
Abstract
Aims
A field experiment was conducted to evaluate the effects of alley cropping systems on microbial activity and soil organic matter (SOM) pools. We hypothesized that enzyme activity and labile pools of SOM are early and sensitive indicators of changes induced by tree introduction in the cropping systems.
Methods
Poplar-alfalfa and alder-gramineous (cereal or ryegrass) associations and their respective control systems (alfalfa and gramineous) were compared in terms of soil carbon (C), nitrogen (N) and water contents, SOM labile pools, NIRS-MIRS spectra and microbial enzyme activity in the topsoil (0–15 cm) for 4 years after tree planting.
Results
After 1 year, tree introduction induced a decrease in soil water content, microbial biomass N and some enzyme activities under alfalfa system. After 4 years, tree introduction resulted in higher soil water contents in both systems (alfalfa and gramineous); higher microbial biomass N and lower C:N in alfalfa-poplar plots compared to control plots. MIRS-NIRS analyses showed a greatest differentiation in SOM quality between alfalfa-based systems.
Conclusions
The effects of temperate agroforestry systems on SOC in the topsoil are relatively weak in the first years after tree introduction. Observed effects were more pronounced in the alfalfa-poplar system, probably due to higher tree growth. Further studies will provide insights into the longer-term effects of these systems on soil functioning.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Agroforestry systems are innovative cropping practices that associate trees with annual and/or perennial herbaceous plants or livestock on the same field. The intentional integration of row trees in herbaceous crops (alley cropping systems) is recognized as an integrated way to increase the sustainability of current agricultural systems and enhance ecosystem services as compared to less diverse cropping systems (Tsonkova et al. 2012; Torralba et al. 2016). It is commonly reported that alley cropping systems increase soil organic carbon (SOC) storage (Cardinael et al. 2018) and thus promote both C sequestration (Nair et al. 2009) and soil fertility components (Rao et al. 1997). The increase of SOC in agroforestry systems is due to a large input of C in the soil through aboveground and belowground deposition (litter, exudates, root turnover).
The tree species used in agroforestry systems have different aerial and root functional traits (litter quality, C:N ratio, proportion of fine roots, etc) that may directly or indirectly alter microbial communities and their associated functions. Tree species depends greatly on climatic zone. Temperate studies are dominated by trees affiliated to Juglans (walnut) or Populus (poplar) genera which were recorded in 55% of publications reviewed by Wolz and DeLucia (2018). These two genera may have different effects on soil properties due to their lower litter C:N ratio and/or higher lignin content. Many studies reported that significant increases in SOC in agroforestry systems may only occur after the systems have been implemented for a long term (Lee and Jose 2003; Oelbermann et al. 2006; Mao et al. 2012; Tian et al. 2013). Thus, more precise, faster, and more reliable measurements than the SOC content might be used to monitor the soil characteristics after introducing trees in cropping systems. These early indicators could be the soil organic matter (SOM) and/or microbial properties. SOM is a continuum of progressively decomposing organic compounds (Lehmann and Kleber 2015), more or less associated with soil minerals, leading to different levels of recalcitrance in relation to decomposition. Different fractions of SOM, described as labile, such as microbial biomass C (MBC), light fraction C, particulate organic matter (POM) and extractable C pools are more sensitive than SOC to changes in land use or soil management strategies both in forestry and agricultural systems (Islam and Weil 2000; Haynes 2000; Shen et al. 2018). The specific composition of this labile pool remains unclear and depends strongly on the extraction methods, but small hydrophilic molecules (e.g. saccharides, amino acids) can be quickly metabolized by microbes (Jones 1998), leading to a MBC increase. Soil microorganisms are involved in both SOM decomposition and stabilization (Six et al. 2004) and are known to synthesize several classes of enzymes that are the proximate agents of SOM decomposition (Sinsabaugh et al. 2008). The synthesis of these enzymes could be modulated by SOM quantity and/or quality and soil nutrient availability, which both affect the stoichiometry of SOM-degrading microbes (Geisseler et al. 2010). Soil microbial activity is more strongly correlated with labile organic carbon pools than with total SOC (Haney et al. 2012), suggesting that these pools can significantly control microbial activity changes on a short-term scale in response to environmental changes due to their accessibility to soil microorganisms (Schimel et al. 2007; Blankinship and Schimel 2018). Enzyme activities are considered as potential indicators of soil quality (Dick et al. 1996) because they are highly sensitive to land use, cropping systems, and agricultural practices such as tillage and fertilization (Klose et al. 1999; Bandick and Dick 1999; Acosta-Martínez et al. 2007; Vong et al. 2007). As enzyme activities are earlier indicators of the effects of soil management changes than SOC contents (Ndiaye et al. 2000), they are commonly monitored in agroforestry systems (Mungai et al. 2005; Mao et al. 2012; Fang et al. 2013; Wang et al. 2017; Beuschel et al. 2019). Studies reporting the temporal evolution of soil parameters (SOM and microbial enzymes activities) in the first years after agroforestry systems were established as compared to the respective agricultural control systems are scarce. The introduction of trees in agricultural systems may induce belowground interactions between associated species. Depending on the nature and the intensity of these interactions, either competition or facilitation for nutrient or water can be evidenced between species. In agroforestry systems, many synchronic studies did not provide any information about the dynamics of ecological interactions and only reported a final comparison many years after installation.
In this context, we conducted a field experiment (under temperate climate conditions) to evaluate the early effects of agroforestry systems on six soil enzyme activities and on different quantitative and qualitative parameters of SOM in comparison with control agricultural systems. Our approach based on localized repeated soil samplings during the first years after system establishment aims to provide data about early changes in the size of labile organic carbon pools or microbial enzyme activities. These potential changes could in turn affect ecophysiological characteristics and productivity of the stand through modifications of soil functioning and nutrient availability. The first objective was to monitor the effect of tree (alder or poplar) introduction in herbaceous crops (gramineous or alfalfa) on these parameters in the first years (one, two, and four years) after tree planting. We hypothesized that enzyme activities and labile pools of SOM (POM, microbial biomass, and soluble extractable C) were early and sensitive indicators of changes induced by tree introduction in cropping systems. The second objective was to investigate the relationships between enzyme activity and quantitative and qualitative parameters of SOM in alder and poplar agroforestry systems vs herbaceous controls. We hypothesized that the differences in growth and litter deposition between the N2-fixing (low C:N ratio for alder N-rich litter) and non N2-fixing (with high C:N ratio for poplar low-N litter) tree species, might differentially alter microbial and SOM parameters.
Materials and methods
Experimental site
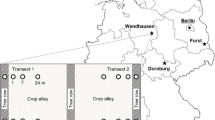
The study site (48°44′19”N, 6°18′50″E, elevation 219 m a.s.l.) is located 12 km north-east of Nancy, in north-eastern France, at the experimental farm of La Bouzule. The area is characterized by a degraded oceanic climate as defined in Joly et al. (2010) and the soil is a Vertic Stagnic Cambisol (IUSS Working Group WRB 2006). Prior to the setting-up of the experimental design, 8 soil cores (0–20 cm) regularly distributed across the site (Fig. 1) were sampled in mid-November 2013 to measure initial soil physico-chemical parameters. The soil is characterized by a clayey texture (Table 1). For the whole site, the mean SOM content was 5.3%, the mean CaCO3 content was 1.8% and the mean pHwater was equal to 7.4 (Table 1).

Experimental design at the La Bouzule site: randomized complete block design
Over the 2008–2013 period, the site was an experimental trial consisting of three blocks (100 × 72 m), divided into eight plots of 50 × 18 m to evaluate the effect of cropping systems on weed diversity (Romillac 2015). The site was managed according to regional recommendations for fertilization and the level of herbicide use was reduced by 50% compared to the regional treatment frequency index.
