
Abstract
Background and aims
Understanding how altered precipitation patterns affect the dynamics of phosphorus (P) fractions in both soil and foliage is crucial for predicting climate-induced changes in plant growth and community structure, especially within tropical forests with P-impoverished soils.
Methods
We conducted a nine-year precipitation experiment in a secondary tropical forest, simulating delayed (DW) and wetter wet season (WW) to reflect potential precipitation changes. We analyzed P fractions in soil (0–10 cm and 10–20 cm) and foliage of four dominant trees, and investigated the correlations among these P fractions and photosynthesis.
Results
DW treatment reduced soil P bioavailability and induced foliar P redistribution, while WW treatment increased soil pH and NH4+-N content but had little influence on soil P form. DW treatment generally reduced foliar total P levels and diminished various P fractions with notable species-specific disparities. Moreover, DW treatment variably reduced area-based photosynthetic carbon assimilation rates (Aarea), exhibiting species-specific effects, while WW treatment inconsistently elevated Aarea across species. The structural equation model revealed that Aarea was directly influenced by foliar P fractions and photosynthetic nutrient efficiency, and indirectly by precipitation treatment on soil properties and P fractions.
Conclusion
Altered seasonal precipitation patterns affect soil bioavailable P forms and/or soil chemical properties, and thereby influence foliar P fractions allocation and photosynthesis of dominant trees. Soil and foliar P fractions responses to precipitation treatments varied across different sampling months, underscoring the complexity of P cycle and suggesting tree acclimation and resilience. These insights improve our understanding of climate effects on nutrient cycles and resilience.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Climate change is altering global precipitation patterns, resulting in an overarching trend of increased precipitation variability (Dore 2005; IPPC 2023). This is evident in the dry tropics where there has been an increased uncertainty in the interannual seasonal rainfall. This unpredictability encompasses variations in rainfall duration, and intensity, and a shift in the seasonal distribution (Feng et al. 2013). In regions with subtropical and tropical monsoon climates, such as southeast China, there is a shift toward wetter conditions with more instances of heavy rainfall during the rainy season, despite a decrease in the number of rainy days (Song et al. 2011; Zhang and Zhao 2022; Zhou et al. 2011). Meanwhile, southern China has observed a delayed onset of the rainy season, even though there haven’t been significant changes in the annual total precipitation (Luo et al. 2008; Zhou et al. 2011). Consequently, in southern China, where tropical forests are located, there is a discernible trend toward a delayed wet season. This shift is coupled with a higher frequency of heavy rainfall events characterized by increased intensity, marking a transition to a wetter wet season.
Tropical forests, covering less than 10% of the Earth's land surface, provide vital ecosystem services such as carbon storage, climate regulation, and support of nearly two-thirds of global biodiversity (Bradshaw et al. 2009; Dirzo and Raven 2003; Spracklen et al. 2018). Additionally, these forests play a critical role in climate change mitigation and exert a substantial influence on local and regional precipitation through their role in the hydrological cycle (Smith et al. 2023). However, these ecosystems are increasingly threatened by altered precipitation patterns, which can disrupt forest soil nutrient cycling and availability by affecting the wet-dry cycle of soils (Afreen et al. 2019). This disruption may profoundly affect plant growth and community composition, potentially exacerbating natural disturbances that damage functional diversity (Aguirre-Gutiérrez et al. 2022; Deb et al. 2018). Furthermore, such changes significantly reduce the carbon sequestration capacity of these forests on a larger scale (Hajek and Knapp 2022; Wang et al. 2018). Thus, it is critical to understand the influence of changing precipitation patterns on tropical forest ecosystems (Afreen et al. 2019; Korell et al. 2021; Zhang et al. 2023).
Phosphorus (P) availability in soil restricts plant productivity in tropical forests with ancient and strongly weathered soils (Cheng et al. 2020; Dietrich et al. 2017). Although total P in soil is typically high compared to its bioavailable forms, much of it is immobile and inaccessible to plants (Lambers 2021; Shen et al. 2011). Soil P is present in various forms, including organic (Po) and inorganic (Pi) compounds. Notably, a substantial portion of soil total P, estimated at approximately 30% to 65%, existed in an organic form, which is not readily available for plant uptake (Hedley et al. 1982; Mwende Muindi 2019; Shen et al. 2011). These soil P fractions are categorized into easily-available (resin Pi, NaHCO3 Pi, and NaHCO3 Po), moderately-available (NaOH Pi, NaOH Po, and DHCl Pi), and non-available (CHCl Pi, CHCl Po, and residual Pt) P, with varying availability and turnover times for plants (Helfenstein et al. 2018; Maranguit et al. 2017; Tiessen et al. 1984). Precipitation, as a critical climatic factor, directly or indirectly influences soil physicochemical properties, thereby impacting P availability through changes in soil P forms (Hou et al. 2018b; Zhu et al. 2021). For instance, increased rainfall in tropical forests of southern China can suppress soil organic P mineralization by enhancing P adsorption and limiting soil P uptake by plants (Sun et al. 2020). Conversely, long-term throughfall exclusion experiments showed a decrease in soil Po, correlating with reduced plant root and soil microbial biomass in this tropical forest (Fan et al. 2021). Transformations in soil P fractions could significantly influence plant P nutrition (Hou et al. 2018a; Lambers 2021; Niederberger et al. 2019). Although soil P has been traditionally categorized into several different chemical fractions, the practical relevance of these fractions for plant utilization remains unproven (Lambers 2021). Hence, in the context of altered precipitation patterns, it’s vital to investigate the impact of soil P fraction redistribution on tropical forest P nutrition and its potential to exacerbate P limitations.
Different plant species employ unique strategies for P acquisition from the soil, targeting not only readily available P fractions but also less accessible and recalcitrant forms (Niederberger et al. 2017; Zhong et al. 2020). Accordingly, the evaluation of P availability to plants should encompass a comprehensive consideration of both easily accessible and variably available fractions. Several studies have reported a negative correlation between foliar P and moderately labile P concentration while demonstrating a positive relationship between foliar P and soil labile P concentration, which significantly contributes to plant P availability compared to other soil-P fractions (Gao et al. 2022a; Niederberger et al. 2019). Conversely, Yan et al. (2019) indicated a stronger correlation between foliar P and soil total P concentration rather than soil resin P concentration. These inconsistent findings underscore the complexity of the relationship between foliar P and soil P fractions. Foliar P, comprising both inorganic phosphate and four organic fractions (metabolite P, nucleic acid P, lipid P, and residual P) (Suriyagoda et al. 2022; Yan et al. 2019), could dynamically reallocate in response to P deficiency (Yan et al. 2021; Yu et al. 2022; Zhang et al. 2021). In a recent study, Gao et al. (2022a) emphasized that foliar-P fractions, rather than total foliar-P concentration, offer a more effective approach for determining whether plants are experiencing P limitation. Although there is a correlation between soil P and foliar P concentration, our understanding of soil P dynamics, particularly P fractions, and their relationship to foliar P fractions in the context of changing seasonal precipitation patterns in tropical forests, remains limited. Therefore, gaining insight into the relationship between soil and foliage P fractions could enhance our understanding of the P supply and demand between soil and plants, thereby facilitating optimized forest management under climate change.
Furthermore, foliar P concentration closely correlates with photosynthetic traits, particularly metabolic P (including small metabolites [e.g. RuBP, NADP, ATP, ADP] and inorganic phosphate), but this relationship is not always consistent across different studies. Foliar P may limit maximum photosynthetic rates due to its key role in the Calvin-Benson cycle, underscoring its importance for studying P limitation (Lambers 2021; Mo et al. 2019; Suriyagoda et al. 2022). Several studies have demonstrated that photosynthesis is generally diminished under the conditions of P deficiency. Specifically, Hidaka and Kitayama (2013) observed that in 10 tropical montane rainforest tree species, both mean mass-based photosynthetic carbon assimilation rate (Amass) and various foliar P fractions are notably lower in nutrient-deprived ultrabasic serpentine soils than in sedimentary soils. In contrast, Mo et al. (2019) found that a six-year P fertilization in a tropical forest failed to significantly boost area-based photosynthetic carbon assimilation rate (Aarea) and Amass in five woody species, even with prominent enhancements in foliar P fractions. These findings suggest a complex interaction between P availability and photosynthetic performance, necessitating further research. Additionally, in response to low P availability, plants tend to increase their photosynthetic P-use efficiency (PPUE). Hayes et al. (2022) found rice achieves high PPUE with low lipid-P investment, maintaining photosynthetic capacity. Similarly, Tsujii et al. (2023) found that the Proteaceae species studied exhibited low P allocations to structural constituents (such as cell membrane lipid) and nucleic acids, presumably to enhance P allocation to photosynthesis, thereby increasing PPUE. However, despite these insights, our knowledge of how plants allocate their leaf P fraction under soil P redistribution, its subsequent impact on photosynthetic capacity and PPUE, and their acclimation to changing precipitation patterns remains limited.
