
Abstract
Context
The relationship between disturbances and diversity remains uncertain, especially in forest landscapes where large spatial extents need to be considered, dynamics are slow, and disturbance interactions are common.
Objectives
To analyse the individual and combined effects of ungulate browsing and wind disturbance on tree species diversity at the landscape scale.
Methods
We used a dynamic forest landscape model to simulate the effects of browsing and wind disturbances (i.e., frequency and windthrow size) on tree species diversity in four mountain landscapes in Central Europe. Using boosted regression trees, we analysed the relative importance of each disturbance type for diversity at different layers (i.e., regeneration versus overstorey, and all tree sizes), the shapes of the diversity–disturbance relationships and the combined effect of wind and browsing disturbances on diversity.
Results
Across all landscapes, browsing and windthrow were equally important for tree species diversity when considering all forest layers, but no consistent patterns could be observed for the regeneration and overstorey layer. The shape of the disturbance–diversity relationships differed between disturbance types. More frequent and severe windthrow events typically increased diversity almost linearly, while browsing showed a non-linear response with the highest diversity at intermediate browsing pressure. However, these relationships were not consistent across the four landscapes.
Conclusions
Tree species diversity can be influenced by both browsing and windthrow disturbances. Forested landscapes are likely to experience multiple disturbances, and their relative influence on diversity needs to consider their different spatial and temporal scales.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Natural disturbances strongly influence forest structure and dynamics (Turner 2010), and may also increase species diversity (e.g., White 1979; Sousa 1984). Species diversity is an important metric of landscapes, as increasing diversity increases the stability of ecosystems and promotes ecosystem functioning (Isbell et al. 2011; Brose and Hillebrand 2016). Understanding the drivers of species diversity is therefore fundamental for understanding ecosystem functioning. Disturbance as a potential driver of species diversity has a long history, however the shape of this diversity–disturbance relationship varies widely (Mackey and Currie 2001; Svensson et al. 2012). Diversity may peak at intermediate disturbance levels (e.g., Martinsen et al. 1990), or show positive or negative relationships to increasing diversity (e.g., Abensperg-Traun et al. 1996). In some cases, diversity is even insensitive to disturbances (e.g., Jullien and Thiollay 1996). These conflicting results may arise from differences in environmental conditions among landscapes (e.g., topography; White and Jentsch 2001), differences in species composition (Silva Pedro et al. 2016), or varying frequencies or severities (i.e., amount of damage) of disturbances (e.g., Miller et al. 2011). Furthermore, communities are often shaped by multiple disturbances that may interact in non-linear and non-additive ways (Paine et al. 1998), making the diversity–disturbance relationship even more complex. Especially in forests, the study of the diversity–disturbance relationship is complicated by the large spatial extents that need to be considered, the slow pace of forest dynamics, and the frequent exposure to multiple disturbances.
Wind is one of the most important disturbance agents in Central European forests, being responsible for half of the total wood damage between 1950 and 2000 (Schelhaas et al. 2003). The severity of wind damage is influenced by a variety of factors including topography, management history, species composition and stand structure (Hanewinkel et al. 2015). At the landscape scale (i.e., 100 s of hectares), disturbances create patches of different shapes and sizes (Everham and Brokaw 1996). These windthrow areas were found to be pivotal for maintaining or promoting tree species diversity in temperate mesic forests (Canham and Loucks 1984) and Central Amazon rain forests (Marra et al. 2014). However, the long-term impact of wind events on species composition and diversity is complex and depends highly on the frequency, spatial extent and severity of wind events (Miller et al. 2011; Xi and Peet 2011). For example, Phillips and Shure (1990) observed a positive effect of windthrow size on diversity in temperate forests however, Peterson et al. (2013) found no significant effect. Alternatively, it was intermediate wind storm frequencies that resulted in the highest tree species diversity in beech forests in Japan (Hiura 1995). Comparative landscape-scale studies on the impact of windthrow regimes on tree species diversity remain scarce, and even rarer are those that disentangle the impacts of wind frequency versus severity.
Browsing by ungulates is another common disturbance in European forests, as deer populations have increased in many regions in recent decades (Côté et al. 2004). Heavy browsing may results in a species shift as the abundance of certain shrub, herb and tree species decrease and less palatable species increase (Horsley et al. 2003). This often can lead to a reduction in understorey diversity (Nuttle et al. 2014). By changing tree recruitment patterns, browsing can also modify overstorey composition in the long term (Bradshaw and Waller 2016). In northern Wisconsin, white-tailed deer significantly reduced sapling recruitment for five out of ten examined taxa (Bradshaw and Waller 2016), and in Germany, deer were responsible for a 52–67% decrease in canopy tree species diversity at the regional scale (Schulze et al. 2014).
When several disturbances co-occur, the relationship between disturbances and diversity can become complicated. For instance, wind can increase species diversity when occurring in isolation (e.g., Hiura 1995), but in combination with ungulate browsing, the net result on species diversity can be negative (Nuttle et al. 2013). In an extreme example, forests in coastal New England transitioned from a diverse mixed-species systems to Fagus grandifolia-dominated forests as a consequence of harvesting followed by intensive browsing and a severe hurricane (Busby et al. 2008). The outcome of multiple disturbances can be difficult to predict due to the multitude of factors that need to be considered, including forest state, species composition, the order of the occurrence of multiple disturbances, and species-specific vulnerability to disturbances. For instance, Picea abies is highly susceptible to windthrow but less sensitive to browsing, whereas Abies alba is particularly sensitive to browsing but not to wind (Gill 1992; Gardiner et al. 2013; Tinner et al. 2013). Even though both wind and browsing are becoming increasingly important (Didion et al. 2011; Seidl et al. 2014), little information is available about their combined effect on tree species diversity at the landscape scale.
Observational and experimental studies are limited in their ability to address the impact of multiple disturbances on forests, especially at the landscape scale, due to the limited natural occurrence of wind events and the lack of data over long time periods and large areas. Dynamic forest landscape models are therefore particularly suited to address this topic as they can simulate large-scale and long-term effects of multiple disturbances on tree species composition, structure and dynamics (e.g., Scheller and Mladenoff 2005; Henne et al. 2013). By using a model, confounding factors such as site history, environmental gradients or initial species diversity can be controlled (Morin et al. 2011). Models further allow us to explore changes in species diversity while systematically varying disturbance frequency, severity and size (e.g., Papaik and Canham 2006; Thom et al. 2017). In previous modelling studies, Papaik et al. (2005) found that beech showed a higher vulnerability to windthrow if infected by beech bark disease, ultimately causing a shift towards more shade-intolerant species. Another modelling study reported that the combination of bark beetles, wind and climate change drove Picea abies to local extinction (Temperli et al. 2013). However, there remains a large potential for using dynamic vegetation models to address uncertainties about the exposure to multiple disturbances on tree species diversity.