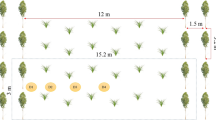
In 2014, the whole experimental site (3.3 ha) was divided into three blocks, each block including one replicate plot (ca. 0.15 ha, 73 m length, 20 to 24 m width) of each treatment (Fig. 1). Soil parameters presented spatial variability over the site with an increase of pH from block 1 to block 3 (Table 1). Also, block 3 is characterized by a lower clay content and a higher sand content than blocks 1 and 2 (Table 1).
Experimental plots were installed during the spring of 2014. Agricultural control plots were 4 years of continuous alfalfa for the first system and the rotation was spring wheat – winter wheat – triticale – ryegrass in the second control system in which these crops were grouped together and defined as “gramineous” hereafter. Within each block, silvoarable agroforestry plots are adjacent to their respective agricultural control plots. Agroforestry systems associated N2-fixing species (alder, alfalfa over the whole 2014–2018 period) to non-fixing species (poplar, cereals over the 2014–2017 period, and perennial ryegrass in 2018). Poplar trees (Populus deltoides × P. nigra, clone Dorskamp) were planted as 20-cm long woody cuttings, while alder trees (Alnus glutinosa) were planted as one-year-old seedlings in April 2014. The experimental site also includes forestry plots (alder monoculture, poplar monoculture, and alder/poplar association) not investigated in the present study, which is focused on the effects of tree introduction in agricultural systems. The experimental system was designed to maximize interactions between associated crops in order to investigate them: the trees were planted at 2-m intervals with a cultivated inter-row of 5 m corresponding to a tree density of 1000 trees per ha (i.e. 10 to 50 times higher than common silvoarable practices under temperate climate). Reduced tillage with disc and rotary harrow was conducted before crop sowing. Alfalfa was sown in 2014 concomitantly with poplar planting. Wheat and triticale were sown between the end of September and mid-October, respectively in 2015 and 2016. Ryegrass was sown in August 2017. Mineral fertilization was applied on gramineous systems associated or not with trees. The annual fertilization averaged 120 kg N ha−1. No fertilization was added on alfalfa and alfalfa-poplar systems. Pesticides were applied on wheat just during the spring of 2015. Cereal straws were exported after grain harvest. Perennial species (alfalfa and ryegrass) were cut two times per year in June and September for livestock feeding.
Climatic data
Daily air temperature, rainfall (R) and potential evapotranspiration (PET, Penman 1948) were recorded at the Champenoux weather station located 2 km away from the experimental site. Daily climatic data were used to calculate mean monthly and annual temperatures (°C) and cumulative R and PET (mm), which were also used to calculate annual water balance values (R-PET).
Tree growth monitoring
Tree growth was monitored on all trees in each agroforestry plot. Tree height was measured yearly at the end of the growing season using a graduated pole to the nearest cm. The trunk diameter at 1.30 m height (Diameter at Breast Height, DBH) was only measurable from 2017, using a digital calliper to the nearest mm.
Soil sample collection
In order to monitor the topsoil organic matter characteristics and microbial parameters, 3 soil cores were sampled (0–15 cm depth, diameter of 8 cm) in the central inter-row of each plot (2.5 m from the tree rows in agroforestry treatment or 10-12 m from the edge of the plots in control treatments), 24, 36, and 48 m from the bottom of each plot in all treatments (Fig. 1). Samplings were performed 1, 2, and 4 years after field establishment (13/05/2015, 12/07/2016, and 03/07/2018, respectively). An initial soil sampling was also performed when the site was installed (on 28/04/2014) 24 and 48 m from the bottom of each plot to assess the spatial variability of SOM by infrared spectroscopy.
Soil analyses
The soil samples taken in 2014, 2016, and 2018 were air-dried and ground before analysis by near-infrared (NIRS) and mid-infrared (MIRS) spectroscopy to investigate potential changes in SOM composition (for the detailed protocol and materials, see Akroume et al. 2016).
In 2015, 2016, and 2018, soil organic C and N contents were measured on soil samples decarbonated with hydrochloric acid using the Dumas dry combustion method according to NF ISO 10694. The soil water content was determined on all samples by the gravimetric method (NF ISO 16586).
Concerning the SOM labile pools, microbial biomass C and N were determined using the fumigation-extraction method (Vance et al. 1987) on field-moist soils. Chloroform-fumigated and non-fumigated control soil samples were extracted for 45 min using 0.5 M K2SO4 (1:5 w/v ratio) and then filtered through Whatman 42 paper. Dissolved C and N in the soil extracts were analyzed using a TOC-TN analyzer (Shimadzu). Conversion factors of 0.45 (Joergensen 1996) and 0.54 (Brookes et al. 1985) were applied to convert extractable C and N flushes into microbial biomass C and N (expressed in mg C or N per kg of soil), respectively. K2SO4 extractions from non-fumigated soil samples were used to estimate extractable organic carbon (EOC, expressed in mg C per kg of soil). Permanganate-oxidizable carbon (POXC) was measured on 2.5 g of soil (2-mm sieved and air-dried) shaken 2 min with 20 mL of 0.1 M KMnO4 solution following the method described in Culman et al. (2012). After 10 min of settling time, 250 μL of supernatant were mixed with 750 μL of deionized water and samples were read for absorbance at 550 nm. POXC was calculated following Weil et al. (2003) and expressed in mg oxidizable C per kg of soil.
Additionally, SOM physical fractionation into particulate organic matter (POM, > 50 μm fraction) and mineral-associated organic matter (MaOM, < 50 μm fraction) was performed on 12 soil samples taken in 2018 (one central sample per treatment within each block) (Gavinelli et al. 1995). Fractionation was performed on 40 g of dry soil sieved at 2 mm. The first step consists in soil dispersion with sodium hexametaphosphate (0.25%) and beads under agitation prior to application of separation criteria. Secondly, a wet-sieving step allows the separation of the 50–2000 μm fractions corresponding to POM fractions. The residual material obtained from the sieving process was dried at 40 °C, weighted and crushed and subsequently analyzed for organic carbon (NF ISO 14235) and nitrogen contents (NF ISO 11261). MaOM and POM fractions were expressed in percentages of total SOM. The N contained in the MaOM and POM fractions was expressed in g N per kg of soil.
Microbial enzyme activities and abundances
After sampling, soils were transported within 1 h to the laboratory and then 5 mm-sieved. They were kept fresh at 4 °C and further analyzed for enzyme activities on field-moist soils within 48-72 h. The potential soil enzyme activities of arylsulfatase (ARS, EC 3.1.6.1), β-glucosidase (BG, EC 3.2.1.21), leucine aminopeptidase (LAP, EC 3.4.11.1), N-acetyl-β-glucosaminidase (NAG, 3.2.1.52), and phosphatase (PH, EC 3.1.3.3) were investigated using the fluorometric microplate assay (Marx et al. 2001). Protease activity (PROT, EC 3.4.2.21–24) was determined spectrophotometrically (Ladd and Butler 1972). Potential enzyme activities were assessed using 200 μM substrate solutions of 4-MUB-sulfate, 4-MUB-β-D-glucopyranoside, L-leucine-7-amido-4-methylcoumarin, 4-MUB-N-acetyl-β-D-glucosaminide, and 4-MUB-phosphate for ARS, BG, LAP, NAG, and PH, respectively, and 2% Na-caseinate for PROT. Soil suspensions were prepared with deionized water in an 8:100 soil-to-water ratio for measuring PROT, and then diluted to obtain suspensions at a 1:100 soil-to-water ratio that were used in fluorometric enzyme assays. Soil suspensions and substrate solutions were mixed in a 1:1 proportion, and enzyme assays were carried out in buffered conditions and at 37 °C except for protease activity incubated at 45 °C. ARS, BG, NAG and PH were assayed in 0.5 M sodium acetate buffer (pH 5.5), LAP and PROT were assayed in 0.1 M Tris-HCl buffer (pH 8.0). The incubation times, preliminarily determined to ensure that the reactions would be linear over time, were 6, 2, 3, 5, 1, and 6 h for ARS, BG, LAP, NAG, PH, and PROT, respectively. Fluorescence (excitation and emission wavelengths set at 360 and 460 nm, respectively) and absorbance (at 680 nm) were measured using a microplate reader (Synergy HT, Bio Tek instruments). The potential enzyme activities were expressed in nmoles of MUB (for ARS, BG, NAG and PH) or AMC (for LAP) formed per gram of soil (dry weight) per hour, or in μg tyrosine equivalent released per gram of soil (dry weight) per hour for PROT.