In this study, we examined seasonal dynamics of soil and foliar P fractions and their interrelationships under two altered seasonal precipitation patterns (i.e., a two-month delay in the onset of the wet season, and a 25% increase of total annual rainfall by heavy events during the wet season) in a secondary tropical forest. Four dominant canopy tree species (Carallia brachiate, Symplocos poilanei, Schefflera heptaphylla, and Tetradium glabrifolium) were selected and foliar P fractions, nitrogen (N) and carbon (C) concentrations, and photosynthetic attributes were measured. Soil P fractions and other soil variables such as pH, total N, and total organic carbon (SOC) were also investigated. This research aimed to elucidate the impacts of these altered precipitation patterns on soil and leaf P fractions and their interrelationships, as well as photosynthetic efficiency and nutrient use efficiency by hypothesized that: (1) a two-month delay in the onset of the wet season could impact the replenishment of Pi concentration by reducing soil Po, leading to lower bioavailable P levels. Increased rainfall would inhibit soil Po mineralization through enhanced P adsorption, thus limiting plant P uptake. (2) These altered precipitation patterns reduced P bioavailability, resulting in reduced leaf P concentrations and the reallocation of leaf P fractions, thereby affecting plant photosynthesis and nutrient use efficiency. The findings from this study could improve our understanding of nutrient cycling and plant-soil interactions in tropical forests under altered seasonal precipitation patterns. Moreover, our insights can inform the development of conservation strategies aimed at enhancing the resilience and sustainability of these critical ecosystems in the face of global precipitation changes.
Materials and methods
Study site and experimental design
The study was conducted in a tropical secondary forest at Xiaoliang Tropical Coastal Ecosystem Research Station (21°27′N, 110°54′E) of the Chinese Academy of Sciences, located at Maoming City, Guangdong Province, China (Fig. 1a). Characterized by a humid tropical climate, the study area experiences distinct wet and dry seasons with an average temperature of 23 ºC and annual precipitation of 1416 mm (Fig. 1b). The total precipitation in 2022 was notably high (1797 mm), and the rainy season from April to September contributed to 82.7% (1486 mm) of the total annual precipitation, exceeding the long-term average. The predominant soil type is latosol soil, which has developed through the weathering of granite. This area has suffered severe erosion primarily due to human disturbance (Ren et al. 2007). In 1959, reforestation efforts were initiated in this area, leading to the establishment of plantations. Presently, the region is home to a broad-leaved mixed forest, which includes tree species such as Symplocos poilanei, Schefflera heptaphylla, Tetradium glabrifolium, Carallia brachiate, and other life forms including shrubs (i.e., Clerodendrum cyrtophyllum, Clerodendrum fortunatum, and Mussaenda pubescens) and ferns (i.e., Dicranopteris dichotoma and Lygodium japonicum).

The geographical location and landscape of Xiaoliang Tropical Coastal Ecosystem Research Station, Guangdong, China (a); the average annual temperature and total annual precipitation during dry and wet seasons (b); the timing of plot treatments and sampling, and view within the plots (c)
An altered precipitation platform was established in the broad-leaved mixed forest in 2013. It consisted of four blocks, each containing three plots (each of 12 m × 12 m). These plots were separated at least 3 m apart and were enclosed by PVC boards. Two precipitation treatments were implemented: a delay of two months in the onset of the wet season (delayed wet season, DW), and a 25% supplementation of total annual precipitation by intense incidents during the wet season (wetter wet season, WW), along with a control (CK) (Fig. 1c). The DW treatment was implemented using a transparent rain-shelter positioned 2 m above the ground, which intercepted 60% of throughfall during April–May. The intercepted water was subsequently added back to the plot in October and November using a sprinkler system. This process was conducted eight times, once per week. The WW treatment, on the other hand, involved the addition of groundwater equivalent to 25% of the annual total precipitation during July and August. The same sprinkler method used in the DW plots was also applied to the WW plots. The altered precipitation platform was established in 2013 and had been operating continuously for nine years when we commenced this study in 2022. During 2022, leaf and soil samples were collected in April, June, September, and November to investigate the effects of rainfall interception or water addition during the period of precipitation treatments.
Soil chemical properties and phosphorus fractions
For each sampling, soil cores from two depths (0–10 cm and 10–20 cm) in each plot were gathered using the five-point method with a 3 cm diameter soil auger after removing surface litter. The collected soil was homogenized, sieved (2 mm mesh), and divided into fresh and air-dried portions. Fresh soil was refrigerated at 4°C for nitrate and ammonium nitrogen (NO3−-N and NH4+-N) analysis using dual-wavelength (Norman et al. 1985) and indophenol blue colorimetric (Dorich and Nelson 1983) methods after filtering 10 g fresh soil in 2 M KCl. The air-dried soil was examined for soil organic carbon (SOC), total nitrogen (TN), pH, and phosphorus (P) fractions. Dichromate oxidation and titration with ferrous ammonium sulfate were used to assess SOC concentration, Kjeldahl methods were utilized to determine TN concentration, and pH was measured in a 1:2.5 soil:water solution (w/v) using a glass electrode (Liu et al. 1996).
For soil P fractionation, the modified Hedley et al. (1982) sequential fractionation method by Tiessen and Moir (2007) was applied. Briefly, 0.5 g air-dried soil is combined with two resin strips in a 50 mL centrifuge tube, shaken for 16 h with 30 mL deionized water, and then the phosphate adsorbed on resin strips was extracted with 20 mL 0.5 M HCl, the soil remaining in the tubes was sequentially extracted by 30 mL of 0.5 M NaHCO3 (pH 8.5), 0.1 M NaOH, and 1 M HCl. After each extraction step, tubes are centrifuged, filtered (< 0.45 mm), and supernatants collected for respective P fractions measurement. Total P (Pt) and inorganic P (Pi) were directly measured from extracts, and the difference between Pt and Pi identified the organic P (Po) concentration. Pi was determined using the molybdate blue colorimetric method (Murphy and Riley 1962), and Pt was analyzed similarly after complete digestion with the addition of persulfate and sterilization at 120°C under autoclave pressure for 1 h (Wang et al. 2021). Seven P fractions, including resin Pi, NaHCO3 Pi, NaHCO3 Po, NaOH Pi, NaOH Po, DHCl Pi, and residual Pt, were examined. The soil total P concentration was determined by the summation of these seven P fractions.
Foliar phosphorus fractions, chemical, and photosynthetic properties
For leaf nutrient analysis, we selected four dominant tree species, including C. brachiate, S. poilanei, S. heptaphylla, and T. glabrifolium. Notably, T. glabrifolium is a deciduous species, whereas the other three species are evergreen. For each precipitation treatment, we selected 4–5 healthy individuals per species as sample trees. A total of 58 sample trees were selected. During each sampling, approximately five sun-facing mature leaves from the canopy of the sampling trees were collected using telescopic pruners, capable of reaching a maximum length of 20 m. These leaves were then divided into two parts for chemical analysis. One part was stored at -80°C until freeze-dried by vacuum freeze drier at least for 48 h, then ground to assess inorganic phosphate and various Po fractions (lipid P, metabolite P, nucleic acids P, and residual P). In brief, a 50 mg subsample was used for inorganic phosphate extraction through the acetic-acid extraction method (Yan et al. 2019; Fig. S1) and determined with the molybdenum blue-based method (Ames 1966). Another 50 mg subsample was employed to evaluate P fractions, including lipid P, nucleic acid P, metabolic P (including inorganic phosphate and small metabolites [e.g. ATP, NADP]), and residual P, following an approach adapted from Hidaka and Kitayama (2013) and further by Yan et al. (2019) (Fig. S2). In the presentation of results, inorganic phosphate was isolated from metabolic P and reported as one of five fractions (as metabolite P). The sum of these five P fractions was taken as the total foliar P concentration. The other part was oven-dried at 60°C for 72 h to determine total nitrogen (using the Kjeldahl method) and carbon (using the potassium dichromate-sulfuric acid oxidation method) concentration.
During each field campaign, we selected healthy, sun-exposed mature leaves from the sample trees to investigate their foliar photosynthetic characteristics in situ between 9:00–12:00 and 14:00–16:00. We measured the net photosynthesis rate (Aarea, μmol CO2 m−2 s−1) using a portable photosynthesis system (Li-6800, Li-Cor, Lincoln, NE, USA). In the leaf chamber, we set photosynthetic photon flux density (PPFD) at 1500 µmol m−2 s−1, ambient temperature (range of values between 25 and 30℃), CO2 concentration at 400 ppm for reference, and flow rate at 500 µmol s−1, and all measurements were made using a 6 cm2 chamber (3 cm × 2 cm). Additionally, approximately 8–15 fresh, mature leaves were harvested from each sampled individual tree and leaf area (LA, in cm2) was quantified using a leaf area meter (LI-3100, Li-Cor Biosciences, Lincoln, NE, USA). Subsequently, these leaves were desiccated at 65ºC for 48 h. The dry weight of the leaves (LDW, g) was assessed to ascertain the leaf mass per unit area (LMA, g cm2), calculated as the quotient of LDW by LA. We calculated photosynthetic nitrogen use efficiency (PNUE) and photosynthetic P use efficiency (PPUE) by converting area-based (Aarea) photosynthetic carbon assimilation rates to mass-based (Amass) using LMA.
