
Abstract
Context
Ungulate browsers often alter plant composition and reduce diversity in forests worldwide, yet our ability to predict browse impact on vegetation remains equivocal. Theory suggests, however, that ungulate distribution and foraging impacts are shaped by scale-dependent decisions based on variation in habitat composition and structure encountered within their home range.
Objective
Examine how variation in habitat composition at landscape (259 ha) scales modulates browse impact on vegetation at local scales.
Methods
We measured vegetation richness and abundance in plots with and without white-tailed deer (Odocoileus virginianus) at 23 northern hardwood forest sites distributed across a 6500 km2 area in Pennsylvania, USA. Experimental sites were embedded within landscapes with varying levels of habitat composition and deer densities.
Results
Browsing reduced vegetation richness and cover by as much as 53 and 70%, respectively; however, we found browse impact was modulated by variation in the relative abundance of managed habitats that alter forage availability. Specifically, relative to fenced areas, browse impact weakened and ultimately disappeared as the proportion of forage-rich habitats (e.g., recent harvests) increased to ≥20%. Conversely, vegetation grew increasingly depauperate as landscapes contained greater proportions of forage-poor habitats (i.e., older harvests), particularly when browsed.
Conclusions
Our results underscore how management actions that alter forage availability to ungulates throughout the landscape (i.e. the foodscape) can shape forest-ungulate interactions and suggest a new paradigm whereby managers evaluate and undertake actions at the appropriate spatio-temporal scales to proactively limit the deleterious impact of browsing on plant biodiversity.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Abundant empirical and theoretical evidence exists demonstrating that ungulate overbrowsing can undermine multiple ecosystem services provided by forests including biodiversity and sustainable wood production (Gill 1992; Russell et al. 2001; Rooney and Waller 2003; Côté et al. 2004). Consequently, research has focused on developing approaches to mitigate browse impacts including identifying population density thresholds compatible with sustaining diversity, population control, and fencing (Porter et al. 1991; Horsley et al. 2003; Vercauteren et al. 2006; Tremblay et al. 2007). While these approaches have merit, the fundamental question of predicting when and how ungulate foraging negatively affects plant biodiversity remains unanswered (Gordon et al. 2004; Wisdom et al. 2006). Thus, land managers are confronted with the dilemma of sustaining forest diversity within unpredictable and variable browsing regimes and often are forced to employ costly measures to maintain biodiversity (Waller and Alverson 1997).
Ecological theory, however, argues that ungulate distributions and foraging behavior are mediated by scale-dependent decisions based on habitat structure, resource availability, and predation risk (Senft et al. 1987; Kie et al. 2002; Hobbs 2003). In particular, spatio-temporal variation in the diversity, distribution, and abundance of habitat patches that provide forage or cover can shape population size, home range, and thus, browsing impacts across the landscape (e.g., Kie et al. 2002; Månsson 2009; Millington et al. 2010; Massé and Côté 2012). The growing awareness of scale-dependency has led to two differing views regarding how habitat diversity and abundance interact with resident ungulate herds to shape foraging impacts.
One view conjectures that variability in habitats at larger scales is critical in moderating ungulate foraging impacts at local scales (Johnson et al. 1995; Takada et al. 2002; Augustine and deCalesta 2003). This view suggests browse impact on vegetation may be weaker in forested landscapes containing a diverse configuration of habitat patch types and sizes, relative to large and relatively homogeneous forested landscapes. Consequently, from a forest management perspective, natural and anthropogenic disturbances that create a diversity of habitats, particularly those that increase forage, should mitigate browse impact (deCalesta and Stout 1997; Månsson 2009; Miller et al. 2009; Hurley et al. 2012; Herfindal et al. 2015).
Alternatively, others hypothesize that ungulate populations will respond numerically via increased fecundity to increases in forest habitat and structural diversity (Cadenasso and Pickett 2000; Kie et al. 2002) and thereby, ultimately intensify browse impact on vegetation (Alverson et al. 1988; Augustine and Jordan 1998; Palmer et al. 2003; Reimoser et al. 2009). Hence, this viewpoint suggests that land-use decisions that minimize disturbance, such as maintaining large, contiguous forested blocks or low-intensity silvicultural systems (e.g., single-tree selection), may mitigate browse risk whereas more extensive and intensively managed areas would ultimately increase browse risk (Reimoser and Gossow 1996; Kramer et al. 2006; Faison et al. 2016).
To date, empirical evidence testing the degree to which browse impact on vegetation is scale-dependent on variation in ungulate densities and habitat composition remains limited and inconsistent. Existing findings suggest browse risk is mitigated (e.g., Hurley et al. 2012; Jarnemo et al. 2014), intensified (e.g., Reimoser and Gossow 1996; Faison et al. 2016), or even unaffected (e.g., Möst et al. 2015; Evans et al. 2016) by variation in landscape composition. Generalizing from existing work is problematic given that most existing studies offer limited replication, rely solely on existing variation in ungulate populations and habitat configuration, or utilize metrics that may inadequately capture the spatial complexities that shape foraging (e.g., one-dimensional metrics such as distance to edge). More importantly, existing studies lack vegetation surveys in paired exclosures. Consequently, it is difficult to separate browsing effects from site-to-site variation in plant diversity or additional, confounding factors (Côté et al. 2004).
Untangling these relationships is critically important to maintain diversity and productivity in forests where wide-ranging large ungulates exist at high density (Millington et al. 2010). Regrettably, Beguin and colleagues (2016) recently concluded that discerning how land management actions across scales shape browse risk remains equivocal. Here, we provide insight into this question using a hybrid experimental approach that incorporates a manipulative (i.e., fence/control) treatment to test how localized (i.e., stand-level) browse impact by white-tailed deer (Odocoileus virginianus) varies among multiple, broadly distributed sites that vary in deer densities and habitat diversity. We hypothesized that variation in habitat types and abundance at the scale of a resident deer’s home range shapes deer foraging behavior, thereby modulating browse impact on plant diversity at local scales. We predict that as the proportion of forage-producing habitat increases within the resident deer’s home range, localized browse impact will decline, and vegetation cover, richness, and diversity will increase. Nonetheless, an alternative hypothesis is also possible wherein deer populations experience a numerical response as a function of habitat quality. If so, we predict increasing proportions of forage-producing habitats will cause a concomitant upsurge in deer populations, intensified browse impact, and decreases in vegetation cover, richness and diversity. In either case, we further predict the combination of habitat variables and deer density will predict browse impact on vegetation better than models containing deer densities alone.
