
Abstract
The search of alternative therapeutic agents for the use of cancer patients has dramatically expanded. Natural products are especially in focus since their structures already function in nature and are more likely to be potent with fewer side effects. Phloretin is a natural product that has been studied for a wide variety of pharmacological actions against human malignancies. This systematic review aims to present up-to-date critical and comprehensive information on the anti-cancer ability of Phloretin with all associated molecular and cellular mechanisms in various forms of cancers. Data retrieved according to PRISMA guidelines from Science Direct, PubMed, and Scopus searching servers by using keywords including Phloretin, cancer name, synergistic, resistance and Pharmacokinetics property was analyzed via some in silico tools. This systematic review comprised 127 articles from different types of study, where Phloretin is hypothesized to be effective against 20 various forms of cancer. Phloretin has been found to inhibit cancer initiation and progression by modulating many imbalanced signalling pathways, including apoptosis, autophagy, necrosis, metastasis, angiogenesis, cell proliferation, glucose absorption, oxidative stress, inflammation, DNA damage, and many other pathways. This wide range of activity may be due to the structural targeting of numerous proteins including, Bcl-2, Bax, Bak, Bad, caspase, cyclins (B1, D1, E) and CDKs (4, 6,7) p18, p21, p27, p53, MMP-2, MMP- 8, MMP-9, Wnt/-catenin, PARP, TNF-α, NF-κB, IκB kinase, IL-1β, TNF-α, phospho-Akt, phosphor-p65, NF-κB, PI3K/Akt, MAPK/ERK, p-mTOR. The introduction of nano-technology-based strategies can improve the efficacy of Phloretin for cancer treatment. Existing evidence shows that Phloretin has synergistic effects with other natural compounds and conventional drugs, and this mechanism assists in reversing the resistance of anticancer drugs by regulating resistance-related proteins. However, Phloretin possesses favorable pharmacokinetic properties with low toxicity in the human body by in silico methods. Therefore, Phloretin could be a potential anti-cancer drug against numerous cancer treatment by mitigating it's toxic effect and enhancing efficacy using nano-technology-based strategies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cancer is a common disease as well as a significant global health issue (Siegel et al. 2023). Although the coronavirus disease 2019 (COVID-19) pandemic created delays in cancer patient detection, new cancer cases and deaths have gradually increased in the last two years(Siegel et al. 2023). The causative agent of cancer is known to all, but therapeutic options are still limited. Some conventional therapies effectively treat numerous cancer patients (Debela et al. 2021; Karpuz et al. 2018; Nobili et al. 2009). Nevertheless, a few of these therapeutic approaches have several drawbacks and restrictions, including several side effects; anaemia, alopecia, poor efficacies, drug resistance, and also the high cost, and unavailability. Therefore, using newly discovered compounds derived from plant sources can provide an innovative and trustworthy therapeutic element for healing a wide variety of human cancers (Shrihastini et al. 2021). Using natural products for cancer treatment is not a new approach. Kurupp et al. reported that a total of 3000 species have anti-cancer characteristics, with plants accounting for more than 60% of presently utilized anti-cancer medicines. (Kuruppu et al. 2019). This is because natural compounds are safe, non-toxic, cost-effective, and readily available from villages to cities and underdeveloped to developed countries (Rodriguez et al. 2015). Our latest study reviewed that natural products regulate cancer progression by targeting apoptosis cell cycle and proliferation, metastasis, angiogenesis, oxidative markers, mammosphere formation, inflammation, regulation enzymatic functions, cell signalling, epigenetics control, and activating immune system (Sohel et al. 2022a, b, c, d; Biswas et al. 2022; Paul et al. 2021; Morshed et al. 2023; Sohel et al. 2023).
Phloretin, chemically known as 3-(4-hydroxyphenyl)-1-(2,4,6-tri hydroxyphenyl) propane-1-one, is a naturally occurring functional phytoestrogen and to the isoflavone category, that is generally found in Rosaceae plants(“Phloretin | C15H14O5—PubChem,”.; C. Wu et al. 2009a, b; Nakhate et al. 2022; Picinelli, Dapena, and Mangas 1995). This compound has several pharmacological activities, including anti-oxidative(Rezk et al. 2002), anti-inflammatory(Chang et al. 2012), anti-microbial(Barreca et al. 2014), anti-allergic(Huang et al. 2017), anti-thrombotic (Mariadoss et al. 2019a, b), neuroprotective(Barreca et al. 2017), and hepatoprotective(Lu et al. 2017) due to strong interaction with multiple receptors(Mariadoss et al. 2019a, b). Medicinal chemists can modify its chemical structure and develop analogs with improved pharmacological properties, potentially leading to the development of more potent and selective anticancer drugs. Regarding cancer, previous studies have shown that consumption of Phloretin as a dietary food reduces the risk of certain cancers, such as lung(Min et al. 2015), oral (Yang et al. 2020), esophageal(Duan et al. 2017), gastric, colon(Kim et al. 2022), liver(Min et al. 2015), breast (He et al. 2020.),pancreartic(Brij B Patel et al. 2016), kidney (J. Kim et al. 2018), glioblastoma(Liu et al. 2016), ovarian(Xintaropoulou et al. 2015) cervical(Hsiao et al. 2019), bladder (Hui 2020), myeloma (McBrayer et al. 2012), skin(Casarini et al. 2020), melanoma(KoBoRI et al. 1999) leukemia(KoBoRI et al. 1999), thyroid(Ma et al. 2022), osteosarcoma(Ogawa et al. 2021), oesophagal (Duan et al. 2017) and prostate cancer (Yang et al. 2022). This wide range of anti-cancer activities is mediated by the interaction of Phloretin with some regulatory molecules and modulators. Phloretin is typically thought of as an antioxidant, its impacts on cancer cells can be complicated. Through selective cytotoxicity, pro-oxidant action, and signaling pathway modification, but it can cause oxidative damage to cancer cells. Due to increased metabolic activity and mitochondrial malfunction, cancer cells frequently have greater ROS level than normal cells. Phloretin can take advantage of this vulnerability by raising ROS levels in cancer cells to levels that exceed their antioxidant tolerance and result in oxidative damage. Phloretin also has the ability to interfere with various signaling pathways and cause oxidative damage by affecting the redox balance within cancer cells. Although some experimental studies of Phloretin, with numerous cancer types, have been in various in vitro and pre-clinical trials, the overall anti-cancer profile against numerous types of human malignancies has not been reviewed yet.
Thus, in different study models, authors emphasized Phloretin's total anti-cancer impact with cellular and molecular mechanisms against numerous cancers. Moreover, we targeted unravelling synergistic profiles with other natural products, proteins, and traditional drugs currently used and reversed the efficacy of existing drugs by alleviating multi-drug resistance. Additionally, this article focused on an up-to-date overview of developing nano-strategies for better efficacies, with some additional information, including a pharmacokinetics study and toxicity profile. So, in general, the information covered here will encourage potent scholars to design novel and potentially clinically proven drugs to fight against cancerous neoplasm.
Methodology
Literature research on phloretin and malignancies
According to PRISMA guidelines, the current systematic review was carried out by using several databases, including Science Direct, PubMed, Scopus, and Google scholar (Dewanjee et al. 2023).
Inclusion criteria
We used different study models, including in silico, in vitro and in vivo, where only the English language was considered for evaluating the anticancer activity of phloretin.
Exclusion criteria
The following exclusion criteria were applied “ (I) conference paper, unpublished paper, dissertation, and thesis book (II) Non-English paper (III) Review article, systematic review, meta-analysis, letter (IV) Original studies without the use of tumour cell lines or animal tumor models.”
Data extraction
At first, 5161 articles were searched by searching protocol, 2900 articles were excluded due to initial selection, 28 review articles were omitted, 852 articles were excluded due to non-relevance and 39 non-English articles were disqualified. Out of 393 selected, 97 were excluded due to whole plant extract testing, and 41 more were removed due to discussion of other biological activities rather than cancer. Finally, data was retrieved from 137 papers, and the literature search and selection method is shown in a flowchart (Fig. 1).

The PRISMA flow chart showing the method used to choose the studies that were included
Pharmacokinetics study analysis
In-silico approaches conducted pharmacokinetics or ADME/Tox prediction through computational tools such as Schrodinger's QuickPro modules, online accessible server admetSAR, and SwissADME were used (Sohel, Sultana, Sultana, Al Amin, et al. 2022).
Result
Sources of phloretin
Phloretin is a natural compound found in various plant sources, particularly in apples and apple-related products like apple juice, apple cider, and apple peel extracts (Szabo et al. 2022). Apple contains up to 2 g of total phenolic content (TPC) per kilogram of wet weight or approximately 400 mg of total phenols per apple (FΑΟ 2013; Choi 2019). Although apples are the most well-known source, small amounts of phloretin can also be found in other fruits like cherries, strawberries, and raspberries, although at much lower levels (Hilt et al. 2003) (Wu et al. 2009a, b; Nakhate et al. 2022; Picinelli, Dapena, and Mangas 1995). Phloretin can be found in a few different plant species, but its concentration tends to be lower than in apples. Malus doumeri, M. domestica Populus candicans are the common plant species for Phloretin(“Phloretin | C15H14O5—PubChem”). Additionally, the manchurian apricot, a fruit in the genus Prunus, contains Phloretin in a small amount (“Manchurian Apricot facts and health benefits,”). Moreover, as a natural sweetener, Phloretin can be found in Lithocarpus Polystachyus and Rehd. (Shang et al. 2022). Moreover, its derivatives, including Phloretin 3′,5′-di-C-glc, phloretin-2'-glucoside, Phloretin 2′-O-xyloglucoside, Phloretin 2′-O-glucuronide are found in various plant sources (Slimestad et al. 2008) (Hagl et al. 2011). Furthermore, some derivatives, food supplements, topical creams, and Dietary Sources contain a small amount of Phloretin. Unlocking the secret source of Phloretin is tabulated in Table 1.
Chemistry of phloretin
The chemical structure of phloretin consists of three main components: a dihydrochalcone core, a bicyclic ring system commonly referred to as A and B and two phenolic hydroxyl groups at positions 2, 4, and 6 (Minsat et al. 2021). Other functional groups, including the carbonyl group (C = O) and ether linkage (C–O–C), contribute to its overall structure. The chemical formula of phloretin is C15H14O5, where IUPAC name is 3-(4-hydroxyphenyl)-1-(2,4,6-tri hydroxyphenyl) propane-1-one and systematic name is 2',4,6'-trihydroxy-3-(4-hydroxyphenyl) propiophenone. It has 20 heavy atom counts, four donor and rotatable hydrogen bond counts, five acceptor hydrogen bond counts, and one covalently bonded unit quantity. There is no formal charge, and the topological polar surface area is 98 Å2. Phloretin has a molecular weight of 274.27 g/mol, while its exact mass is 274.084 g/mol (“Phloretin | C15H14O5—PubChem”). Phloretin has two aromatic phenol rings (rings A and B), hydroxyl groups, and a carbonyl group (Tuli et al. 2022a). Figure 2 portrays the chemical structure of Phloretin.