Bacterial and fungal abundances were estimated by quantitative PCR (qPCR) on 12 soil samples taken in 2018 (one central sample per treatment within each block). Total DNA was extracted from 0.25 g of field-moist soil (2-mm sieved) using the DNeasy powersoil kit (Qiagen) and quantified using the Quant-iT DNA Assay Kit (Invitrogen). Quantitative PCR was used to quantify bacterial 16S rRNA genes (Muyzer et al. 1993) and fungal Internal Transcribed Spacers (White et al. 1990; Schoch et al. 2012) following the procedure described in Soper et al. (2018). Abundances were expressed as the number of copies of 16S rRNA genes or Internal Transcribed Spacers (ITS) per ng DNA.
Data analysis
Kruskal–Wallis and post-hoc (kruskalmc) tests were used 1) to test the effect of blocks on tree height and DBH, for each tree species and for each year, and 2) to compare tree height and DBH between poplar and alder, for each year.
For infrared spectroscopy analyses, the 8000–4000 and 4000–550 cm−1 regions were selected for NIRS and MIRS, respectively. Spectral data were pre-processed using Savitzky-Golay smoothing with a second-order polynomial degree (Savitzky and Golay 1964) and the standard normal variate (SNV) method, successively. Principal component analyses (PCAs) were performed to compare the absorbance spectra of the soil samples for the two frequency ranges. For the 2014 data (i.e. at the time of field establishment), we investigated SOM spatial variability by comparing soil spectra from the 3 blocks (n = 8 spectra per block) of the experimental site. Data from 2016 and 2018 were then used to investigate potential changes after 2 and 4 years of system differentiation by comparing the soil spectra of the agricultural and agroforestry modalities within each block. For the 2016 and 2018 infrared spectroscopy analyses, we analyzed 3 subsamples per soil replicate (n = 9 spectra per treatment and per block), and used them in the PCA projection to integrate the variability of the soil sampling and the analytical method in the comparative analysis. Differences in spectral profiles among crop types (alfalfa vs gramineous) and cropping systems (agroforestry vs agricultural) were determined using permutational analysis of variance (PerMANOVA) based on Bray-Curtis dissimilarities, which was conducted using the “adonis” function from “vegan” package. When significant, a post-hoc test with 999 permutations and a Bonferroni adjustment of p-values was applied using the “pairwise.perm.manova” function (“RVAideMemoire” package) to identify significant differences between treatments.
The potential effects of tree introduction (“system effect”), sampling year (“year effect”) and spatial variability (“block effect”) on the soil and microbial variables were tested within each pair of agricultural control-agroforestry modality by fitting a mixed-effects model to the data (“lme” function from “nlme” package). System, block and year were defined as fixed effects in the model and replicate samples as random effect. Temporal continuous autoregressive correlation structure was integrated in the mixed-effects model using the “corCAR1” function to account for autocorrelation between soils repeatedly sampled at different time intervals. Data were log-transformed when necessary to satisfy the ANOVA assumptions (Zar 1999). Pairwise post-hoc tests were performed using the “emmeans” package to compare the means between systems when a significant system effect was found.
The Response Ratio (RR) as defined in Hedges et al. (1999) was used as a metric to assess for each year the size effect of tree introduction on investigated variables. The natural log value of the RR between agroforestry and control systems and the sampling variance for each block were calculated as described in Jian et al. (2016). The weighted mean RR for the three blocks and 95% confidence intervals (CI) were then calculated using the “rma” function of the “metafor” package. Mean RR were considered significantly different from zero when the CI did not overlap the zero value.
Pearson correlation analyses were performed to investigate the relationships between microbial enzyme activities and variables likely to explain their variations. Pearson correlation p-values were adjusted using the Bonferroni correction.
R version 3.5.2 was used for all statistical analyses (R Core Team 2016). A probability level of 5% was applied.
Results
Climate
The site was characterized by a degraded oceanic climate with cumulative rainfalls (R) ranging between 542 and 808 mm (mean 673 mm) per year over the 2014–2018 period. The mean annual temperature was between 10.3 and 11.4 °C (mean 10.9 °C) with maximum monthly temperatures recorded each year during the June–August period and the coldest season observed during the November–March period (Fig. 2). The annual water balance (R-PET) was −19 mm on average over the investigated period; 2015 was the driest year (R-PET = −178 mm) and 2016 was the wettest one (R-PET = 198 mm). The average temperature and cumulative rainfall over the last 60 days before soil sampling ranged in the following way: 2015 (10.1 °C) < 2016 (16.2 °C) < 2018 (17.7 °C) for temperatures and 2015 (98 mm) < 2018 (136 mm) < 2016 (239 mm) for cumulative rainfall.

Monthly variations in mean air temperature (dots) and cumulative rainfall (histograms) recorded at the Champenoux weather station located 2 km away from the experimental site. Arrows indicate the topsoil sampling dates when the site was established in 2014 and after 1, 2, and 4 years of cropping system differentiation
Tree growth monitoring
Overall, many trees died the two first years and had to be replanted in 2015 (mainly for alder: + 138 trees) and in 2016 (mainly for poplar: + 107 trees). In the first four years, tree growth, evaluated through height and DBH, was significantly greater for poplar trees (associated with alfalfa) than for alder trees (associated with gramineous plants), except for tree height in 2016 (Table 2). In 2018, (i.e. four years after the site was established), mean tree height was 264 cm for alder as compared to 343 cm for poplar trees, and DBH was 26.8 and 18.7 mm for poplar and alder trees, respectively. Tree height and DBH were significantly higher in block 1 for both species than in blocks 2 and 3 (except for 2014, when we did not find a significant difference between blocks 1 and 3). Nevertheless, a significant differentiation in terms of tree growth between blocks 2 and 3 started in 2016. The mean height of alder trees was greater in block 3 than in block 2 contrary to poplar trees that grew better in block 2 than in block 3 (Table 2).
Soil parameters and organic matter pools
PCA analyses of soil infrared spectra showed differences between the three blocks of the experimental site in 2014, both in the MIR (4000 to 550 cm−1, Fig. 3a) and NIR (8000 to 4000 cm−1, Fig. 3b) regions. PerMANOVA confirmed a significant block effect on soil MIR and NIR spectral data (p = 0.001), each block being found to differ significantly from the others (p < 0.05). MIRS and NIRS spectra from the soils sampled in block 3 appeared more diverse within this block in the PCA ordinations than the spectra from blocks 1 and 2. No marked effects of the cropping systems (agroforestry vs control) were observed on the soil spectra in 2016, two years after the trees were planted. However, significant crop effects (alfalfa vs gramineous) were found in each block in 2016 and 2018 (p < 0.05). In 2018, a system effect was found in blocks 2 and 3 for MIRS and NIRS data (p < 0.01), a clear differentiation being notably observed in block 2 between the soils under the alfalfa-poplar system and those from the control system (p < 0.01).