Statistical analysis
Two-way ANOVAs were used to compare the soil and foliar chemical properties, foliar and soil P fractions concentrations and proportion, and photosynthetic characteristics among the CK, DW, and WW treatments. In this analysis, random factors included blocks for soil-related variables and repeated measures on individual trees for plant-related variables, while the precipitation treatments (CK, DW, WW) were designated as fixed factors. ANOVA was performed using SPSS version 23 software (IBM Corp., Armonk, NY, USA). Pearson’s correlation coefficient was calculated using the corrplot package (Wei and Simko 2017) in R software (R 4.2.2) to estimate the relationship between soil and foliar P fractions and their respective properties among the precipitation treatments. Redundancy analysis (RDA) was conducted using the vegan package (Oksanen et al. 2022) in R software (version 4.2.2) to explore the relationship between the foliar P fractions (response variable) and the soil P fractions of two soil layers (explanatory variable). We employed the piecewiseSEM, nlme, and lme4 packages for the structural equation model (SEM) (Bates et al. 2017; Lefcheck 2015). The initially observed variables were divided into composite variables before being incorporated into the SEM framework. To assess the links between pivotal soil elements and foliar traits influencing photosynthesis, we utilized piecewiseSEM to account for random effects of species and month, providing both “marginal” and “conditional” predictor contributions. We assessed model adequacy using Fisher’s C test and iteratively refined the models based on pathway significance (p < 0.05) as well as the goodness of the model (0 ≤ Fisher’s C/df ≤ 2 and 0.05 < p ≤ 1.00).
Results
Variations in soil chemical properties and phosphorus fractions
Altered precipitation patterns had a significant impact on various soil variables, with pH exhibiting pronounced sensitivity to these changes (Table 1). Specifically, the wetter wet seasons (WW) treatment caused a notable increase in soil pH (4.5–4.8) compared to the CK (4.3–4.6). For instance, during April, June, and September, the pH in the 0–10 cm soil layer under the WW treatment (p = 0.018, 0.002, and 0.001, respectively), as well as in the 10–20 cm layer in September under the same condition (p = 0.020), were significantly higher than those in the CK. Conversely, the delayed wet seasons (DW) treatment had a minimal effect on soil pH. Additionally, the WW treatment in September significantly elevated NH4+-N concentration in both the 0–10 cm (13.2 ± 2.7 mg kg−1 vs 3.3 ± 0.6 mg kg−1) and 10–20 cm (9.1 ± 1.1 mg kg−1 vs 4.8 ± 0.8 mg kg−1) soil layers compared to the CK (p = 0.014 and 0.012, respectively).
Generally, the DW treatment decreased the concentrations of Pi fractions, whereas the WW treatment had a minor impact on soil P fractions, and these effects varied with soil depth (Fig. 2). In April, the DW treatment led to lower DHCl Pi and NaOH Po concentrations in the 0–10 cm soil layer compared to the CK (p = 0.044) and WW treatments (p = 0.036), respectively. Under the WW treatment, the NaHCO3 Po concentration in the 10–20 cm layer was significantly lower than under the CK and DW treatment (p = 0.010 and 0.036, respectively). In June, after two months of 60% rainfall interception, the DW treatment showed a decreasing trend for nearly all Pi fractions in the 0–10 cm soil layer compared to the CK, while the Po and residual P fractions increased correspondingly. However, in the 10–20 cm soil layer, NaHCO3 Pi concentration was higher in the DW plots (14.7 mg kg−1) than the CK plots (7.8 mg kg−1), but NaHCO3 Po values in the DW plots (2.5 mg kg−1) were less than that in the CK (6.9 mg kg−1) and WW plots (7.1 mg kg−1). In September and November, only the P fractions in the 0–10 cm soil layer varied with precipitation treatment. NaOH Pi concentrations in November were significantly lower in the DW plots than in the CK plots (p = 0.030). Moreover, resin Pi concentration in September and DHCl Pi concentrations in November within the DW treatment were both marginally significantly lower than those in the CK, with p-values of 0.055 and 0.059, respectively.

Concentrations of distinct soil inorganic phosphorus (Pi) and organic phosphorus (Po) fractions at depth of 0–10 cm and 10–20 cm soil layers in the natural control (CK), delayed wet season (DW), and wetter wet season (WW) treatments from April to November. The number and error bars represent mean and standard error (n = 4), respectively. Different lowercase letters indicate significant differences (p < 0.05) among the CK, DW, and WW treatments for the same fraction within the same depth
Overall, resin Pi comprised the smallest soil P fraction (≤ 1.5%), whereas residual Pt consistently contributed the largest proportion, varying between 30.1% and 68.5% (Fig. 3). In April, for the 0–10 cm soil layer, compared to the CK treatment, the proportions of NaOH Po were significantly higher under the WW treatment (p = 0.027), and the DHCl Pi proportion was much lower under the DW (p = 0.033), respectively. For the 10–20 cm layer, both the proportions of DHCl Pi in the DW plots (1.6%) and NaHCO3 Po in the WW plots (1.6%) were significantly lower than that in the CK plots (4.7% and 6.1%, respectively). But in June, precipitation treatment didn’t significantly affect P fraction proportions in the 0–10 cm soil layer. Yet, in the 10–20 cm layer, NaHCO3 Po proportions were lower in the DW plots (1.1%) than both the CK and WW plots (3.4% and 3.8%, respectively). In September, precipitation treatment did not induce significant shifts in P fraction proportions in either soil layer. In November, the NaOH Po proportions in the 0–10 cm layer were higher under the DW treatment than the CK (p = 0.026).

Relative proportion of different soil inorganic phosphorus (Pi) and organic phosphorus (Po) fractions to total soil P concentrations at 0–10 cm and 10–20 cm soil layers under the natural control (CK), delayed wet season (DW), and wetter wet season (WW) treatments from April to November (n = 4)
Variations in foliar phosphorus fractions and associated chemical and photosynthetic traits
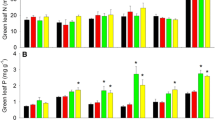
The concentration of foliar P fractions in response to altered precipitation treatments showed species-specific patterns (Fig. 4). In April, precipitation changes significantly impacted only the concentration of foliar P fractions in C. brachiate. The concentration of residual P increased, while that of nucleic acid P decreased under the WW treatment compared to the CK (p = 0.019 and 0.008, respectively). Likewise, the concentration of foliar metabolite P in C. brachiate was lower under the DW treatment than the CK (0.20 mg kg−1 vs 0.34 mg kg−1). In June, the metabolite P concentrations in T. glabrifolium leaves were reduced in the DW plots (p values were < 0.001 and 0.002 for the CK and WW, respectively). In September, all of C. brachiate’s foliar P fractions except metabolite P concentration were lower under the DW treatment than the CK, while S. heptaphylla’s lipid P and T. glabrifolium’s metabolite P concentration increased under the DW treatment compared to the WW treatment. Simultaneously, lipid P, metabolite P, and nucleic acid P concentration in S. poilanei leaves were elevated under the WW treatment compared to the DW treatment. In November, S. poilanei and S. heptaphylla’s residual P concentrations were significantly higher in the DW plots (p = 0.001 and 0.006, respectively), while T. glabrifolium’s residual P concentration was notably lower under the DW treatment (p = 0.002) than the CK. Overall, total foliar P concentration was decreased under the DW treatment. Two species, S. poilanei (June; 1.5 mg g−1) and C. brachiate (September; 2.4 mg g−1), both had significantly lower foliar P concentration under the DW treatment than the CK (1.8 mg g−1and 3.2 mg g−1, respectively; Table 2). Only S. heptaphylla had higher total foliar P concentration under the WW treatment than the CK in November (p = 0.029).

Concentrations of inorganic phosphate and four organic phosphorus fractions in leaves for C. brachiata (CB), S. poilanei (SP), S. heptaphylla (SH), and T. glabrifolium (TG) under the natural control (CK), delayed wet season (DW), and wetter wet season (WW) treatments from April to November. The number and error bars represent mean and standard error (n = 4 or 5), respectively. Different lowercase letters indicate significant differences (p < 0.05) among the CK, DW, and WW treatments for the same fraction within the same species
In April, C. brachiate under the WW treatment increased the proportions of inorganic phosphate and residual P (16.9% and 9.3%, respectively), compared to the CK (10.3% and 5.9%, respectively; Fig. 5). Moreover, its metabolite P proportions were lower under the DW compared to the CK (11.6% vs 19.2%). In June, T. glabrifolium increased the proportions of inorganic phosphate (15.8%) but decreased metabolite P proportions (8.1%) under the DW treatment compared to the CK (12.5% and 14.7%, respectively). Meanwhile, the residual P proportion of T. glabrifolium significantly decreased under the WW treatment compared to the CK treatment (p = 0.026). In September, proportions of inorganic phosphate and residual P in C. brachiate were significantly reduced under the DW treatment compared to the CK (p = 0.006 and 0.033, respectively), whereas T. glabrifolium displayed elevated metabolic P proportions under the DW treatment (p = 0.001). The proportions of inorganic phosphate in C. brachiate and residual P in S. heptaphylla were also higher under the WW treatment than the DW treatment (p = 0.013 and 0.016, respectively), while T. glabrifolium’s metabolic P proportion was significantly lower under the WW treatment than the DW treatment (p = 0.002). In November, T. glabrifolium’s residual P proportion under the DW treatment (2.8%) decreased compared to the CK (6.1%), while both S. poilanei and S. heptaphylla showed an increased residual P proportion under the DW treatment (3.1% and 1.6% in DW, compared to 1.5% and 0.8% in CK, respectively). Under the WW treatment, S. poilanei displayed a higher residual P proportion than the CK (3.1% vs 1.5%), and T. glabrifolium showed a higher residual P proportion than the DW treatment (6.8% vs 2.8%). S. heptaphylla exhibited a lower residual P proportion under the WW treatment than the DW treatment (p = 0.001).

