
Abstract
Background and aims
Phosphorus (P) deficiency is a major problem for alfalfa (Medicago sativa) productivity on alkaline soils on the Loess Plateau, China. Our aim was to investigate growth, morphological and physiological responses of alfalfa to P supply in two alkaline soils when water supply is limited.
Methods
A pot experiment was carried out to grow alfalfa in two alkaline soils supplied with different rates of P. Parameters of plant growth and root morphology, rhizosphere pH and carboxylates, and plant concentrations of mineral nutrients were measured.
Results
Plant growth and nutrient uptake were enhanced by supplying P, but shoot growth was not further increased when P supply was >20 μg P g−1 soil. Specific root length was only responsive to changes in soil P when P supply was low in the loessial soil. The rhizosphere carboxylate amount was significantly greater when no P was supplied than when P was supplied to the loessial soil. The rhizosphere pH was lower than the bulk soil pH, but did not vary with soil P.
Conclusions
A P supply of 20 μg P g−1 soil was optimal for alfalfa growth. The responses of specific root length and rhizosphere carboxylates depended on soil type.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phosphorus (P) plays a vital role in plant growth and physiology, and P deficiency is one of the most limiting factors to crop production in many soils worldwide (Hinsinger 2001; Lambers et al. 2008; Richardson et al. 2011). The major problem with P deficiency is not the total P content in soil, but P availability to plants. In soils, P is often tightly bound to soil particles such as iron (Fe) and aluminum (Al) oxides and hydroxides in acid soils and calcium (Ca) carbonates in alkaline soils, and hence P is relatively immobile and often poorly available to most plants. Orthophosphate (Pi), which is taken up by plants, occurs in soil solutions at very low concentrations, i.e. < 0.6 to 11 μM (Hossner et al. 1973; Yanai 1991; Ron Vaz et al. 1993; Johnston et al. 2014), far lower than the intracellular Pi concentrations (5–20 mM) required for optimal growth of many crop plants (Vance et al. 2003; Fang et al. 2009).
Large amounts of P fertilizers have been applied to many soils globally to improve agricultural productivity. However, much of the P fertilizers applied to agricultural soils in soluble forms are quickly sorbed to the surface of soil particles, or precipitates with Ca, Al, and Fe, and becomes sparingly-available to plants (Simpson et al. 2011; Simpson et al. 2015). Phosphate fertilizers are often applied to soils in excess of plant requirement, and many soils have accumulated large stores of residual P and non-labile inorganic P over time (Vu et al. 2008). Large P inputs to agricultural soils not only increase the cost of production, but also cause serious environmental problems such as eutrophication of terrestrial and aquatic ecosystems (Carpenter 2005; Ulrich et al. 2016). Furthermore, phosphate rock, which is the world’s main source of P, is a non-renewable resource (Cordell et al. 2009; Fixen and Johnston 2012), it is urgently necessary to use P fertilizers in a sustainable way, and improve the P-acquire and use efficiency of cropping systems (Johnston et al. 2014; Cordell and White 2015).
Under P deficiency, plants may alter their root morphology and physiology to improve their capacity for P acquisition. Typical growth and morphological responses to P deficiency include a larger root mass ratio (RMR, the ratio of root dry mass to total plant dry mass), thinner roots and increased specific root length (SRL, root length per unit root dry mass), more and longer root hairs, and formation of mycorrhizal associations. All of these morphological responses are expected to increase total surface area for soil exploration and P acquisition at minimal energy cost (Vance et al. 2003; Lambers et al. 2006). An important physiological mechanism plants use to increase the rate of P absorption per unit root mass and length is the exudation of carboxylates such as citrate, oxalate, and malate, which can increase the mobilisation of sparingly-soluble soil P in the rhizosphere and result in enhanced P uptake (Lambers et al. 2006; Richardson et al. 2011). Some plants can increase P availability by altering the rhizosphere pH, e.g. P availability in alkaline soils is enhanced by rhizosphere acidification resulting from proton release during N2-fixation by some legumes (Hinsinger et al. 2003).
The levels of P supply in soils also affect the accumulation of mineral nutrients other than P by plants. At a high P supply, when more P is taken up, the uptake of some cations is also increased, and a high P concentration in shoots of some plants is accompanied by high concentrations of potassium (K), Ca, magnesium (Mg), and manganese (Mn) (Suriyagoda et al. 2012a; Aziz et al. 2015). The strategies plants deploy to enhance P acquisition can also result in increased uptake of other mineral nutrients (Lynch 2007). Carboxylates released by roots can mobilize micronutrients such as Fe, Mn, copper (Cu), and zinc (Zn) in soil, and, as a consequence, may lead to higher concentrations of these elements in plants (Page et al. 2006; Lambers et al. 2015). Rhizosphere acidification can make Fe, Mn, Cu, and Zn in alkaline soils more available for uptake by roots (Dakora and Phillips 2002). At neutral to alkaline pH, protons released by roots can enhance the dissolution of calcium carbonates to increase Ca2+ for plant uptake (Dakora and Phillips 2002). However, a few organic acids such as citric acid can decrease Ca uptake by roots due to their strong capacity to form stable complexes with Ca in solution at pH 6.0, and roots may not absorb Ca from organic complexes (Nunes et al. 2009).
Alfalfa (Medicago sativa L.) is an important perennial herbaceous forage legume, which has been cultivated for at least 150 years on the Chinese Loess Plateau, where it is an important component of the agroecosystem (Deng et al. 2014; Fan et al. 2015). Most soils on the Loess Plateau are typically alkaline and calcareous, and the productivity of alfalfa is limited by P deficiency and drought on these soils (Fan et al. 2015). Previous studies have demonstrated that, under P deficiency in both acid and alkaline soils, alfalfa can improve its P acquisition by increasing SRL and exuding carboxylates into the rhizosphere (Pang et al. 2010a; Suriyagoda et al. 2010; Fan et al. 2015). In the current study, we carried out a pot experiment to grow alfalfa in two alkaline soils supplied with different rates of P under low soil moisture conditions. The objectives of this study were to explore possible mechanisms that could enhance P acquisition by alfalfa grown in two alkaline soils under a range of P supplies with more emphasis on the deficiency range when soil water supply is limited, and investigate the effects of P supply levels and strategies deployed by plants to enhance P acquisition on the uptake of other mineral nutrients. We hypothesized that the plants would (1) show larger RMR under low P supply; (2) show increased SRL under low P supply; (3) exude more carboxylates into the rhizosphere under low P supply; (4) acidify the rhizosphere under low P supply; and (5) show increased uptake of other mineral nutrients when more P was taken up.
Materials and methods
Plant material and growth conditions
Two alkaline soils, i.e. an aeolian sandy soil (40°02′41″N, 111°22′22″E) and a loessial soil (39°47′35″N, 111°17′19″E) were collected from the top 40-cm layers of two undisturbed sites in the northern part of the Loess Plateau and used as the substrate for the pot experiment. Both the aeolian sandy soil and loessial soil were air-dried and passed through a 2-mm sieve before filling the pots; their physicochemical properties were analyzed and listed in Table 1. Concentration of bicarbonate-extractable P, also considered “plant-available P”, of the aeolian sandy soil and loessial soil was 2.4 and 1.1 μg P g−1 soil, respectively. External P was supplied as calcium superphosphate (chemical pure, Sinopharm Chemical Reagent Co. Ltd., Shanghai, China) at seven rates, i.e. 0, 5, 10, 20, 40, 80, and 160 μg P g−1 dry soil (hereafter referred to as 0P, 5P, 10P, 20P, 40P, 80P, and 160P treatment, respectively) for both soils, with no other fertilizer being supplied throughout the experiment. To prepare the substrate with the designated P-application rates, calcium superphosphate was ground to a powder and weighed, then thoroughly mixed with 1.5 kg of the aeolian sandy soil or loessial soil, and filled to non-transparent PVC tubes, which were of 11-cm diameter and 15-cm height and lined with a plastic bag inside; no drainage was allowed during the experiment. Tubes filled with the aeolian sandy soil were watered to 4% gravimetric soil water content (about 35% field capacity) daily, while tubes filled with the loessial soil were watered to 12% gravimetric soil water content (about 35% field capacity) every two days throughout the experiment, and all substrates prepared were incubated for eight days before sowing the seeds.
Medicago sativa L. cv Golden Empress, an introduced cultivar of alfalfa was used in this study. Seeds were sterilized in 30% (v/v) hydrogen peroxide (H2O2) solution for 5 min, rinsed with cold sterile water three times and then soaked in cold sterile water overnight (Kereszt et al. 2007). Nine seeds were sown in each pot and seedlings were thinned to three plants per pot two weeks after sowing. For each treatment, there were three pots in which plants were grown, and there were three control pots without growing plants. The experiment was carried out from April to July in 2015 in a transparent rain shelter at ambient temperature in the Institute of Soil and Water Conservation (34°16′33″N, 108°04′13″E), Yangling, Shaanxi, China.