Principal component analysis ordinations of soil MIRS (A) and NIRS (B) spectra at the time of tree planting in 2014 and after two (2016) and four years (2018) of system differentiation. Statistical ellipses represent 90% confidence
Results from ANOVAs on the soil C and N contents measured in 2015, 2016 and 2018 (Table 3A and B) confirmed a significant block effect (p < 0.01) on soil C in alfalfa and gramineous-based systems, and on soil N and C:N in the alfalfa-based system (p < 0.05); soil C increased from block 1 to block 3 (Table S1). Significant block effects were found in both alfalfa and gramineous-based systems for microbial biomass C, N and EOC (Table 3A and B). The effect of sampling year was also highly significant for the soil water content and most OM pools. Main effects of system were observed on microbial biomass C:N and soil water content in alfalfa- and gramineous-based systems, respectively, with significant interactive effects between system and year found for the soil water contents in alfalfa and gramineous-based systems (p < 0.05) and for microbial biomass N and C:N in alfalfa plots (p < 0.01). The soil water contents were significantly higher in alfalfa-poplar plots than in controls in 2018 (post-hoc test: p = 0.03) and in gramineous-alder than in controls in 2016 (post-hoc test: p = 0.02). Under alfalfa, microbial biomass N was significantly higher in the agroforestry plots than in their controls in 2018 (post-hoc test: p = 0.01), while microbial biomass C:N was the lowest in agroforestry plots in 2016 and 2018 (post-hoc test: p < 0.05).
Analysis of the response ratios RR (Fig. 4) revealed significant positive effects of tree introduction on soil water contents after 2 years in the gramineous-based system and after 4 years under alfalfa. In the latter system, the water content was nevertheless negatively impacted in 2015 and 2016. Soil C and N responses appeared to slightly increase over time in both cropping systems but the responses did not differ significantly from zero, soil C:N showing opposite trends between alfalfa and gramineous-based systems.

Size effects of tree introduction on soil parameters measured 1, 2, and 4 years after tree planting in 2014. Points refer to the weighted mean response ratio between agroforestry and control systems for the three blocks and the error bars represent the 95% confidence intervals. Confidence intervals overlapping with the dotted line (ln response ratio = 0) indicate no significant effect of tree introduction
A significant negative effect of tree introduction on microbial biomass N was observed in 2015, while a positive effect was later detected in 2018 but only under alfalfa (Fig. 5). Microbial biomass C:N response was found to be negatively affected in both systems after 4 years, the response being more marked under alfalfa. EOC showed an important positive response in 2016 (significant under alfalfa) but the variability of the RR appeared very high. Compared to other variables, the RR of POXC were relatively small.

Size effects of tree introduction on soil organic matter labile pools measured 1, 2, and 4 years after tree planting in 2014. Points refer to the weighted mean response ratio between agroforestry and control systems for the three blocks and the error bars represent the 95% confidence intervals. Confidence intervals overlapping with the dotted line (ln response ratio = 0) indicate no significant effect of tree introduction. Biomass C:N: C to N ratio of microbial biomass, EOC: K2SO4-extractable organic carbon, POXC: permanganate oxidizable carbon
Four years after tree introduction, additional analyses were performed to investigate potential changes in microbial communities and SOM fractions. Results of qPCR analyses (Fig. 6) showed no significant differences in bacterial and fungal abundances between agroforestry and control plots in 2018. However, although non significant, a decrease in fungal ITS can be observed in the alfalfa-based system concurrent to an increase in 16S/ITS ratio and could suggest an emergent effect on microbial assemblages.

Abundances and ratios of bacterial 16S rRNA and fungal Internal Transcribed Spacers (ITS) in the topsoils of control and agroforestry plots after four years of system differentiation. Barplots represent the means of the 3 blocks for each treatment ± SD
Analysis of the SOM fractions revealed that the mean proportion of the MaOM fraction was around 83% as compared to 17% for the POM fraction. The proportions of both fractions and their N contents and C:N ratios did not differ significantly between the agroforestry and agricultural plots, but the trends seemed to be slightly opposite in the alfalfa and gramineous systems (Fig. 7).

Proportions (%), N contents, and C-to-N ratios (C:N) of mineral-associated organic matter (MaOM) and particulate organic matter (POM) fractions in the topsoils of control and agroforestry plots after four years of system differentiation. Barplots represent the means of the 3 blocks for each treatment ± SD
Microbial enzyme activities
ANOVA results revealed a block effect on two out of six enzyme activities (LAP and PH) under alfalfa and on two enzymes under gramineous crops (ARS and LAP). Similarly to most OM pools a significant year effect was found on all investigated enzyme activities (Table 3A and B), which were found to be the highest in 2016 for most enzymes (Table S1). Comparatively to the year effect, few significant effects of tree introduction were found on microbial enzyme activities. A significant system effect was detectable for NAG and PH activities but only in the alfalfa-based system (p < 0.05), these activities being lower in agroforestry plots than in their controls (post-hoc test: p < 0.05).
Response ratios of microbial enzyme activities (Fig. 8) showed a negative effect of tree introduction on some enzymes involved in the C and N cycles (BG, NAG and LAP) after one year (2015) in alfalfa, these negative effect being offset two years after field establishment. A slight positive effect was also observed on PH activity under gramineous crops after four years (2018).

Size effects of tree introduction on soil potential enzyme activities measured 1, 2, and 4 years after tree planting in 2014. Points refer to the weighted mean response ratio between agroforestry and control systems for the three blocks and the error bars represent the 95% confidence intervals. Confidence intervals overlapping with the dotted line (ln response ratio = 0) indicate no significant effect of tree introduction. ARS: arylsulfatase, BG: β-glucosidase, LAP: leucine aminopeptidase, NAG: N-acetyl-β-glucosaminidase, PH: phosphatase and PROT: protease
Four out of six enzyme activities (BG, LAP, PH and PROT) were significantly and positively correlated with the soil water content (0.4 < r value <0.64, p < 0.05) (Figure S1). Enzyme activities related to the N (LAP and NAG) and P (PH) cycles were positively correlated with BG (Figure S1). Overall, enzyme activities were more correlated with microbial biomass C than with the soil C content. Three enzyme activities (BG, LAP and NAG) were thus significantly and positively correlated with microbial biomass C (Pearson r value ranging between 0.60 and 0.79, p < 0.05) and four (BG, LAP, NAG and PH) with the C:N ratio of microbial biomass (0.41 < r value <0.67, p < 0.05). Considering the relationships between enzyme activities and soil parameters or OM pools for each agroforestry system, it should be noted that the significant and positive correlations between enzyme activities and water content or microbial biomass C:N were stronger in the gramineous-alder than in the alfalfa-poplar plots (data not shown). Among the investigated enzymes, ARS and PH showed the most significant and negative correlations with the soil C:N ratio and EOC (−0.43 < r value < −0.49, p < 0.05). Generally, ARS and PH were the lowest in block 3 where soil C:N and EOC contents were higher than in blocks 1 and 2 (Table S1).