Chemical structure of Phloretin
Structure–activity relationship (SAR) of Phloretin linked with anti-cancer activity
The biological activities of a natural product are attributed to having hydroxyl or phenolic structures as a functional group surrounding its core structures. For example, Phloretin is a potent flavanone with significant health advantages because of its polyphenolic group (Tuli et al. 2022a). The total number and arrangement of functional hydroxyl groups considerably affect Phloretin's antioxidant activity, which can scavenge free radicals or chelate metals. Phloretin has four hydroxyl groups at different position of ring A and B. Due to its pharmacophore and 2,6-dihydroxyacetone region, phloretin can eliminate peroxynitrite radicals with IC50 of 3.1 μM (Rezk et al. 2002). This scavenging capacity of Phloretin protects DNA from oxidative damage by raising levels of the antioxidant glutathione (GSH) in human colon cancer Caco-2 and HT-29 cells (Schaefer et al. 2006). The existence of a pair of bonds within phloretin's C2 and C3 atoms has a significant impact on cell proliferation (Jacob et al. 2011). In addition, phloretin's di-OH3',4', double bond at C2–C3, and carbonyl group at C-4 mediate a selective attack on cyclin-dependent kinase 2 (CDK-2), leading to arrest at cell cycle at G1 phase (Mintie et al. 2020). The structural organization of phloretin facilitates the formation of interaction with metal, including Ruthenium. In MCF-7 and MDA-MB-231 breast cancer cell lines, the combination of Ruthenium-phloretin modifies both natural apoptosis and the anti-angiogenic pathway by modulating p53, Bcl-2, and Bax and lowering the PI3K/Akt/mTOR and MMP9-regulated tumour invasive pathways (Roy et al. 2022). Phloretin has a topological polar surface area (TPSA) of approximately 97.99 Å2. There is a strong connection between this TPSA and anti-cancer activity. Prasanna S, Doerksen RJ concluded that natural compound anticancer properties are correlated with normal TPSA through inhibition of the telomerase and aromatase enzymes in cancer cells (Prasanna and Doerksen 2008). Overall, a brief diagram for SAR studies of Phloretin is summarized at the Fig. 3.

Diagram for Structure–Activity Relationship (SAR) studies of Phloretin
Anti-cancer perspectives of phloretin
In silico anti-cancer activity
In silico analysis can predict or analyze a molecule's biological activity without conducting experiments in a laboratory against diseases, including cancer. In silico analysis can provide valuable insights into its potential effects when assessing the anti-cancer activity of a compound like phloretin. It induced fit molecular docking prediction shows that phloretin showed notable binding affinity, especially with carbonic anhydrase I, ENOS, and INOS with variable binding energy. Additionally, Phloretin showcased significant potential against Caspase 3 and NF Kappa, thereby showing its potential as an effective against several anti-cancer therapeutics (Ranjanamala et al. 2021). Anand Mariadoss et al. showed in their study phloretin exhibited a significant binding interaction pattern with several receptors, including EGFR, Bcl-2, NF-КB, c-Kit receptor protein-tyrosine kinase, Farnesyl transferase, PDGFs and VEGFR2 proteins (Anand Mariadoss et al. 2018). However, other types of study, including data mining and system pharmacology, are preliminary steps in drug discovery and development against cancer treatment before starting clinical trials.
In vitro and in vivo anti-cancer perspectives
Existing evidence shows that Phloretin is effective against numerous human malignancies at both in vitro and in vivo (animal) (Figs. 4, 5, 6, 7) through several mechanisms. These specific mechanisms might lead to a potential remedy after the study of this leads in further phases of pharmaceutical development. In the sections, phloretin's antitumor properties and related processes in various malignancies are discussed briefly.

Regulation of apoptosis and cell proliferation by Phloretin. Phloretin must first cross the cellular membrane, which consists of a lipid bilayer embedded with various proteins. Carrier protein is an example of facilitated transporter, which might be used by Phloretin to entry into the cell before mediate function. Section and B denotes induction of apoptosis and cell cycle regulation, respectively

Control of signaling molecules by Phloretin in cancer treatment

Phloretin suppress metastasis and angiogenesis

Effect of Pholretin on glucose absorption by cancer cells via imbibition of GLUTs transporter
Breast cancer
Typically, breast cancer travels to other organs, including bone, liver, brain, and lung, making it a leading cause of death in women, and several factors may contribute to the development of breast cancer (Sun et al. 2017). According to previous research, natural flavonoids such as phloretin can be used to treat breast cancer. Phloretin (60–120 μM) can induce DNA damage and apoptosis while inhibiting cancer cell proliferation, viability, migration, invasion, and development of colonies in MCF-7 and MDA-MB-231 by activating p53, regulating the Bcl2/Bax ratio, and suppressing the PI3K/Akt/mTOR pathway coupled with MMP-9 downregulation (He et al.2020.). Wu et al. described that Phloretin (10–50 mg/kg) inhibits cell cycle progression and cell proliferation by inhibiting paxillin/FAK, Src, and alpha-smooth muscle actin (a-sMA), and the activation of E-cadherin, p53, p21 with decreasing vimentin and N-cadherin levels in MDA-MB-231 tumour xenografts mice(Wu et al. 2018). Phloretin, a GLUT inhibitor, reduced GLUT1 and GLUT12 expression in MCF-7 and MDA-MB-231 cells (Matsui et al. 2017). Phloretin inhibited the mTOR/ULK1, LCB-II pathways in MCF-7 and MDA-MB-231 cell lines, resulting in the control of cell proliferation and activation of autophagy in a separate study (Chen et al. 2021). Phloretin inhibits cell growth in immunocompromised NODSCID mice model while regulating reactive oxygen species (ROS) level control growth of in vitro MDA-MB-231 breast cancer cells (Brij B Patel et al. 2016) (Patel et al. 2017). JNK and p38 MAP kinase activation have been associated with phloretin-induced caspase-3 cleavage in H-ras-transformed human mammary epithelial cells (H-ras-MCF-10A). Additionally, these researchers found that phloretin increased p53 and Bax while lowering Bcl-xl, leading to apoptosis, and decreased H-ras proliferation at MCF-10A cells in a concentration-dependent way (Kim et al. 2009).
Prostate cancer
Prostate cancer develops when abnormal cells form and grow in the prostate gland. According to a previous study, Phloretin activates the apoptosis pathway by suppressing Bcl-xL, XIAP, and surviving, while increasing Bax's expression and reducing cell proliferation by decreasing VEGF, CCND1, and CCNB expression in prostate cancer LNCaP, CWR22Rv1, PC-3 cell line (Yang et al. 2022). Kang et al. reported that Phloretin (0–100 μM) inhibited cyclin B1, XIAP, and Bcl-2 in LNCaP cell line culture to arrest the G2/M phase cell cycle and trigger apoptosis (Kang et al. 2020). Furthermore, phloretin (50–100 μM) was associated with increased MTT cytotoxicity and cell death induction at LNCaP prostate cancer cells (Szliszka et al. 2010). In a different study, phloretin was used to examine how some GLUTs (GLUT1 and GLUT4) are regulated inside of androgen-sensitive (LNCaP) as well as insensitive (PC-3) prostate cancer cell lines, where phloretin (25–39.4 µM) showed anti-proliferative properties by modifying the expression of the glucose transporter (GLUT) or by changing glucose binding while also regulating cell growth (Gonzalez-Menendez et al. 2014).
Colon cancer
Colon cancer is a prominent cancer type around the world (Jin et al. 2020). Phloretin has anti-inflammatory, antioxidant, and anticancer properties (Kim et al. 2022). Jin et al. showed that Phloretin induces apoptosis and DNA fragmentation and decreases cell proliferation via upregulating CAT, SOD, glutathione, and Caspase-3, while Bcl-2, NF-B, MMP-9 and Akt/mTOR signal were downregulated in HT-29 cells(Jin et al. 2020). Phloretin treatment boosted apoptosis in HT-29 Luc cells by raising cleaved poly(ADP ribose) polymerase, Mac-1, caspase-3, and caspase-8 in a dose-dependent manner (J. L. Kim et al. 2022). Phloretin (2.35–18.75 g/ml) boosts the killing effect of γδ T lymphocytes and the production of PFP and GraB in a human SW-1116 cancer cell line by inhibiting the Wnt signalling pathway(Zhu et al. 2013a, b). Park et al. reported that Phloretin (0-100 mol/L) led to apoptosis in HT-29 cells by increasing the levels of Bax, cytochrome c, caspase-8, -9, -7, and -3, Smac/Diablo, and poly(ADP-ribose) polymerase(Park et al. 2007). Phloretin (100 µM) induced apoptosis and arrested the G2/M checkpoint in human carcinoma SW620 and HCT116 cells via elevating phosphor-cdc2 and Myt1 and decreasing cyclin B and cdc2 (Zhou et al. 2018). Phloretin inhibits cell growth and cell invasiveness in both in vivo and in vitro models by causing a cell cycle arrest at the G0/G1phase, which increases the actions of the p21/Cip1 and p27/Kip1 proteins (p53-mediated proteins) and decreases GLUT2 expression(Lin et al. 2016). Phloretin (25–300 µM) causes programmed cell death (apoptosis) as well as reduced cell proliferation, colony formation, and cell migration at HCT-116 and SW-480 colon cancer cells by inhibiting Wnt/β-catenin signaling, which results in the downregulation of cyclinD1, c-Myc, and survivin and the upregulation of ROS and bax expression(Kapoor and Padwad 2023).
Liver cancer
One of the most prevalent and deadly types of cancer is hepatocellular carcinoma (HCC), also known as liver cancer (Wu et al. 2009a, b). Despite significant advancements regarding treatment strategies in recent years, there are no possibilities for curative therapy, and patients with HCC still have few therapeutic options and poorer prognosis (Pope et al. 2019). However, Phloretin has therapeutic effects against HCC. Phloretin (200 µM), inhibits glucose transporter (GLUT) and triggers apoptosis in HepG2 cells by suppressing the Akt and Bcl-2 signalling pathways (Wu et al. 2009a, b). Phloretin might increase the expression of SHP-1 and decrease STAT3 as well as inhibit pAKT/pERK signalling in HCC cells leading to reducing angiogenesis, cell proliferation, and inducing apoptosis (Saraswati et al. 2019). Phloretin (1–200 μM) inhibits the iodothyronine-5'-deiodinase and the mammalian glucose transporter in Hep G2 cell line (Movius et al. 1989). Phloretin (50–150 mM), either alone or in combination, has the power to fight liver cancer by inhibiting the glucose transporter GLUT, Bcl-2, NF-κB, T4 5'-deiodinase, STAT3 expression, SHP-1 expression, pAKT/pERK, Akt/VEGFR2 pathway and activating, caspase-3 and -9 in Hep G2 cells (Yang et al. 2009).
Pancreatic cancer
Pancreatic cancer typically develops in advanced stages and has a limited chance of survival (Rawla, Sunkara, and Gaduputi 2019). Phloretin has recently been shown to be effective in treating pancreatic cancer. Phloretin act as a ROS inhibitor to lessen MAPK signalling and SOD activity and reverse treatment resistance in Panc1 and HTB147 pancreatic cancer cells. In addition, in the tumour xenograft model, by inhibiting CXCL12/CXCR4-dependent MAPK signalling and ROS pathways, phloretin and gemcitabine combined inhibit cell proliferation and migration, reversing the drug susceptibility of Panc-1 lines (Patel et al. 2016).