Collection and analyses of rhizosphere carboxylates
When plants were harvested 110 days after sowing, shoots were severed at the base first, then the plastic bags were lifted from the pots to separate the roots from soil. Soil that was still attached to the roots after gently shaking was defined as rhizosphere soil (Veneklaas et al. 2003). For each pot with plants, about 1.0 g fresh roots and rhizosphere soil was transferred to a beaker containing 20 mL of 0.2 mM CaCl2 solution. Roots were gently stirred in the solution for about 20 min to ensure cell integrity and to remove the rhizosphere soil as much as possible. A 1-mL subsample of the rhizosphere extract was filtered using a 0.22-μm syringe filter into a 1-mL HPLC vial, then acidified with one drop of concentrated phosphoric acid, and kept in a − 20 °C freezer until HPLC analysis. pH of the remaining extract in the beaker was measured. Carboxylates in the rhizosphere extracts were analyzed using a Waters E2695 HPLC equipped with a Waters 2998 detector and Waters X select columns T3 (Waters, Milford MA, USA). The working standards included oxalic, malic, citric, and malonic acids to identify carboxylates at 210 nm (Cawthray 2003). Amounts of rhizosphere carboxylates were expressed per unit root dry mass (RDM), and per unit root length (RL) as well.
Measurement of bulk soil pH
When plants were harvested, bulk soil samples were collected from each pot in which plants were grown, and from each control pot without growing plants. All bulk soil samples were air-dried and passed through a 2-mm sieve, and their pH in a 1:5 soil:water suspension was measured using a pH meter (Little 1992).
Measurement of plant biomass and root morphology
Roots used for rhizosphere carboxylate collection and those not extracted were washed separately, first with tap water then with deionized water to remove soil particles, and scanned separately using an Epson Perfection V750 Pro scanner. Root length (RL) and root surface area (RSA) were measured using the WinRHIZO image analysis system (Regent Instruments, Quebec, Canada). Shoots and non-extracted roots were oven-dried at 60 °C for 48 h, extracted roots were oven-dried at 60 °C for 24 h, and weighed separately. Total RDM, RL and RSA was the sum of the respective parameter of the extracted roots and non-extracted roots, and specific root length (SRL) was calculated as total root length divided by total RDM. Root mass ratio (RMR) was calculated as total RDM divided by the sum of total RDM and shoot dry mass (SDM).
Analyses of mineral nutrients in plants
Oven-dried shoots and non-extracted roots were finely ground. For each sample, about 0.1 g subsample was weighed and digested with aqua regia and H2O2 (Sastre et al. 2002; Lomonte et al. 2008), and analyzed with Inductively Coupled Plasma Mass Spectrometry (ICP-MS) NexION 300 (PerkinElmer, Inc., Waltham, MA, USA) for concentrations of P, K, Ca, Mg, sulfur (S), Fe, Mn, Cu, Zn, nickel (Ni), molybdenum (Mo), and boron (B). Total plant content of each element in each pot was calculated as the sum of shoot element content (SDM per pot × shoot element concentration) and root element content (RDM per pot × root element concentration). Uptake of each element per unit RDM or RL was calculated as total plant content of each element divided by RDM or RL, without considering the content of each element in seeds.
Calculation of P-uptake efficiency and P-utilisation efficiency
For each treatment with external P supply, P-uptake efficiency was calculated as the difference in total plant content of P between this treatment and the 0P treatment divided by the difference in the amount of P applied between this treatment and the 0P treatment (Pang et al. 2010a). P-utilisation efficiency, which is defined as plant biomass production per unit of P uptake (Richardson et al. 2011), was calculated as total plant dry mass (SDM plus RDM) per pot divided by total plant content of P per pot in this study.
Statistical analyses
For all parameters, including plant growth and root morphological parameters, concentrations of mineral nutrients in shoots and roots, total plant contents of mineral nutrients, P-uptake efficiency and P-utilisation efficiency, rhizosphere and bulk soil pH, amounts of rhizosphere carboxylates, and uptake of mineral nutrients per unit RL, the data of three replicates in each treatment were analyzed by performing a one-way (P-application rate) analysis of variance (ANOVA) for the aeolian sandy soil and the loessial soil separately, and the differences between treatments were determined to be significant when P < 0.05, according to the least significant difference (LSD) test for post-hoc means comparisons. For the loessial soil, the data of malonate were only available for one replicate of the 40P treatment; therefore, the 40P treatment was not included in post-hoc means comparisons of the one-way ANOVA for malonate. Correlations between concentrations of different mineral nutrients in shoots and between those in roots, correlations between total plant content of different mineral nutrients, correlations between total plant content of mineral nutrients and root morphological parameters, and correlations between uptake of mineral nutrients per unit RDM or RL and rhizosphere carboxylate amount per unit RDM or RL were analyzed using bivariate Pearson correlations, and the correlations were considered to be significant when P < 0.05. All statistical analyses were performed using the IBM SPSS Statistics 22.0 software package (IBM, Montauk, New York, USA).
Results
Plant growth
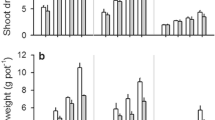
For the aeolian sandy soil, the 5P and 10P treatments did not result in a significant increase in shoot dry mass (SDM) (Fig. 1a); all ≥20P treatments significantly (P < 0.01) increased SDM, with SDM reaching a maximum when P was supplied at 20 μg P g−1 soil and remaining the same or decreasing slightly when P-application rate was further increased; SDM in all ≥20P treatments, except the 160P treatment, were markedly (P < 0.02) greater than those in the 5P and 10P treatments. For the loessial soil, SDM was always significantly (P < 0.001) increased by all ≥5P treatments, i.e. by 315–699%; SDM in all ≥20P treatments were similar, being significantly (P ≤ 0.002) greater than those in the 0P and 5P treatments; SDM in the 10P treatment was significantly (P = 0.040) greater than that in the 5P treatment, and markedly (P = 0.043) less than that in the 40P treatment, but was not significantly different from those in all other ≥20P treatments.

a Shoot dry mass, b Root dry mass, and c Root mass ratio of alfalfa grown in an aeolian sandy soil and a loessial soil supplied with different rates of phosphorus (P). Data are presented as means ± SE (n = 3). Ps-significance (P-value) of P treatment for plants grown in the aeolian sandy soil; LSDS-LSD of P treatment for plants grown in the aeolian sandy soil. P L -significance of P treatment for plants grown in the loessial soil; LSDL-LSD of P treatment for plants grown in the loessial soil
Root dry mass (RDM) of plants grown in the aeolian sandy soil was not significantly affected by any of the ≤20P treatments (Fig. 1b), but was significantly (P ≤ 0.01) increased by all ≥40P treatments, by 74% on average; no significant difference in RDM was found between any two of the ≥5P treatments. For the loessial soil, RDM was always significantly (P ≤ 0.01) increased when P was applied, being 4.1–7.9 times greater than in the 0P treatment; RDM reached a maximum at 10P, and decreased slightly when more P was supplied, with RDM being similar in all ≥10P treatments, but only the 10P treatment showed significantly (P = 0.017) more RDM than the 5P treatment.
Root mass ratio (RMR) of plants grown in the aeolian sandy soil and loessial soil was 0.42–0.54 and 0.34–0.48, respectively (Fig. 1c). For both the aeolian sandy soil and the loessial soil, there was no significant difference in RMR between any two treatments, except that the 10P treatment showed a markedly (P = 0.038) greater RMR than the 40P treatment for the loessial soil.
Root morphology
For plants grown in the aeolian sandy soil, root length (RL) was significantly increased by the 80P and 160P treatments (P = 0.022 and 0.027, respectively), but was not considerably affected by any of the ≤40P treatments; there was no significant difference in RL between any two of the ≥5P treatments (Fig. 2a). When P was applied to the loessial soil, RL was considerably (P < 0.01) increased, i.e. by 1.2–2.2 times, with all ≥5P treatments having similar RL.

a Root length, b Root surface area, and c Specific root length of alfalfa grown in an aeolian sandy soil and a loessial soil supplied with different rates of phosphorus (P). Data are presented as means ± SE (n = 3). Ps-significance (P-value) of P treatment for plants grown in the aeolian sandy soil; LSDS-LSD of P treatment for plants grown in the aeolian sandy soil. P L -significance of P treatment for plants grown in the loessial soil; LSDL-LSD of P treatment for plants grown in the loessial soil
Root surface area (RSA) of plants grown in the aeolian sandy soil was significantly (P < 0.04) increased by all ≥40P treatments; there was no significant difference in RSA between any two of the ≥5P treatments, except that the 80P treatment showed a significantly (P = 0.046) greater RSA than the 5P treatment (Fig. 2b). For plants grown in the loessial soil, RSA in all ≥5P treatments was significantly (P ≤ 0.006) greater than that in the 0P treatment, it increased with increasing P-application rate from 0P to 10P, but slightly decreased when more P was applied, with all ≥5P treatments showing similar RSA, except that the 10P treatment showed a significantly (P = 0.020) greater RSA than the 5P treatment.
