
Abstract
Context
Jack pine (Pinus banksiana)-dominated ecosystems of northern Lower Michigan are the primary breeding habitat for the federally endangered Kirtland’s warbler (Setophaga kirtlandii, KW). Historically, young stands used by KW were produced by stand-replacing wildfires, but fire suppression has necessitated the management of jack pine plantations for KW habitat since the 1970s. Effects of this long-term management on landscape age heterogeneity have previously not been quantified.
Objectives
We hypothesized that forest management has altered the spatial and temporal distribution of jack pine-dominated ecosystems beyond their historic range of variability.
Methods
By developing a diameter-age relationship for jack pine, we estimated ages of pre-European settlement trees found in General Land Office survey notes. We compared pre-European and current landscapes using geostatistical modeling of survey notes, and landscape metrics to quantify changes in pattern.
Results
Three KW management-based age classes (<20, 21–50, >50 years) are now more evenly distributed (31, 39, and 30 %, respectively) compared to the pre-European distribution (5, 19, 76 %) with little variability over time. Landscape metrics suggest the current landscape is younger and more fragmented than the pre-European landscape. These changes indicate restriction of the historic range of age variability, largely due to conversion of older jack pine stands to young KW habitat plantations.
Conclusions
Management has met KW population objectives, but has altered the temporal variability of the landscape’s age structure. Pre-European settlement patterns of stand-ages may provide a foundation for an ecosystem-based management plan for the region that supports both KW and the ecosystems upon which they depend.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The 1973 Endangered Species Act (ESA) encourages protections and associated management priorities for imperiled species, and has achieved varying levels of success at rehabilitating foundering populations (Stokstad 2005; Schwartz 2008). Explicit management practices focused on species of concern are often necessary to allow quick and decisive intervention (Simberloff 1998), and when successful may be replicated elsewhere to maintain at-risk populations. However, even recovered populations may be conservation-reliant in the long term, requiring considerable resources for habitat management or mitigation of population-limiting variables (Doremus and Pagel 2001; Scott et al. 2010). In many cases a narrowly-focused approach may emerge, resulting in landscape management that focuses mainly on a single imperiled species or population. As a result, management actions intended to accommodate species at risk may reduce the long-term variability of landscapes, concomitantly sacrificing other ecosystem properties such as biodiversity or resilience (Holling and Meffe 1996; Churchill et al. 2013), even while achieving or exceeding management objectives for the species or population of concern.
Forest management that produces heterogeneous landscapes has been proposed to support long-term viability of ecosystems, and can sustainably manage both at-risk species and the landscapes they inhabit (Franklin 1993; Churchill et al. 2013). The historic range of variability (HRV) concept is useful as a guiding principle of modern landscape management, where management objectives seek to identify and emulate the heterogeneity of naturally produced landscapes (Hessburg et al. 1999; Keane et al. 2009). Rather than a single, static condition, HRV has been used as a way to sustainably manage landscapes by encompassing a range of ecosystem characteristics that historically were often spatially and temporally variable, and resulted from interactions of disturbances with abiotic factors and vegetation processes (Landres et al. 1999). Given current, rapidly changing climate conditions, the utility of selecting historically relevant management objectives has been questioned (Millar et al. 2007). However, HRV can enhance overall ecosystem health and sustainability by incorporating heterogeneous conditions that differentially interact with future disturbances or climate extremes such as drought (Holling and Meffe 1996).
Site-specific measures of HRV have demonstrated marked changes to forests caused by management practices (Cyr et al. 2009). Determining the characteristics of historical landscapes is difficult, however, because data describing pre-anthropogenic conditions often do not exist, especially at broad scales (White and Walker 1997; Keane et al. 2009). The HRV of fire-adapted forests, in particular, may be especially difficult to determine from remnant stands due to widespread alteration of fire regimes and suppression (Baker 1992). Broad-scale documentation of pre-European settlement conditions may therefore be the best way to provide essential baseline data to determine reference conditions for comparison with modern managed landscapes (Schulte and Mladenoff 2001).
General Land Office (GLO) public land surveys have been widely used to describe landscapes prior to European settlement, particularly in the Great Lakes region (Whitney 1987; Schulte and Mladenoff 2001; Cleland et al. 2004). Originally conducted for township and property designation, the size and species of marker trees were recorded, and areas of recent disturbance such as wildfire or windthrow were documented (Liu et al. 2011). These data have been used as historic metrics of forest structure including species composition, stand density, tree size distributions, and landscape disturbance patterns (Schulte and Mladenoff 2001; Cleland et al. 2004; Williams and Baker 2011). To our knowledge, GLO notes have not been used to determine the age of historic forests, yet historical stand age distributions determined from recorded tree size data may serve as a foundation for management within the HRV. This is especially true on landscapes managed for species that require specific successional stages for habitat, and where the age of the modern managed landscape is likely inconsistent with the historic age range.
