
Abstract
In this article, an eco-epidemiological model with strong Allee effect in prey population growth is presented by a system of delay differential equations. The time lag in terms of the delay parameter corresponds to the predator gestation period. We inspect elementary mathematical characteristic of the proposed model such as uniform persistence, stability and Hopf bifurcation at the interior equilibrium point of the system. We execute several numerical simulations to illustrate the proposed mathematical model and our analytical findings. We use basic tools of nonlinear dynamic analysis as first return maps, Poincare sections and Lyapunov exponents to identify chaotic behavior of the system. We observe that the system exhibits chaotic oscillation due to the increase of the delay parameter. Such chaotic behavior can be suppressed by the strength of Allee effect.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The research on eco-epidemiology is the investigation into ecological systems with the impact of epidemiological parameters. Hadeler Freedman [34] first modeled the spread of disease among predator–prey interacting populations. Chattopadhyay and Arino [13] coined the term eco-epidemiology for such systems. Since then, the research on eco-epidemiology as well as its biological importance has gained great attention [5, 6, 14, 15, 37, 68, 90, 95].
In recent times, significant research has been done on the development of the concept for Allee effect, which corresponds to the positive correlation between population size/density and per-capita growth rate at low population density [1, 17, 49, 60, 62]. Empirical evidence of Allee effects has been reported in many natural populations including plants [29, 33], insects [47], marine invertibrates [83], birds and mammals [20]. Few mechanisms behind this Allee effect are due to complications in finding mates, reproductive facilitation, predation, environment conditioning, inbreeding depression, etc. [22, 42, 53, 71, 80–82]. Allee effects mainly classified into two broad categories: strong Allee effect and weak Allee effect [21, 64, 65, 92]. There is a threshold population level for the strong Allee effect such that the species become extinct below this threshold population density [27]. On the other hand, the weak Allee effect occurs when the growth rate reduces but remains positive at low population density. Ecologists paid significant attention on this topic as it relates to species extinction [3, 8, 26, 72, 88]. Recently, Saha et al. [66], analyzed the time series of two herring populations from the Icelandic and Canadian regions from the Global Population dynamics database with GPDD ID 1765,1759 [59] and observed the presence of strong Allee effect. Furthermore, disease has been considered as one of the main cause for species disappearance, and if it is connected with the Allee effects, the interaction between them has substantial biological importance in nature [38]. Recently, many researchers have been done in Allee effects on interacting population [e.g., see [43, 45, 94]] as well as Allee effects with disease in the population [38, 39, 69, 70, 89, 99]. Many species suffer from Allee effect and disease. For example, the combined effects of disease and Allee effect has been observed in the African wild dogs [11, 19] and the island fox [4, 18]. Thus, understanding the combined impact of Allee effects and disease on population dynamics of predator–prey interactions can enrich us to have better knowledge on species abundance and disease outbreak.
Many ecological and men-made activities in biology, and medicine can be better interpreted with the help of time delays[56, 67, 91, 96]; the classical books such as MacDonald [54], Gopalsamy [32] and Kuang [46] discussed detail topics on the relevance of time delays in practical models. There have been extensive research activities on the dynamical behaviors, periodic oscillation, persistence, bifurcation and chaos of population with retarded systems [7, 23–25, 28, 50, 51, 58, 63, 73, 74, 76–79, 84–86, 100, 102]. Since ignoring time delays means ignoring reality, thus without delays, dynamical models become a worse conjecture of reality. [16, 30–32, 46, 52, 93, 97] tell us that for digesting food the predator requires some time as time delay. As far as our knowledge, there are very few works on time-delayed population dynamics in the presence of Allee effect [9, 10, 12, 61, 98, 101]. According to the authors, this is the first noble attempt of considering the time delay effects for eco-epidemic models with strong Allee in the prey.
Recently, Kang et al. [44] studied a model of a predator–prey interaction with infected prey, and prey is subjected to the strong Allee effects. In this paper, we have considered the same model with a more realistic assumption that the predator takes some time to digest food before having further activities to take place. In addition to the above assumption, we have also considered that the predator population captures only infected prey which contributes toward its positive growth. We like to mention that Lafferty and Morris [48] experimentally established that the predation rate on infected population is thirty- one times higher than the susceptible one. Thus, based on this experimental result, we ignore the predation on susceptible one.
The rest of the article is organized as follows: Sect. 2 deals with the development of the model; in Sect. 3, we discuss the dynamics of the model with no time delay. Detailed mathematical analysis of the time delay model, its boundedness of solutions, existence of switching stability, persistence and permanence, the direction and stability of Hopf bifurcation are discussed in the Sect. 4. Dynamical behavior of the model without Allee effect is studied in 5. Sensitivity analysis and numerical simulations are presented in the Sect. 6. The article ends with a discussion in Sect. 7.
2 The model
In this paper, we have considered the model studied by Kang et al. [44] with the following two important realistic assumptions:
-
1.
We have neglected the predation on susceptible prey by the predator population; thus, in our model, predator feeds only on infected prey. This assumption is supported by the experimental evidence by Lafferty and Morris [48]; they evaluated that the predation rate on infected fish, on an average, is thirty-one times higher than the predation rate on susceptible fish. In addition, we have assumed that the consumption of infected prey contributes positive growth to the predator population.
-
2.
After consumption of prey by predator, some amounts of energy in the form of prey biomass converted into predator biomass through a very complicated internal process in the predators digest system. The whole transformation process requires time; thus, we have taken a constant time lag \(\tau {\,>\,} 0\) for the gestation of predator [16, 30–32, 46, 52, 93, 97].
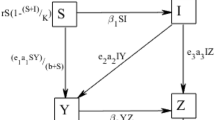
Thus, a delayed predator–prey model with susceptible prey, infected prey and predator, with the strong Allee effects on susceptible prey, is described by the following set of nonlinear differential equations:
Here the infected population does not contribute to the reproduction but compete for resources with susceptible one. We have considered disease transmitted through mass action law, and predator follows Holling type I functional response [40]. All the variables and parameters are positive. The variables and parameters used in Model (1) are presented in the Table 1.
The initial conditions for the system (1) take the form
where \(\psi =(\psi _{1}, \psi _{2}, \psi _{3})^{T}\in {\mathcal {C_{+}}} \) such that \(\psi _{i}(\phi )\ge 0 \) \((i=1, 2, 3)\), \(\forall \) \( \phi \in [-\tau , 0 ] \), and \({\mathcal {C_{+}}}\) denotes the Banach space \( {\mathcal {C_{+}}}\left( [-\tau , 0 ],{\mathbb {R}}^{3}_{+}\right) \) of continuous functions mapping the interval \([-\tau , 0 ]\) into \({\mathbb {R}}^{3}_{+}\) and denotes the norm of an element \(\psi \) in \({\mathcal {C_{+}}}\) by
For biological feasibility, we further assume that \(\psi _{i}(0)> 0\), for \(i=1, 2, 3\).
3 Mathematical analysis of the system (1) with no time lag \(\left( \tau = 0\right) \)
We first study the system (1) with no time lag. The system (1) without delay for gestation of predator can be written as
The system (2) has the following boundary equilibria:
The system (2) has two interior equilibria \(E_1^* = \left( S_1^*, I_1^*, P_1^*\right) \) and \(E_2^* = \left( S_2^*, I_2^*, P_2^*\right) \), where \(I_1^* = \frac{d}{\alpha }=I_2^*\), \(P_1^* = \frac{1}{a}\left( \beta S_1^* - \mu \right) \), \(P_2^* = \frac{1}{a}\left( \beta S_2^* - \mu \right) \) and \(S_1^*,~S_2^*\) are the roots of the quadratic equation
thus,
where \(B = \left( \theta +1-\frac{d}{\alpha }\right) \) and \(C = \left( 1-\frac{d}{\alpha }\right) \theta +\beta \frac{d}{\alpha }\).
Proposition 1
[Local stability of equilibria for the Model (2)] The local stability of equilibria for the Model (2) is summarized in Table 2.
Detailed proof is given in the “Appendix” section.
3.1 Disease-free case for the system (2)
In the absence of disease, the system (2) has only three boundary equilibria (0, 0, 0), \((\theta , 0, 0)\) and (1, 0, 0) where the predator also can not survive, since predator feeds only on infected prey. In this case, (0, 0, 0) and (1, 0, 0) are locally asymptotically stable, whereas \((\theta , 0, 0)\) is always unstable. Existence of the predator population depends on the persistence of the disease among the prey population. If the prey species is disease free, then the system is predator free also. Thus, the infection in prey population is responsible for the survival of the predator.
Remarks
Thus, the system (2) is infection free in the following three situations:
-
1.
Extinction of all the three populations if \(S(0) < \theta \), then \(\lim _{t\rightarrow \infty }\left( S(t), I(t), P(t)\right) = E_0.\)
-
2.
Only susceptible prey is able to survive if \(\frac{\beta }{\mu } < 1\). In this case, both the boundary equilibria \(E_0\) and \(E_1\) are locally asymptotically stable. The system (2) goes to the boundary equilibrium \(E_1\), if initial conditions start from the basin of attraction of \(E_1\); otherwise, it goes to the extinction equilibrium.