Kidney cancer
Kidney cancer is also known as renal cancer, in which kidney cells develop into malignant tumours. Appropriate treatment is needed to prevent kidney cancer; phloretin has recently proven helpful. According to Funari et al. Phloretin (20–40µΜ) can prevent ROS formation and inhibit cell growth in HEK-293 cells (Funari et al. 2011). In human embryonic kidney (HEK) 293-hTLR2 cells, phloretin inhibits TLR1 and TLR2 receptors substantially, decreasing TNF-α and interleukin-8 inflammatory cytokines (Kim et al. 2018). Phloretin (50 μM) markedly decreased the renal ECM deposition level and the α -SMA in renal tubules, which cells indirectly influenced cancer and reduced the release of cytokines, including NOD-like receptor family domain 3 (NLRP3) and interleukin-1β (IL-1β), mitochondrial reactive oxygen species (ROS), and morphological lesions in the kidneys of HUA mice (Cui et al. 2020).
Oral cancer
Oral cancer, which is caused due to several factors, makes up 2–4% of all malignancies in the United States(Suhail et al. 2016). Phloretin inhibits the proliferation of human oral cancer cells, making it a potential therapy for oral carcinoma. Phloretin (12.5 µM) promotes necrosis and reduces cell proliferation with migration by inducing cell cycle arrest at the G0/G1 phase in the SCC-1 cell line in which CDK4, CDK6, and cyclin D1 expression are downregulated (Yang et al. 2020). Phloretin (15.3 g/mL) inhibits glucose transporters (GLUTs), induces apoptosis, and reduces cell proliferation in YD-9 cells by elevating the levels of p53 and Bax (Nam et al. 2017). Above research's results indicate that phloretin, a secondary plant metabolite with tremendous pharmacological potential, might have the capacity to be a therapeutic agent for oral cancer by inducing apoptosis, cell cycle arrest, reactive oxygen species (ROS), and inhibiting cell proliferation, migration, and glucose uptake via associated signaling pathways.
Glioblastoma
Glioblastoma (GBM) is one of the deadliest and most resistant solid malignant tumors(Davis 2016)(Nahar Metu et al.,2023). Phloretin has a chemopreventive effect on glioblastoma cells in humans. Phloretin (0–300 µM) suppresses cell proliferation and induces apoptosis in U87 and U251 glioblastoma cells by decreasing Bcl-2, cyclin D, cyclin E and CDK2, CDK4, CDK6, and increasing Bax, Bak, c-PARP (Liu et al. 2016). Furthermore, Phloretin suppressed specific signalling molecules, including PI3K/AKT/mTOR cascades, resulting in G0-G1 cell cycle arrest (Liu et al. 2016). In SH-SY5Y cells, Phloretin (200 µΜ) lowered cell viability through the JNK-Beclin-1 pathway and triggered autophagy by downregulating p-AKT and p-mTOR (Fan et al. 2022).
Ovarian Cancer
Ovarian cancer is the growth of cells that form in the ovaries. Phloretin (0.6–300 μM) induced apoptosis through activating caspase-3 and suppressed metastasis by reducing the MMP ovarian cancer cell line. Moreover, it stops some cancerous properties, including lowering glycolysis by decreasing the function of HIF1-inducible glycolytic enzymes (GLUT1, HKII, PFKFB3, PDHK1, LDH) (Xintaropoulou et al. 2015).
Bladder cancer
A prevalent form of cancer that starts in the bladder's cells is bladder cancer. Phloretin might be effective in bladder cancer treatment. Hui et al. reported that Phloretin (20 uM) helped to accumulate H2O2, leading to cytotoxicity and preventing migration of AQP9 cell lines (Hui 2020).
Myeloma cancer
A plasma cell cancer called myeloma occurs when the blood cells proliferate uncontrollably. Phloretin (100 μM) inhibits GLUT transporters, including GLUT8 and GLUT11, which reduces cellular proliferation and glycolysis in MM.1S, RPMI 8226 and U266 cell (McBrayer et al. 2012). Friday at el. showed that Phloretin (200 μM) enhanced ROS, elevated TRX activity, and lowered TXNIP activity to enhance apoptosis and inhibit cell growth (Friday et al. 2011).
Melanoma cancer
Melanocytes are the cells that produce melanoma, the most dangerous form of skin cancer. In skin cancer melanoma 4A5 cells, phloretin (25 μM) inhibits elastase, MMP-1, decreases cellular tyrosinase activity and melanin concentration, leads to promotes apoptosis and inhibits metastasis via an unknown mechanism (Casarini et al. 2020). In addition, KoBoRI et al. showed how Phloretin led to apoptosis in HL60 and B16 cells by increasing the expression of BAX protein without affecting p53, Bcl-XL, and Bcl-2, and by inhibiting the activity of protein kinase C (KoBoRI et al. 1999). In addition, in SK MEL28 cell lines, Phloretin induces apoptosis and reduces metastasis, suppressing MMPs in a dose-dependent manner (Casarini et al. 2021).
Thyroid cancer
Malignant cells can develop in the tissues of the thyroid gland, resulting in thyroid cancer. Phloretin exerted its anti-cancer activities in thyroid cancer by controlling the pathway of PI3K-Akt and regulating apoptosis-related proteins, including the downregulation of Bcl-2 and Bcl-xL, while the increased expression of Bax, caspase-3, -8, and -9 in thyroid cancer SW579 cell lines (Ma et al. 2022).
Osteosarcoma
The most prevalent form of cancer that originates in the bones is known as osteosarcoma. Phloretin inhibited the uptake of 2-NBDLG, an inhibitor of the sodium-glucose co-transporter (SGLT) and prevented the absorption of 2-NBDLG in osteosarcoma U2OS cell lines in a dose-dependent manner (Ogawa et al. 2021).
Esophageal carcinoma
Cancer of the oesophagus, also known as oesophagal cancer, is a disease that develops in the esophagus, a tube extending from the throat to the stomach. Phloretin (60–100 µg/ml, 6–48 h) prevents cell proliferation and induces apoptosis, a mitochondria-dependent route with proper regulation of bcl-2 and p53 in EC-109 cell (Duan et al. 2017).
Cervical cancer
Cancer that originates in the cells of the cervix is referred to as cervical cancer. In human SiHa cervical cancer cells, phloretin (60 μM) demonstrated substantial inhibition of invasion and migration by downregulation of matrix metalloproteinase (MMP)-2, MMP-3, cathepsin S and mesenchymal markers such fibronectin, vimentin, and RhoA as well as the epithelial-mesenchymal transition brought on by transforming growth factor-1(Hsiao et al. 2019). Another study showed that phloretin increases BAX, activates caspases to induce apoptosis and decreases Bcl2 and MMPs by stopping cell growth (Tuli et al. 2022a). Phloretin also causes cytotoxic activity to be induced in human HeLa cell lines and suppresses the viability of the cells (Abu-Azzam et al. 2022).
Lung carcinoma
The lungs are the primary site of origin for lung cancer, although the disease can also extend to lymph nodes and other organs. Phloretin had an anti-cancer activity in lung cancer A549 cell lines at a 20 mg/kg dose by inducing apoptosis via the P38 MAPK and JNK1/2 pathways in vivo, while increasing the expression levels of BAX, cleaved caspase-3 and -9, and the degraded form of PARP at a 200 µM dosage in A549 cell line (Min et al. 2015). NSCLC starts at the cellular level and causes aberrant lung cells to proliferate quickly and uncontrollably. Phloretin dramatically increased P38 MAPK and JNK1/2 phosphorylation, which inhibited cell proliferation and increased BAX expression, caspase-3, caspase-9, and the degraded form of PARP, while decreasing Bcl-2 in an in vitro model of NSCLC cell line A549. Furthermore, in A549 xenografts, Phloretin at 20 mg/kg exhibited an increasingly substantial tumour growth-inhibiting effect (Min et al. 2015).
Leukemia
Cancer that affects the blood-forming tissues of the body, including the bone marrow and the lymphatic system, is referred to as leukemia. Phloretin (0.1–0.2 mM) increases the Caspase3 activity in human leukemia HL60 cell lines by stopping PKC activity, leading to inducing apoptosis (KoBoRI et al. 1999). A summary of the chemotherapeutic action of Phloretin against human malignancies is tabulated in Table 2.
Combination of phloretin with other agents in cancer treatment
Although future studies are necessary for strong evidence, according to the previous literature review, phloretin might have therapeutic efficacies against a wide range of cancers. However, a combination of Phloretin with other phytochemical or conventional drugs can show more effectiveness because of their synergistic activities at single or several target sites connected to the regulation of cancer. For example, Phloretin has a synergistic anti-tumour effect when taken with radiation, possibly via inducing apoptosis, reducing cell proliferation and cell growth rate at the same time glucose transport in lung cancer in Lewis lung cancer (LLC) xenograft model (Tang and Gong 2021). Phloretin with atorvastatin combination synergistically effective in colon cancer, induced apoptosis and G2/M phase cell cycle arrest through reducing cyclin B expression and upregulating of phospho-cdc2 and Myt1 at SW620 and HCT116 cancer cells (Zhou et al. 2018). Recent research indicates that the combination of Phloretin and cisplatin enhances the therapeutic efficacy of cisplatin by promoting the inhibition of MMP-2, Bcl-2, and MMP-9 with increased levels of cleaved-caspases in A549 cells (Ma et al. 2016). Abkin and colleagues investigated the anti-tumour activity of recombinant HSP70 Chaperone in combination with Phloretin. They discovered that the penetration of HSP70 into K562 cells increased its anticancer effect at in vitro and in vivo melanoma model (Abkin et al. 2016). Phloretin, in combination with daunorubicin, overcomes hypoxia-induced drug resistance by sensitizing daunorubicin-resistant SW620 and K562 cancer cells to its anti-tumor effects and apoptosis (Cao et al. 2007). Berbamine with phloretin efficiently demonstrated anti-tumor effects in two SORA-resistant hepatocellular carcinoma cell lines by augmenting SHP-1, basal, and IL-6-induced the inactivation of STAT3 (W. Zhao et al. 2020). Phloretin substantially induced cytoprotective autophagy in breast cancer cell lines by modulating the mTOR/ULK1 pathway with chemotherapeutic agents such as tamoxifen and doxorubicin (Chen et al. 2021). Phloretin has been observed to be a good combination with other natural products. For example, Payne et al. showed that Phloretin and phloridzin act as anti-neoplastic agents in the HeLa cell line, indicating a better treatment option for cervical cancer (Badwaik, Tockstein, and Thompson 2018). Ruthenium is an inorganic compound. Ruthenium-phloretin chemotherapy is an effective treatment because it modifies intrinsic apoptosis and the anti-angiogenic pathway by modulating p53, Bcl2, and Bax and reducing the PI3K/Akt/mTOR and MMP9-regulated tumour invasive pathways in MCF-7 and MDA-MB-231 breast cancer cell lines (Roy et al. 2022). Phloretin and gemcitabine inhibit cell proliferation, migration, and reverse drug susceptibility in Panc-1 cells in a tumour xenograft model by involving CXCL12/CXCR4-dependent MAPK signaling and ROS pathways (Brij B Patel et al. 2016). An overview of the synergistic activity of Phloretin with other phytochemicals and conventional chemotherapeutic agents is tabulated in Table 3.