Discussion
Effects of spatial and temporal variabilities on soil parameters
In our study under temperate climatic conditions, we monitored the effect of tree (alder or poplar) introduction in cropping systems on SOC, labile pools of SOM and enzyme activities, one, two, and four years after tree planting. Overall, our results showed few significant main effects of tree introduction on variables measured in the topsoil (0–15 cm layer) whereas significant influences of temporal (year effect) and spatial (block effect) variabilities were evidenced. During the first years, the influence of environmental factors such as climate or soil heterogeneity may potentially prevent detecting subtle effects resulting from agroforestry system implantation. An effect of sampling year was evidenced on microbial pools and enzyme activities concurrently to soil water content, these latter being the highest in 2016, the wettest year. This result is in line with a previous work on global drivers of microbial abundance in soils showing that microbial biomass is positively related with annual moisture availability (Serna-Chavez et al. 2013). The increase in microbial biomass could in turn explain the highest potential enzyme activities measured in 2016 and the correlation found between microbial biomass and most enzyme activities, as previously demonstrated by Acosta-Martínez et al. (2008). Moisture can also play a role in enzymes and substrates diffusion in soil and on plant growth which could affect enzyme activities through the release of root exudates (Burns et al. 2013).
From the rhizosphere to the field scales, one important characteristic of the soil environment is its spatial heterogeneity which may affect at various scales the microbial ecology and the distribution of enzymes in soils (Hinsinger et al. 2005; Baldrian 2014). The experimental site of La Bouzule was characterized by an increasing gradient of SOC and pH. Differences in SOM and soil pH could potentially explain the block effect that we emphasized on OM pools and some enzyme activities. Indeed, some enzyme activities are well known to be related to these soil parameters and notably PH and LAP which are negatively and positively correlated, respectively, with soil pH (Sinsabaugh et al. 2008). Accordingly, we observed that PH activity was the lowest in block 3, where LAP was the highest compared to the other blocks. Soil MIRS and NIRS spectra confirmed the differences in SOM quality between blocks constituting our experimental site and more particularly the greatest variability of soil within the block 3 compared to the other blocks. Infrared spectroscopy analyses have the advantage of being rapid and less expensive than some of the other techniques for soils analyses (Nanni and Demattê 2006) and has been suggested as a tool for characterizing the spatial heterogeneity of soil properties in field experiments (Akroume et al. 2016). However, international soil infrared spectral libraries as well as spectral characterization are needed in order to use infrared reflectance spectroscopy analyses to identify soil properties and SOM fractions for soil quality and fertility assessment (Cécillon et al. 2009; Du and Zhou 2009; Grinand et al. 2012). Four years after the site was established, the analyses of MIRS-NIRS spectral data revealed that the effects of tree introduction were more marked in the alfalfa-based system, which suggest a differentiation in SOM quality.
Effects of tree introduction on soil early indicators
Agroforestry systems can play an important role in greenhouse gas mitigation through the sequestration of atmospheric CO2 in plants and soils. Notably, an increase in SOC stocks is expected when trees are planted in agricultural systems (De Stefano and Jacobson 2018). Nevertheless, studies reporting the temporal evolution of SOC in the first years after tree introduction are relatively scarce and despite higher organic matter inputs in agroforestry systems, differences in SOC between young agroforestry systems and their agricultural control plots are not necessary well marked (Mao and Zeng 2013). Four years after tree introduction, differences in SOC between agroforestry and control systems were relatively small and non-significant (increase of 3 and 5% in SOC contents for alfalfa-poplar and gramineous-alder vs controls, respectively). This result is consistent with the analysis performed by Chatterjee et al. (2018) in which SOC stocks in the topsoil (0–20 cm) were reported to be 6% higher in agroforestry systems aged between 0 and 5 years compared to agricultural ones under temperate climate.
In France, Cardinael et al. (2017) found that the SOC concentration (0–10 cm layer) was significantly higher (increase of 10%) in one 6-year-old agroforestry system (inter-rows) than in the agricultural control plot. Previous studies reported that agroforestry systems can lead to significant increases of SOC when the systems are installed on the long term (Lee and Jose 2003; Oelbermann et al. 2006; Mao et al. 2012; Tian et al. 2013) but also that carbon sequestration rates are higher in tropical climate compared to temperate one (Feliciano et al. 2018).
Compared to SOC, it has been shown that labile fractions of SOM, such as MBC and POM, can be early and more sensitive indicators of changes induced by agroforestry practices (Marquez et al. 1998; Wang et al. 2005; Mao and Zeng 2013), because of their fast turnover time. As C availability is one of the most important drivers of microbial activity, different methods have been proposed to evaluate and characterize the soil soluble organic C (Sparling et al. 1998; Ghani et al. 2003). The amount of these available C pools, which may thus affect microbial enzyme production, depends on the soil type but also on the plant type and seasonal conditions (Uchida et al. 2012). We did not observe significant difference in proportion of POM fractions in the topsoils of agroforestry plots as compared to the agricultural control plots, four years after tree introduction. Likewise, any clear effects on EOC and POXC were found in both agroforestry systems. In the alfalfa-based system, negative effects of tree introduction were observed on microbial biomass N (MBN) and enzyme activities one year after field establishment. The poplar introduction in alfalfa cropping system may have induced a decrease in the topsoil water content in 2015 as compared to the control alfalfa. These changes in soil moisture content during plant growth can markedly affect the soil microbial biomass (Xue et al. 2017) and could explain that MBN and some microbial activities were found to be lower in 2015 in the alfalfa-based agroforestry system as compared to the control. After 4 years, the soil water contents were higher in agroforestry systems compared to their controls but the negative effects observed the first year on microbial activities appeared to have been offset with no differences between systems. MBN was even found to be higher in alfalfa-poplar plots compared to their control ones. Previous studies on longer time-scale reported greater enzymes activities (BG and NAG) and increased OM levels in the tree rows compared to the middle of alleys in agroforestry systems composed of pecan or pin oak associated with grasses 21 and 9 years after tree planting (Mungai et al. 2005; Udawatta et al. 2008). In temperate climate, no difference in enzyme activity (BG and NAG) was observed in alleys (between 1 and 24 m from tree row) in a younger poplar alley cropping system (5–8 years) (Beuschel et al. 2019). Under tropical climate, Chander et al. (1998) directly compared different density (spacings of 5 × 5 to 10 × 10 m) of Sesham (a N2-fixing tree) intercropped with a wheat-cowpea cropping sequence and a control modality without trees. They found higher organic C, MBC and enzyme activities (dehydrogenase and alkaline phosphatase) in the topsoils (0–15 cm) of tree-crop systems than in the system without trees 12 years after tree planting, with an observed increase in these variables with the decrease in the tree spacing. The early effects of tree introduction reported in our study were more pronounced in the poplar-alfalfa system compared to the alder-gramineous system probably because of differences in growth between both tree species, whose respective litter quality may also affect the soil functioning in different ways.