Understanding landscape structure prior to European settlement is particularly relevant in northern Lower Michigan (U.S.A.; hereafter NLM) due to intensive and extensive management for breeding habitat for the federally endangered Kirtland’s warbler (Setophaga kirtlandii Baird; KW). KWs are neotropical migratory songbirds that overwinter in the Bahamas and nest in NLM in large (>32 ha) stands of young (<20 years old) jack pine (Pinus banksiana Lamb.; Walkinshaw 1983). KW habitat historically was maintained by frequent (12–60 years fire return interval; Simard and Blank 1982), stand-replacing wildfires in the region that created a mosaic of dense jack pine stands interspersed with open barrens (Kashian et al. 2003a). This structure provided young trees with low branches to protect KW ground nests, along with foraging opportunities in the open areas. However, due to twentieth century fire suppression, the coverage of naturally regenerated, early-successional jack pine forests was reduced, and KW breeding habitat is currently maintained as large, young, dense jack pine plantations planted in an opposing-wave pattern to incorporate openings (Probst and Weinrich 1993). These plantations, and associated trapping of nest-parasitizing brown-headed cowbirds (Molothrus ater Boddaert), have successfully increased the KW population to more than twice the original management objective, and delisting is likely in the next decade (MDNR et al. 2015).
Management of jack pine plantations by harvesting and reforesting on a 50-year rotation to maintain young stands has likely had a pronounced influence on the landscape pattern of the region, as more than 75,000 ha of public land are managed specifically for KW habitat in NLM (MDNR et al. 2015). Management protocols state that 3830 ac (approx. 1550 ha) of jack pine forests must be available annually as suitably-aged KW habitat, and although the region has been intensely managed in this way for more than 40 years, the potential impacts on landscape age heterogeneity have not been quantitatively assessed. Ecosystem-based management that incorporates ecosystem processes and diversity has recently been identified as central to KW breeding habitat management (Bocetti et al. 2012), thus baseline data that describe historical landscapes are critical for establishing best-practices for future, more holistic management (Corace and Goebel 2010).
To determine the extent that breeding habitat management for KW has altered landscape-scale stand-age distributions in NLM, we described the pre-European settlement age distribution using nonlinear regression techniques that to our knowledge have previously not been used for pre-European survey data. We compared these historic data with the current landscape to answer the following questions:
-
(1)
How well does the current distribution of jack pine-dominated cover types compare with that prior to European settlement?
-
(2)
How does the current stand-age distribution compare to the historical landscape of this region? and;
-
(3)
How does the spatial distribution of stand ages compare between current and historical landscapes?
We predicted that the current, managed landscape would approximate the historical spatial location of jack pine cover types, but that the current landscape would be younger and have a more homogeneous stand-age distribution, as old, wildfire-produced jack pine stands have been replaced by younger plantations for KW habitat.
Methods
Study area
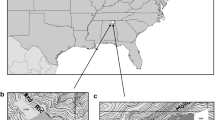
The study area lies mainly within the Grayling Outwash Plain (Sub-subsection VII.2.2) of the Highplains Subsection (VII.2) of the Northern Lacustrine-Influenced Lower Michigan Section (VII) of Region II (NLM; 44°30′N, 84°30′W). This sub-subsection is a broad outwash plain dominated by nutrient-poor, excessively drained sands (Albert 1995; Fig. 1). The vegetation of the study area is dominated by jack pine with northern pin oak (Quercus ellipsoidalis E.J. Hill) and red pine (P. resinosa Sol ex. Aiton) as secondary species (Kashian et al. 2003a). The prevailing disturbance regime in the Highplains subsection was historically high-severity, stand-replacing wildfires with a 59-year rotation (Cleland et al. 2004) or a return interval from 12 to 60 years (Simard and Blank 1982). This disturbance regime created a mosaic of stand ages and forest structures (Leahy and Pregitzer 2003). Although the occurrence of fire has been reduced (Cleland et al. 2004), stand-replacing wildfire is still one of the dominant disturbances in the region (Kashian et al. 2012). Serotinous cones in jack pine allow for rapid post-fire seed dispersal and germination, and facilitate establishment of nearly monotypic stands scattered with shrubby northern pin oak (Whitney 1987). Dense jack pine stands regenerated by fire are interspersed with large openings, or barrens, characterized by grasses and low, woody shrubs typical of dry prairies, such as blueberries (Vaccinium spp.) and sand cherry (Prunus pumila L.; Kashian et al. 2003a). Fire and the dry, acidic soils of the region effectively prevent encroachment of many species that could otherwise out-compete and displace jack pine under more productive conditions (Burns and Honkala 1990). We selected a study area of more than 450,000 ha based on the preponderance of jack pine coverage prior to European settlement (80 % of all jack pine identified in the GLO notes for Lower Michigan; Fig. 1). More than 64 % of the study area falls within the Grayling Outwash Plain sub-subsection, and Kirtland’s warbler management areas (KWMAs) comprise over 72,000 ha in the study area.

The study area used to characterize the pre-European settlement distribution of jack pine forests in northern Lower Michigan (U.S.A.) and the distribution of Michigan Department of Natural Resources land used for cover type analyses. The majority of the study area is encompassed by the Grayling Outwash Plain
Historical versus current jack pine spatial distribution
To determine the extent that modern KW breeding habitat management has changed the distribution of jack pine cover types in NLM, we compared the current reported cover type distribution for the Michigan Department of Natural Resources (MDNR) KWMAs (over 28,600 ha, or approximately 40 %, of the KWMA’s in the study area) with the pre-European distribution of jack pine-dominated cover types within those boundaries. Both plantations and wildfire-origin stands on the current landscape were used for the analyses. Pre-European jack pine coverage was determined using GLO survey data previously digitized and interpreted into cover type polygons by the Michigan Natural Features Inventory (Comer et al. 1995; Austin et al. 1999). Forest cover type categories common to both coverages included jack pine, red pine, and barrens.