Methods
Study area
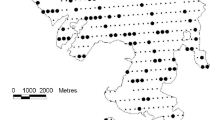
We conducted our study at 23 sites distributed across a 6500 km2 area of Pennsylvania, USA (Fig. 1, Online Appendix 1). Major tree species include red maple (Acer rubrum), sugar maple (A. saccharum), black cherry (Prunus serotina), and American beech (Fagus grandifolia), with lesser abundances of hemlock (Tsuga canadensis), birches (Betula alleghaniensis, B. lenta), and white ash (Fraxinus americana) (Marquis 1975). Understory vegetation is dominated by unpalatable striped maple (A. pensylvanicum) and beech in the shrub layer, and ferns (e.g., Dennstaedtia punctilobula) in the herb layer (Royo et al. 2010b).

Map showing the 23 study sites scattered throughout a four county region in northwest Pennsylvania, USA. Dots represent the approximate location of experimental stands and the center of a 900 m radius (259 ha) buffer within which landscape metrics were calculated
Deer populations within Pennsylvania, as throughout much of the USA, were excessively high for much of the 20th century (i.e., 15–23 deer/km2; Horsley et al. 2003). Beginning in 2003, changes in deer management policies, including additional antlerless licenses and concurrent buck-doe hunting seasons, allowed many of the public and private industrial landowners in the region to intensify hunting on their lands, resulting in deer herd reductions for over a decade (Royo et al. 2010b; Stout et al. 2013; see Online Appendix 1).
Experimental design
At each of the 23 sites, we selected one stand where managers initiated a regeneration sequence as the location of the experimental treatment. In all stands, managers conducted the initial cut of a shelterwood sequence to reduce stand relative density to levels appropriate for hardwood regeneration in mature Allegheny and northern hardwood forests (i.e., <75% relative density; Marquis et al. 1992). With the exception of one site (Regen134), shelterwood harvests occurred 0–5 years prior to fence construction (Online Appendix 1). The harvest reduced basal area across all sites from an average of 26.6–16.6 m2/ha. Additionally, 22 stands received broadcast herbicide treatments (glyphosate and sulfometuron methyl; Marquis et al. 1992) to control dense layers of interfering plant species within 4 years prior to fence construction (Online Appendix 1). By deploying our experiment within managed stands, we can (1) detect browse impacts on vegetation more rapidly and on a greater variety of species than in uncut areas (Horsley et al. 2003); (2) reduce the potential for lagged responses attributed to established recalcitrant layers (Royo et al. 2010b); and (3) directly link our results to common forest management actions.
Within each stand, we established paired 4200 m2 (60 × 70 m2) plots and randomly assigned one plot a deer exclosure (fence) treatment while the other served as an unfenced control. Exclosure construction was completed by September 2013 except for one site (Rush) on which harvest occurred during winter of 2013/14. At this site, the exclosure was constructed in early spring 2014. Centered within each plot, we established a 2000 m2 (40 × 50 m2) sampling area, divided into a 10 × 10 m2 grid, leaving a ~10 m wide buffer between the perimeter of the plots and the sampling area.
Data collection
In summer 2015, two growing seasons after exclosure construction, we censused the plant community in 2 m radius (12.56 m2) subplots located at five randomly chosen nodes of the 10 × 10 m2 grid. Vegetation was censused in the vertical dimension at 20 cm intervals from ground level to 2 m in each of the four cardinal directions (see Collins and Becker 2001; Rohleder 2013). We recorded vascular plant species richness of the vegetation profile in each height interval along a 2-m line-intercept in each cardinal direction and species frequency by height class as the proportion of sample directions in which a species was tallied. Lastly, we estimated total vegetation cover along the vertical profile using a 20 × 20 cm2 cover template placed at the plot center viewed horizontally from a distance of 2 m in each cardinal direction (Online Appendix 2).
For each site, we assessed habitat composition at the landscape scale by developing 259 ha land cover maps centered on each experimental site. Following Putman and colleagues’ (2011) advice, we scaled our assessment of habitat configuration to the typical home range size of the resident ungulate species. Hence, we chose a 259 ha buffer size because within the northern-hardwood region of the eastern U.S., this area encompasses the typical deer mean home range size (range 64–223 ha; Tierson et al. 1985; Campbell et al. 2004). Maps were developed using the 2011 National Land Cover Database (Homer et al. 2015) as a basemap and augmented with aerial imagery, landowner spatial data, and on-the-ground verification. We identified 10 distinct habitats: Mature Forest, Shelterwood, Recent Removal, Older Removals (hereafter, stem exclusion stage), Thinning/Salvage, Conifer, Herbaceous, Water, Developed Areas, and Agriculture. We classified mature forest as any closed canopy hardwood stand that had not received recent timber harvest. Shelterwood was reserved for stands with a shelterwood harvest within the past 5 years. Recent removals were stands that had received a clear-cut ≤5 years prior to the year of classification, whereas areas entering stem exclusion stage were clear-cut areas >5 years but ≤17 years prior to classification. Thinning and salvage timber areas were a distinct classification wherein a portion of the mature overstory was removed, but there were no immediate plans to proceed to a clearcut. Conifer areas contained >90% conifer (primarily hemlock) in the overstory. Herbaceous patches are dominated by non-woody vegetation (e.g., food plots, rights-of-ways, and failed regeneration harvests). Developed areas included roads, buildings, and other infrastructure. Agriculture included pastures and active or fallow cropland. Lastly, water represented areas where water is present during all or portions of the year (e.g., reservoirs, streams). We resampled the basemap to a higher resolution of 5 m following classification and used available landowner data to identify the timing of forest management treatments concluded prior to the start of winter 2015. Questionably classified areas were verified using aerial imagery from USGS National Agriculture Imagery Program (NAIP) and Google Earth for the summers of 2013, 2014, and 2015 and field visits in fall of 2015. Thus, our map captures the diversity and abundance of habitats within the home range of deer in the year vegetation data were sampled (see Online Appendix 1).