Differences in tree growth and soil functioning between alder and poplar agroforestry systems
At the experimental site of La Bouzule, alder and poplar were planted in agroforestry systems at the end of April 2014. Measures of tree growth confirmed the soil spatial variability of the experimental site. The differences in terms of mortality and growth rates among the three blocks suggest more favorable conditions for tree growth in the block 1. The soil characteristics differed from the block 1 to block 3 with an increase of pH and carbon content whereas soil depth decreases, with a significant outcrop of rock material (calcareous) near the soil surface in block 3. During the first two years, the water balance was −72 mm between April and December 2014 and − 178 mm in 2015. In these conditions, the topsoil became dry very early in 2014 and 2015, generating a severe compaction which could have altered the ability of young roots of poplar to grow and colonize soil profile during dry periods (DesRochers and Tremblay 2009). Indeed, an extreme competition for water could have occurred between trees and herbaceous, notably in the case of the poplar-alfalfa system as suggested by the negative effects on soil water contents in 2015 and 2016. As poplar has water requirements exceeding those of crops (Deckmyn et al. 2004) or grasslands (Persson 1997), it is likely that in our experimental site, poplar cuttings that did not have pre-established their root systems could have suffered from competition with alfalfa explaining the high tree mortality. Poplar are fast growing trees, as a result, we measured significant higher height for poplar compared to alder trees, except for 2016 when poplar trees had to be replanted. DBH measured in 2017 and 2018 were also significantly higher for poplars than for alders. The higher tree growth of poplar compared to alder and the potential competition for water with alfalfa could explain that the observed effects of tree introduction on microbial variables were more marked in the alfalfa-based system compared to the gramineous one. After four years, small but non-significant positive effects on SOC were observed in the two agroforestry systems suggesting C depositions at similar levels for poplar- and alder- based systems. However, we observed opposite evolution in soil C:N ratio over time (decreasing trend in alder- and increasing trend in poplar- agroforestry system compared to controls, respectively). This result might be partly explained by the difference between quantity or quality of rhizodeposition and/or litter (root and aerial). Tree litter mass was not collected on the two agroforestry systems due to the young tree age and thus, we did not have valuable data to compare the quantity of C and N inputs in soil in alder and poplar systems. Based on the literature, C:N ratio averaged 34 for poplar trees (Populus deltoides × P. nigra, clone Dorskamp) compared to 21 for N2-fixing trees such as Robinia pseudoacacia (Marron, unpublished data) or 16 for Alnus glutinosa (Angst et al. 2017). The two different types of tree leaf litters, N-rich litter with low C:N ratio for alder (N2-fixing tree) and low-N litter with high C:N ratio for poplar, is expected to influence differently soil microbial communities as evidenced by Sun et al. (2018) in a robinia-based system compared with a poplar-based system.
Significant and strong effects of tree introduction were observed on MBN (positive effect) and C:N ratio (negative effect) but only in the alfalfa-based system. The introduction of poplar in alfalfa could have led to a modification of the microbial diversity and/or a regulation of the microbial C and N use efficiencies (Manzoni et al. 2008; Mooshammer et al. 2014). Microbial decomposers may immobilize soil N when decomposing N-poor litter (such as poplar litter) to meet their stoichiometric requirements (Zhou et al. 2018). The decrease in microbial biomass C:N could suggest a decrease in the relative proportion of fungi vs bacteria in microbial assemblages as the C:N ratio of bacteria is generally lower than that of fungi (Hunt et al. 1987). The results of qPCR tended to confirm this hypothesis with an increase in 16S:ITS ratio resulting from a decrease in fungal ITS copy number in alfalfa-poplar plots compared to their control ones. Further analyses would be needed to ascertain this hypothesis and to understand more in depth the effects of tree introduction on soil microbial functioning.
Conclusion
In the present study, the effects of temperate agroforestry systems on SOC in the topsoil are relatively weak. Observed effects on soil water contents and OM pools were more pronounced in the alfalfa-poplar system, probably due to higher tree growth. After 1 year, tree introduction in the alfalfa system induced a decrease in soil water content and some early microbial indicators (biomass N and some enzyme activities) which appeared to be more sensitive than SOC to land use change. After 4 years, tree introduction resulted in positive effects on soil water contents in both agroforestry systems, while changes in microbial biomass were only found in alfalfa-poplar plots compared to their control plots. This work shows the importance of characterizing the initial state and the variability of the systems and their early changes, which will impact their evolution on the long term. Further studies should be conducted to measure root traits and C inputs (both in terms of quantity and quality) to assess the belowground interactions between trees and herbaceous crops along the soil depth profile. Seasonality and spatial variability should be taken into account to unravel the role of soil microbial diversity and functionality in the facilitation processes between associated species in regard to SOM dynamics and nutrient resources.
References
Acosta-Martínez V, Cruz L, Sotomayor-Ramírez D, Pérez-Alegría L (2007) Enzyme activities as affected by soil properties and land use in a tropical watershed. Appl Soil Ecol 35:35–45. https://doi.org/10.1016/j.apsoil.2006.05.012
Acosta-Martínez V, Acosta-Mercado D, Sotomayor-Ramírez D, Cruz-Rodríguez L (2008) Microbial communities and enzymatic activities under different management in semiarid soils. Appl Soil Ecol 38:249–260. https://doi.org/10.1016/j.apsoil.2007.10.012
Akroume E, Zeller B, Buée M et al (2016) Improving the design of long-term monitoring experiments in forests: a new method for the assessment of local soil variability by combining infrared spectroscopy and dendrometric data. Ann For Sci 73:1005–1013. https://doi.org/10.1007/s13595-016-0572-3
Angst Š, Cajthaml T, Angst G et al (2017) Retention of dead standing plant biomass (marcescence) increases subsequent litter decomposition in the soil organic layer. Plant Soil 418:571–579. https://doi.org/10.1007/s11104-017-3318-6
Baldrian P (2014) Distribution of extracellular enzymes in soils: spatial heterogeneity and determining factors at various scales. Soil Sci Soc Am J 78:11–18. https://doi.org/10.2136/sssaj2013.04.0155dgs
Bandick AK, Dick RP (1999) Field management effects on soil enzyme activities. Soil Biol Biochem 31:1471–1479. https://doi.org/10.1016/S0038-0717(99)00051-6
Beuschel R, Piepho H-P, Joergensen RG, Wachendorf C (2019) Similar spatial patterns of soil quality indicators in three poplar-based silvo-arable alley cropping systems in Germany. Biol Fertil Soils 55:1–14. https://doi.org/10.1007/s00374-018-1324-3
Blankinship JC, Schimel JP (2018) Biotic versus abiotic controls on bioavailable soil organic carbon. Soil Syst 2:10. https://doi.org/10.3390/soilsystems2010010
Brookes PC, Landman A, Pruden G, Jenkinson DS (1985) Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol Biochem 17:837–842. https://doi.org/10.1016/0038-0717(85)90144-0