Here, we used a dynamic forest landscape model to disentangle the impact of two distinctive disturbance agents (i.e., wind disturbance and ungulate browsing) on tree species diversity at the landscape scale. To look for generalities in these patterns, we used four Central European landscapes that differ in species composition and overall species diversity. Specifically, we addressed the following research questions: (1) What is the relative contribution of browsing and wind disturbance for shaping tree species diversity in European forest landscapes? (2) Does the shape of the diversity–disturbance relationship vary across different landscapes? (3) What is the combined effect of browsing and wind on tree species diversity?
Methods
Dynamic forest landscape model LandClim
We used the forest landscape model LandClim (v1.8), a process-based model designed to simulate forest dynamics in a spatially explicit manner under changing climate, disturbances and land-use over long time periods and at large spatial scales (Schumacher et al. 2004). Landscapes are represented as a raster of 25 × 25 m grid cells. Within each cell, a simplified forest gap model simulates tree establishment, growth and mortality in response to abiotic factors and biotic interactions at a yearly time step (Schumacher et al. 2004). LandClim uses a cohort approach, i.e., trees of the same age and species are simulated as one representative individual. Management and spatial interactions between grid cells, such as seed dispersal and disturbances, are simulated at a decadal time step (Schumacher et al. 2006).
LandClim has successfully been used to simulate species composition and forest dynamics in the European Alps (Schumacher et al. 2004; Elkin et al. 2013), in Mediterranean ecosystems (Henne et al. 2015) and in the Rocky Mountains (Schumacher et al. 2006). The model has also been used to simulate the combined impacts of wind, bark beetle attacks and climate change on spruce-dominated forests (Temperli et al. 2013), fire and browsing on vegetation dynamics in the Mediterranean region (Henne et al. 2013), and how wildfire, wind and harvest may impact future forest development in the European Alps (Schumacher and Bugmann 2006). A detailed description of the model can be found in Schumacher (2004). Previous implementations of LandClim simulated density-dependent mortality based on a user-defined maximum biomass parameter. To improve the simulation of processes responsible for species diversity, we have removed this parameter and replaced it with a more mechanistic representation of light competition (see Online Appendix S1 for more details). A brief overview of how the model simulates disturbance processes relevant for this study is provided below.
Establishment and ungulate browsing
In LandClim, trees can establish if species-specific requirements are met (e.g., light and water availability, minimum growing degree-days). Environmental conditions are tracked annually to calculate the number of favourable years that occurred within each decade, which is used to determine the number of saplings that will establish at the end of the decade. Browsing can additionally decrease the number of saplings depending on species-specific browsing tolerances and browsing pressure. The browsing pressure in a landscape is defined by the user and is assumed to be constant over time and space. This assumption is not an accurate representation of reality (i.e., browsing is indeed variable in time and space; Adler et al. 2001), but we consider this simplification appropriate for our purposes (i.e., analysing how diversity responds to differences in browsing intensities at the landscape level).
Wind disturbances
Wind disturbances are simulated as stochastic events based on user-defined distributions of wind frequency and windthrow size. A wind event starts at a random location and spreads to adjacent grid cells until the windthrow size, which is randomly drawn from the size distribution, is reached. In previous LandClim versions (< v1.8), the major factor affecting windthrow risk and magnitude was tree diameter at breast height (DBH) only while the severity within a cell was uncorrelated to the size of the windthrow event. In the current version (v1.8), we revised the wind module to include species-specific wind resistance. Additionally, the method for determining the occurrence of wind events was improved. The major changes of the new submodel are summarized below (see Online Appendix S2 for additional details and model evaluation).
Occurrence of wind events
The frequency of wind events is determined by two user-defined parameters: the mean number of wind events per hectare and decade (mw), and the probability that these wind events occur (Pw). To determine the actual number of wind events (Xw) occurring in a specific decade per hectare, a random number from a negative binomial distribution (NB) is drawn:
.
where rw is a measure of dispersion based on the probability (Pw) and the mean number of wind events (mw) defined as:
Xw (the actual number of wind events per hectare within one decade) is then scaled up to the entire landscape. The negative binomial distribution was chosen as it can be parameterized to represent both frequent and infrequent wind regimes.
Species-specific resistance to wind
We concentrated on the key factors determining susceptibility to wind damage, i.e., tree species, height or DBH (cf. Hanewinkel et al. 2011; Albrecht et al. 2012; Gardiner et al. 2013). Since height is an allometric function of DBH in LandClim, we selected DBH. Species-specific susceptibility to wind within each grid cell was calculated using an equation developed by Canham et al. (2001). Storm severity (S) is used to calculate the log odds ratio for wind mortality probability for each species:
where Pcoh is the probability of windthrow for cohort coh within the cell, DBHcoh is diameter at breast height of the cohort, and a, b and c are species-specific parameters that relate to tolerance to wind. Storm severity (S) ranges from 0 to 1, with 1 representing the most severe wind storm (where 100% of the biomass of the most vulnerable species is removed). Within each cohort, the number of stems killed by a wind event is determined by drawing a random number from a binomial distribution using the species-specific mortality probability (Eq. 3) and the stem number of the cohort. We do not consider the effects of partial crown damage from wind storms. For assigning the parameter values a, b and c (Eq. 3), tree species were grouped into three windthrow tolerance classes (i.e., resistant, intermediate and vulnerable; Table S2 in Online Appendix S2; Gardiner et al. 2013). The parameter values were taken from Canham et al. (2001) based on the most resistant species (Acer saccharum Marsh.), an intermediate (Acer rubrum L.) and the most vulnerable species (Picea rubens Sarg.). Although the Canham et al. (2001) study was based in North America, similar wind tolerance classifications are available for the most dominant Central European tree species. For some species, information about their vulnerability class was not available. These species were mostly small, Mediterranean tree and shrub species that are very rare in the simulated landscapes, and thus were classified as resistant to wind.