Finally, we estimated deer densities throughout each of the 259 areas surrounding each stand using fecal pellet surveys. Data were collected along five, 1.6 km transects spanning the site in spring (April to early May) of 2013–2015. Within each transect, deer pellet group counts were obtained on fifty-two, 4.67 m2 plots spaced 30.5 m apart. Estimates derived from pellet group counts represent the average number of deer over winter for each site and therefore, the number of adult deer surviving into the spring and summer (see deCalesta 2013 for further details on methodology).
Statistical analyses
To avoid model overfitting our analyses proceeded hierarchically. We first screened potential analysis of covariance variables for collinearity using correlation analyses (Online Appendix 3). Additionally, as our hypotheses make specific predictions regarding available forage, we aggregated land cover classes capable of producing forage into a composite variable: propforage. These include recent forest management areas (i.e., shelterwood, recent removal, thinning, and salvage harvests), agricultural areas, and herbaceous habitats. Stem exclusion patches generally lack abundant forage as sapling canopy closes and is increasingly out of reach of deer and the ensuing low-light conditions reduce herbaceous cover to very low levels (Johnson et al. 1995; Roberts 2004). Unmanaged, mature forest strongly covaried (r = −0.75) with propforage as it encompassed most of the habitat remaining across the landscape after accounting for managed forest habitats, and was therefore removed from further consideration.
Following preliminary variable screening, we used an exploratory model selection approach with model averaging to determine whether treatment (fence/control) and the landscape covariables of deer density and proportions of the landscape in forage, stem exclusion, and conifer cover were robust predictors to include in final models. We pooled the data and fit linear models to the mean species richness, total cover (log[cover + 0.1]), and Shannon diversity (H′) of each experimental unit (i.e., average of five subplots within each plot). For these models, treatment was converted to binary variable (0/1) and all possible variable combinations were examined the R function dredge (MuMIn package; Barton 2013). We compared competing models using AICC scores and weights and present AICC-weighted average parameter estimates across the top models (ΔAICC < 2; Burnham and Anderson 2002).
We used a generalized linear mixed model to test how plant community responses varied as a function of deer browsing alone and interactions between deer browsing and selected landscape-level covariates. We modelled treatment (fence/control) and height as fixed effects. The model included one random effect for site and a second random effect of height nested within treatment(site) to account for the hierarchical nature of the design (Milliken 2006). We used an autoregressive first order covariance structure (AR(1)) in the G-matrix to model the dependence in samples between successive height strata. We initially tested the single treatment effect of fence to examine whether localized deer browsing affected vegetation across the entire 2 m profile, as established by Rohleder’s (2013) methods. However, as our sites were fairly early in stand development, vegetation was almost uniformly low in stature with all of the species and 75% of the total plant cover contained in the first 60 cm of height. Hence, to examine whether localized deer browsing impact on the plant community varied as a function of landscape-level covariates, we restricted our analyses to the lowermost 60 cm in height, corresponding to the three lowest height strata.
All analyses included a treatment × covariate term to test the homogeneity of slopes assumption. This interaction is central to our hypothesis testing whether landscape covariates modulate vegetation responses to browsing as a significant interaction would indicate treatment effects do not respond in a similar manner (i.e., non-parallel) to variation in the covariate. In cases where treatment was significant (P < 0.10), we utilized least-squared means to test pair-wise differences and, where treatment × covariate interactions were significant (P < 0.10), we tested for treatment differences along the entire range of the landscape covariate in 5% increments using least-squared means (Proc Glimmix; SAS Institute Inc. 2011). This approach allowed us to assess, for example, at which point along the landscape covariate, vegetation responses to browsing in control plots became indistinguishable from fenced plots.
We did not include the simple main effects of the landscape covariates as these were collinear with the blocking term Site (i.e., each site has unique values for landscape covariates) and were applied to responses in both the fence and control plots. Instead, we examined whether responses against landscape covariates had non-zero slopes. For all analyses, we utilized the Kenward-Rogers denominator degrees of freedom method. Compliance to the ANOVA assumptions of homoscedasticity and normality of the residuals was assessed using boxplots and Levene’s test. Species richness was modelled using a Poisson distribution, percent cover was modelled using a gamma distribution, and diversity (H′) was modelled using a normal distribution.
To examine whether species composition differed across treatments, and treatments × landscape covariate interactions, we ran partial redundancy analyses (RDA) using the Jaccard distance metric on the mean frequency of species throughout 0–60 cm stratum. Redundancy analyses are the canonical extension of principal component analysis (PCA) and have the flexibility of allowing the effects of known covariables, such as spatial blocking effects, to be accounted for (i.e., ‘partialled out’; ter Braak 1988). Species present on <3 plots (42% of species) were culled from the analyses as rare species exert unduly large influence in multivariate analyses and distort interpretation (McCune and Grace 2002). Species frequencies were standardized using the Hellinger standardization following Legendre and Gallagher (2001). To account for the paired nature of the design, site was included as a conditional covariable in the RDA and permuted tests of significance (999 permutations) were constrained within sites using the strata option in the vegan package in R version 3.1.2 (Oksanen et al. 2015). Species associations with the constraining variables were visualized using RDA biplots. Community composition analyses were run using the rda and anova functions in the vegan package.