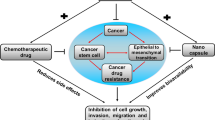
Power of phloretin in alleviating multi-drug resistance of existing chemotherapeutics drugs in numerous cancer types
Due to numerous serious issues, such as multi-drug resistance, treating cancer patients is becoming difficult (So et al. 2009). This resistance mechanism may result from a variety of potential protective mechanisms, including drug efflux (Po and Pc 2012), drug inactivation (Akhdar et al. 2012), drug detoxification (Akhdar et al. 2012), drug target modification (Lynch et al. 2004), involvement of cancer stem cells (Costea et al. 2020), miRNA dysregulation (Si et al. 2019), epigenetic alteration (Mansoori et al. 2017), and other numerous irregular DNA damage/repair mechanisms, tumour microenvironment, and ROS modulation (Si et al. 2019; Costea et al. 2020; Mansoori et al. 2017). Drug resistance is linked to several proteins, including P glycoprotein (P-GP), MRP 1, MRP 1–9, BCRP, and changes in beta-tubulin (Xue and Liang 2012). However, previous research on natural phytochemical-based selective inhibitors of multi-drug resistance protein might improve the effectiveness of cancer chemotherapy(Molnár et al. 2010). However, most had numerous adverse effects and were ineffective at their intended location. The effectiveness of cancer treatment may potentially be improved by blocking MDR-efflux proteins. In human Panc-1 pancreatic cancer cells, co-treatment with phloretin and daunomycin significantly increased the accumulation of daunomycin and vinblastine by inhibiting drug transport via MRP1 (Nguyen et al. 2003). Zhao et al. describe that phloretin is an effective substrate of P-gp, MRP2, and BCRP and works with active transport, efflux protein transport and cell bypass (Zhao et al. 2020). Molnár et al. described that phloretin is an effective modulator to overcome the resistance of anticancer drugs, where phloretin identified as a potent P-gp inhibitor in mouse lymphoma cells (L1210) and human breast cancer cells MDA-MB-231 cell line (Molnár et al. 2010). Furthermore, phloretin inhibited P-gp-mediated cellular efflux in MCF-7 and MDA435/LCC6 breast cancer cell lines (Zhang and Morris 2003). Phloretin suppresses STAT3 expression in sorafenib-sensitive and resistant HCCs via SHP-1-mediated inhibition of STAT3 and AKT/mTOR/JAK2/VEGFR2 pathway, suggesting that Phloretin could potentially become an anticancer agent for hepatocellular carcinoma by overcoming sorafenib resistance (Saraswati et al. 2019). Phloretin inhibited active efflux and reduced steady-state accumulation of VCR in PC-V40 subline in vincristine (VCR)-selected murine erythroleukemia (MEL) PC4 cell lines. In doxorubicin-resistant SNU-1 gastric cancer cells, Phloretin inhibited proliferation by inducing cell cycle arrest at G0/G1 phase by depleting cyclin D1 and D2 expression, activating autophagy by increasing LC3B II and Beclin-1 expression, and blocking the ERK1/2/MAPK signaling pathway (You et al. 2020). Figure 8 describes the synergetic and overcoming resistance mechanism of Phloretin.

Graphical presentation of synergistic and alleviating multidrug resistance by Phloretin
Nano formulation strategies of phloretin with the aim of better bioavailability
According to the literature review, Phloretin has many therapeutic functions, including antioxidant, anti-inflammatory, anti-diabetic and anti-tumour (Crespy et al. 2001; Tuli et al. 2022a). However, its low bioavailability limits its efficacy as a cancer treatment. Nanoformulation strategies can help improve the bioavailability of phloretin and enhance its therapeutic potential in cancer treatment. Phloretin's bioavailability is increased when combined with nanomaterials, a novel method of treating cancer cells. In cancer treatment, the choice of nanoformulation method depends on various factors such as the desired route of administration, therapeutic goals, and the physicochemical properties of Phloretin. Some possible strategies include nanoparticles, nano-emulsions, solid lipid nanoparticles (SLNS), nano-suspensions, self-emulsifying drug delivery systems (SEDDS), co-crystals, nanoparticles with targeting ligands, inclusion complexes, surface modifications and quality control and characterization can ensure the safety and efficacy of the nano-formulation before it can be used for clinical applications. In vitro, phloretin in NLC had a sustained release pattern more suited for absorption than phloretin ethanol solution. Phloretin-loaded chitosan nanoparticles (PhCsNPs) were used to demonstrate PhCsNPs' anticancer effectiveness through the measurement of cytotoxicity, intracellular ROS, mitochondrial dysfunction, lipid peroxidation, antioxidant status, apoptotic-associated gene expression profile, and cell cycle analysis in human oral cancer cell lines (YD-9, CA9-22) by releasing Bax, cytochrome-c, and caspases-3 -9 (Arokia Vijaya Anand Mariadoss et al. 2019a, b; Arokia Vijaya Anand et al. 2019). Phloretin was delivered locally to oral malignancies (YD-9 cell lines) using a fast-dissolving nanofiber (NF) made of poly(vinyl alcohol) (PVA) and D-a-tocopheryl polyethene glycol succinate (TPGS), which also boosted the phloretin's anti-proliferation effectiveness (Nam et al. 2017). In MDA-MB-231 cells (human breast adenocarcinoma cells that express the GLUT1 and CD44 receptors), HACE-d/phloretin NPs showed improved cellular accumulation efficiency, anti-proliferation, and spheroid growth-inhibitory impact at 279 µM concentration. The optical imaging test revealed that the MDA-MB-231 tumour-xenografted mice HACEd/phloretin NPs group had improved anticancer efficacies compared to the other experimental groups (Lee et al. 2017). Flow cytometry was monitored to evaluate the anti-neoplastic effectiveness of the phloretin-attached AuNPs against cervical malignant cell lines (HeLa) (Badwaik, Tockstein, and Thompson 2018). Phloretin-loaded NC-Phl, a novel semi-solid formulation (HG-NCPhl), increased skin penetration and adherence while having a cytotoxic impact on Sk-Mel-28 cells (Casarini et al. 2021). In research, chitosan-coated nanoparticles with phloretin (CS-PLGA/Phl) were examined in vivo using monolayer-growing mouse melanoma cells (B16F10) (Lee et al. 2019). A recent study shows that polymerization-induced self-assembly creates microscopes of cross-linked poly (cyclotriphosphazene-co-phloretin) (PCTPPT). The regulated release of the anti-cancer model medication (CPT) is effectively explored in various pH media (Mehmood et al. 2022). These results imply that phloretin can be introduced into nanoparticles to increase their stability and enhance biological capabilities.
Toxicity of phloretin
Phloretin provides hundreds of therapeutic benefits, however, it may also address several adverse implications or toxic reactions. There may be risks when phloretin is administered to mammalian cells at high dosages, according to a recent study on the substance's related toxicity (de Oliveira 2016). In a corroboration analysis of toxicity, they observed that Phloretin was given at APAP models alone at the maximum dose rate (2.4 mmol/kg), ultimately caused total lethality within 24 h & lower doses were not harmful, while the most significant oral dose alone generated 64% lethality(Geohagen et al. 2018). Despite having cytoprotective properties, Phloretin tended to be toxic in our APAP models (Geohagen et al. 2018). Exposure to 0.1 Mm of phloretin for one hour in primary cultured rat hepatocytes significantly inhibited GSH content and MMP activity, leading to severe cytotoxicity (Frerichs and Ball 1964). Another experiment showed that phloretin could simulate PINK1 knockdown in primary astrocytes at high concentrations, reducing cellular function, viability, and proliferation(Choi et al. 2013). Moreover, Phloretin (30 μM for 60 min before exposure to Amyloid-β) also decreased the toxicity that Aβ peptides induced in cultured PC12 cells. But it's interesting to note that PC12 cells were cytotoxic to phloretin at 300 μM (de Oliveira 2016). One more controversy surrounding Phloretin is its safe consumption levels. The safety of Phloretin consumption remains a topic of discussion. Most people obtain Phloretin from dietary sources like soy and red clover. Consuming moderate amounts of Phloretin from these sources is considered safe for the general population. However, high-dose supplements or concentrated forms of Phloretin may pose risks, particularly if taken by individuals with a history of hormone-related cancers or hormonal imbalances. Additionally, the effects of Phloretin may vary from person to person. Genetic factors, diet, and hormonal status can influence how Phloretin interacts with the body. More investigations on the parent phloretin's safety are needed before human clinical trials can evaluate the bioactive compound's anticancer potential (Fig. 9)

Overview of anti-cancer profile of Phloretin against human malignancies
Pharmacokinetics of Phloretin and future perspective in drug development
Lipinski's "Rule of Five" was employed for the "drug-likeness" test, and compounds met the requirements. More precisely, it was revealed that Phloretin's molecular weight, QPlogPo/w, HBD, and HBA were 274.080 g/mol, 2.1, 4, and 5, respectively, indicating that the compounds had good drug-like properties. The ADMETLAB 2.0 server was used to determine the physicochemical properties of phloretin, including molecular weight (MW), volume, density, nHA, nHD, nRot, nRing, MaxRing, nHet, fChar, nRig, flexibility, stereo centres, TPSA, logS, logP, and logD. These results are illustrated in Fig. 10.

In silico pharmacokinetics evaluation of Phloretin. Note: MW(Molecular weight), Volume(Vander Waals Volume), Density, nHA (The number of hydrogen bond acceptors), nHD (The number of hydrogen bond donors), nRot (Number of ratable bond), nRing (Number of ring), MaxRing (Number of atom in the biggest ring), nHet (Number of hetero atoms), fChar (Formal charge), nRig (number of rigid bond), flexibility, stereo centres, TPSA(Topological polar surface area), logS (Water solubility), logP (Partition coefficient), logD (Distribution co-efficient)
In assessing whether or not to advance a molecule to the clinical stage, the assessment of drug absorption, distribution, metabolism, excretion, and toxicity (ADMET) prediction may assist in minimizing the risk, time, and expenditure entailed (Wu et al. 2020; Lin et al. 2005; Sohel, Sultana, Sultana, Al Amin, et al. 2022). Table 4 depicts the pharmacokinetics profile of phloretin using in-silico techniques like Schrodinger's QuickPro modules and the publicly available server admetSAR, which gives credible information on absorption, distribution, metabolism, excretion, and toxicity.
Drugs must be absorbed to reach the bloodstream and be used daily. In accordance with what we had anticipated, phloretin could be readily absorbed when taken orally and may pass via skin, gut, and kidney cells. Drug movement within the body is referred to as a distribution, and it depends on several variables, along with the permeability of the blood–brain barrier, the permeability of the central nervous system, the drug's capacity to attach to plasma proteins, and the cumulative volume of distribution. Drug concentrations in the bloodstream are reduced dramatically when they bind to plasma proteins such as human serum albumin, lipoprotein, glycoprotein, and globulins. The fact that our intended compounds fall within the recommended range indicating they have a good chance of reaching the target spot and moving freely through the bloodstream. The quantity of bodily fluid necessary for blood plasma to dissolve is known as the volume of distribution. Thankfully, phloretin is highly likely to spread evenly throughout blood and tissue. The central nervous system and blood–brain barrier (QPlogBB) are also critical for medications that address brain disease. However, Phloretin's ability to penetrate the central nervous system and a brain-blood barrier is low.