Burns RG, DeForest JL, Marxsen J et al (2013) Soil enzymes in a changing environment: current knowledge and future directions. Soil Biol Biochem 58:216–234. https://doi.org/10.1016/j.soilbio.2012.11.009
Cardinael R, Chevallier T, Cambou A et al (2017) Increased soil organic carbon stocks under agroforestry: a survey of six different sites in France. Agric Ecosyst Environ 236:243–255. https://doi.org/10.1016/j.agee.2016.12.011
Cardinael R, Guenet B, Chevallier T et al (2018) High organic inputs explain shallow and deep SOC storage in a long-term agroforestry system – combining experimental and modeling approaches. Biogeosciences 15:297–317. https://doi.org/10.5194/bg-15-297-2018
Cécillon L, Barthès BG, Gomez C et al (2009) Assessment and monitoring of soil quality using near-infrared reflectance spectroscopy (NIRS). Eur J Soil Sci 60:770–784. https://doi.org/10.1111/j.1365-2389.2009.01178.x
Chander K, Goyal S, Nandal DP, Kapoor KK (1998) Soil organic matter, microbial biomass and enzyme activities in a tropical agroforestry system. Biol Fertil Soils 27:168–172. https://doi.org/10.1007/s003740050416
Chatterjee N, PKR N, Chakraborty S, Nair VD (2018) Changes in soil carbon stocks across the Forest-agroforest-agriculture/pasture continuum in various agroecological regions: a meta-analysis. Agric Ecosyst Environ 266:55–67. https://doi.org/10.1016/j.agee.2018.07.014
Culman SW, Snapp SS, Freeman MA et al (2012) Permanganate oxidizable carbon reflects a processed soil fraction that is sensitive to management. Soil Sci Soc Am J 76:494–504. https://doi.org/10.2136/sssaj2011.0286
De Stefano A, Jacobson MG (2018) Soil carbon sequestration in agroforestry systems: a meta-analysis. Agrofor Syst 92:285–299. https://doi.org/10.1007/s10457-017-0147-9
Deckmyn G, Muys B, Quijano JG, Ceulemans R (2004) Carbon sequestration following afforestation of agricultural soils: comparing oak/beech forest to short-rotation poplar coppice combining a process and a carbon accounting model. Glob Chang Biol 10:1482–1491. https://doi.org/10.1111/j.1365-2486.2004.00832.x
DesRochers A, Tremblay F (2009) The effect of root and shoot pruning on early growth of hybrid poplars. For Ecol Manag 258:2062–2067. https://doi.org/10.1016/j.foreco.2009.07.059
Dick RP, Breakwell DP, Turco RF (1996) Soil enzyme activities and biodiversity measurements as integrative microbiological indicators. Methods for Assessing Soil Quality, SSSA Special Publication 49:247–271. https://doi.org/10.2136/sssaspecpub49.c15
Du C, Zhou J (2009) Evaluation of soil fertility using infrared spectroscopy: a review. Environ Chem Lett 7:97–113. https://doi.org/10.1007/s10311-008-0166-x
Fang S, Liu D, Tian Y et al (2013) Tree species composition influences enzyme activities and microbial biomass in the rhizosphere: a rhizobox approach. PLoS One 8:e61461. https://doi.org/10.1371/journal.pone.0061461
Feliciano D, Ledo A, Hillier J, Nayak DR (2018) Which agroforestry options give the greatest soil and above ground carbon benefits in different world regions? Agric Ecosyst Environ 254:117–129. https://doi.org/10.1016/j.agee.2017.11.032
Gavinelli E, Feller C, Larré-Larrouy MC et al (1995) A routine method to study soil organic matter by particle-size fractionation: examples for tropical soils. Commun Soil Sci Plant Anal 26:1749–1760. https://doi.org/10.1080/00103629509369406
Geisseler D, Horwath WR, Joergensen RG, Ludwig B (2010) Pathways of nitrogen utilization by soil microorganisms – a review. Soil Biol Biochem 42:2058–2067. https://doi.org/10.1016/j.soilbio.2010.08.021
Ghani A, Dexter M, Perrott KW (2003) Hot-water extractable carbon in soils: a sensitive measurement for determining impacts of fertilisation, grazing and cultivation. Soil Biol Biochem 35:1231–1243. https://doi.org/10.1016/S0038-0717(03)00186-X
Grinand C, Barthès BG, Brunet D et al (2012) Prediction of soil organic and inorganic carbon contents at a national scale (France) using mid-infrared reflectance spectroscopy (MIRS). Eur J Soil Sci 63:141–151. https://doi.org/10.1111/j.1365-2389.2012.01429.x
Haney RL, Franzluebbers AJ, Jin VL et al (2012) Soil organic C:N vs. water-extractable organic C:N. Open J Soil Sci 02:269–274. https://doi.org/10.4236/ojss.2012.23032
Haynes RJ (2000) Labile organic matter as an indicator of organic matter quality in arable and pastoral soils in New Zealand. Soil Biol Biochem 32:211–219. https://doi.org/10.1016/S0038-0717(99)00148-0
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecology 80:1150–1156. https://doi.org/10.2307/177062
Hinsinger P, Gobran GR, Gregory PJ, Wenzel WW (2005) Rhizosphere geometry and heterogeneity arising from root-mediated physical and chemical processes. New Phytol 168:293–303. https://doi.org/10.1111/j.1469-8137.2005.01512.x
Hunt HW, Coleman DC, Ingham ER et al (1987) The detrital food web in a shortgrass prairie. Biol Fertil Soils 3:57–68. https://doi.org/10.1007/BF00260580
Islam KR, Weil RR (2000) Soil quality indicator properties in mid-Atlantic soils as influenced by conservation management. J Soil Water Conserv 55:69–78
IUSS Working Group WRB (2006) World reference base for soil resources 2006. 2nd edition. World Soil Resources Reports No. 103
Jian S, Li J, Chen J et al (2016) Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: a meta-analysis. Soil Biol Biochem 101:32–43. https://doi.org/10.1016/j.soilbio.2016.07.003
Joergensen RG (1996) The fumigation-extraction method to estimate soil microbial biomass: calibration of the kEC value. Soil Biol Biochem 28:25–31. https://doi.org/10.1016/0038-0717(95)00102-6
Joly D, Brossard T, Cardot H et al (2010) Les types de climats en France, une construction spatiale. Cybergeo Eur J Geogr. https://doi.org/10.4000/cybergeo.23155
Jones DL (1998) Organic acids in the rhizosphere – a critical review. Plant Soil 205:25–44. https://doi.org/10.1023/A:1004356007312
Klose S, Moore JM, Tabatabai MA (1999) Arylsulfatase activity of microbial biomass in soils as affected by cropping systems. Biol Fertil Soils 29:46–54. https://doi.org/10.1007/s003740050523
Ladd JN, Butler JHA (1972) Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol Biochem 4:19–30. https://doi.org/10.1016/0038-0717(72)90038-7
Lee K-H, Jose S (2003) Soil respiration and microbial biomass in a pecan — cotton alley cropping system in southern USA. Agrofor Syst 58:45–54. https://doi.org/10.1023/A:1025404019211
Lehmann J, Kleber M (2015) The contentious nature of soil organic matter. Nature 528:60–68. https://doi.org/10.1038/nature16069
Manzoni S, Jackson RB, Trofymow JA, Porporato A (2008) The global stoichiometry of litter nitrogen mineralization. Science 321:684–686. https://doi.org/10.1126/science.1159792
Mao R, Zeng D-H (2013) Effect of land-use change from cropland to poplar-based agroforestry on soil properties in a semiarid region of Northeast China. Fresenius Environ Bull 22:1077–1084
Mao R, Zeng D-H, Li L-J, Hu Y-L (2012) Changes in labile soil organic matter fractions following land use change from monocropping to poplar-based agroforestry systems in a semiarid region of Northeast China. Environ Monit Assess 184:6845–6853. https://doi.org/10.1007/s10661-011-2462-3
Marquez CO, Cambardella CA, Isenhart TM, Schultz RC (1998) Assessing soil quality in a riparian buffer by testing organic matter fractions in Central Iowa, USA. Agrofor Syst 44:133–140. https://doi.org/10.1023/A:1006261519080
Marx M-C, Wood M, Jarvis SC (2001) A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol Biochem 33:1633–1640. https://doi.org/10.1016/S0038-0717(01)00079-7
Mooshammer M, Wanek W, Hämmerle I et al (2014) Adjustment of microbial nitrogen use efficiency to carbon:nitrogen imbalances regulates soil nitrogen cycling. Nat Commun 5:3694. https://doi.org/10.1038/ncomms4694