Peterson et al. (2013) showed that the severity of wind events within windthrow gaps, as measured by fallen basal area, is closely related to gap size. We thus used the logarithmic relationship from this study to calculate storm severity (S) as a function of the disturbed area (D), assuming that for a single wind event the severity index is the same within the entire windthrow area:
Case study landscapes
We used four case study landscapes ranging in size from 1200 to 5300 ha, representing a wide range of forest types and climatic conditions (cf. Fig. 1 for the location, Fig. 2 for the potential natural vegetation (PNV) of each study landscape, and Online Appendix S3 for a description of the landscapes). Moreover, they range from relatively species-rich forests at low elevations (i.e., Origlio) up to high-elevation forests with only few species (i.e., Dischma). LandClim has already been used to successfully simulate PNV in the Feldberg landscape in the Black Forest region (Thrippleton et al. 2016), in Switzerland in the Dischma valley in the Canton of Grisons (Schumacher et al. 2004) and in a part of the Saas valley and the surroundings of the city of Visp in the main valley of the Valais (Elkin et al. 2013). In the landscape around Lake Origlio, LandClim simulates PNV dominated by Fagus sylvatica L., Abies alba Mill. and an admixture of Quercus ilex L. and Ilex aquifolium L. in the understorey. The current landscape is dominated by Castanea sativa Mill. with an admixture of Quercus petraea (Matt.) Liebl., Q. pubescens Willd., Alnus glutinosa (L.) Gaertn., Fraxinus excelsior L., Fagus sylvatica and Tilia cordata Mill. The disappearance of Abies alba and the decline in Tilia cordata and Fagus sylvatica were likely due to fire (Tinner et al. 1999) and in the case of Abies alba also due to increased browsing pressure (Tinner et al. 2013). Hereafter, the study landscapes are referred to as Feldberg, Dischma, Valais and Origlio.

Location of the four case study landscapes (i.e., Dischma, Feldberg, Origlio and Valais)

Simulated potential natural vegetation for the four landscapes (a–d) along an elevational gradient based on biomass after 1500 simulation years without any disturbances, land use history or climate change
These landscapes were selected not only for their diverse and representative species pools, but also because browsing and windthrow events are prominent disturbances in these areas. For instance, large windthrow events in the Black Forest region occur every 10 to 15 years (Hanewinkel et al. 2008), whereas infrequent, large storms have had large impacts in the canton of Grisons, where hurricane Vivian led to a massive amount of wood damage in 1990 (Willi et al. 2014). High ungulate browsing pressure in Switzerland and Germany has strongly suppressed the regeneration of many highly palatable species such as Abies alba, Acer pseudoplatanus L., Quercus spp., Sorbus torminalis (L.) Crantz, S. aucuparia L. or Taxus baccata L. (ForstBW 2013; Rigling et al. 2015).
Simulation settings
To quantify the sensitivity of tree species diversity to browsing and wind disturbances, we ran simulations for 1500 years under a constant climate, starting from bare ground for all four landscapes, with different combinations of three disturbance parameters: windthrow frequency (i.e., mean number of wind events), mean windthrow size (i.e., correlates with the severity of a storm event, Eq. 4), and browsing pressure (see Table 1 for parameter values). Minimum and maximum windthrow size were set to 60% and 1400% of mean windthrow size, creating a log-normal size distribution that is highly skewed towards a higher frequency of small windthrow sizes but with the occasional occurrence of large gaps (e.g., Quine and Bell 1998). The windthrow probability (Pw) was fixed to 0.9 creating a relatively narrow almost normal distribution for all scenarios (see Online Appendix S2 for a justification of the selected wind disturbance ranges). Browsing pressure may differ greatly among landscapes (Fischer et al. 2013), and might even lead to complete establishment failure of certain tree species (Senn and Suter 2003). Thus, a very wide range of browsing pressures (0–0.9) was explored. All simulations included 31 potential tree species for Feldberg, Dischma and Valais, and 38 potential tree species for Origlio, assuming global seed dispersal and no management interventions. Using historical climate data from each landscape as input (from ~ 1950 to 2000), climate records were randomly sampled with replacement to generate 1500-year climate sequences for each simulation run.
Combinations of disturbance parameter values were generated using Latin Hypercube sampling (LHS), which has been shown to be an efficient alternative to a factorial design, and the results are considered robust in spite of much smaller sample sizes (Saltelli 2008). To ensure coverage of the entire parameter space, 1000 parameter combinations were generated using the lhs function in the tgp package v2.4-14 in R (Gramacy and Taddy 2010). For all landscapes, the same 1000 parameter combinations were used.
Output measures
Tree diversity
We used Shannon’s diversity index H′ (Shannon and Weaver 1949) based on the proportion of stem numbers of each species (pi) across the entire landscape. We chose to use stem numbers instead of biomass to account for subdominant species that have a smaller contribution to the total forest biomass:
Disturbances can have a different impact on different layers in the forest (e.g., Tremblay et al. 2007; Gardiner et al. 2013). Wind tends to act as a top-down disturbance mostly affecting the larger trees, whereas browsing is a bottom-up disturbance that affects forests by differential filtering of regeneration. Thus, we calculated Shannon’s diversity index for: (1) all tree sizes (i.e., all individuals), (2) the regeneration layer (i.e., stems < 10 cm DBH), and (3) the overstorey (i.e., stems > 40 cm DBH; Brändli and Speich 2007).
Statistical analysis
To analyse the effect of browsing and wind disturbance on tree species diversity at the landscape scale, we calculated Shannon’s diversity index of the entire landscape after 1500 years of simulation, i.e., when the simulated landscapes had reached a dynamic equilibrium. The area above natural treeline (i.e., tree height < 3 m; Harsch et al. 2009) was not included in the analysis for the Dischma valley. As we focused on the impact of disturbances on tree species diversity changes, we evaluated the difference in diversity between a baseline simulation and the disturbance scenarios. As a baseline, we simulated forest dynamics for all study landscapes in the absence of disturbances, with 10 replicates each, and calculated average diversity in the year 1500 over all replicates for the entire landscape.
To quantify the relative contribution of browsing, wind frequency and windthrow size on Shannon’s diversity index, we used boosted regression trees (BRT) in R v3.4.1 (R Development Core Team 2016). BRT are machine-learning algorithms that combine regression tree models and boosting algorithms (Elith et al. 2008), particularly useful for fitting complex and non-linear responses. We used the gbm package v2.1.3 (Ridgeway 2017) and the dismo package v1.1-4 (Hijmans et al. 2017) following the recommendations by Elith et al. (2008). BRT models were fitted using a tree complexity of 2 and a bag fraction (i.e., how much of the data is used for each iteration) of 0.5. We varied the learning rate to reach at least 1000 trees, as suggested by Leathwick et al. (2006). Browsing pressure, mean windthrow size and wind frequency (i.e., normalized to 1 ha) were used as explanatory variables. Due to the highly stochastic nature of the wind regime, we tracked the number of wind events that were simulated in the previous 100 years (i.e., between the simulation years 1400 to 1500) as well as mean windthrow size over this 100-year period. This was done because two simulations might have the exact same input values, but could still differ in the number and size of wind events simulated. We selected the last 100 years as the number and size of the wind events during this time period, would have had the largest impact on current tree diversity. Thus, we were able to use the actual wind disturbance regime per simulation for our analyses. To enable comparisons across the four landscapes, the number of wind events was always normalized to one hectare.