More than 80% of medications go through the CYP450 enzyme's detoxification process in the liver during first-pass metabolism, and blocking this enzyme increases drug toxicity and causes several side effects. According to the expected metabolic outcome, phloretin is a substrate for many CYP450 subunits, including CYP2C9 and CYP2D6. Phloretin's in silico toxicity profile was predicted using data on hepatotoxicity, cardiotoxicity, skin sensitization, kidney toxicity, eye irritation, and AMES toxicity. Phloretin has demonstrated toxicity against these effects and toxicity against the eye, skin and AMES. Using the total clearance (CLtot) and T 12, we estimated the clearance profile based on the renal clearance property of the compounds. As anticipated, the findings showed that phloretin can be easily eliminated from the human body after performing its therapeutic function. In silico drug-likeness and pharmacokinetics properties profile of Phloretin is summarized at Table 3.
Given that there are fewer toxicity hazards, the pharmacokinetics profile of phloretin predicts good lead and efficiency. So, in the context of cancer treatment and drug discovery, phloretin can serve as an excellent starting point for drug design techniques like docking and pharmacophore-based virtual screening.
Concluding remark and future recommendation
Treating human disorders, including cancer patients, has significantly benefited from using natural products. Due to its inexpensive cost, wide availability, and extended usage history, phloretin may be a promising drug for treating cancer like other plant-based anti-cancer drugs. We believe that our review is the first of its kind to systematically review the remarkable anti-cancer potential of phloretin. across a diverse spectrum of cancer types, diverse mechanisms, combinational effect with conventional drugs, reversing antineoplastic activity of resistance drugs, safety profile.Our systematic review showed that phloretin has an anticancer effect against numerous malignancies, including lung, oral, oesophagal, gastric, colon, liver, breast, pancreatic, kidney, glioblastoma, ovarian, cervical, bladder, myeloma, skin, melanoma, leukaemia, thyroid, osteosarcoma, oesophagal and prostate cancer. It can be seen that phloretin employs multiple mechanisms to halt cancer progressions, such as modulation of the signalling cascade associated with apoptosis, autophagy, necrosis, metastasis, angiogenesis, cell proliferation, glucose absorption, oxidative stress, inflammation and DNA damage (Fig. 9). Phloretin's ability to prevent cancer has been connected in particular to the control of several proteins Bcl-2, Bax, Bak, Bad, caspase, cyclins (B1, D1, E) and CDKs (4, 6,7) p18, p21, p27, p53, MMP-2, MMP- 8, MMP-9, Wnt/-catenin, PARP, TNF-α, NF- κB, IκB kinase, IL-1β, TNF-α, phospho-Akt, phosphor-p65, NF-κB, PI3K/Akt, MAPK/ERK, p-mTOR. Phloretin may have more potent anticancer effects when coupled with other phytochemicals or conventional drugs than when taken alone. Such synergistic mechanisms aid in alleviating resistance to conventional drugs used in cancer treatment. Phloretin has a good combination with some nano-particles, which adds an extra advantage to improve the bioavailability of phloretin in cancer treatments. Moreover, phloretin has a good structure–activity relationship, positively impacting cancer biology therapeutics. Phloretin possesses favourable in silico pharmacokinetic properties such as absorption, distribution, metabolism, excretion, and less toxicity. However, like many potential cancer treatment strategies, Phloretin has limitations, including limited clinical evidence, low bioavailability, specificity of action, side effects, interaction with other drugs, and optimum doses. Therefore, more research on well-designed clinical trials to evaluate the long-term actual safety and efficacy, improving bioavailability, combination with other cancer therapies, such as chemotherapy, radiation, or immunotherapy, and strategies to prevent or overcome resistance should be explored before recognizing phloretin as a standard cancer biology drug.
Abbreviations
- Bcl-2:
-
B-cell leukemia 2
- Bax:
-
Bcl-2-Associated X protein
- PI3K:
-
Phosphoinositide 3-kinase
- Akt:
-
Ak strain transforming
- mTOR:
-
Mammalian target of rapamycin
- MMP9 :
-
Matrix metalloproteinase 9
- GLUT:
-
Glucose transporters
- ULK1 :
-
Unc-51 like autophagy activating kinase
- ERK1/2:
-
Extracellular signal-regulated kinase 1/2
- TPA :
-
Tissue plasminogen activator
- LC3B :
-
Light chain 3B
- BCL-XL :
-
B-cell lymphoma-extra large
- XIAP :
-
X-linked inhibitor of apoptosis protein
- VEGF :
-
Vascular endothelial growth factor
- MTT :
-
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide
- CAT :
-
Chloramphenicol acetyltransferase
- SOD :
-
Superoxide dismutase
- NF-B :
-
Nuclear factor-B
- Mac-1 :
-
Macrophage 1 antigen
- PEP :
-
Phosphoenolpyruvate
- GraB :
-
Granzyme B
- c-Myc :
-
Cellular Myc
- ROS :
-
Reactive oxygen species
- HCC :
-
Hepatocellular carcinoma
- SHP-1 :
-
Src homology region 2 domain-containing phosphatase-1
- STAT3 :
-
Signal transducer and activator of transcription 3
- pERK :
-
Protein kinase RNA-like endoplasmic reticulum kinase
- VEGFR :
-
Vascular endothelial growth factor receptor
- MAPK :
-
Mitogen-activated protein kinase
- CXCL12 :
-
C-X-C motif chemokine ligand 12
- CXCR4 :
-
C-X-C motif chemokine receptor 4
- TLR1 :
-
Toll-like receptor 1
- TLR2:
-
Toll-like receptor 2
- TNF-α :
-
Tumor necrosis factor-alpha
- NOD :
-
Nucleotide-binding oligomerization domain
- NLRP3:
-
Nucleotide-binding domain (NOD)-like receptor protein 3
- CDK4 :
-
Cyclin-dependent kinase 4
- CDK6 :
-
Cyclin-dependent kinase 6
- BAK :
-
Bcl-2 homologous antagonist/killer
- PARP :
-
Poly-ADP ribose polymerase
- HIF1 :
-
Hypoxia-inducible factor-1
- HKII :
-
Hexokinase 2
- PFKFB3 :
-
6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase isozyme 3
- PDHK1 :
-
Pyruvate dehydrogenase kinase 1
- LDH :
-
Lactate dehydrogenase
- Pink1 :
-
PTEN-induced kinase 1
- GSH:
-
Glutathione
- NSCLC :
-
Non-small cell lung cancer
- JNK :
-
C-Jun N-terminal kinase
- TRX:
-
Thioredoxin
- TXNIP :
-
Thioredoxin interacting protein
- 2-NBDLG :
-
2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl) amino]-2-deoxy-D-glucose
- PKC:
-
Protein kinase C
- Myt1 :
-
Myelin transcription factor 1
- cdc2:
-
Cyclin-dependent kinase 2
- HSP70:
-
70 Kilodalton heat shock protein
- IL-6:
-
Interleukin-6
- MRP:
-
Multi-drug resistance associated protein 1
- BCRP:
-
Breast cancer resistance protein
- MDR:
-
Multi-drug resistance
References
Abkin SV, Ostroumova OS, Komarova EY, Meshalkina DA, Shevtsov MA, Margulis BA, Guzhova IV (2016) Phloretin increases the anti-tumor efficacy of intratumorally delivered heat-shock protein 70 KDa (HSP70) in a murine model of melanoma. Cancer Immunol Immunother. https://doi.org/10.1007/s00262-015-1778-1
Abu-Azzam O, Aldalaen S, Nasr M (2022) Potentiation of the cytotoxic activity of nutraceutical phloretin against cervical cancer by formulation into microemulsion. Pharm Chem J 55(11):1215–1218
Akhdar H, Legendre C, Aninat C, More F (2012) Anticancer drug metabolism: chemotherapy resistance and new therapeutic approaches. Top Drug Metabol 22:138–170
Alsanani A, Pate BB, Wondisford S, Johnson N, Banerjee D (2015) Stromal carcinoma associated fibroblasts promote drug resistance of human pancreatic cancer cells by modulation of ROS via CXCR4/CXCL12 signaling. Int J Transl Sci. https://doi.org/10.13052/ijts2246-8765.20151006
Anand M, Suresh K (2014) Biochemical Profiling and chemopreventive activity of phloretin on 7,12-dimethylbenz (A) anthracene induced oral carcinogenesis in male golden syrian hamsters. Toxicol Int. https://doi.org/10.4103/0971-6580.139805
Mariadoss A, Vijaya A, Dhanabalan AK, Munusamy H, Gunasekaran K, David E (2018) In silico studies towards enhancing the anticancer activity of phytochemical phloretin against cancer drug targets. Current Drug Therapy. https://doi.org/10.2174/1574885513666180402134054
Anand AV, Mariadoss VR, Baojun Xu, Karthikumar V, Vijayalakshmi S, Shalini V, Rani BS, Mohamed Sadiq A, Ernest D (2019) Phloretin loaded chitosan nanoparticles enhance the antioxidants and apoptotic mechanisms in DMBA induced experimental carcinogenesis. Chem Biol Interact 308:11–19. https://doi.org/10.1016/j.cbi.2019.05.008
Badwaik, Vivek D, Sarah Tockstein, David H Thompson (2018) Development of Dihydrochalcone-Functionalized Gold Nanoparticles for Augmented Antineoplastic Activity, 1917–26
Barreca D, Bellocco E, Laganà G, Ginestra G, Bisignano C (2014) Biochemical and antimicrobial activity of phloretin and its glycosilated derivatives present in apple and kumquat. Food Chem. https://doi.org/10.1016/j.foodchem.2014.03.118
Barreca D, Currò M, Bellocco E, Ficarra S, Laganà G, Tellone E, Giunta ML et al (2017) Neuroprotective effects of phloretin and its glycosylated derivative on rotenone-induced toxicity in human SH-SY5Y neuronal-like cells. BioFactors. https://doi.org/10.1002/biof.1358
Biswas P, Dey D, Biswas PK, Rahaman TI, Saha S, Parvez A, Khan DA, Lily NJ, Saha K, Sohel Md (2022) A comprehensive analysis and anti-cancer activities of quercetin in ROS-mediated cancer and cancer stem cells. Int J Mol Sci 23(19):11746
Cao X, Fang L, Gibbs S, Huang Y, Dai Z, Wen P, Zheng X, Sadee W, Sun D (2007) Glucose uptake inhibitor sensitizes cancer cells to daunorubicin and overcomes drug resistance in hypoxia. Cancer Chemother Pharmacol. https://doi.org/10.1007/s00280-006-0291-9
Casarini TP, Anunciato LA, Frank TB, Onzi G, Pohlmann AR, Guterres SS (2021) Innovative hydrogel containing polymeric nanocapsules loaded with phloretin: enhanced skin penetration and adhesion. Mater Sci Eng, C 120:111681
Casarini TPA, Frank LA, Pohlmann AR, Guterres SS (2020) Dermatological applications of the flavonoid phloretin. Eur J Pharmacol 889:173593
Chang WT, Huang WC, Liou CJ (2012) Evaluation of the anti-inflammatory effects of phloretin and phlorizin in lipopolysaccharide-stimulated mouse macrophages. Food Chem. https://doi.org/10.1016/j.foodchem.2012.03.002