-
3.
Disease driven extinction : if \(\frac{\beta }{\mu } > \frac{1}{\theta }\) and \(\frac{\alpha (1-\theta )}{d}\le 1\), then the system (2) converges to the extinction equilibrium \(E_0\), for any initial conditions in \({\mathbb {R}}^3_+\) [44]. The biological explanation for such situation is that the transmission of disease is large enough so that the susceptible population density drops below its threshold density (which is known as Allee threshold [27]), and eventually susceptible population goes to extinction, which drive the other populations to extinction.
Allee effect makes the system prone to extinction, and initial conditions are significant characteristics for the existence of susceptible prey as well as other species. If initial condition for susceptible prey population is less than Allee threshold (\(S(0) < \theta \)), then prey population extinct and, consequently, predator population also goes to extinction. Now if \(\mu >\beta \), i.e., death rate of infected prey is higher than disease transmission rate, then disease does not persist in the system. In contrast, if \(\mu <\beta \) and \(\frac{\beta }{\mu }<\frac{1}{\theta }\), then disease starts to persist in the system, and the system converges to predator-free equilibrium. We observe that all the species coexist if \( S_2^*>\frac{\mu }{\beta }\), i.e., \(\beta S_2^*> \mu \) and \(\alpha >d\). Here \(\beta S_2^*\) represents per-capita infection rate and \(\mu \) represents the per-capita death rate. Therefore, all the species persist in the system if the per-capita infection rate is higher than the per-capita death rate of infected prey and the net gain to P by consuming I is greater than the natural death rate of predator.
4 Mathematical analysis of the system (1)
In this section, we have studied the delay model (1). Here we have performed local stability analysis of equilibria, permanence, and existence of switching stability of the delay differential Eq. (1).
4.1 Local stability analysis
Let \(\widetilde{E} ~=~ \left( \widetilde{S},~\widetilde{I},~\widetilde{P}\right) \) be any equilibrium point of the system (1). The linearized system of the system (1) at \(\widetilde{E} ~=~ \left( \widetilde{S},~\widetilde{I},~\widetilde{P}\right) \) is
So, the characteristic equation of the delayed system (1) around any equilibrium point \(\widetilde{E} = (\widetilde{S}, \widetilde{I}, \widetilde{P})\) is given by
The following transcendental equation represents the characteristic equation at the interior equilibrium \(E_2^* = \left( S_2^*, I_2^*, P_2^*\right) \) of the dynamical system (1):
where
For the delay-induced system (1), it is known that if all the roots of the corresponding characteristic Eq. (5) have negative real parts, then the equilibrium point \(E_2{^*}\) will be asymptotically stable. But, we cannot use the classical Routh–Hurwitz criterion to investigate the stability of the dynamical system. We need the sign of the real parts of the roots of the characteristic Eq. (5), to determine the nature of the stability.
Let \(\lambda (\tau )=\zeta (\tau )+i\rho (\tau )\) be the eigenvalue of the characteristic Eq. (5); substituting this value in Eq. (5), we obtain real and imaginary parts, respectively, as
and
A necessary condition for a stability changes of \(E_2{^*}\) is that the characteristic Eq. (5) should have purely imaginary solutions. We set \(\zeta =0\) in (7) and (8). Then, we get
Eliminating \(\tau \) by squaring and adding the Eqs. (9) and (10), we get the algebraic equation for determining \(\rho \) as
Substituting \(\rho ^{2} = \theta \) in Eq. (11), we obtain a cubic equation given by
where
Now \(\sigma _{3}<0\) implies that (12) has at least one positive root. The next theorem gives a criterion for the switching in the stability behavior of \(E_2{^*}\).
Theorem 1
Suppose that \(E_2^{*}\) exists and is locally asymptotically stable for (1) with \(\tau =0\). Also let \(\theta _{0}=\rho ^{2}_{0}\) be a positive root of (12).
-
1.
Then, there exists \(\tau = \tau ^{*}\) such that the interior equilibrium point \(E_2{^*}\) of the delay system (1) is asymptotically stable when \(0\le \tau <\tau ^{*}\) and unstable for \(\tau >\tau ^{*}\).
-
2.
Furthermore, the system will undergo a Hopf bifurcation at \({E_2{^*}}\) when \(\tau =\tau ^{*}\), provided \(Z(\rho )X(\rho )-Y(\rho )W(\rho )>0\).
Proof
We already know that \(\rho ^{}_{0}\) is a solution of the Eq. (11), i.e. the characteristic Eq. (5) has a pair of purely imaginary roots \(\pm i\rho ^{}_{0}\). From Eqs. (9) and (10), we have that \(\tau ^{*}_{p}\) is a function of \(\rho _{0}\) for \(p=0,~ 1,~ 2,\ldots \), which is given by
Now, the system will be locally asymptotically stable around the interior equilibrium point \(E_2{^*}\) for \(\tau =0\), if the condition (27) holds. In that case by Butler’s lemma [30], \(E_2{^*}\) will remain stable for \(\tau <\tau ^{*}\), such that \(\tau ^{*} = \displaystyle {\min _{p\ge 0}} \ \tau ^{*}_{p}\).
Also, we can verify the following transversality condition
Differentiating Eqs. (7) and (8), with respect to \(\tau \) and then put \(\zeta =0\), we obtain
where
Solving the above system, we have \(\frac{\hbox {d}}{\hbox {d}\tau }[\hbox {Re}\{\lambda (\tau )\}]_{\tau =\tau ^{*}, \rho =\rho _0}=\frac{Z(\rho )X(\rho )-Y(\rho )W(\rho )}{X^2(\rho )+Y^2(\rho )}_{\tau =\tau ^{*}, \rho =\rho _0}\), which shows that \(\frac{\hbox {d}}{\hbox {d}\tau }[\hbox {Re}\{\lambda (\tau )\}]_{\tau =\tau ^{*},\rho =\rho _0}>0\) if \(Z(\rho )X(\rho )-Y(\rho )W(\rho )>0\). Therefore, Hopf bifurcation occurs at \(\tau =\tau ^*\) as the transversality condition is satisfied. This proves the theorem.
Remark
For the system (1), at most finite number of stability switching is possible.
We rewrite characteristic Eq. (5) in the form as
where
In our system (1), \(M(\lambda )=\lambda ^{3}+B_{1}\lambda ^{2}+B_{2}\lambda +B_{3}\), \( N(\lambda )= -[B_{4}\lambda ^{2}+B_{5}\lambda +B_{6}]\).
Now, we have the following results:
-
1.
\(M(\lambda )\) and \( N(\lambda )\) have no common imaginary root and both analytic functions in \(\hbox {Re}(\lambda ) >0\).
-
2.
\(\overline{M(-iy)} = M(iy)\), \(\overline{N(-iy)} = N(iy)\) \(\forall \ y\).
-
3.
\(M(0) + N(0) =B_{3}-B_{6}\ne 0\).
-
4.
\(\displaystyle {\limsup _{{|\lambda |\rightarrow +\infty }}} \Big |\frac{N(\lambda )}{M(\lambda )}\Big |=0 < 1\).
-
5.
\({\varPsi }(y) = |M(iy)|^{2} - |N(iy)|^{2} = y^{6} + \sigma _{1}y^{4} + \sigma _{2}y^{2} + \sigma _{3} = 0\), which is a cubic expression in \(y^{2}\), where \(\sigma _{1}=B_{1}^{2}-2B_{2}-B_{4}^{2} \), \(\sigma _{2}=B_{2}^{2}-2B_{1}B_{3}-2B_{4}B_{6}-B_{5}^{2}\) and \(\sigma _{3}=B_{3}^{2}-B_{6}^{2}\). Since \(\sigma _{3}\) is negative, so \({\varPsi }(y)\) must have at least one positive root.
From Kuang [46], we conclude that at most finite number of stability switching is possible for the system (1).
4.2 Uniform persistence of the system
In this section, we present the conditions for uniform persistence of the system (1). We denote by \({\mathbb {R}}^3_+ = \{(S, I, P)\in {\mathbb {R}}^3 : S\ge 0,I\ge 0, P\ge 0\}\) the nonnegative quadrant and by int\(({\mathbb {R}}^3_+) = \Big \{(S, I, P)\in {\mathbb {R}}^3: S>0, I>0, P>0\Big \}\).
Definition
System (1) is said to be uniformly persistent if a compact region \(D \subset \text{ int } ({\mathbb {R}}^3_+)\) exists such that every solution \({\varXi } (t) = \left( S(t), I(t), P(t)\right) \) of the system (1) with initial conditions eventually enters and remains in the region D.
4.3 Boundedness of the solution of the delayed system (1)
The first equation of the system (1) can be written as
Integrating between the limits 0 and t, we have
Similarly, from the second and the third equations of the system, we have
and

where \(S(0)= S_0 > 0\), \(I(0)= I_0 > 0\) and \(P(0)= P_0 > 0\). Therefore, \(S(t) > 0\), \(I(t) > 0\) and \(P(t) > 0\).