For assessing the influence of just wind frequency and windthrow size on tree species diversity, we used only those scenarios that had a low browsing pressure (< 0.2), as the influence of browsing at this level relatively small (Didion et al. 2011). Relying on only those simulations with a browsing pressure of exactly zero would have led to a too small sample size from the LHS. For each study landscape, a local polynomial regression smoother (LOESS) was fitted using mean windthrow size and normalized wind frequency as predictors. As mean windthrow size and the number of wind events were found to have similar effects on tree species diversity (see “Results”), we combined these two parameters into a single metric of the wind rotation period (RP), which is defined as the time needed to disturb an area equal to the entire landscape given a certain average wind frequency and windthrow size (e.g., Turner et al. 2001); it is calculated as
where t is the time of observation (i.e., in our case 100 years) and p is the proportion of the landscape disturbed. Then, we used browsing pressure and RP as predictors in the LOESS function to examine the influence of wind disturbance and browsing on tree species diversity for each landscape.
Results
Relative contribution of browsing and wind disturbance for tree diversity
The relative contribution of browsing pressure, wind frequency and windthrow size on tree species diversity varied among landscapes and forest layers (Fig. 3). In general, tree diversity was more sensitive to wind in Feldberg and Origlio, whereas browsing pressure was more important in the Valais and Dischma. For all tree sizes, the relative contribution of browsing ranged from 32% in Origlio up to 56% in the Valais, and the total effect of wind (i.e., both frequency and size) ranged between 44 and 68%. In comparison to all tree sizes, diversity in the regeneration layer was more sensitive to browsing in Origlio and Dischma (the relative contribution of browsing to the change in diversity in the regeneration layer increased by ~ 30% compared to all tree sizes; Fig. 3b, d). In Feldberg and Valais, the regeneration layer was slightly less sensitive to browsing, compared to all tree sizes (decreased by ~ 10%; Fig. 3a, c). Diversity in the overstory layer was more sensitive to wind disturbances in Feldberg (63%) and Origlio (74%). These patterns are in contrast to those found in the Valais and Dischma, where overstory diversity was much less sensitive to wind (relative contribution of wind was only 28% and 5%, respectively).

Relative contribution of each disturbance to the total change in tree species diversity for all case study landscapes (a–d) based on boosted regression trees. Diversity changes are expressed as the difference in Shannon’s diversity index, comparing the absolute difference between a baseline scenario without disturbances to disturbance scenarios. Diversity was calculated after 1500 years of simulation for all tree sizes (i.e., all individuals), the regeneration layer (i.e., stems < 10 cm DBH) and the overstorey layer (i.e., stems > 40 cm DBH)
Shape of the diversity–disturbance relationship
The predicted response of tree species diversity to increasing browsing pressure was nonlinear (Fig. 4a–c; Online Appendix S4 and S5 for more details), and typically the highest increase in diversity occurred at intermediate browsing pressure (Fig. 4a, c). For the overstorey layer, the relationship between browsing pressure and diversity differed between the four landscapes (Fig. 4b). In the Dischma valley, diversity had a negative linear relationship (i.e., as diversity decreased as browsing pressure increased). In the Valais, there was a singular diversity peak at intermediate browsing pressure, while Feldberg and Origlio had more of a bimodal relationship. The first peak in diversity occurred at a browsing pressure of 0.3, then fell and rose again for values > 0.6.

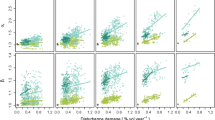
Partial dependence plots for windthrow and browsing disturbance for all tree sizes (i.e., all individuals), the overstorey layer (i.e., DBH > 40 cm) and the regeneration layer (i.e., stems < 10 cm DBH) for the four landscapes. Mean windthrow size and windthrow frequency were calculated for each simulation, based on the wind events that were simulated in last 100 years (see “Methods” for additional details). The difference in Shannon’s diversity index between disturbance scenarios and the baseline (i.e., a scenario without disturbances) was used as the response variable. Values on the y-axis were plotted by fixing all other predictors at their mean value. Note that the difference in species diversity does not reveal how diverse the landscape is in general. For instance, even though Dischma is the species-poorest system, the diversity change can be strong
Across the four landscapes and in all forest layers, there was a positive linear relationship between species diversity and increasing wind frequency and windthrow size (Fig. 4d–i; Online Appendix S5 for more details). Moreover, the highest increase in the diversity index typically occurred under a wind regime with both high frequency and large windthrow size (Fig. 5). The increase in diversity was highest in the low elevation landscape, Feldberg (Fig. 5a–c). The smallest increase in diversity due to wind disturbances was found in the highest elevation landscape, Dischma (Fig. 5j–l). The sensitivity of diversity to wind frequency and size was not consistent between canopy layers. For instance, the overstorey layer in Origlio had a greater diversity increase due to wind than the regeneration layer (Fig. 5e, f), while it was the regeneration layer in Valais that had a greater increase in diversity compared to the overstory (Fig. 5h, i).

Change in species diversity as a function of wind frequency (i.e., number of wind events in the previous 100 years, normalized to 1 ha) and mean windthrow size (ha) for all tree sizes (i.e., all individuals), the overstorey layer (i.e., stems > 40 cm DBH) and the regeneration layer (i.e., stems < 10 cm DBH) for all four landscapes. The surface plots display the absolute difference in Shannon’s diversity index (between disturbance scenarios and a baseline scenario without disturbances) after 1500 years of simulation for a gradient of wind frequency and windthrow size with minor browsing pressure (< 0.2). A local polynomial trend function (LOESS) was fitted to approximate the diversity response for the whole parameter space
Interactions between browsing and wind disturbance
Both disturbance agents influenced diversity at the landscape scale, although the sensitivity of diversity to these two disturbances varied between landscapes (Fig. 6). Once again, the greatest increase in the Shannon’s diversity index was found in the lowest elevation landscape (i.e., Feldberg). In the highest elevation landscape, Dischma, diversity was either unchanged or decreased under various disturbance intensities. There was also evidence that the interaction between the two disturbances led to different diversity responses. For instance, the interaction between increasing browsing pressure and increasing wind disturbances (i.e., shorter rotation periods) had a strong positive effect on tree diversity in the Feldberg landscape (Fig. 6a–c) while the interaction between the two disturbances reduced diversity in the Dischma landscape (Fig. 6j–l). In addition, the peak in diversity at intermediate browsing pressure could shift depending on the severity of the wind regime. In Feldberg and Origlio, the peak in diversity occurred at lower browsing levels under shorter wind rotation periods (Fig. 6c, d). In Dischma, the peak in diversity occurred at higher browsing levels under more intense wind regimes (Fig. 6j). As wind rotation periods became longer, this hump-shape diversity response could change to a linear response (Fig. 6a, b, e).