Chen M, Gowd V, Wang M, Chen F, Cheng KW (2021) The apple dihydrochalcone phloretin suppresses growth and improves chemosensitivity of breast cancer cells: via inhibition of cytoprotective autophagy. Food Funct 12(1):177–190. https://doi.org/10.1039/d0fo02362k
Young CB (2019) Biochemical basis of anti-cancer-effects of phloretin—a natural dihydrochalcone. Molecules 24(2):278
Choi I, Kim J, Jeong HK, Kim B, Jou I, Park M, Chen L, Kang UJ, Zhuang X, Eun hye Joe. (2013) PINK1 deficiency attenuates astrocyte proliferation through mitochondrial dysfunction, reduced AKT and increased P38 MAPK activation, and downregulation of EGFR. Glia. https://doi.org/10.1002/glia.22475
Costea T, Vlad OC, Miclea LC, Ganea C, Szöllősi J, Mocanu MM (2020) Alleviation of multidrug resistance by flavonoid and non-flavonoid compounds in breast, lung, colorectal and prostate cancer. Int J Mol Sci. https://doi.org/10.3390/ijms21020401
Crespy V, Aprikian O, Morand C, Besson C, Manach C, Demigné C, Rémésy C (2001) Bioavailability of phloretin and phloridzin in rats. J Nutr 131(12):3227–3230. https://doi.org/10.1093/jn/131.12.3227
Cui D, Liu S, Tang M, Yongzhi Lu, Zhao M, Mao R, Wang C, Yuan Y, Li L, Chen Y (2020) Phloretin ameliorates hyperuricemia-induced chronic renal dysfunction through inhibiting NLRP3 inflammasome and uric acid reabsorption. Phytomedicine 66:153111
Davis ME (2016) Glioblastoma: overview of disease and treatment. Clin J Oncol Nurs 20(5):1–8. https://doi.org/10.1188/16.CJON.S1.2-8
Debela DT, Muzazu SGY, Heraro KD, Ndalama MT, Mesele BW, Haile DC, Kitui SK, Manyazewal T (2021) New approaches and procedures for cancer treatment: current perspectives. SAGE Open Medicine. https://doi.org/10.1177/20503121211034366
Dewanjee S, Sohel M, Hossain MS, Ansari F, Islam MT, Sultana F, Al Mamun A, Islam MM, Amin MN (2023) A Comprehensive Review on Clinically Proven Natural Products in the Management of Nerve Pain, with Mechanistic Insights. Heliyon
Duan H, Wang R, Yan X, Liu H, Zhang Y, Deguang Mu, Han J, Li X (2017) Phloretin induces apoptosis of human esophageal cancer via a mitochondria-dependent pathway. Oncol Lett 14(6):6763–6768
Fan C, Yilin Zhang Yu, Tian XZ, Teng J (2022) Phloretin enhances autophagy by impairing AKT activation and inducing JNK-beclin-1 pathway activation. Exp Mol Pathol 127(July):104814. https://doi.org/10.1016/j.yexmp.2022.104814
Frerichs H, Ball EG (1964) Studies on the metabolism of adipose tissue. XVI. Inhibition by phlorizin and phloretin of the insulin-stimulated uptake of glucose. Biochemistry 3(7):981–985
Friday E, Ledet J, Turturro F (2011) Response to dexamethasone is glucose-sensitive in multiple myeloma cell lines. J Exp Clin Cancer Res 30:1–7
Funari CS, Passalacqua TG, Rinaldo D, Napolitano A, Festa M, Capasso A, Piacente S, Pizza C, Young MCM, Durigan G (2011) Interconverting flavanone glucosides and other phenolic compounds in lippia salviaefolia Cham. Ethanol Extracts. Phytochemistry 72(16):2052–2061
FΑΟ. 2013. Food Wastage Footprint. Fao
Geohagen BC, Korsharskyy B, Vydyanatha A, Nordstroem L, LoPachin RM (2018) Phloretin cytoprotection and toxicity. Chem Biol Interact 296:117–123
Gonzalez-Menendez P, Hevia D, Rodriguez-Garcia A, Mayo JC, Sainz RM (2014) Regulation of GLUT transporters by flavonoids in androgen-sensitive and-insensitive prostate cancer cells. Endocrinology (united States). https://doi.org/10.1210/en.2014-1260
Grigoras CG, Destandau E, Fougère L, Elfakir C (2013) Evaluation of apple pomace extracts as a source of bioactive compounds. Ind Crops Prod. https://doi.org/10.1016/j.indcrop.2013.06.026
Hagl S, Deusser H, Soyalan B, Janzowski C, Will F, Dietrich H, Albert FW, Rohner S, Richling E (2011) Colonic availability of polyphenols and D-(-)-quinic acid after apple smoothie consumption. Mol Nutr Food Res. https://doi.org/10.1002/mnfr.201000252
Han L, Zhang Y, Li J, Xiao Y, Mei Lu, Li Y, Wang M (2021) Phloretin attenuation of hepatic steatosis: via an improvement of mitochondrial dysfunction by activating AMPK-dependent signaling pathways in C57BL/6J mice and HepG2 cells. Food Funct. https://doi.org/10.1039/d1fo02577e
He C, Wang J, Chakraborty T, Roy S (2020) A newfangled coordinated ruthenium phloretin complex reprogramming breast cancer microenvironment interceded by modulation of PI3K/Akt/Mtor/VEGF pathway and modifying the antioxidant status correlated with intensied apoptotic events. https://doi.org/10.21203/rs.3.rs-42460/v1
Hilt P, Schieber A, Yildirim C, Arnold G, Klaiber I, Conrad J, Beifuss U, Carle R (2003) Detection of phloridzin in strawberries (Fragaria x ananassa Duch) by HPLC− PDA− MS/MS and NMR spectroscopy. J Agric Food Chem 51(10):2896–2899. https://doi.org/10.1021/jf021115k
Hsiao Y-H, Hsieh M-J, Yang S-F, Chen S-P, Tsai W-C, Chen P-N (2019) Phloretin suppresses metastasis by targeting protease and inhibits cancer stemness and angiogenesis in human cervical cancer cells. Phytomedicine 62:152964
Huang WC, Fang LW, Liou CJ (2017) Phloretin attenuates allergic airway inflammation and oxidative stress in asthmatic mice. Front Immunol. https://doi.org/10.3389/fimmu.2017.00134
Hui J (2020) Down-regulation of aquaporin 9 by natural product phloretin mediates luminal bladder cancer migration in vitro. Johns Hopkins University
Wang H (2012) Phloretin induced apoptosis of human hepatoma cells SMMC-7721 and its correlative biological mechanisms. Afr J Pharm Pharmacol. https://doi.org/10.5897/ajpp11.869
Jacob V, Hagai T, Soliman K (2011) Structure-activity relationships of flavonoids. Curr Org Chem. https://doi.org/10.2174/138527211796367309
Jin G, Zhao Z, Chakraborty T, Mandal A, Roy A, Roy S, Guo Z (2020) Decrypting the molecular mechanistic pathways delineating the chemotherapeutic potential of ruthenium-phloretin complex in colon carcinoma correlated with the oxidative status and increased apoptotic events. Oxidative Med Cell Longev. https://doi.org/10.1155/2020/7690845
Kang D, Zuo W, Wu Q, Zhu Q, Liu P (2020) Inhibition of specificity protein 1 is involved in phloretin-induced suppression of prostate cancer. BioMed Res Int. https://doi.org/10.1155/2020/1358674
Kapoor S, Padwad YS (2023) Phloretin induces G2/M arrest and apoptosis by suppressing the β-catenin signaling pathway in colorectal carcinoma cells. Apoptosis. https://doi.org/10.1007/s10495-023-01826-4
Karpuz M, Silindir-Gunay M, Ozer AY (2018) Current and future approaches for effective cancer imaging and treatment. Cancer Biother Radiopharm. https://doi.org/10.1089/cbr.2017.2378
Kim J, Durai P, Jeon D, Jung ID, Lee SJ, Park Y-M, Kim Y (2018) Phloretin as a potent natural TLR2/1 inhibitor suppresses TLR2-induced inflammation. Nutrients 10(7):868
Kim J-L, Lee D-H, Cheol-Ho Pan Su, Park J, Sang-Cheul Oh, Lee S-Y (2022a) Role of phloretin as a sensitizer to TRAIL-induced apoptosis in colon cancer. Oncol Lett 24(3):1–13
Kim JL, Lee DH, Pan CH, Park SJ, Oh SC, Lee SY (2022b) Role of phloretin as a sensitizer to TRAIL-induced apoptosis in colon cancer. Oncol Lett. https://doi.org/10.3892/ol.2022.13441
Kim M-S, Kwon JY, Kang NJ, Lee KW, Lee HJ (2009) Phloretin induces apoptosis in H-Ras MCF10A Human breast tumor cells through the activation of P53 via JNK and P38 mitogen-activated protein kinase signaling. Ann N Y Acad Sci 1171(1):479–483
KoBoRI M, Iwashita K, Shinmoto H, Tojiro TSUSHIDA (1999) Phloretin-induced apoptosis in B16 melanoma 4A5 cells and HL60 human leukemia cells. Biosci Biotechnol Biochem 63(4):719–725
Kuruppu AI, Paranagama P, Goonasekara CL (2019) Medicinal plants commonly used against cancer in traditional medicine formulae in Sri Lanka. Saudi Pharmaceut J. https://doi.org/10.1016/j.jsps.2019.02.004
Lee SY, Park JH, Ko SH, Shim JS, Kim DD, Cho HJ (2017) Mussel-inspired hyaluronic acid derivative nanostructures for improved tumor targeting and penetration. ACS Appl Mater Interfaces 9(27):22308–22320. https://doi.org/10.1021/acsami.7b06582
Lee YJ, Kim WI, Kim SY, Cho SW, Nam HS, Lee SH, Cho MK (2019) Flavonoid morin inhibits proliferation and induces apoptosis of melanoma cells by regulating reactive oxygen species, Sp1 and Mcl-1. Arch Pharmacal Res 42(6):531–542. https://doi.org/10.1007/s12272-019-01158-5
Lin J, Sahakian D, de Morais S, Jinghai Xu, Polzer R, Winter S (2005) The role of absorption, distribution, metabolism, excretion and toxicity in drug discovery. Curr Top Med Chem. https://doi.org/10.2174/1568026033452096
Lin ST, Tu SH, Yang PS, Hsu SP, Lee WH, Ho CT, Wu CH, Lai YH, Chen MY, Chen LC (2016) Apple polyphenol phloretin inhibits colorectal cancer cell growth via inhibition of the type 2 glucose transporter and activation of p53-mediated signaling. J Agric Food Chem 64(36):6826–6837. https://doi.org/10.1021/acs.jafc.6b02861
Liu Y, Fan C, Lv Pu, Wei C, Jin H, Teng Y, Zhao M et al (2016) Phloretin induces cell cycle arrest and apoptosis of human glioblastoma cells through the generation of reactive oxygen species. J Neurooncol 128(2):217–223. https://doi.org/10.1007/s11060-016-2107-z
Lu Y, Chen J, Ren D, Yang X, Zhao Y (2017) Hepatoprotective effects of phloretin against CCl4-induced liver injury in mice. Food Hydrocolloids. https://doi.org/10.1080/09540105.2016.1258546
Lynch TJ, Bell DW, Sordella R, Gurubhagavatula S, Okimoto RA, Brannigan BW, Harris PL et al (2004) Activating mutations in the epidermal growth factor receptor underlying responsiveness of non–small-cell lung cancer to Gefitinib. N Engl J Med 350(21):2129–2139. https://doi.org/10.1056/nejmoa040938
Ma L, Wang R, Nan Y, Li W, Wang Q, Jin F (2016) Phloretin exhibits an anticancer effect and enhances the anticancer ability of cisplatin on non-small cell lung cancer cell lines by regulating expression of apoptotic pathways and matrix metalloproteinases. Int J Oncol 48(2):843–853
Ma P, Wu Y, Tao L, Han J (2022) Effect of Gardenia tea on apoptosis of human thyroid cancer cell line SW579 through PI3K-Akt pathway. Food Sci Technol 42:e73122
“Manchurian Apricot Facts and Health Benefits.”. Accessed April 7, 2023. https://www.healthbenefitstimes.com/manchurian-apricot/.
