomura CT, Perrotta JA et al (2010) Production and characterization of poly-3-hydroxybutyrate from biodiesel-glycerol by Burkholderia cepacia ATCC 17759. Biotechnol Prog 26:424–430. https://doi.org/10.1002/btpr.355
Acknowledgements
The authors would like to acknowledge the support for this project by Qatar National Research fund under the national Priorities Research Program, Grant NPRP11-S-0110-180245.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sali, S., Mackey, H.R. The application of purple non-sulfur bacteria for microbial mixed culture polyhydroxyalkanoates production. Rev Environ Sci Biotechnol 20, 959–983 (2021). https://doi.org/10.1007/s11157-021-09597-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11157-021-09597-7