Change in species diversity relative to a no-disturbance scenario as a function of wind rotation period (i.e., mean time a disturbance regime needs to impact an area equal to the size of the entire study landscape; in log-scale) and browsing pressure for the four case study landscapes. Absolute differences in Shannon’s diversity index are shown for a gradient of rotation period and browsing pressure for all tree sizes (i.e., all individuals), the overstorey layer (i.e., stems > 40 cm DBH) and the regeneration layer (i.e., stems < 10 cm DBH). A local polynomial trend function (LOESS) was fitted to approximate the diversity response for the entire parameter space. Note that shorter rotation periods feature a more intense wind disturbance regime
Discussion
Tree species diversity at the landscape scale can be influenced by multiple disturbance agents, but it is quite challenging to quantify their relative importance in empirical studies. Describing the shape of the diversity–disturbance relationship for forests is equally as challenging, as it is virtually impossible to capture all disturbance intensity levels within the same landscape. Our modelling approach allowed for a systematic analysis of multiple disturbance regimes in different landscapes with different species pools. Although our approach exhaustively covered multiple parameter combinations, several assumptions were necessary. First, each simulation assumed a constant browsing pressure for all parts of the landscape (although browsing pressure did vary between simulations). In reality, certain parts of the landscape will experience higher or lower browsing pressure, spatially as well as temporally (Adler et al. 2001; Reimoser et al. 2009). However, the purpose of this study was not to determine the influence of browsing on species diversity at any one particular location, as this question is better suited for empirical studies (e.g., Royo et al. 2017; Ramirez et al. 2019). Rather, it was to elucidate the more general response of diversity to browsing at all levels. A similar simplifying assumption was made for wind disturbances, which we know in reality to be influenced by topography, aspect, and soil conditions (Everham and Brokaw 1996), which are not considered in our simulations. However, even though the intensity of wind and browsing disturbances were constant within any one particular simulation, they varied between simulations. This allowed us to quantify the relative contribution of wind and browsing on species diversity at the landscape scale, describe the shape of the diversity–disturbance relationship, and understand how these two disturbances interact.
Relative contribution of browsing and wind disturbance for tree diversity
Tree species diversity was influenced equally by both browsing and windthrow across all four landscapes in our simulations when considering all tree sizes (Fig. 3). Thus, both bottom-up (browsing) and top-down (windthrow) disturbances are likely to be important for tree species diversity in European landscapes. Interestingly, the relative influence of wind and browsing differed between landscapes when considering the regeneration and the overstorey layer separately.
One might have presumed that wind would have a higher relative importance for overstorey tree species diversity as strong wind events predominantly kill larger trees, while browsing would have a stronger impact on diversity in the regeneration layer. However, Origlio was the only landscape that showed this response (Fig. 3b). In Valais, the impact of the two disturbances was exactly opposite to the presumed pattern (i.e., browsing was more important for overstorey diversity and wind was more important for regeneration diversity). In Feldberg, wind disturbances had a larger relative impact on tree species diversity in both layers, while browsing was more important for tree species diversity in Dischma, independent of the layer. The reasons for these landscape-specific responses are discussed below.
The four landscapes had different species compositions and structure due to differences in topography and climatic conditions (Fig. 2). These differences translated into different compositions of disturbance vulnerabilities and thus influenced the importance of each disturbance in the study landscapes. For instance, diversity in all forest layers were especially sensitive to increasing browsing pressure in the Dischma valley (Fig. 3d). In this species-poor landscape, browsing caused Abies alba to disappear, and there were no other browsing-tolerant species that could have ‘filled in’ for the species loss (Fig. 4b, c). As the Shannon’s diversity index considers both species abundance and species richness (Shannon and Weaver 1949), the browsing-induced removal of a species had a large influence on the index in this species-poor landscape, and thus it resulted in a high relative importance of browsing in all layers. In Feldberg and Origlio, it was wind disturbances that had the highest relative importance for tree species diversity in the overstorey (Fig. 3a, b). Both of these landscapes are located at lower elevations where tree growth rates are higher. The simulated forests were composed of more trees in larger DBH size classes, which are more susceptible to wind damage. Wind storm damage allowed less competitive species to reach the canopy, and thus increased overstorey diversity. Hence, our findings suggest that wind and browsing are indeed important disturbances for shaping tree species diversity at the landscape scale, but their relative contribution depends strongly on the regional species pool, i.e., species composition (via functional traits) and forest structure, particularly tree size distributions as mediated by growing conditions.
Our analysis was designed to evaluate and compare the relative importance of wind and browsing disturbances for tree species diversity among different landscapes. However, we recognize that the relative importance of wind and browsing was influenced by the disturbance ranges we selected for our study. Even though we restricted the parameters of our disturbance regimes to ranges that are typically observed in temperature forest ecosystems (e.g., Quine and Bell 1998; Senn and Suter 2003; Klopcic et al. 2009), such estimates can also be uncertain. For instance, using more severe wind regimes would most likely have led to a larger importance of wind in shaping tree species diversity. In addition, the range of different disturbance regimes simulated do not represent the actual disturbance regime of each specific landscape. Our results should therefore not be interpreted as a clear indication of the importance of the disturbances in the different landscapes, but as an example of how the importance of disturbances can differ between landscapes.
Shape of the diversity–disturbance relationship
The shape of the diversity–disturbance relationship differed between windthrow and browsing, but was mostly consistent across the four landscapes. While diversity typically peaked at intermediate browsing pressures, it linearly increased with increasing windthrow size and frequency (Figs. 4, 5).
The linear increase in diversity with intensifying wind disturbances was, at first glance, surprising as we expected diversity to decrease under very strong wind regimes. In our simulations, forests without wind disturbances tended to form species-poor stands dominated by a few late-successional species. This was especially true in Feldberg (Fig. 5a–c). Windthrow events disrupted this forest structure by preferentially removing larger trees and allowing for the coexistence of shade-tolerant and -intolerant species. As wind is a spatially explicit disturbance in the model (and in reality), some patches remained undisturbed, leading to a landscape featuring a heterogeneous mosaic of patches with different successional states, and thus higher diversity at the landscape scale. Hence, our results suggest that intensifying wind regimes, within the range common for Central Europe, are likely to enhance tree species diversity by creating more heterogeneous landscapes.
This result is generally well supported by empirical research. For example an increase of diversity due to higher landscape patchiness was reported for natural forest fires in the Yellowstone National Park (Romme 1982). However, empirical studies that separately consider the effects of frequency and windthrow size on tree species diversity are ambiguous. While some studies found a positive effect of increasing gap size on species diversity due to an increase in early-successional species within the gaps (e.g., Phillips and Shure 1990), others found no relationship between windthrow size and diversity (Peterson et al. 2013; Cowden et al. 2014), or the highest diversity under an intermediate disturbance frequency (Hiura 1995). However, these empirical studies often evaluated single wind events or focused only on the disturbed parts of the landscape (i.e., without taking into account the surrounding undisturbed forest). Our landscapes (quite realistically) never experienced a wind event that disturbed the entire area, whereas forests in windthrow plots can be damaged completely, thus this lack of congruence between results at different spatial scales is not surprising.