Proposition 2
All the solutions of the system (1) starting in int\(({\mathbb {R}}^3_+)\) are uniformly bounded with an ultimate bound.
Proof
We know that (for detailed proof, please see the reference Kang et al. [44])
Define a function \(V_1 = \frac{1}{a}I(t-\tau ) + \frac{1}{\alpha }P(t)\).
Taking its time derivative along the solution of the system (1), we get
So,
where \(M=\frac{\beta }{\min \{\mu ,d\}}.\)
4.4 Permanence
In order to prove permanence of the system (1), we use the uniform persistence theory for infinite dimensional systems [35]. Let X be a complete metric space. Suppose that \(X^{0}\) is open and dense in X and \(X^{0}\cap X_{0}={\varPhi }\). Assume that T (t) is a \(C^{0}\) semigroup on X satisfying
Let \(T_{b}(t) = T (t)\big |_{X_{0}}\) and let \(A_{b}\) be the global attractor for \(T_{b}(t)\). To investigate the permanence of the system (1), the following lemmas are useful.
Lemma 1
[35] If T(t) satisfies (17) and we have the following:
-
(i) there is a \(t_{0}\ge 0\) such that T (t) is compact for \( t \ge t_{0}\);
-
(ii) T (t) is a point dissipative in X;
-
(iii) \(\widehat{A_{b}}=\cup _{x\in A_{b}} \omega (x) \) is isolated and thus has an acyclic covering \(\widehat{M}\), where \(\widehat{M}= \{M_{1},M_{2}, \ldots ,M_{n}\}\);
-
(iv) \(W^{s}( M_{i} ) \cap X^{0} = {\varPhi } \) for \(i = 1, 2, \ldots , n.\)
Then, \(X_{0}\) is a uniform repeller with respect to \(X^{0}\), i.e., there is an \(\epsilon > 0\) such that for any \(x \in X^{0}\), \(\displaystyle {\lim _{t\rightarrow \infty }inf \ d(T(t)x, X_{0} )\ge \epsilon }\), where d is the distance of T(t)x from \(X_{0}\).
Lemma 2
[95] Consider the following differential equation:
where \(A,~ B,~ C,~ \tau > 0\); \(x(t) > 0\), for \(-\tau \le t\le 0\). Then, we have
-
(i) if \(A < B\), then \(\displaystyle {\lim _{t\rightarrow \infty } x(t)} = 0;\)
-
(ii) if \(A > B\), then \(\displaystyle {\lim _{t\rightarrow \infty } x(t)} = + \infty .\)
Theorem 2
[Permanence of the model (1)] System (1) is permanent, provided
-
(i) \(\left( 1-\frac{\mu + a\epsilon _1}{\beta }\right) \left( \frac{\mu + a\epsilon _1}{\beta }-\theta \right) >0,\) where \(\epsilon _{1}\) is sufficiently small and
-
(ii) \(\alpha (I_2-\epsilon _2)>d\) where \(I_2=\frac{\left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) }{\frac{\mu }{\beta }+\beta -\theta }.\)
We have given the detailed proof of the above permanence conditions for the model (1) in the “Appendix” section.
4.5 The direction and stability of Hopf-bifurcating periodic solutions
We already know that the system (1) will undergo Hopf bifurcation at endemic equilibrium \(E_{*} = \left( S_*,I_*,P_*\right) \) when \(\tau \) passes through \(\tau ^{*}\) (for our convenience, we assume that \(E_{*} = E_2^*\)). In this section, we investigate the direction, stability and period of Hopf-bifurcating solutions at \(\tau = \tau ^{*}\) for system (1) by using the techniques of normal form theory and center manifold theorem introduce by Hassard et al. [36].
We have given the detailed analysis for the direction and stability of the Hopf bifurcation in the “Appendix” section. From the detailed analysis, we can compute the following quantities, which determine the direction, stability and the periods of the bifurcating periodic solutions of Hopf bifurcation.
We can compute the following quantities (please see in the “Appendix” section):
It is well known that \(\mu _{2}\) and \(\beta _{2}\) determine the direction of Hopf bifurcation and stability of bifurcating periodic solutions. If \(\mu _{2}>0\) \((<0)\), and \(\beta _{2}<0\) \((>0)\), which imply that the Hopf bifurcation is supercritical (subcritical), bifurcating periodic solutions are exist for \(\tau >\tau ^{*}\) (\(\tau < \tau ^{*}\)) and are orbitally stable (unstable). Moreover, \(\tau _{2}\) determines the periods of the bifurcating periodic solutions and the period increases (decreases) if \(\tau _{2}>0\) \((<0)\).
5 Mathematical analysis of the system (1) with no Allee effect
Without Allee effect, the system (1) reduces to
Xiao and Chen [95] already discussed similar kind of model. In this section, we only mention the dynamical features of the system (18).
5.1 Mathematical analysis of the system (18) with no time delay
The system (18) with out delay parameter can be written as
The system (19) has the following boundary equilibria:
The system (19) has one interior equilibria \(E_3^* = \left( S_3^*, I_3^*, P_3^*\right) \), where \(I_3^* = \frac{d}{\alpha }\), \(P_3^* = \frac{1}{a}\left( \beta S_3^* - \mu \right) \) and \(S_3^*=1-(\beta +1)\frac{d}{\alpha }\). Here, \(E^2\) exists if \(1-\frac{\mu }{\beta }>0\), and \(E_3^*\) exists if \(\beta \left( 1-(\beta +1)\frac{d}{\alpha }\right) >\mu \).
Proposition 3
[Local stability of equilibria for the Model (19)] The local stability of equilibria of the Model (19) is summarized in Table 3.
Proof
We omit the proof here, as it is easy to verify.
5.2 Mathematical analysis of the system (18)
The characteristic equation of the delayed system (18) at the interior equilibrium \( E_3{^*} = \left( S_3{^*}, I_3{^*}, P_3{^*}\right) \) is given by
Where
Let \(\lambda (\tau )=i\rho (\tau )\) be the eigenvalue of the characteristic Eq. (20). Substituting this value in Eq. (20) and proceeding as the Sect. 4, we give a criterion for the switching in the stability behavior of \(E_3{^*}\).
Theorem 3
Suppose that \(E_3{^*}\) exists and is locally asymptotically stable for (18) with \(\tau =0\).
-
1.
Then, there exists a \(\tau = \tau _{*}\) such that the interior equilibrium point \(E_3{^*}\) of the delay system (18) is asymptotically stable when \(0\le \tau <\tau _{*}\) and unstable for \(\tau >\tau _{*}\).
-
2.
Furthermore, the system will undergo a Hopf bifurcation at \({E_3{^*}}\) when \(\tau =\tau _{*}\), provided \(Z_1(\rho )X_1(\rho )-Y_1(\rho )W_1(\rho )>0\).
Where
6 Sensitivity analysis and numerical simulation for the model (1)
In this section, we try to find the parameters having significant impact upon the predator density, the response of interest. The extent of impact of the input parameter on response are measured by partial rank correlation coefficients (PRCC) along with their p values, calculated using Latin Hypercube sampling scheme described in Marino et al. [55]. First step is to assume a suitable prior distribution on the parameters. An uniform prior over a biologically feasible range for each parameters is appropriate. The PRCC values and their significance are given in the adjoining Table 1. We set the dimension of the sample in LHS to \(N=100\) and \(N=1000\) with the time points 20,000 h. The model output chosen for P is predator density. Our PRCC analysis suggests that predator density is mainly affected by the parameter a.
6.1 Numerical simulations
In this section, we give the time series diagram, phase plane diagram and bifurcation diagram of the system (1) to compare the results and to support our analytical findings for different values of \(\tau \). Extensive numerical simulations have been performed for various values of parameters to determine the global dynamics. This study is not only provide local stability and Hopf bifurcation but also demonstrate the feasibility of different complex dynamical behavior, including limit cycles and chaos.

Stability of the interior equilibrium for the nondelayed model (2), when \(\beta = 0.395\), \(\mu = 0.05\), \(d = 0.1\), \(a = 0.1\), \(\alpha = 0.85\) and \(\theta = 0.1\), with different initial conditions

The figure depicts local stability of the interior equilibrium for the delayed system (1), with the time delay \(\tau = 1 ~(< \tau ^{*} = 5.39)\), when \(\beta = 0.395\), \(\mu = 0.05\), \(d = 0.1\), \(a = 0.1\), \(\alpha = 0.85\) and \(\theta = 0.1\), with the initial value \([S(0),~I(0),~P(0)] = [0.3,~0.2,~5]\). The first row left figure is the time series for the susceptible prey; the first row right figure is the time series for the infected prey; the second row left figure is the time series for the predator; and the second row right figure is the 3D phase portrait of the stable interior of (1)
Consider the set of the parameter as \(\beta =0.395;~ \mu =0.05;~ d=0.1;~ a=0.1;~ \alpha =0.85; and ~ \theta =0.1.\) For the above set of parameter values, we obtain the interior equilibrium point \(E_1^*=(0.16,0.11,0.15)\) which is an unstable equilibrium and \(E_2^*=(0.82,0.12,2.73)\) is a stable focus as we see that all the trajectories initiating inside the region of attraction approach toward the equilibrium point \(E_2^*=(0.82,0.12,2.73)\) (see Fig. 1). We choose different initial densities of \(\left[ S(0), I(0), P(0)\right] \) as \(\left[ 0.5,0.1,0.2\right] \), \(\left[ 0.3,0.4,0.6\right] \), \(\left[ 0.8,0.1,2\right] \), \(\left[ 0.2,0.2,0.5\right] \) and draw the phase portrait of the system (2) in Fig. 1.