Relative proportion of inorganic phosphate and organic phosphorus fractions to total phosphorus concentration in leaves for C. brachiata (CB), S. poilanei (SP), S. heptaphylla (SH), and T. glabrifolium (TG) under the natural control (CK), delayed wet season (DW), and wetter wet season (WW) treatments from April to November (n = 4 or 5)
Precipitation treatments induced significant changes in photosynthetic characteristics and nitrogen and P use efficiency (PNUE and PPUE) across the four species (Table 3). In April, T. glabrifolium’s Aarea was higher under the WW treatment than the DW treatment and CK (p = 0.009 and 0.010, respectively). S. poilanei showed higher Amass and PNUE values under the WW treatment than the CK (p = 0.042 and 0.002, respectively). In June, S. poilanei’s Aarea and Amass were decreased by the DW treatment (p = 0.021 and 0.036, respectively), whereas C. brachiate’s Aarea was lower under both the DW and WW treatment than the CK (p = 0.032 and 0.002, respectively). But in September, the WW treatment led to higher values of Aarea, Amass, PNUE, and PPUE for S. poilanei's, as well as elevated levels of Aarea, Amass, and PPUE for S. heptaphylla’s, when compared to the CK. In November, both the DW and WW treatments increased Aarea values of S. heptaphylla (p = 0.010 and 0.043, respectively), and its Amass was also higher in the DW treatment compared to the CK (p = 0.047). Moreover, S. poilanei’s PNUE and PPUE under the WW exceeded the DW treatment and CK (p-values for PNUE were 0.004 and 0.002, and for PPUE 0.016 and 0.015, respectively), as did S. heptaphylla’s PNUE under the WW treatment compared to the CK (p = 0.044).
Relationships between soil and foliar characteristics and phosphorus fractions
In the 0–10 cm soil layer, NaHCO3 Pi concentration was negatively correlated with concentrations of NaHCO3 Po, DHCl Pi, residual Pt, total P and N, and SOC (R = -0.7, 0.65, -0.68, -0.65, -0.62, and -0.8, respectively), while NaHCO3 Po concentration was positively correlated with SOC and total N in the soil (R = 0.67 and 0.69, respectively; Fig. 6). DHCl Pi was strongly correlated with residual Pt and total P (R = 0.7 and 0.81, respectively), while concurrently, a positive correlation was found between residual Pt and soil pH (R = 0.6). NaHCO3 Pi was positively correlated with all foliar P fractions except for residual P (R = 0.58, 0.86, 0.7, 0.88, and 0.89, for phosphate, lipid P, metabolite P, nucleic acid P, and total P, respectively). Additionally, NaHCO3 Po, DHCl Pi, residual Pt, and total P in the soil were negatively correlated with foliar lipid P, nucleic acid P, and total P, while soil residual Pt and total P had positive correlations with foliar residual P. Furthermore, PPUE was negatively correlated with NaHCO3 Pi, lipid P, metabolite P, and foliar total P (R = -0.66, -0.83, -0.68, -0.74, respectively), but was positively correlated with DHCl Pi, soil total P, Aarea, Amass and PNUE (R = 0.72, 0.59, 0.63, 0.68, 0.69, respectively).

Pearson correlations between soil and foliar properties under the natural control (CK), delayed wet season (DW), and wetter wet season (WW) treatments. Note: The data were collected from soil P fractions and soil properties in both soil layers (n = 4), foliar P fractions, as well as photosynthetic and chemical characteristics of four species (n = 4 or 5) under the CK, DW, and WW treatments from April to November. Each repetition represents the average of every soil layer per treatment per month for soil in 0–10 cm or 10–20 cm, and the mean of four species in each treatment per month for foliar. Colors correspond to the strength of the correlations (decrease/increase of color intensity = loss of or gain of correlation). ***, **, and * indicate significance at 0.001, 0.01, and 0.05 levels, respectively
In the 10–20 cm soil layer, NaHCO3 Pi was negatively correlated with DHCl Pi, residual Pt, and total P (R = -0.65, -0.66, and -0.66, respectively), but positively correlated with NH4+-N (R = 0.71; Fig. 6). Resin Pi showed a similar pattern as NaHCO3 Pi. It was negatively correlated with soil residual Pt (R = -0.63) and positively correlated with NH4+-N (R = 0.81). Soil total P was positively correlated with NaOH Po, DHCl Pi, and residual Pt (R = 0.61, 0.66, and 0.98, respectively). The relationship between NaHCO3 Pi and foliar attributes largely paralleled the trend observed in the 0–10 cm soil layer. Resin Pi was positively correlated with foliar total C, metabolite P, nucleic acid P, and total P (R = 0.62, 0.64, 0.58, and 0.61, respectively). Conversely, DHCl Pi, residual Pt, and soil total P were negatively correlated with lipid P, nucleic acid P, and total foliar P. Foliar C, lipid P, nucleic acid P, and total foliar P exhibited positive correlations with NH4+-N, while soil pH was negatively correlated with foliar metabolite P but positively correlated with foliar residual P. Among the measured foliar variables, foliar total P had positive correlations with all foliar P fractions except residual P. Specifically, foliar inorganic phosphate was positively correlated with lipid P and nucleic acid P, and lipid P was positively correlated with metabolite P and nucleic acid P. In addition, PPUE showed positive correlations with NaOH Po, DHCl Pi, soil total P and total N, Aarea, Amass, and PNUE (R = 0.64, 0.79, 0.71, 0.63, 0.63, 0.68, and 0.69, respectively), while exhibited negative correlations with lipid P, metabolite P and foliar total P (R = -0.83, -0.68, and -0.74, respectively).
Piecewise SEM was performed to further reveal the direct and indirect pathway factors in photosynthesis affected by precipitation treatment (Fig. 7). Considering the random effects of “species and months”, precipitation treatment, soil properties, and P fractions at both 0–10 cm and 10–20 cm depths, along with foliar P fractions and photosynthetic nutrient use efficiency, uniformly explained 69% of the photosynthesis variation. Besides foliar P fractions and nutrient use efficiency, precipitation treatment mainly influenced photosynthesis indirectly through changes in soil NH4+-N, NO3−-N, pH, and soil P fractions. Furthermore, to concisely summarize the key findings in this study, we depicted the primary responses of the observed indicators (Fig. 8).

Structural equation model (SEM) showing how altered precipitation patterns affect foliar photosynthetic properties. piecewiseSEM evaluates direct and indirect effects of precipitation patterns, soil traits, and soil P fractions ((a) is data from the 0–10 cm soil layer, and (b) from the 10–20 cm layer), photosynthetic nutrient efficiency, and leaf P fractions on photosynthesis (n = 232). Numbers by arrows denote path coefficients (partial regression), indicating standardized effect size. Solid arrows signify paths significant at p < 0.05, while dashed lines represent non-significant ones (p ≥ 0.05). The conditional (C) and marginal (M) R2 show variance explained by predictors, excluding or including random effects of ‘species and months’. Significance: *p < 0.05, **p < 0.01, ***p < 0.001

Conceptual framework of the impacts of altered precipitation patterns (the natural control: CK, delayed wet season: DW, and wetter wet season: WW) on the dynamics of soil and foliar P fractions and their respective characteristics. Upward arrows and downward arrows denote indicators significantly increased and decreased relative to CK, respectively. Indicators with a significance greater than 0.05 compared with CK are not shown in the figure
Discussion
This study unraveled the intricate connections between changes in seasonal precipitation patterns and nutrient cycling in tropical forests, focusing on soil P forms and bioavailability, and their interactions with foliar traits. We discovered that soil P fractions, more than soil total P, strongly reacted to changes in precipitation in a tropical secondary forest. Notably, delaying the wet season by two months significantly decreased soil P availability, altering the concentrations and proportions of various soil P forms. Conversely, a 25% wet season rainfall increase only slightly affected soil P forms but raised soil pH and NH4+-N concentration. These precipitation changes had variable effects on soil P forms over time, mirroring the diverse responses within the irregular foliar P fractions of the four target tree species. Amid these, a significant drop in foliar total P was observed for DW treatment, leading to diverse foliar P fractions changes within species. Concurrently, the DW treatment also led to a reduction in Amass and/or Aarea, while the WW treatment variably elevated Amass and/or Aarea, with these effects showing species-specific patterns. Precipitation changes also indirectly affected photosynthesis by altering soil properties and P fractions. Both Pearson’s correlation coefficient and redundancy analysis underscored a strong link between plant-available soil P fractions and foliar P fractions.