There was no significant difference in specific root length (SRL) between any two treatments for plants grown in the aeolian sandy soil (Fig. 2c). For plants grown in the loessial soil, SRL was significantly (P ≤ 0.001) reduced by all ≥5P treatments, i.e. by up to 60–70% compared with the 0P treatment; there was no significant difference in SRL between any two of the ≥5P treatments.
Amounts of rhizosphere carboxylates
For the aeolian sandy soil, the amount of oxalate, malate, malonate, and the total amount of rhizosphere carboxylates measured relative to RDM was 9–23, 50–104, 32–70, and 103–191 μmol g−1 RDM, respectively (Fig. 3a); there was no significant difference between treatments in the amount of any rhizosphere carboxylate measured relative to RDM, except that the amount of oxalate relative to RDM in the 20P and 160P treatments was significantly greater than that in the 10P treatment (P = 0.029 and 0.036, respectively). For the loessial soil, the amount of oxalate, malate and malonate, and the total amount of rhizosphere carboxylates measured relative to RDM was 23–53, 19–67, 2–103, and 55–224 μmol g−1 RDM, respectively; there was no significant difference between treatments in the amounts of oxalate and malate relative to RDM; the 5P, 10P, 20P, and 80P treatments showed significantly (P < 0.04) less malonate than the 0P treatment, but all ≥5P treatments (except the 40P treatment) showed similar amounts of malonate; the total amounts of rhizosphere carboxylates measured in all ≥5P treatments were similar and significantly (P < 0.05) less than that in the 0P treatment.

a Amounts of carboxylates recovered in 0.2 mM CaCl2 extracts from the rhizosphere of alfalfa grown in an aeolian sandy soil and a loessial soil supplied with different rates of phosphorus (P), expressed on a root dry mass basis. b Amounts of rhizosphere carboxylates expressed on a root length basis. Data are presented as means ± SE (n = 3). Ps ox , Ps ma , Ps mo , and Ps t was the significance (P-value) of P treatment for the amount of oxalate, malate, malonate, and the total amount of rhizosphere carboxylates measured for the aeolian sandy soil; LSDSox, LSDSma, LSDSmo, and LSDSt was LSD of P treatment for the amount of oxalate, malate, malonate, and the total amount of rhizosphere carboxylates measured for the aeolian sandy soil. P Lox , P Lma , P Lmo , and P Lt was significance of P treatment for the amount of oxalate, malate, malonate, and the total amount of rhizosphere carboxylates measured for the loessial soil; LSDLox, LSDLma, LSDLmo, and LSDLt was LSD of P treatment for the amount of oxalate, malate, malonate, and the total amount of rhizosphere carboxylates measured for the loessial soil
For the aeolian sandy soil, the amount of oxalate, malate and malonate, and the total amount of rhizosphere carboxylates measured relative to RL was 0.06–0.14, 0.31–0.58, 0.19–0.45 and 0.64–1.10 μmol m−1 RL, respectively (Fig. 3b); there was no significant difference between treatments in the amount of any rhizosphere carboxylate measured relative to RL, except that the amount of oxalate relative to RL in the 20P treatment was significantly (P = 0.037) greater than that in the 10P treatment. For the loessial soil, the amount of oxalate, malate and malonate, and the total amount of rhizosphere carboxylates measured relative to RL was 0.16–0.34, 0.11–0.50, 0.01–0.70, and 0.32–1.53 μmol m−1 RL, respectively; no significant difference between treatments in the amount of oxalate and malate relative to RL was found; the 5P, 10P, and 80P treatments showed significantly (P < 0.05) less malonate than the 0P treatment, but all ≥5P treatments (except the 40P treatment) showed similar amounts of malonate; the total amounts of rhizosphere carboxylates measured in all ≥5P treatments were similar, and all except that in the 20P treatment were significantly (P < 0.05) less than that in the 0P treatment.
Bulk soil and rhizosphere pH
For the aeolian sandy soil, bulk soil in pots with plants had the highest pH, followed by the soil in control pots without plants, and the rhizosphere pH was the lowest (Fig. 4). The 5P treatment did not significantly affect soil pH of the control pots without plants, but all ≥10P treatments markedly (P ≤ 0.002) reduced the pH, i.e. by 0.24–0.96 unit, with the pH decreasing with increasing P-application rate, and the 160P treatment reducing the pH most (P < 0.001). Bulk soil pH in pots with plants was not significantly affected by the 5P and 40P treatments, but was markedly (P < 0.05) reduced (by 0.27–0.49 unit) by all other ≥10P treatments, with the 10P, 20P, and 80P treatments showing similar bulk soil pH with both the 40P and 160P treatments, but the 160P treatment showing a significantly (P = 0.013) lower bulk soil pH than the 40P treatment did. There was no significant difference in rhizosphere pH between any two treatments.

Rhizosphere and bulk soil pH of an aeolian sandy soil and a loessial soil supplied with different rates of phosphorus (P). SR-rhizosphere aeolian sandy soil, LR-rhizosphere loessial soil, SB-bulk aeolian sandy soil in pots with plants, LB-bulk loessial soil in pots with plants, SC-aeolian sandy soil in control pots without plants, LC-loessial soil in control pots without plants. Data are presented as means ± SE (n = 3). P SR , P SB , and P SC was significance (P-value) of P treatment for SR, SB, and SC; LSDSR, LSDSB, and LSDSC,was LSD of P treatment for SR, SB, and SC. P LR , P LB , and P LC was significance (P-value) of P treatment for LR, LB, and LC; LSDLR, LSDLB, and LSDLC,was LSD of P treatment for LR, LB, and LC
For the loessial soil, soil in the control pots without plants and bulk soil in pots with plants showed a similar pH, which was much higher than the rhizosphere pH (Fig. 4). Soil pH of the control pots without plants was not significantly changed by the 5P and 80P treatments, but was markedly (P ≤ 0.002) affected by all other ≥10P treatments, with the 10P, 20P, and 40P treatments increasing the pH significantly to similar degrees, i.e. by 0.16–0.20 unit, and the 160P treatment showing the lowest pH among treatments (P < 0.001). Bulk soil pH in pots with plants was only significantly affected by the 160P treatment (P = 0.040); there was no significant difference in bulk soil pH between any two of the ≥5P treatments, except that the 80P and 160P treatments showing considerably lower pH than the 5P treatment (P = 0.050 and 0.026, respectively). Rhizosphere pH was not significantly different between any two treatments.
Total contents of mineral nutrients in plants
For plants grown in the aeolian sandy soil, there were significant differences among treatments in total contents of all mineral nutrients measured (Fig. 5). Compared with the 0P treatment, total P content was not significantly affected by the 5P and 10P treatments (Fig. 5a), but was considerably (P ≤ 0.001) increased (by 1.6–2.6 times) by all ≥20P treatments, which also showed significantly (P ≤ 0.034) greater total P contents than the 5P and 10P treatments. The 10P treatment did not influence total K content significantly (Fig. 5b), but the 5P and all ≥20P treatments markedly (P ≤ 0.043) increased total K content to similar degrees, i.e. by 39–51%. Neither total Ca content nor total Mg content was considerably affected by the 5P and 10P treatments (Fig. 5c and d), but both were increased by all ≥20P treatments (P ≤ 0.003 for Ca, and P ≤ 0.006 for Mg) to similar degrees, i.e. by 82–90% for Ca and by 86–111% for Mg; the 20P and 40P treatments showed significantly (P ≤ 0.036) greater total Ca contents than the 5P and 10P treatments did, and the 80P and 160P treatments showed considerably greater total Ca contents than the 10P treatment did (P = 0.040 and 0.038, respectively); total Mg contents in all ≥20P treatments were significantly (P ≤ 0.041) greater than that in the 5P treatment; those in the 40P and 80P treatments were also markedly greater than that in the 10P treatment (P = 0.024 and 0.026, respectively). Total S contents were significantly (P ≤ 0.024) greater in all ≥10P treatments than in the 0P treatment (Fig. 5e), with all ≥20P treatments showing similar total S contents, which were significantly (P ≤ 0.018) greater than that in the 5P treatment. Total Fe contents in the 20P and 40P treatments were significantly greater than that in the 0P treatment (P = 0.049 and 0.036, respectively) (Fig. 5f), but there was no significant difference in total Fe content between any other two treatments. Total contents of Mn, Zn and Ni were not significantly affected by the 5P and 10P treatments (Fig. 5g, i, and j), but were all markedly increased by all ≥20P treatments (P ≤ 0.015 for Mn, P ≤ 0.041 for Zn, and P ≤ 0.030 for Ni), with no significant difference between any two of the ≥20P treatments. There was no significant difference in total Cu content between treatments, except that the 160P treatment showed a significantly greater total Cu content than the 0P, 10P and 80P treatments (P = 0.030, 0.010, and 0.041, respectively) (Fig. 5h). Total Mo content was only considerably (P = 0.013) affected by the 40P treatment (Fig. 5k), which showed a markedly (P ≤ 0.044) greater total Mo content than all other treatments, except the 20P treatment. Total B content was not significantly affected by the 5P and 10P treatments (Fig. 5l), but was markedly (P ≤ 0.003) increased by all ≥20P treatments, for which total B content generally increased with increasing P-application rate, being the greatest in the 160P treatment and significantly (P ≤ 0.046) greater than those in the 5P and 10P treatments.