From our analytical findings, it is observed that \(E_2^*\) is locally asymptotically stable for \(\tau <\tau ^{*}=5.39\). Figure 2 shows the simulation result for the model system (1) with \(\tau =1<\tau ^{*}\). Interior equilibrium point \(E_2^*\) looses its stability as \(\tau \) passes through its critical value \(\tau >\tau ^{*}\), and the system (1) experiences Hopf bifurcation as \(\frac{\hbox {d}}{\hbox {d}\tau }[\hbox {Re}\{\lambda (\tau )\}]_{\tau =\tau ^{*},\,\rho =\rho _0}=0.0030>0.\) From 4.5, the nature of the stability and direction of the periodic solution bifurcating from the interior equilibrium \(E_2^*\) at the critical point \(\tau ^{*}\) can be computed. Through simulations, we compute that






It shows the existence of bifurcating periodic solution, and it is supercritical and stable as evident from Fig. 3. Figure 3 shows that the limit cycle is stable around the coexisting equilibrium point \(E_2^*\). The solutions starting from two different initial values converge to the limit cycle oscillation, while \(\tau =5.5\). Figure 4 shows the limit cycle oscillations for \(\tau =10\) with same parameter values as in Fig. 1 and initial conditions have been fixed to \(S(0)=0.8,~ I(0)=0.1,~ P(0)=2.2\). Further, we increase the value of time delay (\(\tau \)) and observe that the system (1) shows two periodic solutions for \(\tau =15\) (see Fig. 5) and chaotic oscillations for \(\tau =28\) (see Fig. 6). A Poincare map of a typical chaotic region is shown in Fig.7a for \(\tau =28\). The scattered distribution of the sampling points implies the chaotic behavior of the system. We also draw the return map for the system (1) in 7(b). Here we plot \(P_{\mathrm{max}}(n)\) versus \(P_{\mathrm{max}}(n+1)\). The data fall on a smooth one-dimensinal map. We observe that the map is unimodal, like the logistic map.
We plot the Lyapunov exponents with time delay \(\tau \) in Fig. 8a, with the parameter values \(\beta = 0.395\), \(\mu = 0.05\), \(d = 0.1\), \(a = 0.1\), \(\alpha = 0.85\) and \(\theta = 0.1\). We use Wolf algorithm and [75] to calculate Lyapunov exponents. Here the red-colored Lyapunov exponent is positive for \(\tau \ge 20\) (for clear visualization, see Fig. 8b), green is approximately zero and other two Lyapunov exponents are always negative. So, we can conclude that the system (1) is chaotic for \(28\ge \tau \ge 20\).

Bifurcation diagrams with respect to \(\tau \) for the system (1), when the other parameters are fixed as \(\beta = 0.395\), \(\mu = 0.05\), \(d = 0.1\), \(a = 0.1\), \(\alpha = 0.85\) and \(\theta = 0.1\). a Bifurcation diagram for the susceptible prey; b bifurcation diagram for the infected prey; and c bifurcation diagram for the predator

Bifurcation diagrams with respect to \(\theta \) for the system (1). When the other parameters are fixed as above and \(\tau = 20\). a Bifurcation diagram for the susceptible prey; b bifurcation diagram for the infected prey; and c bifurcation diagram for the predator

Waveform plot and phase plane for the delayed system (18), with the time delay \(\tau = 1.75(< \tau _{*} = 3.62)\), with the initial value \([S(0),~I(0),P(0)] = [0.8,~0.1,~2.2]\). The first row left figure is the time series for the susceptible prey; the first row right figure is the time series for the infected prey; the second row left figure is the time series for the predator and the second row right figure is the 3D phase portrait of the stable interior of (18)
To make it straightforward, we draw the bifurcation diagram with respect to time delay \(\tau \) for \(0< \tau \le 28\). The bifurcation diagram with respect to time delay \(\tau \) drawn in Fig. 9 represents the complex dynamical feature from the limit cycle to chaos in our proposed model (1). For the gradual increase in the delay parameter, the system (1) switches its stability from stable focus to limit cycle oscillation to chaotic oscillation. Figure 9 shows that for \(\tau \in [0,5.39)\) the interior equilibria \(E^*\) are stable; for \(\tau \in (5.39,18.4]\), it shows limit cycle oscillations; and for \(\tau \in (18.4,28]\), it exhibits higher periodic and chaotic oscillations.
Next, we draw the bifurcation diagram with respect to the parameter \(\theta \) for \(\theta \in (0,0.42]\) where \(\tau \) is fixed at 20. The complex dynamic behavior including chaos of the delayed system with respect to \(\theta \) is evident from Fig. 10. We observe that for \(\theta \in (0,0.12]\) the system (1) shows chaotic and higher periodic oscillations. As \(\theta \) crosses its threshold value 0.12, the system (1) shows limit cycle behavior for \(\theta \in (0.12,0.32]\) and the system becomes stable for \(\theta \in (0.32,0.42]\). The changes in the bifurcation diagrams suggest that chaos may be reduced/removed with the changes in the strength of Allee effect.

Waveform plot and phase plane for the delayed system (18), with the time delay \(\tau =4\). Figure indicates the existence of limitcycle
Time delay plays a similar kind of role in the model (18) like (1). We give a numerical example to support the Theorem 3 about the switching of stability at the coexistence equilibrium \(E_3^*\). Keeping the parameter values fixed as \(\beta = 0.395\), \(\mu = 0.05\), \(d = 0.1\), \(a = 0.1\) and \(\alpha = 0.85\) with the initial values as \(S(0)=0.8,~ I(0)=0.1,~ P(0)=2.2\), we draw the figures for the model (18). We observe that the system (18) undergoes local Hopf bifurcation at \(\tau =3.62\). We notice that for \(\tau = 1.75 ~(< \tau _{*} = 3.62)\) the coexistence equilibrium \(E_3^*\) is locally stable (Fig. 11). When delay parameter \(\tau \) crosses \(\tau _{*}\) the system exhibits limit cycle. Figure 12 depicts periodic oscillation for the system at \(\tau =4.5>\tau _{*}\). Chaotic behavior for the system is observed at \(\tau =60\) (Fig. 13).

Solution curves and the phase plane showing chaotic attractor at \(\tau =60\) for the model (18)
7 Discussion
In this article, we make an attempt to discuss the impact of a time delay parameter \(\tau \) in the model described by Kang et al. [44]. The model (1) has two interior equilibria \(E_1^*\) and \(E_2^*\), while the second model (18) has only one interior equilibria \(E_3^*\). We keep our main focus on the model (1) as we want to check the effect of Allee on our dynamical system. For our model, the gestation delay plays a crucial role. Time delay can switch the stability of the equilibrium points; that is, for some critical value \(\tau ^*\), the positive equilibrium \(E_2^*\) is stable when \(\tau <\tau ^*\), and it becomes unstable as \(\tau \) crosses through its critical magnitude from lower to higher values. We have proved that the system (1) experiences the Hopf bifurcation as the delay parameter \(\tau \) crosses the critical value \(\tau ^*\). Further increasing in the delay parameter beyond the bifurcation point leads to complex dynamic behavior, including chaos. The explicit formulae which determine the stability, direction and other properties of bifurcating periodic solution are calculated, by using normal form and center manifold theorem and then using numerical simulations; we illustrate our analytical results. We also discussed the complex dynamical behavior of the system (18) without Allee effect.
From ecological aspect, chaos has utmost biological importance. Many theoretical studies reveal that many important ecosystem features such as predictability, species persistence [2] and biodiversity [41] can be affected by chaos. Various chaos control schemes have been explored till now. In both the systems (1) and (18), we observed chaotic oscillations, but Allee effect plays an vital factor in the system (1). The impact of the Allee effect on the stability of population models can show different dynamics which strictly depend on the assumptions of the corresponding model. So, the idea that chaos can be enhanced by the Allee effect ([57]) is not always true but in this context, we have shown chaos can be controlled by Allee threshold \(\theta \). The changes in the bifurcation diagrams (Fig. 10) suggest that chaos may be reduced/removed with the changes in the strength of Allee effect.
To the best of our knowledge, stability and Hopf bifurcation are not discussed in any article, for the delayed dynamical system when the reproduction rate of prey population is influenced by strong Allee effect in the eco-epidemiology model. We hope that our findings in this article will certainly help the ecologists and, as a consequence, it may enrich theoretical ecology.