Results
General patterns
We censused vegetation in plots covering a two-dimensional footprint of ~3141 m2 and a total volume across the entire vegetation profile of 6282 m3. We surveyed 61 taxa of which 16 were tree species. Across the entire 2-m height, deer browsing reduced overall plant species richness by 53% (RichnessTrt: F1,73.3 = 6.31; P = 0.01; Fig. 2a) and overall cover by 70% (CoverTrt: F1,34.4 = 41.73; P < 0.0001; Fig. 2b, see also Table 2 for similar results on lower strata). Progressively taller strata grew increasingly depauperate, exhibiting both fewer species and less cover (RichnessHt: F9,389.3 = 47.6; P < 0.0001 and CoverHt: F9,370.2 = 85.23; P < 0.0001; Fig. 2) and browsing exacerbated this decline for cover (CoverTrt×Ht: F9,370.2 = 6.85; P < 0.0001). Across all strata, the most abundant species were Rubus allegheniensis (16% relative frequency), Dennstaedtia punctilobula (9%) and various grasses and sedges (17% combined). Among the tree species, Prunus pensylvanica (8%), Acer rubrum (8%), and Betula spp. (B. allegheniensis and B. lenta; 4.8%) were the most frequently tallied.

Mean overall (± 1SE) percent cover (a) and species richness (b), by 20 cm interval height strata, with and without ambient browsing. In both responses there were significant (P < 0.05) treatment and height main effects. Percent cover also had a significant treatment × height interaction and therefore we further examined treatment differences within height strata. Asterisks (*) denote a significant pair-wise difference in treatment within a height strata
Habitat composition modulates browse impact
The most plausible (i.e., lowest AICC) models predicting species richness, cover, and diversity all included treatment (fence vs control), forage-producing habitats, and stem exclusion stands as essential landscape variables (Table 1A). In fact, stem exclusion stands were selected in all models receiving competitive support (Δ AICC < 2). Forage-producing habitats were selected in 78% of all alternative, competitive models. None of the remaining landscape variables, including average deer density, were identified in the most plausible (lowest AICc) model. While average deer density and proportion conifer appeared in the confidence set of models, confidence intervals crossed zero. Additionally, models without any landscape variables (i.e., intercept only or an intercept + treatment models) were not identified as competitive or plausible models (e.g., high AICc values, low R2 values, low weights). Excluding deer and increasing proportions of forage habitat were associated with greater richness, cover, and diversity whereas increasing the proportion of stands in stem exclusion diminished these metrics (Table 1B).
Land cover covariates consistently modulated browse impact across the lowermost strata of the vegetation profile (Table 2). Specifically, we found the negative impacts of deer browsing on richness and abundance weakened as the proportion of forage-producing habitats increased across the landscape (significant treatment × propforage interaction; Table 2; Fig. 3). This relationship was not observed within areas where deer were excluded. Comparisons of the two treatments along the propforage covariate found that when landscapes contained ≥20% in forage-producing habitats, the plant community richness cover became indistinguishable from that found within exclosures (Online Appendix 4). Overall species richness, abundance, and diversity grew increasingly depauperate in landscapes with greater proportions of stem exclusion stands, and particularly so in areas with ambient browsing (significant treatment × stmex interaction; Table 2; Fig. 4). However, even within exclosures, we detected richness and diversity declines in sites embedded in landscapes with more area in stem exclusion stands.

Relationships between percent of landscape (259 ha) in forage-producing habitats and overall a species richness and b cover, in the 0–60 cm strata with (filled circle) and without (open circle) ambient browsing. For richness and cover data plotted are back-transformed values of the Poisson and gamma distributions, respectively. Data analysis was based on a hierarchically nested approach wherein each height strata within a plot was a unit of observation and the nestedness and covariance structure among successive heights within plot were accounted for in the model. For convenience, data are presented separately for each height strata (0–20; 20–40; 40–60 cm). See Table 2 for analyses

Relationships between percent of landscape (259 ha) in stem exclusion habitat and overall a species richness and b cover, in the 0–60 cm strata with (filled circle) and without (open circle) ambient browsing. For richness and cover data plotted are back-transformed values of the Poisson and gamma distributions, respectively. Data analysis was based on a hierarchically nested approach wherein each height strata within a plot was a unit of observation and the nestedness and covariance structure among successive heights within plot were accounted for in the model. For convenience, data are presented separately for each height strata (0–20; 20–40; 40–60 cm). See Table 2 for analyses
Deer browsing significantly altered species composition (RDATrt: F1,20 = 2.19, P = 0.004; Online Appendix 5). The RDA indicated that in areas with ambient browsing, ferns and graminoids (D. punctilobula, Carex spp., T. noveboracencis) were 20, 37, and 300% more frequent, respectively. In contrast, eliminating browsing increased the frequency of P. pensylvanica, F. grandifolia, Magnolia acuminata, and Sambucus spp. by 177, 192, 358 and 204%, respectively. Additionally, in areas with ambient browsing, stands embedded in landscapes with greater proportions of forage-producing habitats resulted in greater frequencies of some species including Polygonum cilinode, Fragaria virginiana, and Eurybia divaricata (RDATrt×PF: F1, 20 = 1.60, P = 0.057).
Discussion
Dynamic foodscapes
Our experiment demonstrates that variation in the relative abundance of key habitats at large spatial scales modulates localized browse impact in two diametrically opposed ways: disturbances that open up forest canopies and create early-successional habitats initially mitigate browse impact whereas expanding mid-successional habitats intensify browsing. In the largely forested landscapes where we conducted our study, land cover changes are mainly created by harvesting, although herbaceous areas and agricultural patches occur and natural disturbances are common (Online Appendix 1, Royo et al. 2016b). Overstory tree harvests, coupled with the herbicide applications, increased light availability and disturbed the soil surface thereby promoting plant germination, growth, flowering, and fruiting within the lower vegetation strata. The concomitant changes in composition and structure caused by harvesting creates high quality forage patches favored by wildlife (reviewed by Greenberg et al. 2011) and mitigate localized browse impact in the short term (deCalesta and Stout 1997; Månsson 2009; Miller et al. 2009; reviewed by Hurley et al. 2012; Gerhardt et al. 2013; Herfindal et al. 2015). Our study extends and enhances these prior findings by unequivocally isolating deer impact across nearly two dozen sites that vary greatly in habitat composition and deer densities. Moreover, our study identifies target levels of area under forest management beyond which negative effects of browsing on plant community richness, diversity, and abundance are completely mitigated. Our analyses indicate that, under the range of deer densities observed in our study, when approximately 20% of the landscape (52/259 ha) is in forage-producing habitats (e.g., forest harvests, agriculture, herbaceous openings), browse impact on cover and diversity of the plant community is indistinguishable from that found within a fence.