Distribution of jack pine stand ages
We used GLO public land survey notes previously digitized into point coverages of surveyed line trees (Maclean and Cleland 2003) to model the pre-European distribution of jack pine stand ages in NLM. The extent of jack pine forests on the pre-European landscape was determined from the GLO point coverage using a two-step process. First, line trees were refined by species to select only jack pine across the study area. Second, to exclude forests where scattered jack pines were present but did not dominate the canopy, the pre-European data were extracted using overlaid mapped, historic land cover type polygons that were dominated by jack pine (Comer et al. 1995; Austin et al. 1999).
GLO survey notes include the species name and diameter at breast height (dbh) of trees, but not tree age. We estimated tree age from dbh using nonlinear regression analysis of data taken from living, naturally-regenerated jack pines (N = 1790 trees) across the study area. We used residual plots to assess the assumption of equal variances, and histograms, skewness and kurtosis, and normal q–q plots to confirm normality of the randomly-sampled response variables (Whitlock and Schluter 2009) and found no violations of these assumptions. Tree age (determined from increment cores) and diameter data were fit to a Michaelis–Menten function, which was then applied to tree diameters in the GLO notes to determine the age of 6847 surveyed trees on the historic landscape. Given the extended time period over which surveys were conducted in the region (1837–1858), tree ages were adjusted to the age they would have been in 1858, the latest year surveys were conducted in the region, to provide a standardized dataset. While this technique increased the age of the surveyed landscape, most of the data points (57 %) required fewer than 10 years added to their age because most of the area was surveyed after 1848, and the standardized dataset could more accurately reflect the overall age distribution of the landscape. Additionally, areas noted by surveyors as “burned” or “recently burned” were delimited and added as young forest (Austin et al. 1999). These polygons were not associated with specific survey years in our data, and polygons were therefore added to the pre-European surface as points arbitrarily assigned an age of three years in order to remove zero values from the data set.
To transform the grid of pre-European data points into polygons to map stand ages, areas between data points (average distance = 455 m) were interpolated using empirical Bayesian kriging (EBK) in ArcMap 10.2 (ESRI 2011). Kriging uses known spatial data to estimate unknown data through semivariogram analyses of the known values. EBK differs from traditional kriging methods in that it simulates a number of semivariograms to determine the best fit for the data, thus incorporating uncertainty of the estimated semivariogram model into the modeling process (Krivoruchko 2012). While spatial autocorrelation is necessary for the process of kriging, somewhat nonstationary data, such as individual tree data points, may be modeled robustly in EBK using local subsets of data during the process (Krivoruchko 2012). We confirmed the assumptions of spatial autocorrelation, moderate stationarity, and absence of directional trends using semivariogram clouds, Voronoi maps, and the trend analysis tool in ArcMap, respectively (Krivoruchko 2012), and found no violations of these assumptions. We then used EBK with 100 semivariogram simulations per subset, a subset size of 100 points, and a prediction output surface of polygons to model the pre-European landscape. This process provided a description of the age of the pre-European landscape across an approximately 450,000 ha region.
During the modeling process in ArcMap, validation points are created by the program by systematically removing individual points, modeling without the point, then determining the value at that point. After confirming the normality of the paired differences using a normal q–q plot and finding no violations of normality (Whitlock and Schluter 2009), we tested input survey data and the model-generated validation points for differences using a paired t test. Accuracy of the kriged prediction surface was estimated using classification error matrices of omission and commission of the same points. The overall percent accuracy of each of the matrices was calculated, as well as the coefficient of agreement, kappa. Kappa is a measure of the similarity of matrices, and assumes independent, paired measurements organized into symmetric categorical matrices (Landis and Koch 1977). The statistic ranges between (−1) and (1), with (−1) having complete disagreement, and (1) having complete agreement; (0) indicates the amount of agreement expected due to pure chance (Landis and Koch 1977; Bonham-Carter 2014). While the assumption of independent samples is not strictly met in this case, as moderate spatial autocorrelation exists, these measures are still useful as investigations of the correlation between maps (Bonham-Carter 2014).
Current forest species and age distributions on public lands managed for KW were provided in the form of stand-age polygon maps by management agencies operating in NLM: MDNR, U.S. Forest Service, and U.S. Fish and Wildlife Service. We clipped regions overlaid by current KWMAs from the kriged pre-European coverage, allowing for comparison of discrete areas of historic and current land cover. We divided stand ages across both landscapes into three categories based on their role in the current management regime: stands <20 years old (young, generally KW occupied), 21–50 years old (intermediate, abandoned by KW but remain within the management rotation), and >50 years old (mature, stands awaiting harvesting and planting to produce new plantation habitat). The percent cover of each age class was determined for both coverages, and the mean ages of a subsample of 1395 randomly-chosen points from each coverage were compared using the non-parametric Wilcoxon signed-rank test; the data display moderate non-normality (leptokurtosis), but lack skewness, thus making the Wilcoxon test appropriate (using R moments package; R Core Team 2014; Whitlock and Schluter 2009). All statistical analyses were performed with alpha = 0.05, and using R (v. 3.1.1; R Core Team 2014) and SPSS (v. 22; IBM Corp. 2013).