Total plant content of mineral nutrients of alfalfa grown in an aeolian sandy soil and a loessial soil supplied with different rates of phosphorus (P). Data are presented as means ± SE (n = 3). Ps-significance (P-value) of P treatment for plants grown in the aeolian sandy soil; LSDS-LSD of P treatment for plants grown in the aeolian sandy soil. P L -significance of P treatment for plants grown in the loessial soil; LSDL-LSD of P treatment for plants grown in the loessial soil
For plants grown in the loessial soil, total contents of all mineral nutrients measured were significantly affected by P-application rate (Fig. 5). Total P content was significantly (P ≤ 0.014) increased by all ≥5P treatments, i.e. by 5.6–20.3 times, reaching a maximum at 160P (Fig. 5a); total P contents in the 20P, 40P and 80P treatments were similar, being markedly (P ≤ 0.011) greater than those in the 5P and 10P treatments, which were not markedly different. Total K content was markedly (P ≤ 0.023) increased by all ≥5P treatments, i.e. by 3.1–7.2 times (Fig. 5b); total K contents in the 10P, 40P, 80P and 160P treatments were similar, being significantly (P ≤ 0.038) greater than that in the 5P treatment; total K content in the 20P treatment was not significantly different from that in any other of the ≥5P treatments. Total Ca content was considerably (P ≤ 0.008) increased, i.e. by 3.5–6.5 times, when P was applied (Fig. 5c); total Ca contents in almost all ≥5P treatments were similar, except that the 20P treatment showed a markedly (P = 0.016) greater total Ca content than the 5P treatment did. Total Mg and S contents were significantly (P ≤ 0.001 for Mg, P ≤ 0.005 for S) increased by all ≥5P treatments, i.e. by 3.6–7.5 times for Mg and 3.1–6.7 times for S, respectively (Fig. 5d and e); for both Mg and S, the total contents in all ≥10P treatments were similar, being considerably (P ≤ 0.001 for Mg, P ≤ 0.003 for S) greater than that in the 5P treatment. Total Fe content was not significantly affected by the 5P and 160P treatments (Fig. 5f), but was considerably (P ≤ 0.046) increased by all other ≥10P treatments, i.e. 3.6–5.4 times; all ≥5P treatments showed similar total Fe contents. Total Mn content was significantly (P ≤ 0.016) affected by the 10P, 20P and 40P treatments (Fig. 5g), which increased total Mn content to similar degrees, i.e. by 5–7 times; there was no significant difference in total Mn content between any two of the ≥5P treatments, except that the 20P treatment had a considerably (P = 0.042) greater total Mn content than the 160P treatment. When P was supplied, total Cu content was always significantly (P ≤ 0.024) increased, i.e. by 1.9–3.9 times (Fig. 5h); the 10P, 40P and 80P treatments had markedly (P ≤ 0.041) greater total Cu contents than the 5P treatment; there was no significant difference in total Cu content between any two of the ≥10P treatments. Total Zn content was markedly (P ≤ 0.031) increased by all ≥10P treatments, i.e. by 3.3–6.0 times, but was not significantly affected by the 5P treatment (Fig. 5i); the 40P and 80P treatments showed considerably greater total Zn contents than the 5P treatment (P = 0.045 and 0.034, respectively); no significant difference was found between any two of the ≥10P treatments. Total Ni content was always significantly (P ≤ 0.018) increased when P was supplied, i.e. by 3.3–6.0 times (Fig. 5j); the 10P treatment showed a significantly (P = 0.044) greater total Ni content than the 5P treatment did; there was no significant difference in total Ni content between any two of the ≥10P treatments. Total Mo content was increased (P ≤ 0.047) by all ≥5P treatments, i.e. by 1.2–2.4 times (Fig. 5k); almost all ≥5P treatments showed similar total Mo contents, except that the 20P treatment had a significantly (P = 0.036) lower total Mo content than the 10P treatment did. The 5P treatment did not markedly affect total B content (Fig. 5l), but all ≥10P treatments significantly (P ≤ 0.001) increased total B content to similar degrees, i.e. by 10–14 times, and showed markedly (P ≤ 0.024) greater total B contents than the 5P treatment did.
Uptake of mineral nutrients per unit root length
For plants grown in the aeolian sandy soil, uptake of K, Ca, Mg, Fe, Mn, Cu and Zn per unit RL was not significantly affected by P-application rate (Fig. 6). Uptake of P per unit RL was significantly (P ≤ 0.016) increased by the 20P, 40P and 80P treatments, but was not affected by other treatments (Fig. 6a); the 20P, 40P and 80P treatments also showed significantly (P ≤ 0.033) greater P uptake per unit RL than the 5P treatment; there was no significant difference in P uptake per unit RL between any two of the ≥10P treatments, except that the 80P treatment showed a considerably (P = 0.041) greater P uptake per unit RL than the 10P treatment. Uptake of S per unit RL was not significantly different between any two treatments, except that the 20P treatment showed a significantly greater S uptake per unit RL than the 0P and 5P treatments (P = 0.021 and 0.026, respectively) (Fig. 6e). Uptake of Ni per unit RL was not significantly affected by P-application rate in almost all cases, except that the 20P treatment showed a markedly (P = 0.030) greater Ni uptake per unit RL than the 10P treatment (Fig. 6j). Uptake of Mo per unit RL was considerably (P ≤ 0.037) reduced by the 5P, 80P and 160P treatments, but was not markedly affected by other treatments (Fig. 6k); the 40P treatment showed a significantly (P ≤ 0.050) greater Mo uptake per unit RL than the 10P, 80P and 160P treatments; Mo uptake per unit RL in the 160P treatment was significantly (P = 0.047) less than that in the 5P treatment. Uptake of B per unit RL was considerably (P ≤ 0.046) increased by all ≥20P treatments to similar degrees (Fig. 6l), but was not significantly affected by the 5P and 10P treatments; all ≥40P treatments showed markedly (P ≤ 0.038) greater B uptake per unit RL than the 5P treatment; the 40P and 160P treatments also showed significantly greater B uptake per unit RL than the 10P treatment (P = 0.035 and 0.003, respectively).

Uptake of mineral nutrients per unit root length by alfalfa grown in an aeolian sandy soil and a loessial soil supplied with different rates of phosphorus (P). Data are presented as means ± SE (n = 3). Ps-significance (P-value) of P treatment for plants grown in the aeolian sandy soil; LSDS-LSD of P treatment for plants grown in the aeolian sandy soil. P L -significance of P treatment for plants grown in the loessial soil; LSDL-LSD of P treatment for plants grown in the loessial soil
For plants grown in the loessial soil, uptake of all mineral nutrients measured, except Mo, per unit RL were significantly affected by P-application rate (Fig. 6). The 5P and 10P treatments did not result in significant change in P uptake per unit RL (Fig. 6a), but all ≥20P treatments significantly (P ≤ 0.020) increased P uptake per unit RL similarly, i.e. by 5.0–8.6 times, with the 160P treatment showing much greater P uptake per unit RL than the 5P and 10P treatments did (P = 0.004 and 0.008, respectively). The 5P treatment did not significantly affect K uptake per unit RL (Fig. 6b), but all ≥10P treatments markedly (P ≤ 0.042) increased K uptake per unit RL to similar degrees, i.e. by 80–200%. Uptake of Ca and Mg per unit RL was not markedly affected by the 5P treatment (Fig. 6c and d), but was significantly (P ≤ 0.037 for Ca, and P ≤ 0.016 for Mg) increased by all ≥10P treatments to similar degrees, i.e. by 1.1–2.0 times for Ca and 1.7–2.4 times for Mg, with the 20P and 160P treatments showing significantly greater Mg uptake per unit RL than the 5P treatment did (P = 0.046 and 0.045, respectively). Uptake of S per unit RL was significantly (P ≤ 0.023) increased by all ≥10P treatments to similar degrees, i.e. by 1.6–2.3 times (Fig. 6e), with the 160P treatment showing markedly (P = 0.038) greater S uptake per unit RL than the 5P treatment did. Uptake of Fe per unit RL was not considerably affected by P-application rate in almost all cases, except that the 20P treatment showed significantly (P = 0.023) greater Fe uptake per unit RL than the 0P treatment did (Fig. 6f). The difference in Mn uptake per unit RL was not significant between any two treatments, except that the 20P treatment showed a considerably greater Mn uptake per unit RL than the 0P, 80P and 160P treatments (P = 0.006, 0.039, and 0.047, respectively) (Fig. 6g). There was no significant difference in Cu uptake per unit RL between any two treatments, except that the 80P treatment showed a markedly (P = 0.049) greater Cu uptake per unit RL than the 0P treatment did (Fig. 6h). The difference in Zn uptake per unit RL was not significant between any two treatments, except that the 40P and 80P treatments showed considerably greater Zn uptake per unit RL than the 0P treatment did (P = 0.027 and 0.019, respectively) (Fig. 6i).The 5P and 160P treatments did not significantly affect Ni uptake per unit RL, but all other ≥10P treatments significantly (P ≤ 0.043) increased Ni uptake per unit RL to similar degrees, i.e. by 95–166% (Fig. 6j). Uptake of B per unit RL was not markedly affected by the 5P treatment, but was significantly (P ≤ 0.024) increased by all ≥10P treatments to similar degrees, i.e. by 2.9–5.3 times, with the 20P treatment showing a considerably (P = 0.004) greater B uptake per unit RL than the 5P treatment did (Fig. 6l).