Our findings also indicate that interactions between browsing and variation in relative abundance of forage-producing habitats create communities with contrasting plant composition. Excluding deer benefited fast-growing, highly preferred species trees, particularly P. pensylvanica (Horsley et al. 2003). Under ambient browsing, however, species composition in areas embedded in landscapes with high amounts of alternative forage did not mirror that found within a fence. Instead, the habitat-mediated reductions in browse impact allowed moderately preferred species, including Polygonum cilinode, Fragaria virginiana, and Eurybia divaricata (Atwood 1941) to persist. In contrast, as forage became less available to resident deer herds, browsing intensified and constricted species composition toward the least nutritious, browse tolerant and highly recalcitrant species, including ferns and sedges (Royo and Carson 2006). These findings suggest that deer may suppress or promote species coexistence, depending on landscape context. Although herbivory and stand-level disturbances are known to promote coexistence where they co-occur (Royo et al. 2010a; Faison et al. 2016), such interactions occurring at larger spatial scales remain largely unexplored.
Our findings also suggest the role of disturbance in modulating browsing is temporally dynamic as changes to vegetation structure, composition, and abundance that occur during succession eventually reverse and intensify browse impact. We found plant communities became increasingly depauperate when embedded in landscapes with higher proportions of stem exclusion stands. We suggest this effect is caused by both historic and present intensification of browse pressure. During stem exclusion, plant biomass becomes increasingly concentrated in the competing tree canopy and, concomitantly, woody and herbaceous biomass available to deer in lower strata declines (Christensen and Peet 1984; Johnson et al. 1995; Howard and Lee 2003; Roberts 2004). Hence, from an ungulate browser’s perspective, stem exclusion stands present a tradeoff between suboptimal, forage-poor habitat and dense hiding cover, both of which will concentrate foraging activity in forage hotspots created by harvesting (Partl et al. 2002; Vospernik and Reimoser 2008; Månsson 2009). Within our study areas, stem exclusion stands predate the building of the exclosures. Hence, we suggest our sites experienced past browse pressure that intensified in landscapes with abundant stem exclusion stands which may have negatively impacted plant richness, cover and diversity, even in areas that were ultimately fenced.
Considered in their entirety, our findings suggest habitat-mediated effects on foraging activity and browse pressure are both scale-dependent and dynamic. Our findings indicate that in largely forested areas, the application of even-aged forest management practices that stimulate vegetation establishment and growth in hardwoods initially mitigates and ultimately intensifies browse impacts on vegetation as successional changes to forage availability cause deer to concentrate browsing. Hence, the modulating effect of disturbance-driven habitat heterogeneity on browsing changes as the ‘foodscape’ (sensu Searle et al. 2007) throughout the home range of the ungulate population may wax and wane both spatially and temporally.
Reconciling opposing predictions
Our experimental findings support the hypothesis that variation in habitat composition, and particularly the relative abundance of forage patches, mitigates the negative effect of browsing on plant cover and species richness. This modulating effect assumes deer populations are relatively stable throughout the landscape. However, natural or anthropogenic disturbance that increases productivity and forage availability increase carrying capacity and, thereby, may eventually increase deer populations (Gill et al. 1996; Kramer et al. 2006) and, thereby, ultimately intensify browse impacts (Alverson et al. 1988; Augustine and Jordan 1998; Reimoser et al. 2009). Moreover, some have argued that any modulating effects of increasing forage throughout a landscape may be overwhelmed at high deer densities (Augustine and Jordan 1998; Beguin et al. 2016). Our findings cannot refute these alternative predictions as our experiment was carried out under conditions where landowners and game management policies encouraged strong top-down control over deer population growth by increasing hunting opportunities and courting hunter participation (Stout et al. 2013). In fact, although average deer densities in our study were higher than precolonial estimates (<4 deer/km2; McCabe and McCabe 1997), they are intermediate between thresholds identified to maintain overall plant biodiversity (~4 deer/km2; Alverson et al. 1988; deCalesta and Stout 1997) and negative thresholds identified for tree regeneration (~8 deer/km2; Horsley et al. 2003; Tremblay et al. 2007). Moreover, within our study areas, deer populations remained stable over time, even in areas with high forage (see Online Appendix 1 and 3). Thus, our results apply to managed forests in which deer densities are moderately high (median = 6.3 deer/km2; range 3.2–12.3 deer/km2) and which are largely open to seasonal hunting pressure. Therefore, it is not clear the degree to which our conclusions would apply in landscapes with weak top-down control (i.e., lax hunting, absence of predators) where deer populations would be free to respond positively to increasing forage availability. Nonetheless, even under those conditions, populations may build over a few years as a result of lags in juvenile recruitment and low emigration from nearby social groups (Fryxell et al. 1991; Miller et al. 2010). Understanding spatiotemporal dynamics and interplay between vegetation development and ungulate populations should be a focus of future studies.