Landscape patterns of stand ages
Age class- and landscape-level metrics for both the current and pre-European landscapes were calculated from raster maps exported at a resolution of 15.24 m (50 ft. using the State Plane projection) using FRAGSTATS v3.3 (McGarigal et al. 2002). We calculated the proportion of the landscape, number of patches, largest patch index (the percent of the landscape composed of the largest patch in that class), and Euclidean nearest neighbor distance for each age class on both landscapes. Landscape-scale metrics calculated for each landscape were: number of patches, mean patch size, patch contagion (a measure of patch connectedness), and interspersion and juxtaposition index (to describe the adjacency of patches of different types; McGarigal et al. 2002).
Results
Jack pine spatial distribution
The current distribution of KWMAs in NLM is consistent with pre-European jack pine-dominated cover types. Greater than 99 % of the area within KWMAs is located on landforms where jack pine-dominated ecosystems occurred prior to European settlement (glacial outwash plains). On MDNR land, the proportion of jack pine-dominated cover types has increased by approximately 6 %, from 27,203 ha on the pre-European settlement landscape to 28,963 ha currently. The other major cover types on the pre-European landscape have decreased in the KWMAs, indicating homogenization of the landscape over time; red pine decreased 29 % from 3816 to 2707 ha, and barrens have been reduced by approximately 67 % (9650 to 3188 ha) from their pre-European distribution.
Determination of jack pine stand ages
The ages of field-sampled trees ranged from 5 to 105 years, and diameters ranged from 0.2 to 50.5 cm (Y = (289.43x)/(83.24 + x)). The coefficient of determination between tree age and diameter was high for all 1790 trees (R 2 = 0.81, p < 0.001), although variation of diameter at a given age increased with stand age (Fig. 2). These trends suggest that size inequality, or size hierarchies, for jack pine are more common in older stands (Kenkel et al. 1997), and thus prediction of tree age from diameter is most accurate for younger stands.

Age and diameter at breast height (dbh) of field-collected data, showing Michaelis–Menten regression line (Y = (289.43x)/(83.24 + x)) and 95 % CI (R 2 = 0.808, p < 0.001). The age data ranged from 5 to 100 years. Pre-European settlement data derived from the equation ranged from 3 to 161 years
Differences in means between the original input data and the validation points predicted from the model were not statistically significant. Kappa statistics and error matrices showed moderate agreement between the data sets and overall accuracy of the kriged values (see Landis and Koch 1977). We constructed two matrices using the three management-based age classes (<20, 21–50, >50 years; Table 1a) and also using five, equal-interval age classes to more precisely determine age classes with the most error (Table 1b). The model tends to under-predict the age of the pre-European forest in the 3-class scenario, and to over-predict the age in the 5-class scenario. The 3-class configuration kappa was 0.494 (T = 54.5, p < 0.001), 95 % CI (0.476, 0.512), and overall accuracy was 69.8 %. The 5-class configuration showed kappa was 0.516 (T = 77.5, p < 0.001), 95 % CI (0.500, 0.532), and overall accuracy was 63.0 %. The 3-class matrix exhibited the most error in the middle (21–50 years) age class, whereas the 5-class matrix displayed a more uniform distribution of disagreement (Table 1a, b). Error in the 21–50 years age class is likely a result of a lack of data between 21 and 40 years, because most data (89 %) in the broader 21–50 years old class is >40 years.
Comparison of the proportions of the landscape in the three, management-based age classes within existing managed areas shows significant changes to the distribution (Wilcoxon signed-rank test; V = 652382.5, p < 0.001). The pre-European distribution had 5 % of the landscape in the youngest age class, 19 % in the intermediate age class, and 76 % in the mature age class; the current landscape, in comparison, contains 31, 39, and 30 % in the three classes, respectively (Fig. 3a). Using equal, 20-year-interval age classes to more closely examine the distribution of ages, the pre-European landscape was dominated by the 61–80 year age class (47 %; Fig. 3b). Stands 0–20 years represented 5 %, stands 21–40 years 10 %, 41–60 years 36 %, and stands > 80 years represented 2 % of the landscape.

Percent cover of pre-European settlement and current jack pine stand-age classes within current Kirtland’s warbler management areas expressed as a three management-based age classes, and b 20-year age intervals. There is a marked decrease in the coverage of older (>50 years) stand ages on the current landscape
Landscape patterns of stand ages
The spatial arrangement and configuration of age classes within KWMAs differed markedly between pre-European and current landscapes (Fig. 4a, b). As a whole, the current landscape is much more fragmented than the pre-European landscape, with over twice as many patches (2029 current, 999 pre-settlement), half the mean patch size (35.8, 71.3 ha), lower aggregation of stands within an age class (contagion = 47.3, 68 %), and much higher interspersion of patches (interspersion and juxtaposition index = 94.1, 62.1 %). When analyzed by age class, metrics show a pronounced change in the distribution of the three age classes between the two time periods (Table 2). The youngest age class currently has a much larger mean patch size, consistent with the establishment of large plantations in KWMAs. While the intermediate age class has exhibited the least change in mean patch size, the increase in largest patch index indicates the presence of very large patches on the modern landscape. The extent of the mature age class (which is being replaced by plantations in the current management plan) has been reduced by 60 % from pre-European settlement, has had a 74 % reduction in mean patch size, and has exhibited a 15-fold reduction in largest patch index, suggesting the presence of very large, mature patches on the historical landscape that are no longer present.

Distribution of jack pine stand age classes in Kirtland’s warbler management areas on a pre-European settlement and b current landscapes. The extent represents Michigan Department of Natural Resources, USDA Forest Service, and USDI Fish and Wildlife Service management areas in northern Lower Michigan (U.S.A.)