References
Allee, W.C.: Animal Aggregations. A Study in General Sociology. University of Chicago Press, Chicago (1931)
Allen, J.C., Schaffer, W.M., Rosko, D.: Chaos reduces species extinctions by amplifying local population noise. Nature 364, 229–232 (1993)
Amarasekare, P.: Interactions between local dynamics and dispersal: insights from single species models. Theor. Popul. Biol. 53, 44–59 (1998)
Angulo, E., Roemer, G.W., Berec, L., Gascoigen, J., Courchamp, F.: Double Allee effects and extinction in the island fox. Conserv. Biol. 21, 1082–1091 (2007)
Bairagi, N., Roy, P.K., Chattopadhyay, J.: Role of infection on the stability of a predator–prey system with several response functions—a comparative study. J. Theor. Biol. 248, 10–25 (2007)
Bate, M.A., Hilker, M.F.: Disease in group-defending prey can benefit predators. Theor. Ecol. 7, 87–100 (2014)
Beretta, E., Kuang, Y.: Convergence results in a well-known delayed predator–prey system. J. Math. Anal. Appl. 204, 840–853 (1996)
Berezovskaya, F.S., Song, B., Castillo-Chavez, C.: Role of prey dispersal and refuges on predator–prey dynamics. SIAM J. Appl. Math. 70, 1821–1839 (2010)
Biswas, S., Sasmal, S.K., Samanta, S., Saifuddin, Md, Ahmed, Q.J.K., Chattopadhyay, J.: A delayed eco-epidemiological system with infected prey and predator subject to the weak Allee effect. Math. Biosci. 263, 198–208 (2015)
Biswas, S., Sasmal, S.K., Saifuddin, Md, Chattopadhyay, J.: On existence of multiple periodic solutions for Lotka–Volterra’s predator–prey model with Allee effects. Nonlinear Stud. 22(2), 189–199 (2015)
Burrows, R., Hofer, H., East, M.L.: Population dynamics, intervention and survival in African wild dogs (Lycaon pictus). Proc. R. Soc. B Biol. Sci. 262, 235–245 (1995)
Celik, C., Merdan, H., Duman, O., Akin, O.: Allee effects on population dynamics with delay. Chaos Solitons Fractals 37, 65–74 (2008)
Chattopadhyay, J., Arino, O.: A predator–prey model with disease in the prey. Nonlinear Anal. 36, 747–766 (1999)
Chattopadhyay, J., Bairagi, N.: Pelicans at risk in Salton Sea—an eco-epidemiological study. Ecol. Model. 136, 103–112 (2001)
Chattopadhyay, J., Pal, S.: Viral infection on phytoplankton–zooplankton system—a mathematical model. Ecol. Model. 151, 15–28 (2002)
Chen, Y., Yu, J., Sun, C.: Stability and Hopf bifurcation analysis in a three-level food chain system with delay. Chaos Solitons Fractals 31, 683–694 (2007)
Chunjin, W., Chen, L.: Periodic solution and heteroclinic bifurcation in a predator–prey system with allee effect and impulsive harvesting. Nonlinear Dyn. 76(2), 1109–1117 (2014)
Clifford, D.L., Mazet, J.A.K., Dubovi, E.J., Garcelon, D.K., Coonan, T.J., Conrad, P.A., Munson, L.: Pathogen exposure in endangered island fox (Urocyon littoralis) populations: implications for conservation management. Biol. Conserv. 131, 230–243 (2006)
Courchamp, F., Clutton-Brock, T., Grenfell, B.: Multipack dynamics and the Allee effect in the African wild dog (Lycaon pictus). Anim. Conserv. 3, 277–285 (2000)
Courchamp, F., Grenfell, B., Clutton-Brock, T.: Impact of natural enemies on obligately cooperatively breeders. Oikos 91, 311–322 (2000)
Courchamp, F., Berec, L., Gascoigne, J.: Allee Effects in Ecology and Conservation. Oxford University Press, Oxford (2008)
Courchamp, F., Clutton-Brock, T., Grenfell, B.: Inverse density dependence and the Allee effect. Trends Ecol. Evol. 14, 405–410 (1999)
Cushing, J.M.: Periodic time-dependent predator–prey systems. SIAM J. Appl. Math. 32, 82–95 (1997)
Das, P., Mukandavire, Z., Chiyaka, C., Sen, A., Mukherjee, D.: Bifurcation and chaos in S-I-S epidemic model. Differ. Equ. Dyn. Syst. 17(4), 393–417 (2009)
Dong, T., Liao, X.: Bogdanov–Takens bifurcation in a trineuron BAM neural network model with multiple delays. Nonlinear Dyn. 71(3), 583–595 (2013)
Drake, J.: Allee effects and the risk of biological invasion. Risk Anal. 24, 795–802 (2004)
Elaydi, S., Sacker, R.J.: Population models with Allee effect: a new model. J. Biol. Dyn. 4, 397–408 (2010)
Faria, T.: Stability and bifurcation for a delay predator–prey model and the effect of diffusion. J. Math. Anal. Appl. 254, 433–463 (2001)
Ferdy, J., Austerlitz, F., Moret, J., Gouyon, P., Godelle, B.: Pollinator-induced density dependence in deceptive species. Oikos 87, 549–560 (1999)
Freedman, H.I., Rao, V.S.H.: The tradeoff between mutual interference and time lag in predator prey models. Bull. Math. Biol. 45, 991–1004 (1983)
Freedman, H.I., So, J., Waltman, P.: Coexistence in a model of competition in the chemostat incorporating discrete time delays. SIAM J. Appl. Math. 49, 859–870 (1989)
Gopalsamy, K.: Stability and Oscillation in Delay Differential Equation of Population Dynamics. Kluwer Academic, Dordrecht (1992)
Groom, M.: Allee effects limit population viability of an annual plant. Am. Nat. 151, 487–496 (1998)
Hadeler, K.P., Freedman, H.I.: Predator-prey populations with parasitic infection. J. Math. Biol. 27, 609–631 (1989)
Hale, J.K., Waltman, P.: Persistence in infinite-dimensional systems. SIAM J. Math. Anal. 20(2), 388–395 (1989)
Hassard, B., Kazarinof, D., Wan, Y.: Theory and Applications of Hopf Bifurcation. Cambridge University Press, Cambridge (1981)
Hethcote, H.W., Wang, W., Han, L., Ma, Z.: A predator–prey model with infected prey. Theor. Popul. Biol. 66, 259–268 (2004)
Hilker, F.M., Langlais, M., Petrovskii, S.V., Malchow, H.: A diffusive SI model with Allee effect and application to FIV. Math. Biosci. 206, 61–80 (2007)
Hilker, F.M., Langlais, M., Malchow, H.: The allee effect and infectious diseases: extinction, multistability, and the (dis-)appearance of oscillations. Am. Nat. 173, 72–88 (2009)
Holling, C.S.: Some characteristics of simple types of predation and parasitism. Can. Entomol. 91, 385–398 (1959)
Huisman, J., Weissing, F.J.: Biodiversity of plankton by species oscillations and chaos. Nature 402, 407–410 (1999)
Jacobs, J.: Cooperation, optimal density and low density thresholds: yet another modification of the logistic model. Oecologia 64, 389–395 (1984)
Jang, S.R.J.: Allee effects in a discrete-time host-parasitoid model. J. Differ. Equ. Appl. 12, 165–181 (2006)
Kang, Y., Sasmal, S.K., Bhowmick, A.R., Chattopadhyay, J.: Dynamics of a predator–prey system with prey subject to Allee effects and disease. Math. Biosci. Eng. 11(4), 877–918 (2014)
Kang, Y., Sasmal, S.K., Bhowmick, A.R., Chattopadhyay, J.: A host-parasitoid system with predation-driven component Allee effects in host population. J. Biol. Dyn. 9, 213–232 (2015)
Kuang, Y.: Delay Differential Equation with Applications in Population Dynamics. Academic Press, New York (1993)
Kuussaari, M., et al.: Allee effect and population dynamics in the Glanville fritillary butterfly. Oikos 82, 384–392 (1998)
Lafferty, K.D., Morris, A.K.: Altered behaviour of parasitized killifish increases susceptibility to predation by bird final hosts. Ecology 77, 1390–1397 (1996)
Leonel, R.J., Prunaret, D.F., Taha, A.K.: Big bang bifurcations and Allee effect in blumbergs dynamics. Nonlinear Dyn. 77(4), 1749–1771 (2014)
Liao, M.X., Tang, X.H., Xu, C.J.: Dynamics of a competitive Lotka–Volterra system with three delays. Appl. Math. Comput. 217, 10024–10034 (2011)
Liao, M.X., Tang, X.H., Xu, C.J.: Bifurcation analysis for a three-species predator–prey system with two delays. Commun. Nonlinear Sci. Numer. Simul. 17, 183–194 (2012)
Liu, Z., Yuan, R.: Stability and bifurcation in a delayed predator–prey system with Beddington–DeAngelis functional response. J. Math. Anal. Appl. 296, 521–537 (2004)
Lamont, B., Klinkhamer, P., Witkowski, E.: Population fragmentation may reduce fertility to zero in Banksia goodii-demonstration of the Allee effect. Oecologia 94, 446–450 (1993)