Mansoori B, Mohammadi A, Davudian S, Shirjang S, Baradaran B (2017) The different mechanisms of cancer drug resistance: a brief review. Adv Pharmaceut Bull. https://doi.org/10.15171/apb.2017.041
Mariadoss AVA, Vinyagam R, Rajamanickam V, Sankaran V, Venkatesan S, David E (2019a) Pharmacological aspects and potential use of phloretin: a systemic review. Mini-Rev Med Chem. https://doi.org/10.2174/1389557519666190311154425
Mariadoss AV, Anand RV, Senthilkumar V, Paulpandi M, Murugan K, Baojun Xu, Gothandam KM, Kotakadi VS, David E (2019b) Phloretin loaded chitosan nanoparticles augments the ph-dependent mitochondrial-mediated intrinsic apoptosis in human oral cancer cells. Int J Biol Macromol 130(June):997–1008. https://doi.org/10.1016/J.IJBIOMAC.2019.03.031
Matsui C, Takatani-Nakase T, Maeda S, Nakase I, Takahashi K (2017) Potential roles of GLUT12 for glucose sensing and cellular migration in MCF-7 human breast cancer cells under high glucose conditions. Anticancer Res 37(12):6715–6722. https://doi.org/10.21873/anticanres.12130
McBrayer SK, Cheng JC, Singhal S, Krett NL, Rosen ST, Shanmugam M (2012) Multiple myeloma exhibits novel dependence on GLUT4, GLUT8, and GLUT11: implications for glucose transporter-directed therapy. Blood J Am Soc Hematol 119(20):4686–4697
Mehmood S, Haojie Yu, Li Wang Md, Uddin A, Amin BU, Haq F, Fahad S, Haroon M (2022) Cross-linked poly(cyclotriphosphazene-co-phloretin) microspheres and their application for controlled drug delivery. Macromol Res 30(9):623–630. https://doi.org/10.1007/S13233-022-0066-0/METRICS
Min J, Li Xu, Huang K, Tang H, Ding X, Qi C, Qin X, Zhifei Xu (2015) Phloretin induces apoptosis of non-small cell lung carcinoma A549 cells via JNK1/2 and P38 MAPK pathways. Oncol Rep 34(6):2871–2879
Minsat L, Peyrot C, Brunissen F, Renault JH, Allais F (2021) Synthesis of biobased phloretin analogues: an access to antioxidant and anti-tyrosinase compounds for cosmetic applications. Antioxidants. https://doi.org/10.3390/antiox10040512
Mintie CA, Singh CK, Ahmad N (2020) Whole fruit phytochemicals combating skin damage and carcinogenesis. Transl Oncol 13(2):146–156
Molnár J, Engi H, Hohmann J, Molnár P, Deli J, Wesolowska O, Michalak K, Wang Q (2010) Reversal of multidrug resistance by natural substances from plants. Curr Top Med Chem 10(17):1757–1768
Morshed AKM, Helal SP, Hossain A, Tuli Basak Md, Sanower Hossain Md, Mehedi Hasan Md, Hasibuzzaman Al, Rahaman TI, Mia MAR, Shing P (2023) Baicalein as promising anticancer agent: a comprehensive analysis on molecular mechanisms and therapeutic perspectives. Cancers 15(7):2128
Movius EG, Phyillaier MM, Robbins J (1989) Phloretin Inhibits cellular uptake and nuclear receptor binding of triiodothyronine in human hep G2 hepatocarcinoma cells. Endocrinology. https://doi.org/10.1210/endo-124-4-1988
Nahar Metu CL, Sutihar SK, Sohel M, Zohora F, Hasan A, Miah MT, Rani Kar T, Hossain MA, Rahman MH (2023) Unraveling the signaling mechanism behind astrocytoma and possible therapeutics strategies: a comprehensive review. Cancer Reports. 6(10):e1889
Nair SV, Ziaullah Rupasinghe HV (2014) Fatty acid esters of phloridzin induce apoptosis of human liver cancer cells through altered gene expression. PLoS ONE 9(9):e107149
Nakhate KT, Badwaik H, Choudhary R, Sakure K, Agrawal YO, Sharma C, Ojha S, Goyal SN (2022) Therapeutic potential and pharmaceutical development of a multitargeted flavonoid phloretin. Nutrients 14(17):3638
Nam S, Lee SY, Cho HJ (2017) Phloretin-loaded fast dissolving nanofibers for the locoregional therapy of oral squamous cell carcinoma. J Colloid Interface Sci. https://doi.org/10.1016/j.jcis.2017.08.030
Nguyen H, Zhang S, Morris ME (2003) Effect of flavonoids on MRP1-mediated transport in panc-1 cells. J Pharm Sci 92(2):250–257. https://doi.org/10.1002/jps.10283
Nobili S, Lippi D, Witort E, Donnini M, Bausi L, Mini E, Capaccioli S (2009) Natural compounds for cancer treatment and prevention. Pharmacol Res. https://doi.org/10.1016/j.phrs.2009.01.017
Ogawa T, Sasaki A, Ono K, Ohshika S, Ishibashi Y, Yamada K (2021) Uptake of fluorescent D-and L-glucose analogues, 2-NBDG and 2-NBDLG, into human osteosarcoma U2OS cells in a phloretin-inhibitable manner. Hum Cell 34:634–643
Oliveira MR (2016) Phloretin-induced cytoprotective effects on mammalian cells: a mechanistic view and future directions. BioFactors 42(1):13–40
Park SY, Kim EJ, Shin HK, Kwon DY, Kim MS, Surh YJ, Park JHY (2007) Induction of apoptosis in HT-29 colon cancer cells by phloretin. J Med Food 10(4):581–586. https://doi.org/10.1089/jmf.2007.116
Patel BB, Alsanani A, Wondisford S, Johnson N, Banerjee D (2016) Stromal carcinoma associated fibroblasts promote drug resistance of human pancreatic cancer cells by modulation of ROS via CXCR4/CXCL12 signaling. Int J Transl Sci 2016(1):107–130
Patel BB, Ackerstaff E, Serganova IS, Kerrigan JE, Blasberg RG, Koutcher JA, Banerjee D (2017) Tumor stroma interaction is mediated by monocarboxylate metabolism. Exp Cell Res 352(1):20–33. https://doi.org/10.1016/j.yexcr.2017.01.013
Paul P, Biswas P, Dey D, Saikat ASM, Islam MA, Sohel Md, Hossain R et al (2021) Exhaustive plant profile of ‘dimocarpus longan lour’ with significant phytomedicinal properties: a literature based-review. Processes. https://doi.org/10.3390/pr9101803
Payne JN, Badwaik VD, Waghwani HK, Moolani HV, Tockstein S, Thompson DH, Dakshinamurthy R (2018) Development of dihydrochalcone-functionalized gold nanoparticles for augmented antineoplastic activity. Int J Nanomed. https://doi.org/10.2147/IJN.S143506
“Phloretin | C15H14O5 - PubChem.”.
“
“Phloretin CF | Topical Vitamin C | Vitamin C Gel | SkinCeuticals.”.
Picinelli A, Dapena E, Mangas JJ (1995) Polyphenolic pattern in apple tree leaves in relation to scab resistance a preliminary study. J Agric Food Chem. https://doi.org/10.1021/jf00056a057
Po, Ughachukwu, and Unekwe Pc. 2012. “Efflux Pump _ Mediated Resistance in Chemotherapy” 2 (2): 191–98. https://doi.org/10.4103/2141-9248.105671.