Mungai NW, Motavalli PP, Kremer RJ, Nelson KA (2005) Spatial variation of soil enzyme activities and microbial functional diversity in temperate alley cropping systems. Biol Fertil Soils 42:129–136. https://doi.org/10.1007/s00374-005-0005-1
Muyzer G, de Waal EC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695–700
Nair PKR, Kumar BM, Nair VD (2009) Agroforestry as a strategy for carbon sequestration. J Plant Nutr Soil Sci 172:10–23. https://doi.org/10.1002/jpln.200800030
Nanni MR, Demattê JAM (2006) Spectral reflectance methodology in comparison to traditional soil analysis. Soil Sci Soc Am J 70:393–407. https://doi.org/10.2136/sssaj2003.0285
Ndiaye EL, Sandeno JM, McGrath D, Dick RP (2000) Integrative biological indicators for detecting change in soil quality. Am J Altern Agric 15:26–36. https://doi.org/10.1017/S0889189300008432
Oelbermann M, Voroney RP, Thevathasan NV et al (2006) Soil carbon dynamics and residue stabilization in a Costa Rican and southern Canadian alley cropping system. Agrofor Syst 68:27–36. https://doi.org/10.1007/s10457-005-5963-7
Penman HL (1948) Natural evaporation from open water, bare soil and grass. Proc R Soc Lond A 193:120–145. https://doi.org/10.1098/rspa.1948.0037
Persson G (1997) Comparison of simulated water balance for willow, spruce, grass ley and barley. Hydrol Res 28:85–98. https://doi.org/10.2166/nh.1997.0006
R Core Team (2016) R: A Language and Environment for Statistical Computing. WwwR-Proj
Rao MR, Nair PKR, Ong CK (1997) Biophysical interactions in tropical agroforestry systems. Agrofor Syst 38:3–50. https://doi.org/10.1023/A:1005971525590
Romillac N (2015) Effets de l’introduction du pois dans une succession de cultures sur certaines communautés végétales et bactériennes et leurs fonctions écosystémiques associées. Thesis, Université de Lorraine
Savitzky A, Golay M (1964) Smoothing and differentiation of data by simplified least squares procedures. Anal Chem 36:1627
Schimel J, Balser TC, Wallenstein M (2007) Microbial stress-response physiology and its implications for ecosystem function. Ecology 88:1386–1394
Schoch CL, Seifert KA, Huhndorf S et al (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc Natl Acad Sci 109:6241–6246. https://doi.org/10.1073/pnas.1117018109
Serna-Chavez HM, Fierer N, van Bodegom PM (2013) Global drivers and patterns of microbial abundance in soil. Glob Ecol Biogeogr 22:1162–1172. https://doi.org/10.1111/geb.12070
Shen Y, Cheng R, Xiao W et al (2018) Labile organic carbon pools and enzyme activities of Pinus massoniana plantation soil as affected by understory vegetation removal and thinning. Sci Rep 8:573. https://doi.org/10.1038/s41598-017-18812-x
Sinsabaugh RL, Lauber CL, Weintraub MN et al (2008) Stoichiometry of soil enzyme activity at global scale. Ecol Lett 11:1252–1264
Six J, Bossuyt H, Degryze S, Denef K (2004) A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res 79:7–31. https://doi.org/10.1016/j.still.2004.03.008
Soper FM, Sullivan BW, Nasto MK et al (2018) Remotely sensed canopy nitrogen correlates with nitrous oxide emissions in a lowland tropical rainforest. Ecology 99:2080–2089. https://doi.org/10.1002/ecy.2434
Sparling G, Vojvodić-Vuković M, Schipper LA (1998) Hot-water-soluble C as a simple measure of labile soil organic matter: the relationship with microbial biomass C. Soil Biol Biochem 30:1469–1472. https://doi.org/10.1016/S0038-0717(98)00040-6
Sun H, Koal P, Gerl G et al (2018) Microbial communities and residues in robinia- and poplar-based alley-cropping systems under organic and integrated management. Agrofor Syst 92:35–46. https://doi.org/10.1007/s10457-016-0009-x
Tian Y, Cao F, Wang G (2013) Soil microbiological properties and enzyme activity in Ginkgo–tea agroforestry compared with monoculture. Agrofor Syst 87:1201–1210. https://doi.org/10.1007/s10457-013-9630-0
Torralba M, Fagerholm N, Burgess PJ et al (2016) Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric Ecosyst Environ 230:150–161. https://doi.org/10.1016/j.agee.2016.06.002
Tsonkova P, Böhm C, Quinkenstein A, Freese D (2012) Ecological benefits provided by alley cropping systems for production of woody biomass in the temperate region: a review. Agrofor Syst 85:133–152. https://doi.org/10.1007/s10457-012-9494-8
Uchida Y, Nishimura S, Akiyama H (2012) The relationship of water-soluble carbon and hot-water-soluble carbon with soil respiration in agricultural fields. Agric Ecosyst Environ 156:116–122. https://doi.org/10.1016/j.agee.2012.05.012
Udawatta RP, Kremer RJ, Adamson BW, Anderson SH (2008) Variations in soil aggregate stability and enzyme activities in a temperate agroforestry practice. Appl Soil Ecol 39:153–160. https://doi.org/10.1016/j.apsoil.2007.12.002
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707. https://doi.org/10.1016/0038-0717(87)90052-6
Vong P-C, Piutti S, Benizri E et al (2007) Water-soluble carbon in roots of rape and barley: impacts on labile soil organic carbon, arylsulphatase activity and Sulphur mineralization. Plant Soil 294:19–29. https://doi.org/10.1007/s11104-007-9201-0
Wang H, Huang Y, Huang H et al (2005) Soil properties under young Chinese fir-based agroforestry system in mid-subtropical China. Agrofor Syst 64:131–141. https://doi.org/10.1007/s10457-004-0718-4
Wang Y, Ji H, Wang R et al (2017) Impact of root diversity upon coupling between soil C and N accumulation and bacterial community dynamics and activity: result of a 30year rotation experiment. Geoderma 292:87–95. https://doi.org/10.1016/j.geoderma.2017.01.014
Weil RR, Islam KR, Stine MA et al (2003) Estimating active carbon for soil quality assessment: a simplified method for laboratory and field use. Am J Altern Agric 18:3–17. https://doi.org/10.1079/AJAA200228
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: a Guide to Methods and Applications 18(1):315–322
Wolz KJ, DeLucia EH (2018) Alley cropping: global patterns of species composition and function. Agric Ecosyst Environ 252:61–68. https://doi.org/10.1016/j.agee.2017.10.005
Xue R, Shen Y, Marschner P (2017) Soil water content during and after plant growth influence nutrient availability and microbial biomass. J Soil Sci Plant Nutr 17:702–715. https://doi.org/10.4067/S0718-95162017000300012
Zar JH (1999) Biostatistical analysis. Prentice Hall, Upper Saddle River
Zhou G, Zhang J, Qiu X et al (2018) Decomposing litter and associated microbial activity responses to nitrogen deposition in two subtropical forests containing nitrogen-fixing or non-nitrogen-fixing tree species. Sci Rep 8:12934. https://doi.org/10.1038/s41598-018-30666-5
Acknowledgements
The UMR Silva, UR BEF and the experimental site are supported by the French National Research Agency through the Cluster of Excellence ARBRE (ANR-11-LABX-0002-01). This Research was also funded by La Fondation de France and the French Environment and Energy Management Agency (ADEME) through postdoctoral grants to H. Clivot and C. Petitjean. The experiment was supported by the mobile lab (M-POETE) of ANAEE-France. ANAEE-France is an infrastructure of the French Investment for the Future (‘Investissements d’Avenir’) program, overseen by the French National Research Agency (ANR-11-INBS-0001). We thank M.C. Géhin (INRA) for assistance in NIRS-MIRS analyses. We are also grateful to all the people who helped for maintain the experimental site and for field and lab work. This work is dedicated to the memory of our colleague Bernard Amiaud, Professor at the Université de Lorraine (1969-2018).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Remi Cardinael.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Clivot, H., Petitjean, C., Marron, N. et al. Early effects of temperate agroforestry practices on soil organic matter and microbial enzyme activity. Plant Soil 453, 189–207 (2020). https://doi.org/10.1007/s11104-019-04320-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-019-04320-6