Effects of altered seasonal precipitation on soil P fractions
Previous studies have extensively documented the substantial impact of variances in soil water conditions on the alteration of soil P forms (Helfenstein et al. 2018; Sun et al. 2020; Wood et al. 2016). Nevertheless, contradictory evidence suggested that a reduction of approximately 20.6% in soil water availability does not necessarily induce significant changes in the levels of soil available P in Mediterranean shrubland (Sardans et al. 2006). In our study, we found that altered precipitation treatments did not significantly affect total P concentration in the soil. However, they did lead to noticeable changes in P forms, and these changes varied across sampling times throughout the experiment (Fig. 2). For instance, after intercepting 60% of the precipitation for two months (April–May) within the DW treatment, a significant decrease in NaHCO3 Po concentration was observed within the 10–20 cm soil layers in June. Interestingly, the DW treatment simultaneously led to a significant increase in NaHCO3 Pi concentration for the 10–20 cm soil layer. Fan et al. (2021) likewise confirmed that long-term rainfall exclusion significantly reduced the concentration of total extractable Po, which was associated with reductions in plant roots and soil microbial biomass. Similarly, four-year field drought experiments reported a notable elevation in NaOH Pi concentration (Zhang et al. 2020a). Additionally, our study also observed that in November, after the DW plots were reirrigated with 60% of the intercepted total precipitation, the NaOH Pi levels in the top 10 cm soil were significantly lower under the DW treatment than the CK. Furthermore, in April, the DW treatment led to a significant decrease in DHCl Pi concentration in the 0–10 cm soil layer. Previous research revealed that the stability of HCl-extractable P, influenced by its long residence time and low turnover, varies with soil pH. It tends to be stable in high-pH soils but less so in acidic ones (Helfenstein et al. 2020; Hou et al. 2018b; Zhang et al. 2020a). This could partially elucidate the significant alterations in DHCl Pi observed within the DW treatment. Moreover, similar to the findings by Fan et al. (2021), our study observed a pervasive decline tendency in resin Pi across almost every sampling and soil layer in the DW plots relative to the CK plots, with a marginally significant (p = 0.055) difference evidenced in September.
Several studies reported that increased rainfall intensity directly affects the runoff of P, leading to a substantial rise in total P loss (Shigaki et al. 2007; Zhang et al. 2019). An experimental study in temperate forests found that a 30% increase in annual precipitation negatively affects primary mineral P but enhances secondary minerals (Zhang et al. 2020b). This increase notably elevated NaHCO3 Pi, NaOH Pi, and total P concentration, while notably decreasing DHCl Pi. Furthermore, based on the precipitation platform of our study, Sun et al. (2020) found that the concentrations of NaOH Po, residual Pt, and total Po sampled in August in the WW plots are significantly elevated compared to those in the CK plots. Yet, in this study, we found that the WW treatment did not significantly modify the concentration of soil P fractions, except for a notable decrease in NaHCO3 Po within the 10–20 cm soil layer in April. These findings highlight the intricate interplay between the P cycle and shifts in precipitation patterns, a dynamic that can be further complicated by the timing of sample collection, potentially leading to variations in observations. Therefore, conducting additional long-term observational studies is imperative for achieving a comprehensive understanding of these dynamics. Moreover, we observed that the WW treatment led to a higher soil pH value. This elevation in pH was likely attributed to the dilution and leaching of topsoil salts, where deep percolation facilitated salt removal, simultaneously decreased the concentration of exchangeable H+ ions in the soil solution, leading to an increase in pH level (Rengel 2002, 2011).
Effect of altered seasonal precipitation on the foliar P allocation and photosynthesis
In synchronization with the soil P fraction concentration, the DW treatment variably reduced foliar total P concentration across different samples, with these reductions reflected in varying P fraction concentrations of certain plant species (Figs. 4 and 8). This aligns with observations by Gao et al. (2022a) and Hidaka and Kitayama (2011), indicating a general decline in plant foliar P concentration as soil P availability decreases. However, it is important to note that these changes exhibit significant variability, depending on both the species and the season. This was confirmed by Liu et al. (2023) who reported species-specific and seasonal differences in foliar P allocation across 18 species from three families during wet and dry seasons in Western Australia. Additionally, Yan et al. (2019) reported a strong positive correlation between soil pH and leaf P concentrations in two dune plant species from Western Australia. In our study, soil pH values were much higher in the WW plots than in the DW and CK plots in June and September, yet there was no observed enhancement in the corresponding leaf P concentration for all studied species. Conversely, in November, a higher total leaf P concentration in S. heptaphyla under the WW treatment was observed. Moreover, Mo et al. (2015, 2019) documented that the understory species within the same study area exhibited relatively high leaf N:P ratios (ranging from 10 to 40). In contrast, the N:P ratios of the investigated four canopy species in this study were relatively low with the values not exceeding 15.
Different P fractions serve distinct roles in leaves and are closely related to life history strategies including growth, reproduction, and stress resilience (Hidaka and Kitayama 2011). For example, lipid, integral to the structure of cell membranes, may undergo hydrolysis or be replaced by non-phosphorylated counterparts under P scarcity (Lambers et al. 2012; Shimojima et al. 2013). Nevertheless, exceptions have been documented, as demonstrated by Liang et al. (2023), where certain Myrtaceae species displayed low lipid P without a corresponding increase in P-free lipids. In our study, across most sampling times and species, lipid P constituted the majority of all P fractions (ranging from 32.4% to 56.7%), followed by nucleic acid P (Fig. 5). These findings indicated that the primary expenditure of P inputs lies in essential 'infrastructure' within leaves of the target species, such as membranes and other cellular structures. Previous research suggests that nucleic acid P is the predominant fraction (Chapin III and Kedrowski 1983; Veneklaas et al. 2012), especially at low foliar P levels (P < 1 mg g−1) (Crous and Ellsworth 2020). Yet, the proportions of specific P fractions in leaves and soil vary by species and soil conditions (Yan et al. 2019; Mo et al. 2019). Gao et al. (2022b) further corroborated that interspecies differences yield divergent foliar P fractions. Moreover, metabolomics data show that optimized lipid P and nucleic acid P reserves enhance P utilization efficiency in maize leaves (Han et al. 2022). This could potentially explain our consistent observations of high levels of lipid P and nucleic acid P. Chapin III and Kedrowski (1983) did not identify significant seasonal variations in leaf P fractions between deciduous and evergreen trees. In our study, we observed a similar distribution of P fraction in the deciduous T. glabrifolium and the three evergreen species. Previous studies indicated higher total P and foliar P fractions in annuals than perennials, potentially due to enhanced metabolic activity or luxury consumption (Suriyagoda et al. 2022). Similarly, T. glabrifolium showed higher P levels compared to evergreen species and displayed consistent P fraction distribution patterns across species. Consequently, in our observation, the foliar P concentration and distribution of foliar P fractions were influenced by soil P fraction levels and treatment conditions. This influence pattern varied depending on the plant species and sampling times. The plasticity of P allocation to its fractions plays a crucial role in adapting to limited soil P availability, while species-specific P allocation is essential for coexisting with other species (Suriyagoda et al. 2022). Therefore, our findings suggested that such strategic acclimations in P allocation might represent survival mechanisms employed by these tropical tree species under conditions of varying soil P availability.
Although there are frequent reports of a strong correlation between foliar P concentration and photosynthetic traits (Hidaka and Kitayama 2009, 2013), this association is not consistently observed across different studies (Mo et al. 2019). Here, we found that the DW treatment not only reduced foliar P concentration but also decreased photosynthetic rates, particularly the Aarea and Amass for S. poilanei and Aarea for C. brachiata in June, compared to the CK (Table 3). However, we did not find that any specific foliar P fractions were associated with these decreased Aarea or Amass. There were also some exceptions to this trend. In the case of the DW treatment, it led to significant elevations in Amass for S. poilanei in September and both Amass and Aarea for S. heptaphyla in November relative to the CK. Conversely, although the WW treatment didn’t significantly boost plant P availability, it variably enhanced photosynthetic traits. Specifically, under the WW treatment, the Aarea for T. glabrifolium in April, both Aarea and Amass for S. heptaphyla and S. poilanei in September, as well as Aarea for S. heptaphyla in November, were significantly elevated compared to those of CK. Previous studies have shown that increasing PPUE is an effective acclimation strategy for trees, especially for tropical tree species growing in P-poor soil (Hidaka and Kitayama 2009). Thus, these increases in photosynthetic traits may be linked to rises in PPUE, particularly for S. poilanei species, as the WW treatment in September significantly increased PPUE while increasing Aarea and Amass. This is further confirmed by the results of our structural equation model (Fig. 7). Furthermore, the SEM results indicate that these photosynthetic traits are influenced not only directly by photosynthetic nutrient use efficiency and foliar P fractions but also indirectly by soil properties and soil P fractions modulated by precipitation treatments.