MacDonald, N.: Biological Delay Systems: Linear Stability Theory. Cambridge University Press, Cambridge (1989)
Marino, S., Hogue, I.B., Ray, C.J., Kirschner, D.E.: A methodology for performing global uncertainty and sensitivity analysis in systems biology. J. Theor. Biol. 254, 178–196 (2008)
Meng, X.Y., Huo, H.F., Zhang, X.B., Xiang, H.: Stability and hopf bifurcation in a three-species system with feedback delays. Nonlinear Dyn. 64, 349–364 (2011)
Morozov, A., Petrovskii, S., Li, B.L.: Bifurcations and chaos in a predator–prey system with the allee effect. Proc. R. Soc. B 271, 1407–1414 (2004)
Mukandavire, Z., Garira, W., Chiyaka, C.: Asymptotic properties of an HIV/AIDS model with a time delay. J. Math. Anal. Appl. 330(2), 916–933 (2007)
NERC: The global population dynamics database version 2. Centre for population biology, Imperial College. http://www.sw.ic.ac.uk/cpb/cpb/gpdd.html (2010)
Pablo, A.: A general class of predation models with multiplicative allee effect. Nonlinear Dyn. 78(1), 629–648 (2014)
Pal, P.J., Saha, T., Sen, M., Banerjee, M.: A delayed predator–prey model with strong Allee effect in prey population growth. Nonlinear Dyn. 68, 23–42 (2012)
Peng, F., Kang, Y.: Dynamics of a modified lesliegower model with double allee effects. Nonlinear Dyn. 80, 1051–1062 (2015)
Ruan, S., Wei, J.: On the zeros of transcendental functions with applications to stability of delay differential equations with two delays. Dyn. Contin. Discrete Impuls. Syst. Ser. A 10, 863–874 (2003)
Rocha, J.L., Fournier-Prunaret, D., Taha, A.K.: Strong and weak Allee effects and chaotic dynamics in Richards’ growths. Discrete Contin. Dyn. Syst. Ser. B 18(9), 2397–2425 (2013)
Rocha, J.L., Fournier-Prunaret, D., Taha, A.K.: Big bang bifurcations and allee effect in blumbergs dynamics. Nonlinear Dyn. 77(4), 1749–1771 (2014)
Saha, B., Bhowmick, A.R., Chattopadhyay, J., Bhattacharya, S.: On the evidence of an Allee effect in herring populations and consequences for population survival: a model-based study. Ecol. Model. 250, 72–80 (2013)
Sarwardi, S., Haque, M., Mandal, P.K.: Ratio-dependent predator–prey model of interacting population with delay effect. Nonlinear Dyn. 69, 817–836 (2012)
Sasmal, S.K., Chattopadhyay, J.: An eco-epidemiological system with infected prey and predator subject to the weak Allee effect. Math. Biosci. 246, 260–271 (2013)
Sasmal, S.K., Bhowmick, A.R., Al-Khaled, K., Bhattacharya, S., Chattopadhyay, J.: Interplay of functional responses and weak Allee effect on pest control via viral infection or natural predator: an eco-epidemiological study. Differ. Equ.Dyn. Syst. 24(1), 21–50 (2016)
Sasmal, S.K., Kang, Y., Chattopadhyay, J.: Intra-specific competition in predator can promote the coexistence of an eco-epidemiological model with strong Allee effects in prey. BioSyst. 137, 34–44 (2015)
Schreiber, S.J.: Allee effects, extinctions, and chaotic transients in simple population models. Theor. Popul. Biol. 64, 201–209 (2003)
Shi, J., Shivaji, R.: Persistence in reaction diffusion models with weak Allee effect. J. Math. Biol. 52, 807–829 (2006)
Song, Y., Han, M.: Stability and Hopf bifurcation in a competitive Lokta-Volterra system with two delays. Chaos Solitons Fractals 22, 1139–1148 (2004)
Song, Y., Wei, J.: Local Hopf bifurcation and global periodic solutions in a delayed predator–prey system. J. Math. Anal. Appl. 301, 1–21 (2005)
Sprott, J.C.: A simple chaotic delay differential equation. Phys. Lett. A 366, 397–402 (2007)
Sun, C., Loreau, M.: Dynamics of a three-species food chain model with adaptive traits. Chaos Solitons Fractals 41(5), 2812–2819 (2009)
Sun, C., Han, M., Lin, Y.: Analysis of stability and Hopf-bifurcation for a delayed logistic equation. Chaos Solitons Fractals 31(3), 672–682 (2007)
Sun, C., Han, M., Lin, Y., Chen, Y.: Global qualitative analysis for a predator–prey system with delay. Chaos Solitons Fractals 32(4), 1582–1596 (2007)
Sun, K., Liu, X., Zhu, C., Sprott, J.C.: Hyperchaos and hyperchaos control of the sinusoidally forced simplified lorenz system. Nonlinear Dyn. 69, 1383–1391 (2012)
Sther, B.-E., Ringsby, T., Rskaft, E.: Life history variation, population processes and priorities in species conservation: towards a reunion of research paradigms. Oikos 77, 217–226 (1996)
Stephens, P., Sutherland, W.: Consequences of the Allee effect for behaviour, ecology and conservation. Trends Ecol. Evol. 14, 401–405 (1999)
Stephens, P.A., Sutherland, W.J., Freckleton, R.P.: What is the Allee effect? Oikos 87, 185–190 (1999)
Stoner, Allan W., Ray-Culp, Melody: Evidence for Allee effects in an over-harvested marine gastropod: density-dependent mating and egg production. Mar. Ecol. Prog. Ser. 202, 297–302 (2000)
Tang, X., Cao, D., Zou, X.: Global attractivity of positive periodic solution to periodic Lotka–Volterra competition systems with pure delay. J. Differ. Equ. 128(2), 580–610 (2006)
Tang, X.H., Zou, X.: 3/2-type criteria for global attractivity of LotkaVolterra competition system without instantaneous negative feedbacks. J. Differ. Equ. 186(2), 420–439 (2002)
Tang, X.H., Zou, X.: Global attractivity of non-autonomous Lotka–Volterra competition system without instantaneous negative feedback. J. Differ. Equ. 192(2), 502–535 (2003)
Taylor, A.E., David, C.L.: Introduction to Functional Analysis, vol. 2. Wiley, New York (1958)
Taylor, C., Hastings, A.: Allee effects in biological invasions. Ecol. Lett. 8, 895–908 (2005)
Thieme, H.R., Dhirasakdanon, T., Han, Z., Trevino, R.: Species decline and extinction: synergy of infectious diseases and Allee effect? J. Biol. Dyn. 3, 305–323 (2009)
Venturino, E.: Epidemics in predator-prey models: disease in the predators. IMA J. Math. Appl. Med. Biol. 19, 185–205 (2002)
Wang, J., Jiang, W.: Bifurcation and chaos of a delayed predator–prey model with dormancy of predators. Nonlinear Dyn. 69, 1541–1558 (2012)
Wang, M.H., Kot, M.: Speeds of invasion in a model with strong or weak Allee effects. Math. Biosci. 171, 83–97 (2001)
Wang, W., Ma, Z.: Harmless delays for uniform persistence. J. Math. Anal. Appl. 158, 256–268 (1991)
Wang, J., Shi, J., Wei, J.: Predator–prey system with strong Allee effect in prey. J. Math. Biol. 62, 291–331 (2011)
Xiao, Y., Chen, L.: Modelling and analysis of a predator–prey model with disease in the prey. Math. Biosci. 171, 59–82 (2001)
Xu, C., Tang, X., Liao, M., He, X.: Bifurcation analysis in a delayed lotka-volterra predator–prey model with two delays. Nonlinear Dyn. 66, 169–183 (2011)
Xua, R., Gan, Q., Ma, Z.: Stability and bifurcation analysis on a ratio-dependent predator–prey model with time delay. J. Comput. Appl. Math. 230, 187–203 (2009)
Yan, J., Zhao, A., Yan, W.: Existence and global attractivity of periodic solution for an impulsive delay differential equation with Allee effect. J. Math. Anal. Appl. 309, 489–504 (2005)
Yakubu, A.A.: Allee effects in a discrete-time SIS epidemic model with infected newborns. J. Differ. Equ. Appl. 13, 341–356 (2007)
Zhang, L., Guo, S.J.: Hopf bifurcation in delayed van der Pol oscillators. Nonlinear Dyn. 71(3), 555–568 (2013)
Zhang, X., Zhang, Q.L., Xiang, Z.: Bifurcation analysis of a singular bioeconomic model with Allee effect and two time delays. In: Abstract and Applied Analysis 2014 (2014)
Zhao, T., Kuang, Y., Smith, H.L.: Global existence of periodic solutions in a class of delayed Gause- type predator–prey system. Nonlinear Anal. 28, 1373–1394 (1997)
Acknowledgments
SB’s and MD’s research is supported by the junior research fellowship from the University Grants Commission, Government of India. SS’s research work is supported by NBHM postdoctoral fellowship. JC’s research is partially supported by a DAE project (Ref No. 2/48(4)/2010-R & D II/8870).