Including more frequent or severe wind events in our simulations may have led to a negative effect on diversity under these conditions. However, such wind regimes are arguably beyond what would be realistic for forests of Central Europe (e.g., Schreiner et al. 1996; Klopcic et al. 2009), and thus they were not included. Even though winter storms occur relatively frequently in Central Europe, their destructive power is simply not comparable to the hurricanes and tornados common in North America (Fischer et al. 2013). Therefore, a severe wind regime for Central Europe may not be classified as being severe in other parts of the world. In addition, we evaluated diversity of the entire landscape. Thus, simulating a more severe wind regime would still have created a patchy landscape that would most likely still increase diversity, thus not altering our conclusions.
While species diversity consistently showed a positive relationship with increasing wind disturbances, the highest diversity was found at intermediate levels of browsing across (almost) all landscapes and forest layers (Fig. 4a–c). Under low browsing pressure, the dominant species were often browsing-intolerant. Increasing browsing pressure had a positive impact on species diversity by gradually removing these dominant species and allowing subdominant species to establish in the landscape. As browsing pressure continued to increase, only a few highly browsing-tolerant species were able to thrive, thus reducing diversity under the highest browsing pressure. In Dischma, the shape of this relationship differed from the other landscapes. In this species-poor valley, the removal of Abies alba had a larger influence on the forest composition leading to a decrease in diversity with a higher browsing pressure across all forest layers. In contrast to wind, browsing may permanently remove species from the entire landscape, thus fundamentally altering competition between the remaining species and leading to a non-linear response of tree species diversity.
Browsing was constant over time and space in our simulations, and thus our results can be compared to empirical plot-scale studies. Cook-Patton et al. (2014) also found that browsing increased tree species diversity in the regeneration layer due to the elimination of dominant but browsing-intolerant species. However, browsing may also reduce diversity due to the local depletion of certain species (Gill and Beardall 2001). Our results also showed a subsequent diversity change over a longer period and demonstrated that the impacts on the regeneration layer cascaded into the overstorey, as corroborated by other studies (e.g., Didion et al. 2009; Bradshaw and Waller 2016). Our results also highlight the importance of the spatial scale of disturbances (i.e., in our simulations browsing was assumed to disturb every patch whereas wind throws were stochastically distributed across the landscape). These differences in spatial scales are likely to influence the shape of the diversity–disturbance relationship, and may be one reason why so many different responses have been reported for terrestrial ecosystems (e.g., Martinsen et al. 1990; Abensperg-Traun et al. 1996).
Interactions between browsing and wind disturbance
The diversity–disturbance patterns as discussed in the previous sections assumed that the other disturbance did not vary (see “Methods”). Our results showed that the shape of the diversity–disturbance relationships could change if the severity of the second disturbance was altered (Fig. 6). For example, under more intense wind disturbances (short rotation periods), the peak in diversity occurred at lower browsing pressures. Under longer wind rotation periods, the peak in diversity occurred at higher browsing levels (e.g., Fig. 6c, d, f–i) or the relationship became more linear (e.g., Fig. 6a, b, e). Plant communities are commonly exposed to multiple disturbances, yet the interaction between disturbances can be difficult to predict (Mouillot et al. 2013) and we are only beginning to understand how multiple disturbances shape ecosystems and communities (e.g., D’Amato et al. 2011).
The results from our modelling study support the hypotheses that disturbances can interact in non-linear and non-additive ways (Paine et al. 1998). More importantly, we found that the interaction between disturbances was not consistent between landscapes. Even though the same disturbances were simulated in all four landscapes in our modelling study, the effect of the disturbances was quite different (compare Feldberg and Dischma, for example). Thus, the composition of the local species communities can substantially alter the response to disturbances. Using a functional approach as suggested by Mouillot et al. (2013) would be an important next step, to generalize these diversity-disturbance relationships based on species traits. Understanding how multiple, interacting disturbances influence species diversity is integral for understanding ecosystem functioning and stability.
References
Abensperg-Traun M, Smith GT, Arnold GW, Steven DE (1996) The effects of habitat fragmentation and livestock-grazing on animal communities in remnants of gimlet Eucalyptus salubris woodland in the Western Australian wheatbelt. I. Arthropods. J Appl Ecol 33(6):1281–1301
Adler PB, Raff DA, Lauenroth WK (2001) The effect of grazing on the spatial heterogeneity of vegetation. Oecologia 128(4):465–479
Albrecht A, Hanewinkel M, Bauhus J, Kohnle U (2012) How does silviculture affect storm damage in forests of south-western Germany? Results from empirical modeling based on long-term observations. Eur J For Res 131(1):229–247
Bradshaw L, Waller DM (2016) Impacts of white-tailed deer on regional patterns of forest tree recruitment. For Ecol Manage 375:1–11
Brändli UB, Speich S (2007) Swiss NFI glossary and dictionary. Swiss Federal Research Institute WSL, Birmensdorf
Brose U, Hillebrand H (2016) Biodiversity and ecosystem functioning in dynamic landscapes. Philos Trans R Soc B 371(1694):9
Busby PE, Motzkin G, Foster DR (2008) Multiple and interacting disturbances lead to Fagus grandifolia dominance in coastal New England. J Torrey Bot Soc 135(3):346–359
Canham CD, Loucks OL (1984) Catastrophic windthrow in the presettlement forests of Wisconsin. Ecology 65(3):803–809
Canham CD, Papaik MJ, Latty EF (2001) Interspecific variation in susceptibility to windthrow as a function of tree size and storm severity for northern temperate tree species. Can J For Res 31:1–10
Cook-Patton SC, LaForgia M, Parker JD (2014) Positive interactions between herbivores and plant diversity shape forest regeneration. Proc R Soc B 281:20140261. https://doi.org/10.1098/rspb.2014.0261