Correlations among foliar and soil phosphorus fractions and their respective characteristics
While numerous studies have validated the correlation between plant P concentration and soil P fractions, results vary for specific P forms (Galván-Tejada et al. 2014; Niederberger et al. 2019). For instance, Gao et al. (2022a) found that labile P forms like resin Pi and NaHCO3 Pi are positively correlated with foliar P fractions, whereas Yan et al. (2019) reported resin P and foliar total P are unrelated. Additionally, the influence of various soil P forms on foliar P fractions is species-dependent, notably affecting Alhagi sparsifolia through active Pi and Po, and Karelinia caspia through active Pi coupled with enzyme activity (Gao et al. 2022b). Generally, our study revealed positive relationships of soil resin Pi and NaHCO3 Pi concentrations with most foliar P fractions (except for residual P, which exhibits a negative correlation) and total foliar P (Fig. 6). However, soil DHCl Pi and residual Pt concentration display an inverse trend. These findings underscore the importance of soil resin Pi and NaHCO3 P as vital plant accessible P sources. While our investigation explored the general correlations between soil and foliar P fractions in tropical forests, it did not differentiate these relationships on a species-specific basis (see Fig. S2 for detailed species correlations). Interestingly, partly supporting the findings of Yan et al. (2019), the correlation between soil resin Pi and foliar total P concentration, as well as various foliar P fractions in our study, was indistinct in the 0–10 cm soil layer but significantly apparent in the 10–20 cm soil layer. Thus, it is necessary to consider soil layer variations when evaluating soil nutrient properties in association with plant nutrient characteristics.
Available soil nitrogen such as NH4+-N and NO3−-N could exert substantial influences on soil P availability (Zhang et al. 2004). In contrast to the findings of Zhang et al. (2004) who reported a decline in inorganic P fractions within the root zone following NH4+-N fertilizer application, our study revealed more complex relationships between soil P fractions and available N. Specifically, we observed that the WW treatment significantly and linearly increased pH and NH4+-N concentration in both soil layers, and the elevated NH4+-N concentration was positively correlated with both resin Pi and NaHCO3 Pi concentration, although this correlation was exclusive to the 10–20 cm soil layer. This suggests that their interrelation may not follow a purely linear pattern, warranting further comprehensive exploration. Soil pH is crucial for plant available P, yet its effects are mediated by both plant traits and soil factors (Lambers 2021). Hou et al. (2018b) emphasized the substantial impact of pH on soil Po forms, while other studies noted that decreasing pH increases both root P uptake and soil P sorption, creating a counterbalance that limits root P acquisition (Lambers 2021; Smyth and Sanchez 1980). Our findings provided further insight into this complexity, revealing a significant positive correlation between pH and residual P concentration in the foliage and top 0–10 cm soil layer. Concurrently, pH value in the 10–20 cm layer was positively correlated with foliar residual P but inversely correlated with foliar metabolite P concentration. Hidaka and Kitayama (2013) reported a positive correlation between Amass and both total foliar P and metabolic P concentrations. Contrary to their findings, our study did not detect significant correlations between Amass and any foliar P fractions. Previous studies have indicated that under low-P conditions, plants can maintain their photosynthetic capacity by reducing the lipid P allocation to enhance the PPUE (Hayes et al. 2022). In this study, we observed a negative correlation between the PPUE and both lipid P and metabolite P, along with foliar total P. Contrary to expectations of stable photosynthetic capacity, we observed variations in photosynthesis.
Conclusions
In this study, we investigated how simulated changes in seasonal precipitation affected soil and leaf phosphorus (P) fractions in a tropical secondary forest. Unexpectedly, soil total P remained unaffected by precipitation treatments, whereas P forms and bioavailability were highly sensitive, particularly under delayed wet season (DW) treatment that reduced soil P bioavailability. Yet, wetter wet season (WW) treatment insignificantly altered soil P form but increased soil pH and NH4+-N. We observed that the effects of precipitation treatments on soil and foliar P fractions, as well as photosynthetic traits (Amass and Aarea), varied by species and sampling time. The structural equation model (SEM) indicated that photosynthetic attributes were modulated by nutrient use efficiency and foliar P fractions, with an indirect influence from soil parameters affected by precipitation. Significant correlations were noted between plant-accessible soil P and foliar P fractions. Overall, our study underscored the intricate linkage between changing precipitation patterns and nutrient cycling in tropical forest ecosystems. In conclusion, this study contributes to the advancement of our knowledge to guide forest conservation and restoration strategies under climate change scenarios, ensuring the sustained provision of ecosystem services in tropical regions.
Data availability
The data generated and/or analyzed during the present study are accessible from the corresponding author upon reasonable request.
References
Afreen T, Singh H, Singh JS (2019) Influence of changing patterns of precipitation and temperature on tropical soil ecosystem. In: Garkoti SC, Van Bloem SJ, Fulé PZ, Semwal RL (eds) Tropical ecosystems: structure, functions and challenges in the face of global change. Springer, pp 11–26
Aguirre-Gutiérrez J, Berenguer E, Oliveras Menor I, Bauman D, Corral-Rivas JJ, Nava-Miranda MG, Both S, Ndong JE, Evouna Ondo F, N’ssi Bengone N, Mihinhou V, Dalling JW, Heineman K, Figueiredo A, González-M R, Norden N, Hurtado-M AB, González D, Salgado-Negret B, Reis SM, Moraes de Seixas MM, Farfan-Rios W, Shenkin A, Riutta T, Girardin CAJ, Moore S, Abernethy K, Asner GP, Bentley LP, Burslem DFRP, Cernusak LA, Enquist BJ, Ewers RM, Ferreira J, Jeffery KJ, Joly CA, Marimon-Junior BH, Martin RE, Morandi PS, Phillips OL, Bennett AC, Lewis SL, Quesada CA, Schwantes Marimon B, Kissling WD, Silman M, Teh YA, White LJT, Salinas N, Coomes DA, Barlow J, Adu-Bredu S, Malhi Y (2022) Functional susceptibility of tropical forests to climate change. Nat Ecol Evol 6:878–889. https://doi.org/10.1038/s41559-022-01747-6
Ames BN (1966) Assay of inorganic phosphate, total phosphate and phosphatases. Methods Enzymol 8:115–118. https://doi.org/10.1016/0076-6879(66)08014-5
Bates D, Maechler M, Bolker B, Walker S (2017) lme4: linear mixed-effects models using Eigen and S4. R package version 1.1–34
Bradshaw CJA, Sodhi NS, Brook BW (2009) Tropical turmoil: a biodiversity tragedy in progress. Front Ecol Environ 7(2):79–87. https://doi.org/10.1890/070193
Chapin FS III, Kedrowski RA (1983) Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 64(2):376–391. https://doi.org/10.2307/1937083
Cheng Y, Zhang H, Zang R, Wang X, Long W, Wang X, Xiong M, John R (2020) The effects of soil phosphorus on aboveground biomass are mediated by functional diversity in a tropical cloud forest. Plant Soil 449(2):51–63. https://doi.org/10.1007/s11104-020-04421-7
Crous KY, Ellsworth DS (2020) Probing the inner sanctum of leaf phosphorus: measuring the fractions of leaf P. Plant Soil 454:77–85. https://doi.org/10.1007/s11104-020-04657-3
Deb JC, Phinn S, Butt N, McAlpine CA (2018) Climate change impacts on tropical forests: identifying risks for tropical Asia. J Trop For Sci 182–194. https://doi.org/10.26525/jtfs2018.30.2.182194
Dietrich K, Spohn M, Villamagua M, Oelmann Y (2017) Nutrient addition affects net and gross mineralization of phosphorus in the organic layer of a tropical montane forest. Biogeochemistry 136:223–236. https://doi.org/10.1007/s10533-017-0392-z
Dirzo R, Raven PH (2003) Global state of biodiversity and loss. Annu Rev Environ Resour 28(1):137–167. https://doi.org/10.1146/annurev.energy.28.050302.105532
Dore MH (2005) Climate change and changes in global precipitation patterns: what do we know? Environ Int 31(8):1167–1181. https://doi.org/10.1016/j.envint.2005.03.004