Discussion
Our analyses suggest that while the spatial locations of KWMAs in northern Lower Michigan are consistent with those of pre-European settlement jack pine-dominated cover types, the stand age distribution of jack pine forests on the current landscape differs significantly (Fig. 3). Comparatively, the current landscape is characterized by widespread reductions of mature jack pine stands and an even distribution of stands among age classes (Table 2). Assuming the landscape during the pre-European survey period (1837–1858) to be within the HRV, the current management regime of harvesting and planting jack pine on a 50-year rotation has greatly reduced the landscape-scale variability of stand ages. This supports other conclusions that extensive plantation management for KW breeding habitat is likely to homogenize the landscape (Spaulding and Rothstein 2009). Given that the management regime has resulted in a near replacement of wildfire-regenerated stands by plantations on public lands in the region (Donner et al. 2008), the effects of KW management on the landscape may be extensive.
While our study focuses on landscape structure rather than the structure of individual stands, it has been shown that plantations at young and intermediate ages differ significantly from wildfire-regenerated stands in important attributes such as the variability in stem density among stands, internal patchiness, the number of snags, the amount of coarse woody debris, and forest floor biomass (Spaulding and Rothstein 2009). These important stand structural components begin to resemble those of wildfire-regenerated stands after approximately 40 years, but mature stands > 50 years have been significantly reduced (Figs. 3, 4) on the current landscape. Mature jack pine forests in the region are important for ecosystem services such as carbon storage and nutrient accumulation (Rothstein et al. 2004; LeDuc and Rothstein 2010), and may affect biodiversity of both bird (Corace et al. 2010, 2016) and plant communities (Kashian et al. 2003b). Together with these results, our data suggest that KW habitat management using a short rotation age and targeted removal of stands >50 years old has created a landscape that at multiple spatial scales departs markedly from that resulting from the natural disturbance regime in NLM.
Despite dominance by a short-lived tree species and a frequent, stand-replacing fire regime (Simard and Blank 1982; Cleland et al. 2004), our analyses suggest that the pre-European landscape was not dominated by early-successional forests during the period of the GLO surveys. We acknowledge that the 20-year period of the survey represents only a short period of time, but we suggest that the landscape is perhaps better characterized as dynamic, or temporally variable, rather than strictly early-successional. The dominance of the landscape by older age classes may have resulted from a single large fire or a series of fires in close succession, but the landscape was clearly dominated by young stands just a few decades prior to the GLO surveys. Thus, the pre-European stand age distribution likely varied through time rather than remaining at any one age distribution or successional stage, and that variation may have been important for maintaining ecosystem processes and biodiversity over long temporal scales (Petraitis et al. 1989). This temporal variability is lacking on the current landscape because of consistent annual harvesting and planting that maintains a relatively constant, or slowly increasing, proportion of young stands. While a 50-year harvesting rotation may approximate the historical 59-year fire rotation (Cleland et al. 2004), annual and decadal variability in the area burned, with much area burned in some years but little or none in others, is equally important for re-creating the stand age HRV. Fire rotations are determined by the overall amount of burned land over time, but do not necessarily indicate that the entire landscape has burned within that time frame. Overlapping fires in the same locations (perhaps driven by locally shorter rotations at smaller scales) would lead to variably-burned landscapes, with some areas remaining unburned and others remaining relatively young for long periods of time. Current management practices have modified the age distribution of large swaths of the landscape, without the variability in age that would likely characterize a landscape produced by a historically-relevant fire regime. Although the current proportion of older stands on the landscape may be within the HRV when large fire years are considered, harvesting and planting an equal area of jack pine each year will dampen and eventually eliminate the variability of stand ages over time (Fig. 5).

Conceptual representation of the historic range of jack pine stand-age variability in northern Lower Michigan (U.S.A.), illustrating the effects of warbler habitat management on the temporal variability of stand age-distributions. “KW” indicates the beginning of Kirtland’s warbler habitat management using plantations. Representation of landscape age prior to warbler management is hypothetical based on the periodic occurrence of large wildfires of variable size and timing on a 59-year rotation
Our analyses further suggest that the spatial configuration of stand ages in current jack pine forests has been affected by prolonged KW breeding habitat management. Patches of young forests are much larger, on average, than during the pre-European period (Table 2). This suggests that the current distribution of large plantations does not reflect the variability of a natural fire regime, where a range of fire sizes over time would likely yield some large stands but also many smaller stands. Conversely, the average sizes of intermediate and, more dramatically, mature patches have been reduced over this time period (Table 2). Increasing smaller patches on a landscape results in the loss of forest interior areas, and could have implications for wildlife or plant communities that depend upon such sites (White and Host 2008; Cullinane-Anthony et al. 2014). This observation is reinforced by the reduction in landscape contagion over this time period, suggesting that similarly-aged patches are overall less aggregated into large stands, even as the size of young stands has increased. The current management protocol recommends plantations of at least 121 ha (300 ac), but preferably 202 ha (500 ac), for the provision of KW breeding habitat (MDNR et al. 2015); this protocol will likely continue to increase the mean patch size of young forests, reduce the size of older forests as more old stands are converted to plantations, and further reduce the variability of stand sizes. Thus, in addition to a homogenization of stand-level characteristics (Spaulding and Rothstein 2009), our data suggest that landscape composition and structure have been homogenized by KW breeding habitat management in terms of the extent and configuration of stand ages.