Côté SD, Rooney TP, Tremblay JP, Dussault C, Waller DM (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Sci 35:113–147
Cowden MM, Hart JL, Schweitzer CJ, Dey DC (2014) Effects of intermediate-scale wind disturbance on composition, structure, and succession in Quercus stands: implications for natural disturbance-based silviculture. For Ecol Manage 330:240–251
D’Amato AW, Fraver S, Palik BJ, Bradford JB, Patty L (2011) Singular and interactive effects of blowdown, salvage logging, and wildfire in sub-boreal pine systems. For Ecol Manage 262(11):2070–2078
Didion M, Kupferschmid AD, Bugmann H (2009) Long-term effects of ungulate browsing on forest composition and structure. For Ecol Manage 258:S44–S55
Didion M, Kupferschmid AD, Wolf A, Bugmann H (2011) Ungulate herbivory modifies the effects of climate change on mountain forests. Clim Change 109(3):647–669
Elith J, Leathwick JR, Hastie T (2008) A working guide to boosted regression trees. J Anim Ecol 77(4):802–813
Elkin C, Gutiérrez AG, Leuzinger S, Manusch C, Temperli C, Rasche L, Bugmann H (2013) A 2 °C warmer world is not safe for ecosystem services in the European Alps. Glob Chang Biol 19:1827–1840
Everham EM, Brokaw NVL (1996) Forest damage and recovery from catastrophic wind. Bot Rev 62:113–185
Fischer A, Marshall P, Camp A (2013) Disturbances in deciduous temperate forest ecosystems of the northern hemisphere: their effects on both recent and future forest development. Biodivers Conserv 22:1863–1893
ForstBW (2013) Forstliches Gutachten 2013-2015 zum Rehwildabschussplan. ForstBW, Schwetzingen, p 35
Gardiner B, Schuck A, Schelhaas MJ, Orazio C, Blennow K, Nicoll B (2013) Living with storm damage to forests. European Forest Institute, Joensuu
Gill RMA (1992) A review of damage by mammals in North Temperate Forests. 1. Deer. Forestry 65(2):145–169
Gill RMA, Beardall V (2001) The impact of deer on woodlands: the effects of browsing and seed dispersal on vegetation structure and composition. Forestry 74(3):209–218
Gramacy RB, Taddy M (2010) Categorical inputs, sensitivity analysis, optimization and importance tempering with tgp Version 2, an R package for treed gaussian process models. J Stat Softw 33(6):1–48
Hanewinkel M, Albrecht A, Schmidt M (2015) Können windwurfschäden vermindert werden? Eine analyse von einflussgrössen. Schweiz Z Forstwes 166(3):118–128
Hanewinkel M, Breidenbach J, Neeff T, Kublin E (2008) Seventy-seven years of natural disturbances in a mountain forest area—the influence of storm, snow, and insect damage analysed with a long-term time series. Can J For Res 38:2249–2261
Hanewinkel M, Hummel S, Albrecht A (2011) Assessing natural hazards in forestry for risk management: a review. Eur J For Res 130:329–351
Harsch MA, Hulme PE, McGlone MS, Duncan RP (2009) Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol Lett 12(10):1040–1049
Henne PD, Elkin C, Colombaroli D, Samartin S, Bugmann H, Heiri O, Tinner W (2013) Impacts of changing climate and land use on vegetation dynamics in a Mediterranean ecosystem: insights from paleoecology and dynamic modeling. Landscape Ecol 28:819–833
Henne PD, Elkin C, Franke J, Colombaroli D, Calo C, La Mantia T, Pasta S, Conedera M, Dermody O, Tinner W (2015) Reviving extinct Mediterranean forest communities may improve ecosystem potential in a warmer future. Front Ecol Environ 13(7):356–362
Hijmans RJ, Phillips S, Leathwick JR, Elith J (2017) Dismo: species distribution modeling. R package version 1.1-4
Hiura T (1995) Gap formation and species diversity in Japanese beech forests: a test of the intermediate disturbance hypothesis on a geographic scale. Oecologia 104(3):265–271
Horsley SB, Stout SL, deCalesta DS (2003) White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol Appl 13(1):98–118
Isbell F, Calcagno V, Hector A, Connolly J, Harpole WS, Reich PB, Scherer-Lorenzen M, Schmid B, Tilman D, van Ruijven J, Weigelt A, Wilsey BJ, Zavaleta ES, Loreau M (2011) High plant diversity is needed to maintain ecosystem services. Nature 477(7363):199–202
Jullien M, Thiollay J-M (1996) Effects of rain forest disturbance and fragmentation: comparative changes of the raptor community along natural and human-made gradients in French Guiana. J Biogeogr 23(1):7–25
Klopcic M, Poljanec A, Gartner A, Boncina A (2009) Factors related to natural disturbances in mountain Norway Spruce (Picea abies) forests in the Julian. Ecoscience 16(1):48–57
Leathwick JR, Elith J, Francis MP, Hastie T, Taylor P (2006) Variation in demersal fish species richness in the oceans surrounding New Zealand: an analysis using boosted regression trees. Mar Ecol Prog Ser 321:267–281
Mackey RL, Currie DJ (2001) The diversity–disturbance relationship: is it generally strong and peaked? Ecology 82(12):3479–3492
Marra DM, Chambers JQ, Higuchi N, Trumbore SE, Ribeiro GHPM (2014) Large-scale wind disturbances promote tree diversity in a Central Amazon forest. PLoS ONE 9(11):e103711
Martinsen GD, Cushman JH, Whitham TG (1990) Impact of pocket gopher disturbance on plant–species diversity in a shortgrass prairie community. Oecologia 83(1):132–138
Miller AD, Roxburgh SH, Shea K (2011) How frequency and intensity shape diversity–disturbance relationships. Proc Natl Acad Sci 108(14):5643–5648
Morin X, Fahse L, Scherer-Lorenzen M, Bugmann H (2011) Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol Lett 14:1211–1219
Mouillot D, Graham NAJ, Villeger S, Mason NWH, Bellwood DR (2013) A functional approach reveals community responses to disturbances. Trends Ecol Evol 28(3):167–177
Nuttle T, Ristau TE, Royo AA (2014) Long-term biological legacies of herbivore density in a landscape-scale experiment: forest understoreys reflect past deer density treatments for at least 20 years. J Ecol 102(1):221–228
Nuttle T, Royo AA, Adams MB, Carson WP (2013) Historic disturbance regimes promote tree diversity only under low browsing regimes in eastern deciduous forest. Ecol Monogr 83(1):3–17
Paine RT, Tegner MJ, Johnson EA (1998) Compounded perturbations yield ecological surprises. Ecosystems 1(6):535–545
Panayotov M, Bebi P, Tsvetanov N, Alexandrov N, Laranjeiro L, Kulakowski D (2015) The disturbance regime of Norway spruce forests in Bulgaria. Can J For Res 45(9):1143–1153
Papaik MJ, Canham CD (2006) Species resistance and community response to wind disturbance regimes in northern temperate forests. J Ecol 94:1011–1026
Papaik MJ, Canham CD, Latty EF, Woods KD (2005) Effects of an introduced pathogen on resistance to natural disturbance: beech bark disease and windthrow. Can J For Res 35(8):1832–1843
Peterson CJ, Krueger LM, Royo AA, Stark S, Carson WP (2013) Disturbance size and severity covary in small and mid-size wind disturbances in Pennsylvania northern hardwoods forests. For Ecol Manage 302:273–279