Dorich RA, Nelson DW (1983) Direct colorimetric measurement of ammonium in potassium chloride extracts of soils. Soil Sci Soc Am J 47(4):833–836. https://doi.org/10.2136/sssaj1983.03615995004700040042x
Fan Y, Lu S, He M, Yang L, Hu W, Yang Z, Liu X, Hui D, Guo J, Yang Y (2021) Long-term throughfall exclusion decreases soil organic phosphorus associated with reduced plant roots and soil microbial biomass in a subtropical forest. Geoderma 404:115309. https://doi.org/10.1016/j.geoderma.2021.115309
Feng X, Porporato A, Rodriguez-Iturbe I (2013) Changes in rainfall seasonality in the tropics. Nat Clim Chang 3(9):811–815. https://doi.org/10.1038/nclimate1907
Galván-Tejada NC, Peña-Ramírez V, Mora-Palomino L, Siebe C (2014) Soil P fractions in a volcanic soil chronosequence of Central Mexico and their relationship to foliar P in pine trees. Eur J Soil Sci 177(5):792–802. https://doi.org/10.1002/jpln.201300653
Gao Y, Tariq A, Zeng F, Graciano C, Zhang Z, Sardans J, Peñuelas J (2022a) Allocation of foliar-P fractions of Alhagi sparsifolia and its relationship with soil-P fractions and soil properties in a hyperarid desert ecosystem. Geoderma 407:115546. https://doi.org/10.1016/j.geoderma.2021.115546
Gao Y, Zeng F, Islam W, Zhang Z, Du Y, Zhang Y, Chai X, Liu Y (2022b) Coexistence desert plants respond to soil phosphorus availability by altering the allocation patterns of foliar phosphorus fractions and acquiring different forms of soil phosphorus. J Plant Growth Regul 42(6):3770–3784. https://doi.org/10.1007/s00344-022-10836-6
Hajek OL, Knapp AK (2022) Shifting seasonal patterns of water availability: ecosystem responses to an unappreciated dimension of climate change. New Phytol 233(1):119–125. https://doi.org/10.1111/nph.17728
Han Y, Hong W, Xiong C, Lambers H, Sun Y, Xu Z, Schulze WX, Cheng L (2022) Combining analyses of metabolite profiles and phosphorus fractions to explore high phosphorus utilization efficiency in maize. J Exp Bot 73(12):4184–4203. https://doi.org/10.1093/jxb/erac117
Hayes PE, Adem GD, Pariasca-Tanaka J, Wissuwa M (2022) Leaf phosphorus fractionation in rice to understand internal phosphorus-use efficiency. Ann Bot 129(3):287–302. https://doi.org/10.1093/aob/mcab138
Hedley MJ, Stewart JWB, Chauhan BS (1982) Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Sci Soc Am J 46(5):970–976. https://doi.org/10.2136/sssaj1982.03615995004600050017x
Helfenstein J, Tamburini F, von Sperber C, Massey MS, Pistocchi C, Chadwick OA, Vitousek PM, Kretzschmar R, Frossard E (2018) Combining spectroscopic and isotopic techniques gives a dynamic view of phosphorus cycling in soil. Nat Commun 9(1):3226. https://doi.org/10.1038/s41467-018-05731-2
Hidaka A, Kitayama K (2009) Divergent patterns of photosynthetic phosphorus-use efficiency versus nitrogen-use efficiency of tree leaves along nutrient-availability gradients. J Ecol 97(5):984–991. https://doi.org/10.1111/j.1365-2745.2009.01540.x
Hidaka A, Kitayama K (2011) Allocation of foliar phosphorus fractions and leaf traits of tropical tree species in response to decreased soil phosphorus availability on Mount Kinabalu. Borneo J Ecol 99(3):849–857. https://doi.org/10.1111/j.1365-2745.2011.01805.x
Hidaka A, Kitayama K (2013) Relationship between photosynthetic phosphorus-use efficiency and foliar phosphorus fractions in tropical tree species. Ecol Evol 3(15):4872–4880. https://doi.org/10.1002/ece3.861
Hou E, Chen C, Luo Y, Zhou G, Kuang Y, Zhang Y, Heenan M, Lu X, Wen D (2018a) Effects of climate on soil phosphorus cycle and availability in natural terrestrial ecosystems. Glob Change Biol 24(8):3344–3356. https://doi.org/10.1111/gcb.14093
Hou E, Wen D, Kuang Y, Cong J, Chen C, He X, Heenan M, Lu H, Zhang Y (2018b) Soil pH predominantly controls the forms of organic phosphorus in topsoils under natural broadleaved forests along a 2500 km latitudinal gradient. Geoderma 315:65–74. https://doi.org/10.1016/j.geoderma.2017.11.041
IPCC (2023) Synthesis Report of the IPCC Sixth Assessment Report (AR6). In: Core Writing Team, Pachauri RK and Meyer LA (eds) Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva
Korell L, Auge H, Chase JM, Harpole WS, Knight TM (2021) Responses of plant diversity to precipitation change are strongest at local spatial scales and in drylands. Nat Commun 12(1):2489. https://doi.org/10.1038/s41467-021-22766-0
Lambers H (2021) Phosphorus acquisition and utilization in plants. Annu Rev Plant Biol 73:17–42. https://doi.org/10.1146/annurev-arplant-102720-125738
Lambers H, Cawthray GR, Giavalisco P, Kuo J, Laliberte E, Hulshof CM, Pearse SJ, Scheible WR, Stitt M, Teste F, Turner BL (2012) Proteaceae from severely phosphorus-impoverished soils extensively replace phospholipids with galactolipids and sulfolipids during leaf development to achieve a high photosynthetic phosphorus-use-efficiency. New Phytol 196(4):1098–1108. https://doi.org/10.1111/j.1469-8137.2012.04285.x
Lefcheck JS (2015) PiecewiseSEM: piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol Evol 7:573–579. https://doi.org/10.48550/arXiv.1509.01845
Liang GH, Butler OM, Warren CR (2023) Lipid profiles of plants and soil microbial communities are shaped by soil parent material in Australian sclerophyll forests. Plant Soil. https://doi.org/10.1007/s11104-023-06075-7
Liu GS, Jiang NH, Zhang LD, Liu ZL (1996) Soil physical and chemical analysis and description of soil profiles. Standards Press of China, Beijing [in Chinese]
Liu ST, Gille CE, Bird T, Ranathunge K, Finnegan PM, Lambers H (2023) Leaf phosphorus allocation to chemical fractions and its seasonal variation in south-western Australia is a species-dependent trait. Sci Total Environ 901:166395. https://doi.org/10.1016/j.scitotenv.2023.166395
Luo Y, Liu S, Zhou G (2008) Trends of precipitation in Beijiang River Basin, Guangdong Province, China. Hydrol Process 22(13):2377–2386. https://doi.org/10.1002/hyp.6801
Maranguit D, Guillaume T, Kuzyakov Y (2017) Land-use change affects phosphorus fractions in highly weathered tropical soils. CATENA 149:385–393. https://doi.org/10.1016/j.catena.2016.10.010
Mo Q, Zou B, Li Y, Chen Y, Zhang W, Mao R, Ding Y, Wang J, Lu X, Li X, Li Z, Wang F (2015) Response of plant nutrient stoichiometry to fertilization varied with plant tissues in a tropical forest. Sci Rep 5:14605. https://doi.org/10.1038/srep14605
Mo Q, Li Z, Sayer EJ, Lambers H, Li Y, Zou B, Tang J, Heskel M, Ding Y, Wang F (2019) Foliar phosphorus fractions reveal how tropical plants maintain photosynthetic rates despite low soil phosphorus availability. Funct Ecol 33(3):503–513. https://doi.org/10.1111/1365-2435.13252
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36. https://doi.org/10.1016/S0003-2670(00)88444-5
Mwende Muindi E (2019) Understanding soil phosphorus. Int J Plant Soil Sci 31(2):1–18. https://doi.org/10.9734/ijpss/2019/v31i230208
Niederberger J, Kohler M, Bauhus J (2019) Distribution of phosphorus fractions with different plant availability in German forest soils and their relationship with common soil properties and foliar P contents. Soil 5(2):189–204. https://doi.org/10.5194/soil-5-189-2019
Niederberger J, Kohler M, Bauhus J (2017) The relevance of different soil phosphorus fractions for short-term tree nutrition: results from a mesocosm bioassay. For: Int J For Res 90(2):258–267. https://doi.org/10.1093/forestry/cpw050
Norman RJ, Edberg JC, Stucki JW (1985) Determination of nitrate in soil extracts by dual-wavelength ultraviolet spectrophotometry. Soil Sci Soc Am J 49(5):1182–1185. https://doi.org/10.2136/sssaj1985.03615995004900050022x
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2022) Vegan: community ecology package. R Package Version 2.6–4. https://rdocumentation.org/packages/vegan/versions/2.6-4. Accessed 30 Jun 2023
Ren H, Li Z, Shen W, Yu Z, Peng S, Liao C, Ding M, Wu J (2007) Changes in biodiversity and ecosystem function during the restoration of a tropical forest in south China. Sci China Ser C Life Sci 50(2):277–284. https://doi.org/10.1007/s11427-007-0028-y
Rengel Z (2002) Role of pH in availability of ions in soil. In: Rengel Z (ed) Handbook of plant growth. pH as a master variable in plant growth. Marcel Dekker, New York, pp 323–350
Rengel Z (2011) Soil pH, soil health and climate change. In: Singh BP, Cowie AL, Chan KY (eds) Soil health and climate change. Soil Biology, vol 29. Springer, Berlin, Heidelberg