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
1.1 Proof of the Proposition 1:
Proof
The Jacobian matrix of the model (2) at its any equilibrium point \((S_{*}, I_{*}, P_{*})\) is described as follows
After substituting \(E_i = \left( S^*,I^*,P^*\right) ,~ i~=~0,~\theta ,~1,~2\) into (23), we obtain the eigenvalues for each equilibrium:
-
1.
\(E_0 = \left( 0, 0, 0\right) \) is always locally asymptotically stable since eigenvalues associated with (23) at \(E_0\) can be presented as follows:
$$\begin{aligned} \lambda _1= & {} -\theta ~~ \left( <0\right) ,\quad \lambda _2 = -\mu ~~ (<0) \quad \hbox {and}\\ \lambda _3= & {} -d~~ (<0). \end{aligned}$$ -
2.
\(E_\theta = \left( \theta , 0, 0\right) \) is always unstable since eigenvalues associated with (23) at \(E_1\) are given by,
$$\begin{aligned} \lambda _1= & {} \theta (1-\theta ) ~~(>0),\\ \lambda _2= & {} \beta \theta -\mu \biggl \{^{<0\quad \text{ if } \beta \theta <\mu }_{>0\quad \text{ if } \beta \theta >\mu }\quad \text{ and } \lambda _3 = -d \left( <0\right) . \end{aligned}$$ -
3.
\(E_1 = \left( 1, 0, 0\right) \) is locally asymptotically stable if \(\beta < \mu \) since eigenvalues associated with (23) at \(E_1\) are given by,
$$\begin{aligned} \lambda _1= & {} \theta -1 ~~(<0),\\ \lambda _2= & {} \beta -\mu \biggl \{^{<0\quad \text{ if } \beta <\mu }_{>0\quad \text{ if } \beta >\mu }~~~~ \text{ and } \lambda _3 = -d \left( <0\right) . \end{aligned}$$ -
4.
\(E_{2} = \left( \frac{\mu }{\beta }, \frac{\left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) }{\frac{\mu }{\beta }+\beta -\theta },0\right) \) is locally asymptotically stable if \(\alpha \frac{\left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) }{\frac{\mu }{\beta }+\beta -\theta } -d<0\) and \(1<\frac{\beta }{\mu }<\min \Bigg \{\frac{1}{\theta },\frac{\beta -\theta +\sqrt{\beta ^2-\beta \theta +\beta }}{\beta +\beta \theta -\theta ^2}\Bigg \}\). Since the eigenvalues associated with (23) at \(E_{2}\) are given by
$$\begin{aligned} \lambda _3= & {} \alpha \frac{\left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) }{\frac{\mu }{\beta }+\beta -\theta }\\&-d\biggl \{^{<0\quad \text{ if } \alpha \frac{\left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) }{\frac{\mu }{\beta }+\beta -\theta } -d<0}_{>0\quad \text{ if } \alpha \frac{\left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) }{\frac{\mu }{\beta }+\beta -\theta } -d>0} \end{aligned}$$and the other two eigenvalues are the roots of the equation
$$\begin{aligned} \lambda ^2 - A\lambda +B = 0. \end{aligned}$$Where
$$\begin{aligned} \begin{array}{lll} A &{}=&{} \frac{\mu }{\beta }\left( 1-2\frac{\mu }{\beta }-\frac{\left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) }{\frac{\mu }{\beta }+\beta -\theta }+\theta \right) \\ &{}=&{} \frac{\left( \beta +\beta \theta -\theta ^2\right) \left( \frac{\beta }{\mu }\right) ^2-2\frac{\beta }{\mu }(\beta -\theta )-1}{\left( \frac{\beta }{\mu }\right) ^2\left( 1+\frac{\beta }{\mu }(\beta -\theta )\right) }\\ B &{}=&{} \mu \left( \frac{\mu }{\beta }-\theta \right) \left( 1-\frac{\mu }{\beta }\right) >0 \end{array} \end{aligned}$$(24)Thus, we have
$$\begin{aligned}&A>0\quad \text{ if } \frac{\beta }{\mu }>\frac{\beta -\theta +\sqrt{\beta ^2-\beta \theta +\beta }}{\beta +\beta \theta -\theta ^2} \text{ while } \\&A<0\quad \text{ if } \frac{\beta }{\mu }<\frac{\beta -\theta +\sqrt{\beta ^2-\beta \theta +\beta }}{\beta +\beta \theta -\theta ^2}. \end{aligned}$$Therefore, \(E_2\) exists and is locally asymptotically stable if
$$\begin{aligned} 1<\frac{\beta }{\mu }<\min \Bigg \{\frac{1}{\theta },\frac{\beta -\theta +\sqrt{\beta ^2-\beta \theta +\beta }}{\beta +\beta \theta -\theta ^2}\Bigg \} \end{aligned}$$
Again, the Jacobian matrix of the model (2) at its interior equilibrium point \(\widetilde{E} = (\widetilde{S}, \widetilde{I}, \widetilde{P})\) can be written as follows
where its characteristic equation reads as follows:
with \(\lambda _i, i = 1,2,3\) being roots of (26). If all the real parts of \(\lambda _i, i=1,2,3\) are negative, then we have
Thus, interior equilibrium is locally asymptotically stable if the following conditions hold
So, from (27), we see that \(E_2^*\) is stable and \(E_1^*\) is unstable.
1.2 Proof of the permanence of the model (1)
Proof
In this section, we shall prove that the boundary planes of \({\mathbb {R}}^{3}_{+}\) repel the positive solutions of system (1) uniformly. Let us define
where \(C([-\tau , 0],~{\mathbb {R}}^{3}_{+})\) denote the space of continuous function mapping \([-\tau , 0]\) in to \({\mathbb {R}}^{3}_{+}\).
If \(C_{0} = C_{1}\cup C_{2}\cup C_{3}\) and \(C^{0} = int C\left( [-\tau , 0], {\mathbb {R}}^{3}_{+}\right) \), it suffices to show that there exists an \(\epsilon _{0} >0 \) such that for any solution \(u_{t}\) of system (1) initiating from \(C_{0}\), \(\displaystyle {\lim _{t\rightarrow \infty }\inf \ d(u_{t}, C_{0} )\ge \epsilon _{0}}\).
Now, we verify below that the conditions of Lemma 1 are satisfied. By definition of \(C_{0}\) and \(C^{0}\) and system (1), it is easy to see that \(C_{0}\) and \(C^{0}\) are positively invariant. Moreover, it is clear that conditions (i) and (ii) of Lemma 1 are satisfied. Thus, we need to confirm conditions (iii) and (iv).
Three constant solutions in \(C_{0}\) corresponding to \(\left( S(t) = 0, I(t) = 0, P(t) = 0\right) \), \(\left( S(t)=1, I(t)=0, P(t)=0\right) \) and \(\left( S(t) = S_{2}, I(t) = I_{2}, P(t) = 0\right) \) are respectively \(E_{0}\), \(E_{1}\) and \(E_{2}\).
If \(\left( S(t), I(t), P(t)\right) \) is any solution of system (1) initiating from \(C_{1}\) with \(\psi _{1}(0) = 0\) then \(S(t)\rightarrow 0\), \(I(t)\rightarrow 0\), \(P(t)\rightarrow 0\) as \(t\rightarrow \infty \). If \(\left( S(t), I(t), P(t)\right) \) is a solution of system (1) initiating from \(C_{2}\) with \(\psi _{1}(0) > 0\), it follows that \(S(t)\rightarrow 1\), \(I(t)\rightarrow 0\), \(P(t)\rightarrow 0\) as \(t\rightarrow \infty \). If \(\left( S(t), I(t), P(t)\right) \) is a solution of system (1) initiating from \(C_{3}\) with \(\psi _{1}(0) \psi _{2}(0) > 0\), it follows that \(S(t)\rightarrow S_{2}\), \(I(t)\rightarrow I_{2}\), \(P(t)\rightarrow 0\) as \(t\rightarrow \infty \).
This shows that invariant sets \(E_{0}\), \(E_{1}\) and \(E_{2}\) are isolated invariant, and then, \(E_{0}\), \(E_{1}\) and \(E_{2}\) are an isolated as well as an acyclic covering, satisfying condition (iii) of Lemma 1.