Pope ED, Kimbrough EO, Vemireddy LP, Surapaneni PK, Copland JA, Mody K (2019) Aberrant lipid metabolism as a therapeutic target in liver cancer. Expert Opin Ther Targets. https://doi.org/10.1080/14728222.2019.1615883
Prasanna S, Doerksen R (2008) Topological polar surface area: a useful descriptor in 2D-QSAR. Curr Med Chem 16(1):21–41. https://doi.org/10.2174/092986709787002817
Ranjanamala T, Vanmathiselvi K, Shreemaya R, Rajeswari SN, Sangeetha CC, Ghidan AY, Ghidan FY (2021) Virtual screening of anticancer efficacy of phloretin against apoptotic targets–An In Silico molecular docking study. J Phytol 13:152–160. https://doi.org/10.25081/JP.2021.V13.7268
Rawla P, Sunkara T, Gaduputi V (2019) epidemiology of pancreatic cancer: global trends, etiology and risk factors. World J Oncol 10(1):10–27. https://doi.org/10.14740/wjon1166
Rezk BM, Haenen GRMM, Van der Vijgh WJF, Bast A (2002) The antioxidant activity of phloretin: the disclosure of a new antioxidant pharmacophore in flavonoids. Biochem Biophys Res Commun. https://doi.org/10.1016/S0006-291X(02)00618-6
Rodriguez EB, Flavier ME, Rodriguez-Amaya DB, Amaya-Farfán J (2015) Phytochemicals and functional foods. Current situation and prospect for developing countries”. Segurança Alimentar e Nutricional. https://doi.org/10.20396/san.v13i1.1841
Roy S, Mondru AK, Chakraborty T, Das A, Dasgupta S (2022) Apple polyphenol phloretin complexed with ruthenium is capable of reprogramming the breast cancer microenvironment through modulation of PI3K/Akt/MTOR/VEGF pathways. Toxicol Appl Pharmacol. https://doi.org/10.1016/j.taap.2021.115822
Saraswati S, Alhaider A, Abdelgadir AM, Tanwer P, Korashy HM (2019) Phloretin attenuates STAT-3 activity and overcomes sorafenib resistance targeting SHP-1-mediated inhibition of STAT3 and Akt/VEGFR2 pathway in hepatocellular carcinoma. Cell Commun Signaling. https://doi.org/10.1186/s12964-019-0430-7
Schaefer S, Baum M, Eisenbrand G, Dietrich H, Will F, Janzowski C (2006) Polyphenolic apple juice extracts and their major constituents reduce oxidative damage in human colon cell lines. Mol Nutr Food Res. https://doi.org/10.1002/mnfr.200500136
Schiavano GF, De Santi M, Brandi G, Fanelli M, Bucchini A, Giamperi L, Giomaro G (2015) Inhibition of breast cancer cell proliferation and in vitro tumorigenesis by a new red apple cultivar. PLoS ONE. https://doi.org/10.1371/JOURNAL.PONE.0135840
Shang Ao, Liu H-Y, Min Luo Yu, Xia XY, Li H-Y, Ding-Tao Wu, Sun Q, Geng F, Gan R-Y (2022) Sweet Tea (Lithocarpus Polystachyus Rehd.) as a new natural source of bioactive dihydrochalcones with multiple health benefits. Crit Rev Food Sci Nutr 62(4):917–934
Shrihastini V, Muthuramalingam P, Adarshan S, Sujitha M, Chen JT, Shin H, Ramesh M (2021) Plant derived bioactive compounds, their anti-cancer effects and in silico approaches as an alternative target treatment strategy for breast cancer: an updated overview. Cancers. https://doi.org/10.3390/cancers13246222
Si W, Shen J, Zheng H, Fan W (2019) The role and mechanisms of action of MicroRNAs in cancer drug resistance. Clin Epigenetics. https://doi.org/10.1186/s13148-018-0587-8
Siegel RL, Miller KD, Wagle NS, Jemal A (2023) Cancer Statistics, 2023. CA Cancer J Clin. https://doi.org/10.3322/caac.21763
Slimestad R, Fossen T, Verheul MJ (2008) The flavonoids of tomatoes. J Agric Food Chem 56(7):2436–2441
So, Felicia V, Najla Guthrie, Ann F Chambers, Madeleine Moussa, Kenneth K Carroll, Felicia V So, Najla Guthrie, et al. 2009. “Inhibition of Human Breast Cancer Cell Proliferation and Delay of Mammary Tumorigenesis by Flavonoids and Citrus Juices Inhibition of Human Breast Cancer Cell Proliferation and Delay of Mammary Tumorigenesis by Flavonoids and Citrus Juices” 5581 (November 2017). https://doi.org/10.1080/01635589609514473.
Sohel M, Aktar S, Biswas P, Amin MA, Hossain MA, Ahmed N, Mim MI, Islam F, Mamun AA (2023) Exploring the anti-cancer potential of dietary phytochemicals for the patients with breast cancer: a comprehensive review. Cancer Med 12(13):14556–14583
Sohel Md, Partha Biswas Md, Al Amin Md, Hossain A, Sultana H, Dey D, Aktar S et al (2022a) Genistein, a potential phytochemical against breast cancer treatment-insight into the molecular mechanisms. Processes. https://doi.org/10.3390/pr10020415
Sohel Md, Islam MN, Hossain MA, Sultana T, Amit Dutta Md, Rahman S, Aktar S, Islam K, Mamun AA (2022b) Pharmacological Properties to Pharmacological Insight of Sesamin in Breast Cancer Treatment: A Literature-Based Review Study. International Journal of Breast Cancer. https://doi.org/10.1155/2022/2599689
Sohel M, Sultana H, Sultana T, Al Amin M, Aktar S, Ali MC, Rahim ZB, Hossain MA, Al Mamun A, Amin MN, Dash R (2022c) Chemotherapeutic potential of hesperetin for cancer treatment, with mechanistic insights: a comprehensive review. Heliyon. 8(1):e08815
Sohel Md, Sultana H, Sultana T, Mamun AA, Amin MN, Md Hossain Md, Ali SA, Sultana A, Rahim ZB (2022cd) Chemotherapeutic activities of dietary phytoestrogens against prostate cancer: from observational to clinical studies. Curr Pharm Des 28(19):1561–1580
Suhail S, Gupta S, Kumar V, Gupta OP (2016) Role of curcumin in oral cancer prevention. Fract Geomet Nonlinear Anal Med Biol. https://doi.org/10.15761/fgnamb.1000119
Sun YS, Zhao Z, Yang ZN, Fang Xu, Hang Jing Lu, Zhu ZY, Shi W, Jiang J, Yao PP, Zhu HP (2017) Risk factors and preventions of breast cancer. Int J Biol Sci. https://doi.org/10.7150/ijbs.21635
Szabo K, Mitrea L, Călinoiu LF, Teleky BE, Martău GA, Plamada D, Pascuta MS, Nemeş SA, Varvara RA, Vodnar DC (2022) Natural polyphenol recovery from apple-, cereal-, and tomato-processing by-products and related health-promoting properties. Molecules. https://doi.org/10.3390/molecules27227977
Szliszka E, Czuba ZP, Mazur B, Paradysz A, Krol W (2010) Chalcones and Dihydrochalcones Augment TRAIL-Mediated Apoptosis in Prostate Cancer Cells. Molecules 15(8):5336–5353
Tang J, Gong Y (2021) Synergistic effect of phloretin combined with radiotherapy on lung cancer. Int J Radiat Oncol Biol Phys. https://doi.org/10.1016/j.ijrobp.2021.07.809
Tuli HS, Rath P, Chauhan A, Ramniwas S, Vashishth K, Varol M, Jaswal VS, Haque S, Sak K (2022a) Phloretin, as a potent anticancer compound: from chemistry to cellular interactions. Molecules 27(24):8819
Tuli HS, Rath P, Chauhan A, Ramniwas S, Vashishth K, Varol M, Jaswal VS, Haque S, Sak K (2022b) Phloretin, as a potent anticancer compound: from chemistry to cellular interactions. Molecules. https://doi.org/10.3390/molecules27248819
Ulaszewska M, Vázquez-Manjarrez N, Garcia-Aloy M, Llorach R, Mattivi F, Dragsted LO, Praticò G, Manach C (2018) Food intake biomarkers for apple, pear, and stone fruit lars dragsted. Genes Nutr. https://doi.org/10.1186/s12263-018-0620-8
Wu C-H, Ho Y-S, Tsai C-Y, Wang Y-J, Tseng H, Wei P-L, Lee C-H, Liu R-S, Lin S-Y (2009a) In vitro and in vivo study of phloretin-induced apoptosis in human liver cancer cells involving inhibition of type ii glucose transporter. Int J Cancer 124(9):2210–2219
Wu CH, Ho YS, Tsai CY, Wang YJ, Tseng H, Wei PL, Lee CH, Liu RS, Lin SY (2009b) In vitro and in vivo study of phloretin-induced apoptosis in human liver cancer cells involving inhibition of type II glucose transporter. Int J Cancer. https://doi.org/10.1002/ijc.24189
Wu F, Zhou Y, Li L, Shen X, Chen G, Wang X, Liang X, Tan M, Huang Z (2020) Computational approaches in preclinical studies on drug discovery and development. Front Chem. https://doi.org/10.3389/fchem.2020.00726
Wu KH, Ho CT, Chen ZF, Chen LC, Whang-Peng J, Lin TN, Ho YS (2018) The apple polyphenol phloretin inhibits breast cancer cell migration and proliferation via inhibition of signals by type 2 glucose transporter. J Food Drug Anal 26(1):221–231. https://doi.org/10.1016/j.jfda.2017.03.009
Xintaropoulou C, Ward C, Wise A, Marston H, Turnbull A, Langdon SP (2015) A comparative analysis of inhibitors of the glycolysis pathway in breast and ovarian cancer cell line models. Oncotarget 6(28):25677
Xue X, Liang XJ (2012) Overcoming drug efflux-based multidrug resistance in cancer with nanotechnology. Chin J Cancer. https://doi.org/10.5732/cjc.011.10326
Yang C-H, Yen-Chuan Ou, Lin C-C, Lin Y-S, Tung M-C, Chia-Cheng Yu, Lin J-T, Wen C-Y (2022) Phloretin in benign prostate hyperplasia and prostate cancer: a contemporary systematic review. Life 12(7):1029
Yang G, Yin X, Ma D, Su Z (2020) Anticancer Activity of Phloretin against the Human Oral Cancer Cells Is Due to G0/G1 Cell Cycle Arrest and ROS Mediated Cell Death. J BUON 25:344–349
Yang KC, Tsai CY, Wang YJ, Wei PL, Lee CH, Chen JH, Chih Hsiung Wu, Ho YS (2009) Apple polyphenol phloretin potentiates the anticancer actions of paclitaxel through induction of apoptosis in human hep G2 cells. Mol Carcinog. https://doi.org/10.1002/mc.20480
You Q, Xu JP, Zhu ZX, Hu Z, Cai QP (2020) Phloretin flavonoid exhibits selective antiproliferative activity in doxorubicin-resistant gastric cancer cells by inducing autophagy, inhibiting cell migration and invasion, cell cycle arrest and targeting ERK1/2 MAP pathway. J BUON 25:308–313
Zhang S, Morris ME (2003) Effects of the flavonoids biochanin a, morin, phloretin, and silymarin on p-glycoprotein-mediated transport. J Pharmacol Exp Ther. https://doi.org/10.1124/jpet.102.044412
Zhao W, Bai B, Hong Z, Zhang X, Zhou B (2020) Berbamine (BBM), a natural STAT3 inhibitor, synergistically enhances the antigrowth and proapoptotic effects of sorafenib on hepatocellular carcinoma cells. ACS Omega. https://doi.org/10.1021/acsomega.0c03527
Zhao Y, Fan Y, Wang M, Wang J, Cheng J, Zou J, Zhang X, Shi Y, Guo D (2020) Studies on pharmacokinetic properties and absorption mechanism of phloretin: in vivo and in vitro. Biomed Pharmacother 132:110809
Zhou Mo, Zheng J, Bi J, Xinye Wu, Lyu J, Gao K (2018) Synergistic inhibition of colon cancer cell growth by a combination of atorvastatin and Phloretin. Oncol Lett 15(2):1985–1992. https://doi.org/10.3892/ol.2017.7480
Zhu S-P, Liu G, Xiao-Ting Wu, Chen F-X, Liu J-Q, Zhou Z-H, Zhang J-F, Fei S-J (2013a) The effect of Phloretin on human Γδ T cells killing colon cancer SW-1116 cells. Int Immunopharmacol 15(1):6–14
Zhu SP, Liu G, Xiao Ting Wu, Chen FX, Liu JQ, Zhou ZH, Zhang JF, Fei SJ (2013b) The effect of phloretin on human Γδ t cells killing colon cancer SW-1116 cells. Int Immunopharmacol 15(1):6–14. https://doi.org/10.1016/j.intimp.2012.11.001
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sohel, M., Nity, N.U., Sarker, M.R. et al. Exploring the chemotherapeutic potential and therapeutic insight of phloretin against human malignancies: a systematic review. Phytochem Rev (2024). https://doi.org/10.1007/s11101-024-09938-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11101-024-09938-8