Deer matter, but density estimates lack precision
Contrary to expectations, we found estimated deer densities were not a robust predictor of vegetation responses, even when coupled with landscape predictors. However, we do not conclude deer are unimportant in driving vegetation dynamics in this system. Indeed, our results demonstrate that browsing limited species richness, abundance, diversity, restricted recruitment of species into taller strata, and altered species composition. We suggest the reason average deer densities fail to consistently emerge as a robust predictor in this and other studies may lie in the precision of the variable itself relative to the response (Royo et al. 2010b). Indeed, although fecal surveys provide a comparative population density index at the scale critical to our analyses (Forsyth et al. 2007), variability in fecal deposition and persistence rates may render them unreliable as predictors of localized impact (Putman et al. 2011). Indeed, estimates based on standing crop (i.e., without removing prior year’s pellets) may be biased because pellets may take years to disappear (Alves et al. 2013). Moreover, indices based on over winter fecal deposition rates reflect deer habitat use during a time when deer limit their movement and seek thermal cover (e.g., conifer stands). Thus, the temporal disconnect between winter thermoregulation needs and summer browse foraging in harvested areas may limit the utility of these estimates. Additionally, although we used a 3-year average density estimate in our models, observed browsing impacts at individual sites may be highly idiosyncratic, resulting from foraging bouts that occurred in the past, present, or both, hence further complicating interpretation. Finally, efforts to relate aggregate plant community metrics (e.g., richness, abundance) to densities of selective foragers like deer is problematic as variation in the composition and abundance of the plant community may mask interpretation. Given these complications, it is not surprising that studies that detect relationships between deer density and vegetation are generally focused on highly palatable species (e.g., Augustine and Jordan 1998), specific growth forms (e.g., woody sprouts; Royo et al. 2016a), subsets of the plant community (e.g., native herb cover; Hurley et al. 2012), or analyses that are sensitive to changes in species composition (Bachand et al. 2015).
Proactive management of deer impact
Our work provides empirical evidence to substantiate a paradigm shift in how land managers can sustainably manage their forests under resident ungulate herds (Reimoser and Gossow 1996). Currently, management options to sustain and promote plant diversity are reactive and costly, including the removal of matriarchal social groups from localized areas, localized culls using sharpshooters, or the widespread use of fencing (Porter et al. 1991; Vercauteren et al. 2006; Miller et al. 2010). Our findings inform guidelines for a distinct alternative whereby managers manipulate the landscape on the appropriate temporal and spatial scales to mitigate deer impacts on plant biodiversity at the stand level. Specifically, we suggest that in forested landscapes managed under even-aged silviculture in which deer are hunted, land managers might consider entering approximately 20% of the area within the average home range of deer into a regeneration sequence within 5 years in order to buffer plant communities from browsing. This proportion may be adjusted downward, according to the degree to which alternative forage patches already exist within the landscape (e.g., agriculture). Following these initial entries, we recommend managers limit any additional harvesting within the area as browse risk will increase over the next couple decades as stands transition into stem exclusion. Indeed, stem exclusion patches are an important consideration, as we show browse risk intensifies as the relative abundance of this habitat type increases. Harvests in landscapes containing abundant stem exclusion stands may require increased deer harvest pressure or fencing to mitigate potential risks to biodiversity.
Our recommendation regarding the proportion of the landscape in forage producing habitats is higher than Miller and colleagues’ (2009) recommendation of 14% in <10 year old stands and Stout and colleagues’ (2013) finding that 15% of early successional forests are sufficient to ensure diverse regeneration without fencing and enhance growth and reproduction of browse sensitive herbs. The lower proportions identified by Miller et al. (2009) and Stout et al. (2013) relative to ours may arise as a consequence of their harvests all being recent removals which stimulate abundant regeneration, whereas our classification combines recent removals with partial harvests containing a more subdued regeneration response, and hence, less forage (Horsley et al. 2003).
Our findings are applicable to forests across the eastern United States and elsewhere where forest managers practice even-aged silviculture and contend with browse impact from white-tailed deer. Collectively, our results underscore the importance of understanding how management practices exert scale-dependent influences on ungulate foraging which may intensify or mitigate conflicts between deer and forest management (Jarnemo et al. 2014; Beguin et al. 2016). More broadly, our results support the growing consensus of the benefits of early successional forest habit to biodiversity conservation (Swanson et al. 2011) and demonstrate that forest management may mitigate deer impact and maintain biodiversity while circumventing the contentious socioeconomic-political debates that often surround ungulate management in forests.
References
Alverson WS, Waller DM, Solheim SL (1988) Forests too deer: edge effects in northern Wisconsin. Conserv Biol 2(4):348–358
Alves J, Alves da Silva A, Soares AMVM, Fonseca C (2013) Pellet group count methods to estimate red deer densities: precision, potential accuracy and efficiency. Mammalian Biol 78(2):134–141
Atwood EL (1941) White-tailed deer foods of the United States. J Wildl Manag 5(3):314–332
Augustine DJ, deCalesta D (2003) Defining deer overabundance and threats to forest communities: from individual plants to landscape structure. Ecoscience 10(4):472–486
Augustine DJ, Jordan PA (1998) Predictors of white-tailed deer grazing intensity in fragmented deciduous forests. J Wildl Manag 62(3):1076–1085