P-uptake efficiency and P-utilisation efficiency
For the aeolian sandy soil, the 5P, 10P and 20P treatments showed similar P-uptake efficiency (Fig. 7a), which was significantly (P ≤ 0.020) greater than that in the 160P treatment;, the 20P treatment also showed a considerably (P = 0.036) greater P-uptake efficiency than the 80P treatment; P-uptake efficiency in the 40P treatment was not significantly different from that in any other treatments, and all ≥40P treatments had similar P-uptake efficiency. For the loessial soil, P-uptake efficiency generally decreased with increasing P-application rate (Fig. 7a), with the difference between any two treatments, except that between the 80P and 160P treatments, being significant (P ≤ 0.014).

P-uptake efficiency and P-utilisation efficiency of alfalfa grown in an aeolian sandy soil and a loessial soil supplied with different rates of phosphorus (P). Data are presented as means ± SE (n = 3). Ps-significance (P-value) of P treatment for plants grown in the aeolian sandy soil; LSDS-LSD of P treatment for plants grown in the aeolian sandy soil. P L -significance of P treatment for plants grown in the loessial soil; LSDL-LSD of P treatment for plants grown in the loessial soil
For the aeolian sandy soil, P-utilisation efficiency was not significantly affected by the 5P treatment (Fig. 7b), but was markedly (P ≤ 0.024) reduced by all ≥10P treatments, i.e. by 19–50%; the 5P treatment showed a considerably (P ≤ 0.011) greater P-utilisation efficiency than all ≥20P treatments; P-utilisation efficiency in the 10P treatment was markedly greater than those in the 40P and 80P treatments (P = 0.020 and 0.001, respectively); the 160P treatment had a markedly (P = 0.039) greater P-utilisation efficiency than the 80P treatment. For the loessial soil, P-utilisation efficiency was significantly (P ≤ 0.003) reduced by all ≥5P treatments (Fig. 7b), i.e. by 33–67%; the 5P treatment showed a markedly greater P-utilisation efficiency than the 20P and 160P treatments (P = 0.048 and 0.003, respectively); the 10P treatment had a significantly (P = 0.004) greater P-utilisation efficiency than the 160P treatment; P-utilisation efficiency in the 20P, 40P and 80P treatments were similar to those in the 10P and 160P treatments.
Correlations between parameters
There were numerous strong correlations between total plant content of mineral nutrients and root morphological parameters (Table 2). For the aeolian sandy soil, total plant contents of all mineral nutrients measured, except those of Cu and Mo, were significantly positively correlated with both RL (0.494 ≤ r ≤ 0.684, P ≤ 0.023) and RSA (0.521 ≤ r ≤ 0.742, P ≤ 0.015); there was no strong correlation between total plant content of any of the mineral nutrients studied and SRL. For the loessial soil, total plant contents of all mineral nutrients studied were significantly positively correlated with both RL (0.527 ≤ r ≤ 0.799, P ≤ 0.023) and RSA (0.465 ≤ r ≤ 0.860, P ≤ 0.023); of note, however, the correlations between total plant contents of all mineral nutrients studied and SRL turned out to be strongly negative (−0.790 ≤ r ≤ −0.574, P ≤ 0.007) rather than positive. Correlations between total plant contents of mineral nutrients per unit RDM and rhizosphere carboxylate amount per unit RDM were not significant in most cases (Online Resource 1); for both the aeolian sandy soil and loessial soil, there was no significant correlation between plant P content and rhizosphere carboxylate amount.
There were a number of significant correlations between concentrations of other mineral nutrients and P concentration in shoots and roots, and between total plant contents of other mineral nutrients and total plant P content (Table 3). In shoots of plants grown in the aeolian sandy soil, concentrations of Mg (r = 0.579 and P = 0.006) and B (r = 0.753 and P ≤ 0.001) significantly positively correlated with P concentration, but K concentration and P concentration showed a significant negative correlation (r = −0.479 and P = 0.028). In roots of plants grown in the aeolian sandy soil, Mg concentration was strongly positively (r = 0.649 and P = 0.001) correlated with P concentration, but concentrations of Ca (r = −0.449 and P = 0.041), Cu (r = −0.659 and P = 0.001) and Zn (r = −0.478 and P = 0.029) were significantly negatively correlated with P concentration. For the aeolian sandy soil, total plant contents of all other mineral nutrients measured, except those of Cu and Mo, were significantly positively (0.554 ≤ r ≤ 0.933, P ≤ 0.009) correlated with total plant P content. The concentration of any other mineral nutrients measured in shoots of plants grown in the loessial soil did not correlate significantly with P concentration, but shoot Fe (r = −0.481 and P = 0.027), Zn (r = −0.434 and P = 0.049), and Ni (r = −0.718 and P ≤ 0.001) concentrations showed significant negative correlations with shoot P concentration (Table 3). For roots of plants grown in the loessial soil, there was no significant positive correlation between concentration of any other mineral nutrients measured and P concentration, but there were significant negative (−0.488 ≤ r ≤ −0.721, P ≤ 0.025) correlations between concentrations of all other mineral nutrients measured, except those of Mg and S, and P . For the loessial soil, total plant content of all other mineral nutrients, except those of Fe and Mo, showed significant positive (0.437 ≤ r ≤ 0.835, P ≤ 0.048) correlations with total plant P content.
Discussion
By carrying out a pot experiment growing alfalfa in two alkaline soils supplied with different rates of P when soil water supply was limited, parameters of plant growth, root morphology, and physiology were measured. The aims were to better understand rhizosphere processes and nutrient dynamics, to reveal possible mechanisms that could enhance P acquisition by alfalfa under P limitation, and investigate the effects of these mechanisms on the uptake of other mineral nutrients. The results and relevant implications are discussed below.
Plant growth in response to soil P
When soil P is limiting, plants tend to show a positive growth response to P addition (Elser et al. 2007). When alfalfa was grown in river sand at three P supply levels (3, 10 and 30 μg P g−1 dry sand), maximum plant growth (both SDM and RDM) was achieved at 30 μg P g−1 dry sand (Suriyagoda et al. 2010). Pang et al. (2010b) found that alfalfa achieved maximum growth (the sum of SDM and RDM) at 24 μg P g−1 dry sand when it was grown in river sand supplied with 0, 6, 12, 24, 48, 96, 192 and 384 μg P g−1 dry sand. By growing an introduced and a locally-selected cultivar of alfalfa in an alkaline loessial soil on the Loess Plateau supplied with four rates of P (0, 4.2, 8.4 and 16.8 μg P g−1 dry soil), Fan et al. (2015) found that plant growth was maximal at the highest P supply. In the present study, for both the aeolian sandy soil and the loessial soil, SDM was considerably increased in most cases when external P was supplied. Applying P significantly increased RDM of plants grown in the aeolian sandy soil when P supply was ≥40 μg P g−1, and considerably increased RDM of plants grown in the loessial soil when P supply was ≥5 μg P g−1. However, SDM did not further increase when P supply was >20 μg P g−1 in both soils, and RDM did not further increase when P supply was >40 μg P g−1 in the aeolian sandy soil and >10 μg P g−1 in the loessial soil. Pang et al. (2010b) also found that total DM of alfalfa was relatively constant when P supply was >24 μg P g−1 sand, and plants did not show visual P-toxicity symptoms at high P supply, similar to what was observed in our study. As the water supply in our study and that of Pang et al. (2010b) was low (about 35% and 50% of the field capacity, respectively), it is very likely that drought was another important factor limiting alfalfa growth (Suriyagoda et al. 2010, 2014; Fan et al. 2015). If the plants were provided with sufficient water, even greater SDM and RDM might have been achieved at higher P supply. Under the conditions of the current experiment, 20 μg P g−1 soil can be considered as optimal for shoot growth of alfalfa grown in both the aeolian sandy soil and the loessial soil, while 40, 80 and 160 μg P g−1 soil were supra-optimal.