We acknowledge that limitations exist in the use of GLO notes to characterize pre-European settlement landscapes, such as surveyor biases in tree species and size selection (Manies et al. 2001), and uncertainty in the quality of data recorded by individual surveyors who were not trained botanists or ecologists (Schulte and Mladenoff 2001). However, relatively homogenous site conditions consisting of nearly-monotypic jack pine stands persist across much of the Grayling Sub-subsection, such that gross misidentification errors were likely minimal. Bias against using jack pines as marker trees have been reported for GLO notes in Wisconsin, suggesting that surveyors chose trees of larger, more easily marked species where possible (Liu et al. 2011). Although such a bias would underreport the presence of jack pines in a given area, our focus on monotypic jack pine stands, rather than mixed-pine stands, reduces the importance of this bias for our results. Further, our exclusive use of surveyed line trees reduces the risk of biases associated with blazing, as these trees were not required to be marked (Liu et al. 2011).
Of greater concern is the lack of small, young jack pine in the survey notes, which fell beneath the minimum diameter for measurement along section lines; 3 in. (7.6 cm) dbh was the smallest reported in our dataset (Manies et al. 2001). Our representation of surveyed, recently burned areas as young stands (3 years old) added young forest to the landscape to ensure representation of young stands that were described by surveyors but would not be otherwise captured by GLO data. The distinct lack of trees in the 21–40 years age class (Table 1b) is associated solely with the survey data, and demonstrates the paucity of young trees in the survey notes. The misinterpretation of the 21–50 years age class by the model (Table 1a), which inaccurately classified most of the trees in the age class, is likely because the majority of the surveyed trees in that class (89 %) were older than 40 years (Table 1b), causing even relatively small errors in the model estimates to frequently fall above the upper limit of the 21–50 years age class. While this sampled age distribution may have implications for the modeling process, it also likely represents a lack of young stands on the actual pre-European landscape. Overall, the frequent, high-severity fire regime in NLM created a landscape composed chiefly of patches of even-aged stands. Such a landscape would have offered little size variation at scales small enough to warrant much bias in surveyor preference for age or size. Wildfire-produced stands in the region often include unburned living trees within burned areas (Kashian et al. 2012), but the number of data points that were surveyed as old trees cannot be accounted for by this heterogeneity alone. While uncertainty exists, the relative simplicity of the study area and the considerations made to add young trees likely ameliorate many of the effects of that uncertainty.
It is increasingly recognized that variability in landscape structure and successional stage is intimately associated with ecosystem function and sustainability (Franklin 1993; Lindenmayer et al. 2000). Landscape heterogeneity is important for maintaining both resistance and resilience to disturbances (Churchill et al. 2013; Moritz et al. 2014; Seidl et al. 2014), and components of stand structure such as complexity and heterogeneity have been associated with increased landscape biodiversity (Lindenmayer et al. 2000). Therefore, historically-relevant variability may be vital for the future sustainability of managed ecosystems (Grumbine 1994; Keane et al. 2009). The efficacy of managing for HRV objectives has been questioned given the overwhelming effects of global climate change on ecosystems (Millar et al. 2007); indeed there is much uncertainty about the effects of climate change in the region (Handler et al. 2014). Given this uncertainty, restoration of historic heterogeneity may be a way to create more robust landscapes that are resilient given the ecosystem properties that are known and documented, while allowing for adaptive management practices and future research and projections of climate effects to shape management practices going forward (Keane et al. 2009).
In NLM in particular, restoration of historically relevant patterns and processes may aid management both ecologically and financially; current management is not self-sustaining in that young jack pine scheduled for harvest under the current protocol has little economic value. Although jack pine is an important pulpwood species, there has been little pressure to harvest for that purpose due to protection of the area as KW breeding habitat under the ESA. As KW delisting approaches, there may be an increase in harvesting for pulpwood or a burgeoning biomass market, though KW habitat protection will continue and interagency agreements have been made to restore or increase more traditional KW management should the population begin to fall (MDNR et al. 2015). Future management will be overseen by a conservation team comprised of management practitioners from multiple agencies and other interested parties, and non-profit fundraising is already under way (MDNR et al. 2015). Such efforts can be facilitated by restoring historic ecosystem heterogeneity across several properties (age, structure, disturbance processes, etc.) to provide a stable and sustainable ecosystem.
This study highlights a need to reconcile endangered species management with the heterogeneity created by natural disturbance regimes in systems intensively managed for very specific objectives. In the case of KW, there exists a complex tradeoff between the recovery and maintenance of an endangered species and the conservation and restoration of a forested landscape within the historical range of age variability, as represented by the relatively old stand-ages of the survey period. We have shown that the spatial and temporal variability of the age distribution of jack pine forests has been homogenized by decades of intensive and extensive management, resulting in a fixed, young landscape that has little historically-consistent age variability. As the overwhelming success of KW recovery moves it towards removal from ESA listing, there is an opportunity to re-evaluate the management regime of the region while maintaining the knowledge gained from a highly successful recovery program. Management that better emulates historic spatial and temporal age variability may provide a foundation for a more ecosystem-based management plan moving forward that supports both the KW population and the viability of jack pine-dominated ecosystems in the region.