Bachand M, Pellerin S, Tremblay JP, Cote SD, Poulin M (2015) Compositional and functional trajectories of herbaceous communities after deer density control in clear-cut boreal forests. Can J For Res 45(6):758–763
Barton K (2013) MuMIn: Multi-model inference. R package version http//CRAN.R-project.org/package = lme4
Beguin J, Tremblay JP, Thiffault N, Pothier D, Côté SD (2016) Management of forest regeneration in boreal and temperate deer–forest systems: challenges, guidelines, and research gaps. Ecosphere 7(10):e01488
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Cadenasso ML, Pickett STA (2000) Linking forest edge structure to edge function: mediation of herbivore damage. J Ecol 88(1):31–44
Campbell TA, Laseter BR, Ford WM, Miller KV (2004) Topographic home ranges of white-tailed deer in the central Appalachians. Southeast Nat 3(4):645–652
Christensen NL, Peet RK (1984) Convergence during secondary forest succession. J Ecol 72:25–36
Collins WB, Becker EF (2001) Estimation of horizontal cover. J Range Manag 54:67–70
Côté SD, Rooney TP, Tremblay JP, Dussault C, Waller DM (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35(1):113–147
deCalesta DS (2013) Reliability and precision of pellet-group counts for estimating landscape-level deer density. Human Wildl Interact 7(1):60–68
deCalesta DS, Stout SL (1997) Relative deer density and sustainability: a conceptual framework for integrating deer management with ecosystem management. Wildl Soc Bull 25(2):252–258
Evans JP, Oldfield CA, Cecala KK, Hiers JK, Van De Ven C, Armistead MM (2016) Pattern and drivers of white-tailed deer (Odocoileus virginianus) herbivory on tree saplings across a plateau landscape. Forests 7(5):101
Faison EK, DeStefano S, Foster DR, Motzkin G, Rapp JM (2016) Ungulate browsers promote herbaceous layer diversity in logged temperate forests. Ecol Evol 6(13):4591–4602
Forsyth DM, Barker RJ, Morriss G, Scroggie MP (2007) Modeling the relationship between fecal pellet indices and deer density. J Wildl Manag 71(3):964–970
Fryxell JM, Hussell DJT, Lambert AB, Smith PC (1991) Time lags and population fluctuations in white-tailed deer. J Wildl Manag 55:377–385
Gerhardt P, Arnold JM, Hackländer K, Hochbichler E (2013) Determinants of deer impact in European forests—A systematic literature analysis. For Ecol Manag 310:173–186
Gill RMA (1992) A review of damage by mammals in north temperate forests: 1. Deer. Forestry 65(2):145–169
Gill RMA, Johnson AL, Francis A, Hiscocks K, Peace AJ (1996) Changes in roe deer (Capreolus capreolus L.) population density in response to forest habitat succession. For Ecol Manag 88(1–2):31–41
Gordon IJ, Hester AJ, Festa Bianchet M (2004) The management of wild large herbivores to meet economic, conservation and environmental objectives. J Appl Ecol 41(6):1021–1031
Greenberg CH, Perry RW, Harper CA, Levey DJ, McCord JM (2011) The role of young, recently disturbed upland hardwood forest as high quality food patches. In: Greenberg CH, Collins BS, Thompson FR III (eds) Sustaining young forest communities. Springer, Dordrecht, pp 121–141
Herfindal I, Tremblay J-P, Hester AJ, Lande US, Wam HK (2015) Associational relationships at multiple spatial scales affect forest damage by moose. For Ecol Manag 348:97–107
Hobbs NT (2003) Challenges and opportunities in integrating ecological knowledge across scales. For Ecol Manag 181(1–2):223–238
Homer CG, Dewitz JA, Yang L, Jin S, Danielson P, Xian G, Coulston J, Herold N, Wickham J, Megown L (2015) Completion of the 2011 National Land Cover Database for the conterminous United States-Representing a decade of land cover change information. Photogr Eng Remote Sens 81(5):345–354
Horsley SB, Stout SL, deCalesta DS (2003) White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol Appl 13(1):98–118
Howard LF, Lee TD (2003) Temporal patterns of vascular plant diversity in southeastern New Hampshire forests. For Ecol Manag 185(1–2):5–20
Hurley PM, Webster CR, Flaspohler DJ, Parker GR (2012) Untangling the landscape of deer overabundance: reserve size versus landscape context in the agricultural Midwest. Biol Conserv 146:62–71
Jarnemo A, Minderman J, Bunnefeld N, Zidar J, Månsson J (2014) Managing landscapes for multiple objectives: alternative forage can reduce the conflict between deer and forestry. Ecosphere 5(8):1–14
Johnson AS, Hale PE, Ford WM, Wentworth JM, French JR, Anderson OF, Pullen GB (1995) White-tailed deer foraging in relation to successional stage, overstory type and management of southern Appalachian forests. Am Midl Nat 133(1):18–35
Kie JG, Bowyer RT, Nicholson MC, Boroski BB, Loft ER (2002) Landscape heterogeneity at differing scales: effects on spatial distribution of mule deer. Ecology 83(2):530–544
Kramer K, Groot Bruinderink G, Prins HHT (2006) Spatial interactions between ungulate herbivory and forest management. For Ecol Manag 226(1–3):238–247
Legendre P, Gallagher ED (2001) Ecologically meaningful transformations for ordination of species data. Oecologia 129(2):271–280
Månsson J (2009) Environmental variation and moose Alces alces density as determinants of spatio-temporal heterogeneity in browsing. Ecography 32(4):601–612
Marquis DA (1975) The Allegheny hardwood forests of Pennsylvania. USDA Forest Service, General Technical Report NE-15, Broomall, PA
Marquis DA, Ernst RL, Stout SL (1992) Prescribing silvicultural treatments in hardwood stands of the Alleghenies (revised). USDA Forest Service, General Technical Report NE-96, Radnor, PA
Massé A, Côté SD (2012) Linking habitat heterogeneity to space use by large herbivores at multiple scales: from habitat mosaics to forest canopy openings. For Ecol Manag 285:67–76
McCabe TR, McCabe RE (1997) Recounting whitetails past. In: McShea WJ, Underwood HB, Rappole JH (eds) The science of overabundance: deer ecology and population management. Smithsonian Institution Press, Washington, DC, pp 11–26
McCune B, Grace JB (2002) Analysis of ecological communities. MJM Software Design, Gleneden Beach
Miller BF, Campbell TA, Laseter BR, Ford WM, Miller KV (2009) Whitetailed deer herbivory and timber harvesting rates: implications for regeneration success. For Ecol Manag 258(7):1067–1072
Miller BF, Campbell TA, Laseter BR, Ford WM, Miller KV (2010) Test of localized management for reducing deer browsing in forest regeneration areas. J Wildl Manag 74(3):370–378