Under P deficiency, root growth is less inhibited than shoot growth, and increased relative allocation to root growth, i.e. higher RMR, is thought to be beneficial for P acquisition (Lynch and Ho 2005; Marschner and Rengel 2012). However, we observed that RMR was not significantly affected by soil P levels, so our first hypothesis that plants would show larger RMR under low P supply was not supported. When chickpea (Cicer arietinum) was grown in soil supplied with four rates of P (0, 1, 5 and 40 μg P g−1 dry soil), Wouterlood et al. (2004b) found that RMR was only markedly reduced at 40 μg P g−1 dry soil, while there was no response of RMR at 1 and 5 μg P g−1 dry soil. Fan et al. (2015) observed that, for two alfalfa cultivars grown in an alkaline soil, higher P supply resulted in larger RMR under severe drought stress (30–35% field capacity). Our experiment was carried out under severe drought stress (35% field capacity), but showed different results of RMR in response to soil P supply from those of Fan et al. (2015), possibly because different alfalfa cultivars and soil types were used.
Relationships between root morphology, soil P, and uptake of mineral nutrients
The nutrient-acquisition capacity of plants, especially P-acquisition capacity, is often closely associated with root morphological parameters such as RL, RSA, SRL, and mycorrhizal associations (Vance et al. 2003; Lambers et al. 2006; Lynch 2007). For most plants grown in P-deficient soils, “root-foraging strategies” can enable the plant to explore large volumes of soil and thereby acquire more P from an enlarged P-depletion zone around the roots (Richardson et al. 2011). Plants tend to have greater SRL to maximize exploitation of soil P under P deficiency (Lambers et al. 2006; Pang et al. 2010a; Fan et al. 2015). In the present study, SRL of plants grown in the aeolian sandy soil was not significantly affected by P-application rate. For plants grown in the loessial soil, SRL was significantly reduced when P was supplied, but all ≥5P treatments had similar SRL. The results indicate that SRL was only responsive to changes in soil P when P supply was low; such response also depended on soil type, and the same may be applicable to RL and RSA. The lack of significant response in SRL to changes in soil P supply in the aeolian sandy soil was very likely because the soil water content was very low, and drought stress may have weakened the effects of soil P levels on SRL. It has been reported that SRL increases significantly with decreasing soil P availability, but decreases with decreasing water availability (Fan et al. 2015), and we would expect the same for the lack of responses in RL and RSA. Our second hypothesis that plants would show increased SRL under low P supply was only partly supported.
Total contents of most mineral nutrients in plants grown in the aeolian sandy soil, and those of all mineral nutrients in plants grown in the loessial soil were significantly positively correlated with RL and RSA, indicating that both RL and RSA played an important role in nutrient acquisition (Vance et al. 2003; Lambers et al. 2006). No significant positive correlation was found between SRL and total plant content of any of the mineral nutrients, suggesting that SRL was not a key factor in determining the uptake of these mineral nutrients. We did not investigate the mycorrhizal colonization status of the roots, and hence we do not know how much mycorrhizas contributed to the acquisition of P and other mineral nutrients.
Relationships between rhizosphere carboxylates, soil P, and uptake of mineral nutrients
Exudation of carboxylates is a P-mining strategy plants use to enhance desorption or solubilisation of P from sparingly-available sources in soil (Lambers et al. 2006; Richardson et al. 2011). A number of plant species, including alfalfa, show an increased exudation of carboxylates into the rhizosphere in response to P deficiency (Pang et al. 2010a; Suriyagoda et al. 2012a; Fan et al. 2015), as well as Fe and Zn deficiency (Broadley et al. 2010; Carvalhais et al. 2011). In the present study, the amounts of carboxylates recovered from the rhizosphere of the aeolian sandy soil did not vary considerably among treatments, but the amount of malonate and the total amount of rhizosphere carboxylates were greater when no P was supplied than when P was supplied to the loessial soil. Therefore, our third hypothesis, that plants would exude more carboxylates into the rhizosphere under small P supply, was partly supported. Pang et al. (2015) reported that amounts of rhizosphere carboxylates of three herbaceous perennial legumes either increased or did not change in response to increased P supply. Suriyagoda et al. (2012a) found that with increasing soil P supply, rhizosphere carboxylate amounts decreased for alfalfa, but first increased and then decreased for Kennedia nigricans (Fabaceae), and did not change for Ptilotus polystachyus (Amaranthaceae). Wouterlood et al. (2004a) found that amounts of carboxylates in the rhizosphere of lateral roots of chickpea increased during plant development, but were not correlated with soil or plants P status, and suggested that plant development is more likely to influence exudation of carboxylates into the rhizosphere of chickpea than P concentration, and carboxylate exudation in chickpea is a constitutive property evolved as an adaptation to the infertile soils of its native habitat. There are a number of studies suggesting that the qualitative and quantitative compositions of rhizosphere carboxylates are affected by abiotic factors such as pH, type and nutrient availability of soil, as well as biotic factors such as microorganisms in the soil, plant species and plant age (Mimmo et al. 2011; Suriyagoda et al. 2012b; Pang et al. 2015). Our results suggest that the response in carboxylate exudation to soil P supply depended on soil type and carboxylate type as well.
Carboxylates with three carboxyl groups, such as citrate, are more effective than carboxylates with two carboxyl groups, such as malate, oxalate, and malonate, at mobilizing P from soil particles (Oburger et al. 2009; Mimmo et al. 2011). In a number of studies, plants exude mainly citrate and malate under P deficiency, and citrate is the main contributor to the positive relationship between total P uptake by plants and rhizosphere carboxylates (Ryan et al. 2012; Kidd et al. 2016). However, in our study, citrate was below the detection limit, but oxalate, malate, and malonate were exuded in great amounts, with malate being the predominant carboxylate measured in most cases. Oxalate can mobilize P as effectively as citrate and malate, and synthesis of the two-carbon oxalate requires less carbon than that of the six-carbon citrate; therefore, plants with a large oxalate fraction of total root-exuded carboxylates have a low P-mobilisation cost (Dong et al. 2004; Hoffland et al. 2006; Abrahao et al. 2014). Malonate exuded by alfalfa in our study was considerable in most cases. Under P deficiency, malonate is the main carboxylate exuded by chickpea (Wouterlood et al. 2004b) and non-mycorrhizal Scots pine (Pinus sylvestris) (van Scholl et al. 2006). Pang et al. (2010a) reported that Kennedia prorepens and Macroptilium bracteatum reduced citrate and increased malonate exudation when P supply was lower. Fan et al. (2015) also found that alfalfa exuded more malonate than citrate. Kidd et al. (2016) reported that Ornithopus spp. had a mixture of citrate (about 40%) and malonate (about 60%). It has been suggested that malonate may have a role in protecting other carboxylates from microbial degradation (Li and Copeland 2000). However, Oburger et al. (2009) found that at high concentrations, malonate decreased sorption of citrate, malate and oxalate on soil particles, and malonate had an additive rather than synergistic effect on P mobilisation by these carboxylates. Furthermore, a high concentration of malonate in the rhizopshere might be sufficient to mobilize P when citrate concentration was not high enough (Kabir et al. 2015).
A few studies have reported positive relationships between amount of rhizosphere carboxylates and plant P uptake (Veneklaas et al. 2003; Ryan et al. 2012). The contribution of rhizosphere carboxylate exudation to P mobilisation varies among both soil type and carboxylate type (Oburger et al. 2009). The ability of rhizosphere carboxylates to mobilize P also depends partly on the amount and form of P present in soil (Lambers et al. 2002; Pang et al. 2015), soil water content (Suriyagoda et al. 2010; Suriyagoda et al. 2012a), and temperature (Suriyagoda et al. 2012b). The amount of carboxylates recovered from the rhizosphere of alfalfa in our study was much greater than that in the study of Pang et al. (2010a) and Fan et al. (2015) which was 25–40 and 6–14 μmol g−1 RDM, respectively, but there is no evidence that carboxylate exudation in alfalfa is constitutive. There was no strong correlation between total rhizosphere carboxylate amount per unit RDM or per unit RL and uptake of P per unit RDM or per unit RL (Online Resource 1), and there was no strong correlation between rhizosphere carboxylate amount per unit RDM or per unit RL and total plant P content (data not shown). However, significant positive correlations were observed for a few other mineral nutrients. In alkaline soils, rhizosphere carboxylates such as oxalate may not promote P mobilisation due to a high free Ca2+ concentration and the formation of sparingly-soluble calcium oxalate minerals (Wang et al. 2012; He et al. 2014). Therefore, rhizosphere carboxylate exudation may not be a key factor in determining the uptake capacity of P and other mineral nutrients by alfalfa under the current experimental conditions.