References
Albert DA (1995) Regional landscape ecosystems of Michigan, Minnesota and Wisconsin: a working map and classification. General Technical Report NC-178. St. Paul, MN: UDSA-FS, North Central Forest Experiment Station
Austin MB, Leibfried TR, Korroch KM (1999) Land use circa 1800 [Digital map]. Michigan Natural Features Inventory, Lansing
Baker WL (1992) Effects of settlement and fire suppression on landscape structure. Ecology 73:1879–1887
Bocetti CI, Goble DD, Scott JM (2012) Using conservation management agreements to secure post-recovery perpetuation of conservation-reliant species: the Kirtland’s Warbler as a case study. BioScience 62:874–879
Bonham-Carter GF (2014) Geographic information systems for geoscientists: modelling with GIS. Elsevier Science, Amsterdam
Burns RM, Honkala BH (1990) Silvics of North America. Department of Agriculture, Forest Service, U.S. G.P.O, Washington
Churchill DJ, Larson AJ, Dahlgreen MC, Franklin JF, Hessburg PF, Lutz JA (2013) Restoring forest resilience: from reference spatial patterns to silvicultural prescriptions and monitoring. For Ecol Manag 291:442–457
Cleland DT, Crow TR, Saunders SC, Dickmann DI, Maclean AL, Jordan JK, Watson RL, Sloan AM, Brosofske KD (2004) Characterizing historical and modern fire regimes in Michigan (USA): a landscape ecosystem approach. Landscape Ecol 19:311–325
Comer PJ, Albert DA, Corner RA, Wells HA, Hart BL, Raab JB, Price DL, Kashian DM (1995) Michigan’s presettlement vegetation, as interpreted from the General Land Office surveys 1816–1856. Michigan Natural Features Inventory, Lansing. Digital map
Corace RG III, Goebel PC (2010) An ecological approach to forest management for wildlife: integrating disturbance ecology patterns into silvicultural treatments. Wildl Profess Spring, pp 38–40
Corace RG III, Korte JL, Shartell LM, Kashian DM (2016) Upland sandpiper: a flagship for jack pine barrens restoration in the upper Midwest? Ecol Restor 34:1
Corace RG III, Seefelt NE, Goebel PC, Shaw HL (2010) Snag longevity and decay class development in a recent jack pine clearcut in Michigan. North J Appl For 27:125–131
Cullinane-Anthony BL, Seefelt NE, Corace RG III, Kashian DM, Gehring TM (2014) Influence of residual forest patches on post-fire bird diversity patterns in jack pine-dominated ecosystems of northern Lower Michigan. For Ecol Manag 331:93–103
Cyr D, Gauthier S, Bergeron Y, Carcaillet C (2009) Forest management is driving the eastern North American boreal forest outside its natural range of variability. Front Ecol Environ 7:519–524
Donner DM, Probst JR, Ribic CA (2008) Influence of habitat amount, arrangement, and use on population trend estimates of male Kirtland’s warblers. Landscape Ecol 23:467–480
Doremus H, Pagel JE (2001) Why listing may be forever: perspectives on delisting under the U.S. endangered species act. Conserv Biol 15:1258–1268
ESRI (2011) ArcGIS desktop: release 10.2. Environmental Systems Research Institute, Redlands
Franklin JF (1993) Preserving biodiversity: species, ecosystems, or landscapes? Ecol Appl 3:202–205
Grumbine RE (1994) What is ecosystem management? Conserv Biol 8:27–38
Handler S, Duveneck MJ, Iverson L, Peters E, Scheller RM, Wythers KR, Brandt L, Butler P, Janowiak M, Shannon PD, Swanston C (2014) Michigan forest ecosystem vulnerability assessment and synthesis: a report from the Northwoods Climate Change Response Framework project. Gen. Tech. Rep. NRS-129. Newtown Square, PA: U.S. Department of Agriculture, Forest Service, Northern Research Station
Hessburg PF, Smith BG, Salter RB (1999) Detecting change in forest spatial patterns from reference conditions. Ecol Appl 9:1232–1252
Holling CS, Meffe GK (1996) Command and control and the pathology of natural resource management. Conserv Biol 10:328–337
IBM Corp. Released 2013. IBM SPSS Statistics for Windows, Version 22.0. Armonk, NY: IBM Corporation
Kashian DM, Barnes BV, Walker WS (2003a) Landscape ecosystems of northern Lower Michigan and the occurrence and management of the Kirtland’s warbler. For Sci 49:140–159
Kashian DM, Barnes BV, Walker WS (2003b) Ecological species groups of landform-level ecosystems dominated by jack pine in northern Lower Michigan, USA. Plant Ecol 166:75–91
Kashian DM, Corace RG III, Shartell LM, Donner DM, Huber PW (2012) Variability and persistence of post-fire biological legacies in jack pine-dominated ecosystems of northern Lower Michigan. For Ecol Manag 263:148–158
Keane RE, Hessburg PF, Landres PB, Swanson FJ (2009) The use of historical range and variability (HRV) in landscape management. For Ecol Manag 258:1025–1037
Kenkel NC, Hendrie ML, Bella IE (1997) A long-term study of Pinus banksiana population dynamics. J Veg Sci 8:241–254
Krivoruchko K (2012) Empirical Bayesian Kriging implemented in arc GIS geostatistical analyst. Arc User 15:6–10
Landis JR, Koch GG (1977) The measurement of observer agreement for categorical data. Biometrics 33:159–174
Landres PB, Morgan P, Swanson FJ (1999) Overview of the use of natural variability concepts in managing ecological systems. Ecol Appl 9:1179–1188
Leahy MJ, Pregitzer KS (2003) A comparison of presettlement and present-day forests in northeastern Lower Michigan. Am Midl Nat 149:71–89