Phillips DL, Shure DJ (1990) Patch-size effects on early succession in Southern Appalachian forests. Ecology 71(1):204–212
Quine CP, Bell PD (1998) Monitoring of windthrow occurrence and progression in spruce forests in Britain. Forestry 71(2):87–97
R Development Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ramirez JI, Jansen PA, den Ouden J, Goudzwaard L, Poorter L (2019) Long-term effects of wild ungulates on the structure, composition and succession of temperate forests. For Ecol Manage 432:478–488
Reimoser S, Partl E, Reimoser F, Vospernik S (2009) Roe-deer habitat suitability and predisposition of forest to browsing damage in its dependence on forest growth-model sensitivity in an alpine forest region. Ecol Model 220(18):2231–2243
Ridgeway G (2017) Generalized boosted regression models. Documentation on the R package ‘gbm’, Version 2-1-3
Rigling A, Bundesamt Schweiz, Schweiz Bundesamt für Umwelt, Eidgenössische Forschungsanstalt für Wald Schnee und Landschaft (2015) Waldbericht 2015 Zustand und Nutzung des Schweizer Waldes. BUWAL, Bern
Romme WH (1982) Fire and landscape diversity in subalpine forests of Yellowstone National Park. Ecol Monogr 52(2):199–221
Royo AA, Kramer DW, Miller KV, Nibbelink NP, Stout SL (2017) Spatio-temporal variation in foodscapes modifies deer browsing impact on vegetation. Landscape Ecol 32(12):2281–2295
Saltelli A (2008) Sensitivity analysis. Wiley, New York
Schelhaas M-J, Nabuurs G-J, Schuck A (2003) Natural disturbances in the European forests in the 19th and 20th centuries. Glob Chang Biol 9:1620–1633
Scheller RM, Mladenoff DJ (2005) A spatially interactive simulation of climate change, harvesting, wind, and tree species migration and projected changes to forest composition and biomass in northern Wisconsin, USA. Glob Chang Biol 11(2):307–321
Schreiner M, Aldinger E, Bantle P (1996) Standort und Sturmwuf 1990—dargestellt am östlichen odenwald und nordöstlichen schwarzwald. Mitt Ver Forst Standortskd Forstpflanzenztg 38:27–36
Schulze ED, Bouriaud O, Wäldchen J, Eisenhauer N, Walentowski H, Seele C, Heinze E, Pruschitzki U, Danila G, Marin G, Hessenmöller D, Bouriaud L, Teodosiu M (2014) Ungulate browsing causes species loss in deciduous forests independent of community dynamics and silvicultural management in Central and Southeastern Europe. Ann For Res 57(2):267–288
Schumacher S (2004) The role of large-scale disturbances and climate for the dynamics of forested landscapes in the European Alps. Dissertation, ETH Zurich
Schumacher S, Bugmann H (2006) The relative importance of climatic effects, wildfires and management for future forest landscape dynamics in the Swiss Alps. Glob Chang Biol 12:1435–1450
Schumacher S, Bugmann H, Mladenoff DJ (2004) Improving the formulation of tree growth and succession in a spatially explicit landscape model. Ecol Model 180:175–194
Schumacher S, Reineking B, Sibold J, Bugmann H (2006) Modeling the impact of climate and vegetation on fire regimes in mountain landscapes. Landscape Ecol 21:539–554
Seidl R, Schelhaas MJ, Rammer W, Verkerk PJ (2014) Increasing forest disturbances in Europe and their impact on carbon storage. Nat Clim Change 4(9):806–810
Senn J, Suter W (2003) Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: beliefs in search of supporting data. For Ecol Manage 181(1):151–164
Shannon CE, Weaver W (1949) The mathematical theory of communication. University of Illinois Press, Urbana
Silva Pedro M, Rammer W, Seidl R (2016) A disturbance-induced increase in tree species diversity facilitates forest productivity. Landscape Ecol 31(5):989–1004
Sousa WP (1984) The role of disturbance in natural communities. Annu Rev Ecol Syst 15:353–391
Svensson JR, Lindegarth M, Jonsson PR, Pavia H (2012) Disturbance–diversity models: what do they really predict and how are they tested? Proc R Soc B 279(1736):2163–2170
Temperli C, Bugmann H, Elkin C (2013) Cross-scale interactions among bark beetles, climate change, and wind disturbances: a landscape modeling approach. Ecol Monogr 83:383–402
Thom D, Rammer W, Dirnbock T, Muller J, Kobler J, Katzensteiner K, Helm N, Seidl R (2017) The impacts of climate change and disturbance on spatio-temporal trajectories of biodiversity in a temperate forest landscape. J Appl Ecol 54(1):28–38
Thrippleton T, Bugmann H, Kramer-Priewasser K, Snell RS (2016) Herbaceous understorey: an overlooked player in forest landscape dynamics? Ecosystems 19:1240–1254
Tinner W, Colombaroli D, Heiri O, Henne PD, Steinacher M, Untenecker J, Vescovi E, Allen JRM, Carraro G, Conedera M, Joos F, Lotter AF, Luterbacher J, Samartin S, Valsecchi V (2013) The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol Monogr 83(4):419–439
Tinner W, Hubschmid P, Wehrli M, Ammann B, Conedera M (1999) Long-term forest fire ecology and dynamics in southern Switzerland. J Ecol 87(2):273–289
Tremblay J-P, Huot J, Potvin F (2007) Density-related effects of deer browsing on the regeneration dynamics of boreal forests. J Appl Ecol 44(3):552–562
Turner MG (2010) Disturbance and landscape dynamics in a changing world. Ecology 91(10):2833–2849
Turner MG, Gardner RH, O’Neill RV (2001) Landscape ecology in theory and practice pattern and process. Springer, New York
White PS (1979) Pattern, process, and natural disturbance in vegetation. Bot Rev 45(3):229–299
White PS, Jentsch A (2001) The search for generality in studies of disturbance and ecosystem dynamics. In: Esser K, Lüttge U, Kadereit JW, Beyschlag W (eds) Progress in botany: genetics physiology systematics ecology. Springer, Berlin, pp 399–450
Willi C, Blaser L, Schulze T (2014) Gefährdungsanalyse Kanton Graubünden. Amt für Militär und Zivilschutz, Graubünden, p 127
Xi W, Peet RK (2011) The complexity of catastrophic wind impacts on temperate forests. In: Lupo A (ed) Recent hurricane research—climate dynamics and societal impacts. IntechOpen, London, p 616
Acknowledgements
This work was supported by the Swiss State Secretariat for Research, Education and Innovation (SERI) under Grant No. C13.0069 in the context of the COST Action FP1206 EuMIXFOR. We thank especially Paul Henne for providing input data for LandClim, Timothy Thrippleton for helpful comments on an earlier draft of this manuscript, and Dominic Michel for his programming and general IT support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Schuler, L.J., Bugmann, H., Petter, G. et al. How multiple and interacting disturbances shape tree diversity in European mountain landscapes. Landscape Ecol 34, 1279–1294 (2019). https://doi.org/10.1007/s10980-019-00838-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-019-00838-3