Roles of rhizosphere pH
Soil pH is a major factor determining the availability of mineral nutrients in soils (Lambers et al. 2008). In the present study, the pH drop of the bulk soil at high P supply was very likely the result of soil acidification caused by superphosphate application (Bolan et al. 2003). The loessial soil had a finer texture and a higher calcium carbonate content and thus greater pH-buffering capacity, so its pH may have been less affected than that of the aeolian sandy soil (Wang et al. 2015). Furthermore, the presence of plants could partly offset the effects of external P application on bulk soil pH. Hassan et al. (2012) reported that, when plants of five grain legumes were at flowering and maturity, pH of the bulk soil from the unplanted pots and the rhizosphere was significantly less than the initial pH (8.8) of a non-calcareous loamy sand with a P supply of 80 μg P g−1 soil. In our study, rhizosphere pH was always lower than bulk soil pH in pots with plants, very likely due to acidification resulting from release of protons during rhizosphere processes such as carboxylate exudation, greater uptake of cations relative to anions, and root respiration (Hinsinger et al. 2003). However, rhizosphere pH did not vary considerably among treatments for either the aeolian sandy soil or the loessial soil. The uptake of phosphate anions could also cause alkalinization of the rhizosphere, due to the net release of hydroxyl ions to balance the charge. It is likely that the alkalinization caused by phosphate uptake negated the acidification resulting from greater uptake of cations by more robust root systems at higher P supply (Hinsinger et al. 2003). Therefore, our fourth hypothesis that plants would acidify the rhizosphere under low P supply was not fully supported.
For many plant species, variation in pH induced by roots is considered to have the most significant effects on the availability of mineral nutrients such as P, Mn, Cu and Zn in the rhizosphere (Rengel and Marschner 2005; Marschner and Rengel 2012). Rhizosphere acidification could increase dissolution of poorly-available P forms such as Ca-P (Hinsinger et al. 2003). Furthermore, in the present study, rhizosphere pH was always neutral, ranging between 6.8 and 7.2, a range within which a few mineral nutrients such as P, K, Ca, Mg, and Mo are optimally available to plants (Lambers et al. 2008).
Uptake of mineral nutrients
Accumulation of one element often has consequences for the accumulation of others which seems to be partly driven by a need to maintain charge balance (Conn and Gilliham 2010; Aziz et al. 2015). When P (as an anion) is taken up in greater quantities, uptake of cations such as K, Ca, and Mg is also increased (Suriyagoda et al. 2012a; Aziz et al. 2015). In the present study, concentrations of some mineral nutrients in shoots and roots (Online Resource 2 and Online Resource 3), and total plant content of almost all mineral nutrients were significantly affected by soil P supply. The P concentration in plants was generally higher at higher P supply. For both the aeolian sandy soil and loessial soil, the correlations between concentrations of other mineral nutrients and P concentration in both shoots and roots were not significant in most cases, but the correlation between total plant contents of almost all other mineral nutrients was strongly and positively correlated with total plant P content. Therefore, our fifth hypothesis that plants would show increased uptake of other mineral nutrients when more P was taken up was substantially supported.
Conclusions
Plant growth, tissue P concentrations and total plant P content were increased when P was supplied. A P supply of 20 μg P g−1 soil to the aeolian sandy soil and the loessial soil should be optimal for shoot growth of alfalfa when soil water content is low. The response of root mass ratio and rhizosphere pH did not vary significantly with soil P supply. Specific root length, root length, and root surface area were only responsive to soil P when P supply was low, and such responses also depended on soil type. Rhizosphere carboxylates were exuded in relatively large amounts (130 μmol g−1 RDM on average for the total amount of carboxylates measured), mainly including malate, oxalate and malonate, and possibly contributed to P mobilisation in the loessial soil when soil P supply was low. The response in carboxylate exudation and carboxylate type to soil P supply depended on soil type. Rhizosphere pH was much lower than bulk soil pH and always neutral, and such a decrease in pH would enhance the availability of a range of mineral nutrients including P. Total plant contents of almost all other mineral nutrients were also increased when P was supplied, and positively correlated with total plant P content, mainly because increasing P supply promoted plant growth and increased the nutrient acquisition area of the root systems.
References
Abrahao A, Lambers H, Sawaya A, Mazzafera P, Oliveira RS (2014) Convergence of a specialized root trait in plants from nutrient-impoverished soils: phosphorus-acquisition strategy in a nonmycorrhizal cactus. Oecologia 176:345–355
Aziz T, Lambers H, Nicol D, Ryan MH (2015) Mechanisms for tolerance of very high tissue phosphorus concentrations in Ptilotus polystachyus. Plant Cell Environ 38:790–799
Bolan NS, Adriano DC, Curtin D (2003) Soil acidification and liming interactions with nutrient and heavy metal transformation and bioavailability. Adv Agron 78:215–272
Broadley MR, Rose T, Frei M, Pariasca-Tanaka J, Yoshihashi T, Thomson M, Hammond JP, Aprile A, Close TJ, Ismail AM, Wissuwa M (2010) Response to zinc deficiency of two rice lines with contrasting tolerance is determined by root growth maintenance and organic acid exudation rates, and not by zinc-transporter activity. New Phytol 186:400–414
Carpenter SR (2005) Eutrophication of aquatic ecosystems: Bistability and soil phosphorus. Proc Natl Acad Sci U S A 102:10002–10005
Carvalhais LC, Dennis PG, Fedoseyenko D, Hajirezaei M-R, Borriss R, von Wirén N (2011) Root exudation of sugars, amino acids, and organic acids by maize as affected by nitrogen, phosphorus, potassium, and iron deficiency. J Plant Nutr Soil Sci 174:3–11
Cawthray GR (2003) An improved reversed-phase liquid chromatographic method for the analysis of low-molecular mass organic acids in plant root exudates. J Chromatogr A 1011:233–240
Conn S, Gilliham M (2010) Comparative physiology of elemental distributions in plants. Ann Bot 105:1081–1102
Cordell D, White S (2015) Tracking phosphorus security: indicators of phosphorus vulnerability in the global food system. Food Secur 7:337–350
Cordell D, Drangert J-O, White S (2009) The story of phosphorus: global food security and food for thought. Glob Environ Change-Human Policy 19:292–305
Dakora FD, Phillips DA (2002) Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil 245:35–47
Deng L, Wang KB, Li JP, Shangguan ZP, Sweeney S (2014) Carbon storage dynamics in alfalfa (Medicago sativa) fields in the hilly-gully region of the Loess Plateau, China. Clean-Soil Air Water 42:1253–1262
Dong DF, Peng XX, Yan XL (2004) Organic acid exudation induced by phosphorus deficiency and/or aluminium toxicity in two contrasting soybean genotypes. Physiol Plant 122:190–199
Elser JJ, Bracken MES, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10:1135–1142
Fan JW, Du YL, Turner NC, Wang BR, Fang Y, Xi Y, Guo XR, Li FM (2015) Changes in root morphology and physiology to limited phosphorus and moisture in a locally-selected cultivar and an introduced cultivar of Medicago sativa L. growing in alkaline soil. Plant Soil 392:215–226
Fang Z, Shao C, Meng Y, Wu P, Chen M (2009) Phosphate signaling in Arabidopsis and Oryza sativa. Plant Sci 176:170–180
Fixen PE, Johnston AM (2012) World fertilizer nutrient reserves: a view to the future. J Sci Food Agric 92:1001–1005
Hassan HM, Marschner P, McNeill A, Tang CX (2012) Growth, P uptake in grain legumes and changes in rhizosphere soil P pools. Biol Fertil Soils 48:151–159
He H, Veneklaas EJ, Kuo J, Lambers H (2014) Physiological and ecological significance of biomineralization in plants. Trends Plant Sci 19:166–174
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237:173–195
Hinsinger P, Plassard C, Tang CX, Jaillard B (2003) Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: a review. Plant Soil 248:43–59
Hoffland E, Wei C, Wissuwa M (2006) Organic anion exudation by lowland rice (Oryza sativa L.) at zinc and phosphorus deficiency. Plant Soil 283:155–162