Sardans J, Peñuelas J, Estiarte M (2006) Warming and drought alter soil phosphatase activity and soil P availability in a Mediterranean shrubland. Plant Soil 289(1–2):227–238. https://doi.org/10.1007/s11104-006-9131-2
Shen J, Yuan L, Zhang J, Li H, Bai Z, Chen X, Zhang W, Zhang F (2011) Phosphorus dynamics: from soil to plant. Plant Physiol 156(3):997–1005. https://doi.org/10.1104/pp.111.175232
Shigaki F, Sharpley A, Prochnow LI (2007) Rainfall intensity and phosphorus source effects on phosphorus transport in surface runoff from soil trays. Sci Total Environ 373(1):334–343. https://doi.org/10.1016/j.scitotenv.2006.10.048
Shimojima M, Watanabe T, Madoka Y, Koizumi R, Yamamoto MP, Masuda K, Yamada K, Masuda S, Ohta H (2013) Differential regulation of two types of monogalactosyldiacylglycerol synthase in membrane lipid remodeling under phosphate-limited conditions in sesame plants. Front Plant Sci 4:469. https://doi.org/10.3389/fpls.2013.00469
Smith C, Baker JCA, Spracklen DV (2023) Tropical deforestation causes large reductions in observed precipitation. Nature 615(7951):270–275. https://doi.org/10.1038/s41586-022-05690-1
Smyth TJ, Sanchez PA (1980) Effects of lime, silicate, and phosphorus applications to an oxisol on phosphorus sorption and ion retention. Soil Sci Soc Am J 44(3):500–505. https://doi.org/10.2136/sssaj1980.03615995004400030017x
Song Y, Achberger C, Linderholm HW (2011) Rain-season trends in precipitation and their effect in different climate regions of China during 1961–2008. Environ Res Lett 6(3):034025. https://doi.org/10.1088/1748-9326/6/3/034025
Spracklen DV, Baker JCA, Garcia-Carreras L, Marsham JH (2018) The effects of tropical vegetation on rainfall. Annu Rev Environ Resour 43:193–218. https://doi.org/10.1146/annurev-environ-102017-030136
Sun F, Song C, Wang M, Lai DYF, Tariq A, Zeng F, Zhong Q, Wang F, Li Z, Peng C (2020) Long-term increase in rainfall decreases soil organic phosphorus decomposition in tropical forests. Soil Biol Biochem 151:108056. https://doi.org/10.1016/j.soilbio.2020.108056
Suriyagoda LDB, Ryan MH, Gille CE, Dayrell RLC, Finnegan PM, Ranathunge K, Nicol D, Lambers H (2022) Phosphorus fractions in leaves. New Phytol 233(1):5–20. https://doi.org/10.1111/nph.18588
Tiessen H, Moir JO (2007) Characterization of available P by sequential extraction. In: Carter MR, Gregorich EG (eds) Soil sampling and methods of analysis, 2nd edn. CRC Press, Boca Raton, pp 293–306
Tiessen H, Stewart JWB, Cole CV (1984) Pathways of phosphorus transformations in soils of differing pedogenesis. Soil Sci Soc Am J 48(4):853–858. https://doi.org/10.2136/sssaj1984.03615995004800040031x
Tsujii Y, Fan B, Atwell BJ, Lambers H, Lei Z, Wright IJ (2023) A survey of leaf phosphorus fractions and leaf economic traits among 12 co-occurring woody species on phosphorus-impoverished soils. Plant Soil 489:107–124. https://doi.org/10.1007/s11104-023-06001-x
Veneklaas EJ, Lambers H, Bragg J, Finnegan PM, Lovelock CE, Plaxton WC, Price CA, Scheible WR, Shane MW, White PJ, Raven JA (2012) Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol 195:306–320. https://doi.org/10.1111/j.1469-8137.2012.04190.x
Wang J, Sun Z, Hui D, Yang L, Wang F, Liu N, Ren H (2018) Responses of seedling performance to altered seasonal precipitation in a secondary tropical forest, southern China. For Ecol Manag 410:27–34. https://doi.org/10.1016/j.foreco.2017.12.035
Wang C, Xue L, Jiao R (2021) Soil phosphorus fractions, phosphatase activity, and the abundance of phoC and phoD genes vary with planting density in subtropical Chinese fir plantations. Soil Tillage Res. https://doi.org/10.1016/j.still.2021.104946
Wei T, Simko V (2017) R package “corrplot”: visualization of a correlation matrix. https://github.com/taiyun/corrplot. Accessed 3 Jul 2023
Wood TE, Matthews D, Vandecar K, Lawrence D (2016) Short-term variability in labile soil phosphorus is positively related to soil moisture in a humid tropical forest in Puerto Rico. Biogeochemistry 127(1):35–43. https://doi.org/10.1007/s10533-015-0150-z
Yan L, Zhang X, Han Z, Pang J, Lambers H, Finnegan PM (2019) Responses of foliar phosphorus fractions to soil age are diverse along a 2 Myr dune chronosequence. New Phytol 223(3):1621–1633. https://doi.org/10.1111/nph.15910
Yan L, Sunoj VSJ, Short AW, Lambers H, Elsheery NI, Kajita T, Wee AKS, Cao KF (2021) Correlations between allocation to foliar phosphorus fractions and maintenance of photosynthetic integrity in six mangrove populations as affected by chilling. New Phytol 232(6):2267–2282. https://doi.org/10.1111/nph.17770
Yu Q, Ni X, Cheng X, Ma S, Tian D, Zhu B, Zhu J, Ji C, Tang Z, Fang J (2022) Foliar phosphorus allocation and photosynthesis reveal plants’ adaptative strategies to phosphorus limitation in tropical forests at different successional stages. Sci Total Environ 846:157456. https://doi.org/10.1016/j.scitotenv.2022.157456
Zhang A, Zhao X (2022) Changes of precipitation pattern in China: 1961–2010. Theor Appl Climatol 148(3–4):1005–1019. https://doi.org/10.1007/s00704-022-03986-w
Zhang F, Kang S, Zhang J, Zhang R, Li F (2004) Nitrogen fertilization on uptake of soil inorganic phosphorus fractions in the wheat root zone. Soil Sci Soc Am J 68:1890–1895. https://doi.org/10.2136/sssaj2004.1890
Zhang R, Li M, Yuan X, Pan Z (2019) Influence of rainfall intensity and slope on suspended solids and phosphorus losses in runoff. Environ Sci Pollut Res 26:33963–33975. https://doi.org/10.1007/s11356-018-2999-6
Zhang H, Shi L, Lu H, Shao Y (2020a) Drought promotes soil phosphorus transformation and reduces phosphorus bioavailability in a temperate forest. Sci Total Environ 732:139295. https://doi.org/10.1016/j.scitotenv.2020.139295
Zhang H, Shi L, Fu S (2020b) Effects of nitrogen deposition and increased precipitation on soil phosphorus dynamics in a temperate forest. Geoderma 380:114650. https://doi.org/10.1016/j.geoderma.2020.114650
Zhang L, Luo X, Liu N, Zang X, Lambers H, Zhang G, Xiao M, Wen D (2021) Effects of elevated CO2 concentration and nitrogen addition on foliar phosphorus fractions of Mikania micranatha and Chromolaena odorata under low phosphorus availability. Physiol Plant 173:2068–2080. https://doi.org/10.1111/ppl.13555
Zhang J, Feng Y, Maestre FT, Berdugo M, Wang J, Coleine C, Sáez-Sandino T, García-Velázquez L, Singh BK, Delgado-Baquerizo M (2023) Water availability creates global thresholds in multidimensional soil biodiversity and functions. Nat Ecol Evol 7:1002–1011. https://doi.org/10.1038/s41559-023-02071-3
Zhong H, Zhou J, Azmi A, Arruda AJ, Doolette AL, Smernik RJ, Lambers H (2020) Xylomelum occidentale (Proteaceae) accesses relatively mobile soil organic phosphorus without releasing carboxylates. J Ecol 109:246–259. https://doi.org/10.5061/dryad.05qfttdz7
Zhou G, Wei X, Wu Y, Huang Y, Yan J, Zhang D, Zhang Q, Liu J, Meng Z, Wang C, Chu G, Liu S, Tang X, Liu X (2011) Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China. Glob Change Biol 17:3736–3746. https://doi.org/10.1111/j.1365-2486.2011.02499.x
Zhu J, Wu A, Zhou G (2021) Spatial distribution patterns of soil total phosphorus influenced by climatic factors in China’s forest ecosystems. Sci Rep 11:5357. https://doi.org/10.1038/s41598-021-84166-0
Acknowledgements
The authors are indebted to Ms. Chunqing Long for laboratory work, and to Mr. Ruyou Zhong, Kangyu Liang, and Liangdong Ren for their assistance throughout the fieldwork.
Funding
This study was supported by the National Natural Science Foundation of China (32171594), and the Youth Innovation Promotion Association CAS (2019340, 2021347). DH was supported by the US NSF and USDA projects.
Author information
Authors and Affiliations
Contributions
X.H., H.R., and J.W. conceptualized and orchestrated the experimental design. Field experiments were carried out by X.H., K.Y., and J.W. Data analysis was primarily conducted by X.H., who also composed the initial manuscript draft. D.H., F.W., D.Q., Z.L., H.L., H.R., and J.W. provided critical feedback and contributed substantially to the manuscript’s refinement.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no competing interests in the work submitted for publication.
Additional information
Responsible Editor: Roberta L C Dayrell.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
He, X., Hui, D., Wang, F. et al. Dynamics of soil and foliar phosphorus fractions in a secondary tropical forest under altered seasonal precipitation patterns. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06774-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06774-9