Milliken GA (2006) Introduction to design and analysis of hierarchical models. Course Notes. SAS Institute Inc., Cary
Millington JDA, Walters MB, Matonis MS, Liu J (2010) Effects of local and regional landscape characteristics on wildlife distribution across managed forests. For Ecol Manag 259(6):1102–1110
Möst L, Hothorn T, Müller J, Heurich M (2015) Creating a landscape of management: unintended effects on the variation of browsing pressure in a national park. For Ecol Manag 338:46–56
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner, H (2015) Vegan: community ecology package. R package version 2.2-1. http://cran.r-project.org/web/packages/vegan/index.html
Palmer SCF, Hester AJ, Elston DA, Gordon IJ, Hartley SE (2003) The perils of having tasty neighbors: grazing impacts of large herbivores at vegetation boundaries. Ecology 84(11):2877–2890
Partl E, Szinovatz V, Reimoser F, Schweiger-Adler J (2002) Forest restoration and browsing impact by roe deer. For Ecol Manag 159(1–2):87–100
Porter WF, Mathews NE, Underwood HB, Sage RWJ, Behrend DF (1991) Social organization in deer: implications for localized management. Environ Manag 15(6):809–814
Putman R, Watson P, Langbein J (2011) Assessing deer densities and impacts at the appropriate level for management: a review of methodologies for use beyond the site scale. Mamm Rev 41:197–219
Reimoser F, Gossow H (1996) Impact of ungulates on forest vegetation and its dependence on the silvicultural system. For Ecol Manag 88(1–2):107–119
Reimoser S, Partl E, Reimoser F, Vospernik S (2009) Roe-deer habitat suitability and predisposition of forest to browsing damage in its dependence on forest growth–Model sensitivity in an alpine forest region. Ecol Modelling 220(18):2231–2243
Roberts MR (2004) Response of the herbaceous layer to natural disturbance in North American forests. Can J Bot 82(9):1273–1283
Rohleder L (2013) The vertical dimension of deer browse effects on forest understories. Dissertation, Rutgers University
Rooney TP, Waller DM (2003) Direct and indirect effects of white-tailed deer in forest ecosystems. For Ecol Manag 181(1–2):165–176
Royo AA, Carson WP (2006) On the formation of dense understory layers in forests worldwide: consequences and implications for forest dynamics, biodiversity, and succession. Can J For Res 36(6):1345–1362
Royo AA, Collins R, Adams MB, Kirschbaum C, Carson WP (2010a) Pervasive interactions between ungulate browsers and disturbance regimes promote temperate forest herbaceous diversity. Ecology 91(1):93–105
Royo AA, Kramer DW, Miller KV, Nibbelink NP, Stout SL (2016a) The canary in the coal mine: sprouts as a rapid indicator of browse impact in managed forests. Ecol Indic 69:269–275
Royo AA, Peterson CJ, Stanovick JS, Carson WP (2016b) Evaluating the ecological impacts of salvage logging: can natural and anthropogenic disturbances promote coexistence? Ecology 97(6):1566–1582
Royo AA, Stout SL, deCalesta DS, Pierson TG (2010b) Restoring forest herb communities through landscape-level deer herd reductions: is recovery limited by legacy effects? Biol Conserv 143(11):2425–2434
Russell FL, Zippin DB, Fowler NL (2001) Effects of white-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: a review. Am Midl Nat 146(1):1–26
SAS Institute Inc. (2011) SAS system for Windows, 9.3rd edn. SAS Institute Inc, Cary
Searle KR, Hobbs NT, Gordon IJ (2007) It’s the “Foodscape”, not the landscape: using foraging behavior to make functional assessments of landscape condition. Israel J Ecol Evol 53(3–4):297–316
Senft RL, Coughenour MB, Bailey DW, Rittenhouse LR, Sala OE, Swift DM (1987) Large herbivore foraging and ecological hierarchies. Bioscience 37(11):789–799
Stout SL, Royo AA, deCalesta DS, McAleese K, Finley JC (2013) The Kinzua Quality Deer Cooperative: can adaptive management and local stakeholder engagement sustain reduced impact of ungulate browsers in forest systems? Boreal Environ Res 18:50–64
Swanson ME, Franklin JF, Beschta RL, Crisafulli CM, DellaSala DA, Hutto RL, Swanson FJ (2011) The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front Ecol Environ 9(2):117–125
Takada M, Asada M, Miyashita T (2002) Cross-habitat foraging by sika deer influences plant community structure in a forest-grassland landscape. Oecologia 133(3):389–394
ter Braak CJ (1988) Partial canonical correspondence analysis. In: Bock HH (ed), Classification and related methods of data analysis. North Holland, Amsterdam, The Netherlands, pp. 551–558
Tierson WC, Mattfeld GF, Sage RWJ, Behrend DF (1985) Seasonal movements and home ranges of white-tailed deer in the Adirondacks. J Wildl Manag 49:760–769
Tremblay JP, Huot J, Potvin F (2007) Density-related effects of deer browsing on the regeneration dynamics of boreal forests. J Appl Ecol 44(3):552–562
Vercauteren KC, Lavelle MJ, Hygnstrom S (2006) Fences and deer-damage management: a review of designs and efficacy. Wildl Soc Bull 34(1):191–200
Vospernik S, Reimoser S (2008) Modelling changes in roe deer habitat in response to forest management. For Ecol and Manag 255(3–4):530–545
Waller DM, Alverson WS (1997) The white-tailed deer: a keystone herbivore. Wildl Soc Bull 25(2):217–226
Wisdom MJ, Vavra M, Boyd JM, Hemstrom MA, Ager AA, Johnson BK (2006) Understanding ungulate herbivory-episodic disturbance effects on vegetation dynamics: knowledge gaps and management needs. Wildl Soc Bull 34(2):283–292
Wood SN (2004) Stable and efficient multiple smoothing parameter estimation for generalized additive models. J Am Stat Assoc 99(467):673–686
Acknowledgements
This work was supported by the USDA-AFRI Award #12-IA-11242302-093 and the USDA Forest Service Northern Research Station. We are indebted to the Allegheny National Forest, the Pennsylvania Bureau of Forestry, Bradford Water Authority, Forest Investment Associates, Generations Forestry, Hancock Forest Management, Landvest, and Kane Hardwoods for field sites. We thank Charles Vandever for assistance with data collection, John Stanovick for his insightful statistical consultations and Scott Stoleson, Todd Ristau, Patricia Raymond, Jean-Pierre Tremblay, and Susan Wright for valuable editorial suggestions.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Royo, A.A., Kramer, D.W., Miller, K.V. et al. Spatio-temporal variation in foodscapes modifies deer browsing impact on vegetation. Landscape Ecol 32, 2281–2295 (2017). https://doi.org/10.1007/s10980-017-0568-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-017-0568-x