Hossner LR, Freeouf JA, Folsom BL (1973) Solution phosphorus concentration and growth of rice (Oryza sativa L.) in flooded soils. Soil Sci Soc Am J 37:405–408
Johnston AE, Poulton PR, Fixen PE, Curtin D (2014) Phosphorus: its efficient use in agriculture. Adv Agron 123:177–228
Kabir ME, Johansen C, Bell RW (2015) Subsoil rhizosphere modification by chickpea under a dry topsoil: implications for phosphorus acquisition. J Plant Nutr Soil Sci 178:904–913
Kereszt A, Li D, Indrasumunar A, Nguyen CDT, Nontachaiyapoom S, Kinkema M, Gresshoff PM (2007) Agrobacterium rhizogenes - mediated transformation of soybean to study root biology. Nat Protoc 2:948–952
Kidd DR, Ryan MH, Haling RE, Lambers H, Sandral GA, Yang ZJ, Culvenor RA, Cawthray GR, Stefanski A, Simpson RJ (2016) Rhizosphere carboxylates and morphological root traits in pasture legumes and grasses. Plant Soil 402:77–89
Lambers H, Juniper D, Cawthray GR, Veneklaas EJ, Martinez-Ferri E (2002) The pattern of carboxylate exudation in Banksia Grandis (Proteaceae) is affected by the form of phosphate added to the soil. Plant Soil 238:111–122
Lambers H, Shane MW, Cramer MD, Pearse SJ, Veneklaas EJ (2006) Root structure and functioning for efficient acquisition of phosphorus: matching morphological and physiological traits. Ann Bot 98:693–713
Lambers H, Thijs LP, Chapin FS (2008) Plant physiological ecology. Springer, New York
Lambers H, Hayes PE, Laliberte E, Oliveira RS, Turner BL (2015) Leaf manganese accumulation and phosphorus-acquisition efficiency. Trends Plant Sci 20:83–90
Li J, Copeland L (2000) Role of malonate in chickpeas. Phytochemistry 54:585–589
Little IP (1992) The relationship between soil pH measurements in calcium chloride and water suspensions. Aust J Soil Res 30:587–592
Lomonte C, Gregory D, Baker AJM, Kolev SD (2008) Comparative study of hotplate wet digestion methods for the determination of mercury in biosolids. Chemosphere 72:1420–1424
Lynch JP (2007) Roots of the second green revolution. Aust J Bot 55:493–512
Lynch JP, Ho MD (2005) Rhizoeconomics: carbon costs of phosphorus acquisition. Plant Soil 269:45–56
Marschner P, Rengel Z (2012) Nutrient availability in soils. In: Marschner P (ed) Mineral nutrition of higher plants, 3rd edn. Elsevier, Amsterdam
Mimmo T, Hann S, Jaitz L, Cesco S, Gessa CE, Puschenreiter M (2011) Time and substrate dependent exudation of carboxylates by Lupinus albus L. and Brassica napus L. Plant Physiol Biochem 49:1272–1278
Nunes FC, Miyazawa M, Pavan MA (2009) Organic acid effect on calcium uptake by the wheat roots. Braz Arch Biol Techn 52:11–15
Oburger E, Kirk GJD, Wenzel WW, Puschenreiter M, Jones DL (2009) Interactive effects of organic acids in the rhizosphere. Soil Biol Biochem 41:449–457
Page V, Weisskopf L, Feller U (2006) Heavy metals in white lupin: uptake, root-to-shoot transfer and redistribution within the plant. New Phytol 171:329–341
Pang JY, Ryan MH, Tibbett M, Cawthray GR, Siddique KHM, Bolland MDA, Denton MD, Lambers H (2010a) Variation in morphological and physiological parameters in herbaceous perennial legumes in response to phosphorus supply. Plant Soil 331:241–255
Pang JY, Tibbett M, Denton MD, Lambers H, Siddique KHM, Bolland MDA, Revell CK, Ryan MH (2010b) Variation in seedling growth of 11 perennial legumes in response to phosphorus supply. Plant Soil 328:133–143
Pang JY, Yang JY, Lambers H, Tibbett M, Siddique KHM, Ryan MH (2015) Physiological and morphological adaptations of herbaceous perennial legumes allow differential access to sources of varyingly soluble phosphate. Physiol Plant 154:511–525
Rengel Z, Marschner P (2005) Nutrient availability and management in the rhizosphere: exploiting genotypic differences. New Phytol 168:305–312
Richardson AE, Lynch JP, Ryan PR, Delhaize E, Smith FA, Smith SE, Harvey PR, Ryan MH, Veneklaas EJ, Lambers H, Oberson A, Culvenor RA, Simpson RJ (2011) Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 349:121–156
Ron Vaz MD, Edwards AC, Shand CA, Cresser MS (1993) Phosphorus fractions in soil solution: influence of soil acidity and fertiliser additions. Plant Soil 148:175–183
Ryan MH, Tibbett M, Edmonds-Tibbett T, Suriyagoda LDB, Lambers H, Cawthray GR, Pang J (2012) Carbon trading for phosphorus gain: the balance between rhizosphere carboxylates and arbuscular mycorrhizal symbiosis in plant phosphorus acquisition. Plant Cell Environ 35:2170–2180
Sastre J, Sahuquillo A, Vidal M, Rauret G (2002) Determination of cd, cu, Pb and Zn in environmental samples: microwave-assisted total digestion versus aqua regia and nitric acid extraction. Anal Chim Acta 462:59–72
van Scholl L, Hoffland E, van Breemen N (2006) Organic anion exudation by ectomycorrhizal fungi and Pinus sylvestris in response to nutrient deficiencies. New Phytol 170:153–163
Simpson RJ, Oberson A, Culvenor RA, Ryan MH, Veneklaas EJ, Lambers H, Lynch JP, Ryan PR, Delhaize E, Smith FA, Smith SE, Harvey PR, Richardson AE (2011) Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 349:89–120
Simpson RJ, Stefanski A, Marshall DJ, Moore AD, Richardson AE (2015) Management of soil phosphorus fertility determines the phosphorus budget of a temperate grazing system and is the key to improving phosphorus efficiency. Agric Ecosyst Environ 212:263–277
Suriyagoda LDB, Ryan MH, Renton M, Lambers H (2010) Multiple adaptive responses of Australian native perennial legumes with pasture potential to grow in phosphorus- and moisture-limited environments. Ann Bot 105:755–767
Suriyagoda LDB, Lambers H, Renton M, Ryan MH (2012a) Growth, carboxylate exudates and nutrient dynamics in three herbaceous perennial plant species under low, moderate and high phosphorus supply. Plant Soil 358:100–112
Suriyagoda LDB, Ryan MH, Renton M, Lambers H (2012b) Adaptive shoot and root responses collectively enhance growth at optimum temperature and limited phosphorus supply of three herbaceous legume species. Ann Bot 110:959–968
Suriyagoda LDB, Ryan MH, Renton M, Lambers H (2014) Plant responses to limited moisture and phosphorus availability: a meta-analysis. Adv Agron 124:143–200
Ulrich AE, Malley DF, Watts PD (2016) Lake Winnipeg Basin: advocacy, challenges and progress for sustainable phosphorus and eutrophication control. Sci Total Environ 542:1030–1039
Vance CP, Uhde-Stone C, Allan DL (2003) Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytol 157:423–447
Veneklaas EJ, Stevens J, Cawthray GR, Turner S, Grigg AM, Lambers H (2003) Chickpea and white lupin rhizosphere carboxylates vary with soil properties and enhance phosphorus uptake. Plant Soil 248:187–197
Vu DT, Tang C, Armstrong RD (2008) Changes and availability of P fractions following 65 years of P application to a calcareous soil in a Mediterranean climate. Plant Soil 304:21–33
Wang L, Ruiz-Agudo E, Putnis CV, Menneken M, Putnis A (2012) Kinetics of calcium phosphate nucleation and growth on calcite: implications for predicting the fate of dissolved phosphate species in alkaline soils. Environ Sci Technol 46:834–842
Wang X, Tang C, Mahony S, Baldock JA, Butterly CR (2015) Factors affecting the measurement of soil pH buffer capacity: approaches to optimize the methods. Eur J Soil Sci 66:53–64
Wouterlood M, Cawthray GR, Scanlon TT, Lambers H, Veneklaas EJ (2004a) Carboxylate concentrations in the rhizosphere of lateral roots of chickpea (Cicer arietinum) increase during plant development, but are not correlated with phosphorus status of soil or plants. New Phytol 162:745–753
Wouterlood M, Cawthray GR, Turner S, Lambers H, Veneklaas EJ (2004b) Rhizosphere carboxylate concentrations of chickpea are affected by genotype and soil type. Plant Soil 261:1–10
Yanai RD (1991) Soil solution phosphorus dynamics in a whole-tree-harvested northern hardwood forest. Soil Sci Soc Am J 55:1746–1752
Acknowledgements
This work was financially supported by the National Natural Science Foundation of China (41301570), the West Light Foundation of the Chinese Academy of Sciences, and Fundamental Research Funds for the Central Universities.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Tim S. George.
Electronic supplementary material
ESM 1
(DOCX 12.7 kb)
Rights and permissions
About this article

Cite this article
He, H., Peng, Q., Wang, X. et al. Growth, morphological and physiological responses of alfalfa (Medicago sativa) to phosphorus supply in two alkaline soils. Plant Soil 416, 565–584 (2017). https://doi.org/10.1007/s11104-017-3242-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-017-3242-9