LeDuc SD, Rothstein DE (2010) Plant-available organic and mineral nitrogen shift in dominance with forest stand age. Ecology 91:708–720
Lindenmayer DB, Margules CR, Botkin DB (2000) Indicators of biodiversity for ecologically sustainable forest management. Conserv Biol 14:941–950
Liu F, Mladenoff DJ, Keuler NS, Moore LS (2011) Broadscale variability in tree data of the historical Public Land Survey and its consequences for ecological studies. Ecol Monogr 81:259–275
Maclean AL, Cleland DT (2003) Determining the spatial extent of historical fires with geostatistics in northern Lower Michigan. In: Fire, fuel treatments, and ecological restoration. Conference proceedings, pp 16–18
Manies KL, Mladenoff DJ, Nordheim EV (2001) Assessing large-scale surveyor variability in the historic forest data of the original U.S. Public Land Survey. Can J For Res 31:1719–1730
McGarigal K, Cushman SA, Neel MC, Ene E (2002) FRAGSTATS v3: computer software program produced by the authors at the University of Massachusetts, Amherst. http://www.umass.edu/landeco/research/fragstats/fragstats.html
Michigan Department of Natural Resources, U.S. Fish and Wildlife Service, U.S. Forest Service (2015) Kirtland’s Warbler breeding range conservation plan. MI DNR, Lansing
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecol Appl 17:2145–2151
Moritz MA, Batllori E, Bradstock RA, Gill AM, Handmer J, Hessburg PF, Leonard J, McCaffrey S, Odion DC, Schoennagel T, Syphard AD (2014) Learning to coexist with wildfire. Nature 515:58–66
Petraitis PS, Latham RE, Niesenbaum RA (1989) The maintenance of species diversity by disturbance. Q Rev Biol 64:393–418
Probst JR, Weinrich J (1993) Relating Kirtland’s warbler population to changing landscape composition and structure. Landscape Ecol 8:257–271
R Core Team R: a language and environment for statistical computing. Austria. https://cran.r-project.org/
Rothstein DE, Yermakov Z, Buell AL (2004) Loss and recovery of ecosystem carbon pools following stand-replacing wildfire in Michigan jack pine forests. Can J For Res 34:1908–1918
Schulte LA, Mladenoff DJ (2001) The original US public land survey records: their use and limitations in reconstructing presettlement vegetation. J For 99:5–10
Schwartz MW (2008) The performance of the endangered species act. Annu Rev Ecol Evol Syst 39:279–299
Scott JM, Goble DD, Haines AM, Wiens JA, Neel MC (2010) Conservation-reliant species and the future of conservation. Conserv Lett 3:91–97
Seidl R, Rammer W, Spies TA (2014) Disturbance legacies increase the resilience of forest ecosystem structure, composition, and functioning. Ecol Appl 24:2063–2077
Simard AJ, Blank RW (1982) Fire history of a Michigan jack pine forest. Mich Acad 15:59–71
Simberloff D (1998) Flagships, umbrellas, and keystones: is single-species management passé in the landscape era? Biol Conserv 83:247–257
Spaulding SE, Rothstein DE (2009) How well does Kirtland’s warbler management emulate the effects of natural disturbance on stand structure in Michigan jack pine forests? For Ecol Manag 258:2609–2618
Stokstad E (2005) What’s wrong with the endangered species act? Science 309:2150–2152
Walkinshaw LH (1983) Kirtland’s warbler: the natural history of an endangered species. Cranbrook Institute of Science, Bloomfield Hills
White MA, Host GE (2008) Forest disturbance frequency and patch structure from pre-European settlement to present in the Mixed Forest Province of Minnesota, USA. Can J For Res 38:2212–2226
White PS, Walker JL (1997) Approximating nature’s variation: selecting and using reference information in restoration ecology. Restor Ecol 5:338–349
Whitlock MC, Schluter D (2009) The analysis of biological data. Roberts and Company, Greenwood Village
Whitney GG (1987) An ecological history of the Great Lakes forest of Michigan. J Ecol 75:667–684
Williams MA, Baker WL (2011) Testing the accuracy of new methods for reconstructing historical structure of forest landscapes using GLO survey data. Ecol Monogr 81:63–88
Acknowledgments
We wish to acknowledge the assistance of several people instrumental in procuring data for this research: T. Greco, Michigan Department of Natural Resources; P. Huber and L. McNichols, US Forest Service; D. Rothstein, Michigan State University; and E. Schools, Michigan Natural Features Inventory. We also thank the Kirtland’s Warbler Recovery Team for helpful feedback. T. Dowling, D.R. Kashian, D. Rothstein, and two anonymous reviewers made helpful comments on earlier versions of this manuscript, and P. Huber and S. Sjogren provided constructive feedback in the earliest stages of this study. This project was funded with a grant from the Joint Fire Science Program to R.G.C. III and D.M.K. The findings and conclusions of this article are those of the authors and do not necessarily represent the views of the U. S. Fish and Wildlife Service or other agencies and institutions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Tucker, M.M., Corace, R.G., Cleland, D.T. et al. Long-term effects of managing for an endangered songbird on the heterogeneity of a fire-prone landscape. Landscape Ecol 31, 2445–2458 (2016). https://doi.org/10.1007/s10980-016-0413-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-016-0413-7