We now show that \(W^{s}(E_{0}) \cap C^{0} = {\varPhi }\), \(W^{s}(E_{1}) \cap C^{0} = {\varPhi }\) and \(W^{s}(E_{2}) \cap C^{0} = {\varPhi }\). The proof for the first part is simple, so we ignore it. We shall prove the second part through contradiction. Let us assume that \(W^{s}(E_{1}) \cap C^{0} \ne {\varPhi }\), then there exists a positive solution \(\left( S(t), I(t), P(t)\right) \) of system (1) such that \(\left( S(t), I(t), P(t)\right) \rightarrow (1, 0, 0)\) as \(t\rightarrow +\infty \). Let us choose \(\epsilon _{1} > 0 \) small enough such that
for some \(t>t_{1}\), where \(t_{1}\) be sufficiently large. Then, from first and second equations of the system (1), we have for \(t>t_{1}\)
Now let us consider
Let \(V = (v_1,v_2)\) and \(\zeta >0\) be small enough such that \(\zeta v_1<S(t_1),\zeta v_2<I(t_1)\). If \(\left( y_1(t),y_2(t)\right) \) is a solution of system (29) satisfying \(y_i(t_1) = \zeta v_i, i = 1, 2\). We know from comparison theorem that \(S(t) > y_1(t),~ I(t) > y_2(t)\) for all \(t>t_1\). We can check easily that the system (29) has a unique positive equilibrium
Now \(S(t) > y_1(t), I(t) > y_2(t)\) for all \(t>t_1\) and \(\displaystyle {\lim _{t\rightarrow \infty } y_2(t)}=y^*_2\). This is a contradiction. Hence, \(W^{s}(E_{2} ) \cap C^{0} = {\varPhi }\).
Let \(W^{s}(E_{2}) \cap C^{0} \ne {\varPhi }\). Then, there exists a positive solution \(\left( S(t), I(t), P(t)\right) \) of the system such that \(\left( S(t), I(t), P(t)\right) \rightarrow (S_{2}, I_{2}, 0)\) as \(t\rightarrow \infty \). Let us choose \(\epsilon _2>0\) small enough such that \(I_2-\epsilon _2 < I(t) < I_2+\epsilon _2\) for \(t > t_2-\tau \).
Then, from third equation of the system (1), we have for \(t > t_{2}-\tau \)
Now, let us consider
Let \(u_{1}\) and \(v>0\) be small enough such that \(vu_{1}<P(t_{2})\). If \(z_{1}\) is a solution of system (31) satisfying \(z_{1}(t_{2})=wu_{1}\), we know from comparison theorem, \(P(t)\ge z_{1}(t)\) for all \(t >t_{2}-\tau \). We also observe that the solution \(z_{1}\) of Eq. (31) satisfies \(\displaystyle {\lim _{t\rightarrow \infty } z_{1}(t)} \rightarrow +\infty \) (from condition (ii)).
Since \(P(t)\ge z_{1}(t)\) for all \( t >t_{2} \), so \(\displaystyle {\lim _{t\rightarrow \infty } P(t)}\nrightarrow 0\). This contradicts that \(W^{s}(E_{2} ) \cap C^{0} = {\varPhi }\). From Lemma 1, we conclude that \(C_{0}\) repels the positive solutions of (1) uniformly. Hence, the system (1) is permanent. This proves the theorem.
1.3 Direction and stability of Hopf bifurcation of model (1)
We consider the transformation \(z_{1}(t) = S(\tau t)-S_{*}\), \(z_{2}(t) = I(\tau t)-I_{*}\), \(z_{3}(t) = P(\tau t)-P_{*}\).
Let \(\tau =\tau ^{*}+\mu \), \(\mu \in \mathbf {R}\). Then, \(\mu =0\) is the Hopf bifurcation value of the system (1). The Eq. (1) can be written in the form
where \(z(t)=(z_{1}(t), z_{2}(t), z_{3}(t))^{T}\in \mathbf {R^{3}}\). For \(\psi =(\psi _{1}, \psi _{2}, \psi _{3})^{T}\in \mathbf {C}([-1, 0], \mathbf {R^{3}_{+}})\); \(L_{\mu }: \mathbf {C}\rightarrow \mathbf {R}\) and \(F: \mathbf {R}\times \mathbf {C}\rightarrow \mathbf {R}\) are given by
and
where
By the Riesz representation theorem [87], there exists a function \(\eta (\theta , \mu )\) of bounded variation for \(\theta \in [-1, 0]\), such that
In fact, we can choose
where \(\delta \) is defined by \(\delta (\theta )=\Big \{^{1, \ \ \theta =0,}_{0, \ \ \theta \ne 0.} \)
For \(\psi \in \mathbf {C}^{1}\left( [-1, 0], \mathbf {R^{3}_{+}}\right) \), define
and
Then, the system (32) is of the form
where \(z_{t}(\theta )=z_{t}(t+\theta )\) for \(\theta \in [-1, 0]\).
For \(\phi \in \mathbf {C}^{1}([0, 1], (\mathbf {R^{3}_{+}})^{*})\), define
and a bilinear inner product
where \(\eta (\theta )=\eta (\theta , 0)\). Clearly, A(0) and \(A^{*}\) are adjoint operators. We know that \(\pm i\rho _{0}\tau ^{*}\) are eigenvalues of A(0). So, they are also eigenvalues of \(A^{*}\). Now we search for the eigenvector of A(0) and \(A^{*}\) corresponding to \(i\rho _{0}\tau ^{*}\) and \(-i\rho _{0}\tau ^{*}\) respectively.
We assume that \(q(\theta )=(1, u, w)^{T} e^{i\rho _{0}\tau ^{*}\theta }\) and \(q^{*}(s)\) are the eigenvectors of A(0) and \(A^{*}\) corresponding to \(i\rho _{0}\tau ^{*}\) and \(-i\rho _{0}\tau ^{*}\), respectively. Then, we have \(A(0)q(\theta )=i\rho _{0}\tau ^{*}q(\theta )\). By the definition of A(0) and from (35) and (36), it follows that
Then, we can get \(q(0)=(1, u, w)^{T}\),
where
Similarly, we can obtain
We choose D in such a way that \(\langle q^{*}(s), q(\theta ) \rangle =1\), \(\langle q^{*}(s), \overline{q}(\theta ) \rangle =0\).
Hence
Thus, we can choose D as \(D=\frac{1}{1+\overline{u}u^{*}+\overline{w}w^{*}+ \tau ^{*} \overline{w} \alpha (u^{*}P_*+ w^{*}I_*) e^{i\rho _{0}\tau ^{*}}}\).
To describe the center manifold \(\mathbf {C}_{0} \) at \(\mu =0 \), we compute the coordinates by using the same notations and procedures as proposed by Hassard et al. [36].
Let \( z_{t}\) be the solution of of Eq. (32) when \(\mu =0\).
Define
On the center manifold \(\mathbf {C}_{0} \), we have
where
\(\text {z}\) and \(\overline{\text {z}}\) are local coordinates for center manifold \(\mathbf {C}_{0}\) in the direction of \(q^{*}\) and \(\overline{q}^{*}\). Here W is real when \(z_{t}\) is real. Now, we consider only real solutions. For solution \(z_{t} \in \mathbf {C}_{0}\) of Eq. (32), since \(\mu =0\), we can obtain
we rewrite this equation as \(\dot{\text {z}}=i\rho _{0}\tau ^{*}\text {z}+g(\text {z}, \overline{\text {z}})\) with
Then, from Eq. (40), we have
Thus, from Eq. (41), we can get
Comparing with the coefficients with (41), we can obtain
To calculate the value of \(g_{21}\), we need to compute the values of \(W_{20}(\theta )\) and \(W_{11}(\theta )\). From Eqs. (37) and (40), we have
where
Expanding the above series and comparing the corresponding coefficients, we obtain
From Eq. (45), we know that for \(\theta \in [-1, 0),\)
Comparing the coefficients with (46) gives that
and
Since \(q(\theta )=(1, u, w)^{T} e^{i\rho _{0}\tau ^{*}\theta }\), hence
where \(E_{1}=(E_{1}^{(1)}, E_{1}^{(2)}, E_{1}^{(3)}) \in \mathbf {R}^{3}\) is a constant vector.
Similarly, from Eqs. (47) and (50), we get
where \(E_{2}=(E_{2}^{(1)}, E_{2}^{(2)}, E_{2}^{(3)}) \in \mathbf {R}^{3}\) is a constant vector.
In what follows, we shall seek appropriate \(E_{1}\) and \(E_{2}\) in (51) and (52) respectively. From the definition of A and (47), we obtain
and
where \(\eta (\theta )=\eta (0, \theta )\). From (47), we have
and
Noting that
and
and putting (51) and (55) into (53), we get
which implies that
It follows that
where
Similarly putting (52) and (56) into (54), we have
which implies that
and hence,
where
From the above analysis, we can compute the following quantities:
The above three quantities \(\mu _2\), \(\beta _2\) and \(\tau _2\) will determine the direction, stability and the periods of the bifurcating periodic solutions.
Rights and permissions
About this article

Cite this article
Biswas, S., Saifuddin, M., Sasmal, S.K. et al. A delayed prey–predator system with prey subject to the strong Allee effect and disease. Nonlinear Dyn 84, 1569–1594 (2016). https://doi.org/10.1007/s11071-015-2589-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11071-015-2589-9