
Abstract
The vascular endothelial growth factor (VEGF) is an essential factor to pathologic angiogenesis. Disruption of VEGF/VEGF receptor interaction in cancer patients inhibits the development of new and pre-existing tumor blood vessels. Consequently, VEGF becomes an important therapeutic target for handling solid tumors. In this work, human VEGF was produced in the culture supernatant of SiHa cells transduced with a replication-defective adenoviral vector (pAdhVEGF121) encoding this molecule. The 35 kDa VEGF121 homodimer was obtained from clarified culture media as a glycosylated protein. VEGF121 expression levels were strictly dependent on the adenoviral viral load used. VEGF121 was produced with purity over 98% after a single step chromatography by immobilized metal affinity chromatography. Additionally, VEGF121 binds Bevacizumab antibody with a KD of 7 nM. Biological characterization by mitogenic assay in HUVEC and ECV-304 cells showed that VEGF121 stimulates cell proliferation in a dose-dependent manner in both cells. Finally, the neovascularization activity of VEGF121 was demonstrated by vascular permeability assays in matrigel plug-bearing mice, showing significantly increased vasculature leakage after treatment with VEGF121. Consequently, transduction of SiHa cells with adenovirus is a suitable alternative for manufacture heterologous proteins of therapeutic interest.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The vascular endothelial growth factor-A (VEGF or VEGF-A) and its receptors play a fundamental role in angiogenesis. VEGF is a 40 kDa glycoprotein that is secreted to the extracellular compartment. The structure of VEGF is homodimeric, formed by two disulfide-linked monomers arranged in an antiparallel direction, displaying two binding sites at each extreme of the molecule [1]. In mammals, VEGF belongs to a family of five members: VEGF-A, VEGF-B, VEGF-C, VEGF-D, and PlGF (Placental growth factor). The biological functions of VEGF proteins are mediated by a family of tyrosine kinase receptors, known as the VEGF receptors (VEGFR) [1,2,3]. The human VEGF-A gene is arranged in 8 exons and is expressed as multiple isoforms due to differential splicing. The four principal isoforms describe are hVEGF121, hVEGF165, hVEGF189 and hVEGF206 [4,5,6,7]. VEGF121 lacks exons 6 and 7 responsible for the heparin-binding domain and freely diffuse through the extracellular matrix [5].
VEGF is a key regulator of vasculogenesis and angiogenesis during embryogenesis [8, 9]. However, in adults, physiological angiogenesis occurs only during wounds healing and the menstrual cycle [10,11,12]. When VEGF regulatory control is lost, angiogenesis contributes to numerous pathological conditions, such as rheumatoid arthritis [13] psoriasis [14], atherosclerosis [15], macular degeneration [16] and tumorigenesis [17].
VEGF induces cell proliferation and promotes survival and migration of endothelial cells. These biological effects are essential for tumorigenesis as angiogenesis is required for tumor growth and progression [17,18,19,20]. Cancer cells secrete VEGF promoting new blood vessels formation for nutrient and oxygen supply [21,22,23] Particularly, VEGF121 isoforms overexpression has been detected in colorectal cancer and in a cancer mouse model VEGF121 overexpression but not VEGF165 overexpression induced tumor growth [24].
The interest of expressing biologically active VEGF121 by recombinant DNA technology lies in the function of this molecule and its different uses: (i) To study and development of new therapies based on inhibiting VEGF/VEGFR interaction [25,26,27,28,29] and (ii) advanced treatments of patients with diabetic foot and ischemic diseases [30,31,32,33,34,35,36].
Recombinant VEGF has been expressed on different systems, including Escherichia coli [37,38,39], yeast [40, 41], insect cells [42] Chinese hamster ovary cells (CHO) [43]. Nevertheless, VEGF purification in these expression systems requires multiple-steps purification protocols increasing the final product cost [37, 44]. VEGF yields obtained on these systems are in the range of mg/l. However, VEGF expression has focused on VEGF165 isoform.
In this work, we expressed VEGF121 in SiHa cells transduced with adenoviral vectors. The use of replication-defective adenoviral vectors for transient gene expression of human proteins in mammal cell cultures is based on the fact that it is an efficient and safe method for gene transfer and protein expression in vitro and in vivo [45,46,47,48,49,50,51]. We analyzed the biological activity of purified VEGF121 based on proliferation and induction of vascular permeability. VEGF121 proliferative properties were analyzed on human umbilical cord endothelial cells (HUVEC) and ECV-304 cell line. The neoangiogenic activity was studied by in vivo murine model of vascular permeability.
2 Materials and Methods
2.1 Generation of Ad-VEGF Adenoviral Vector
The AdEasy vector system was used to generate the replication-defective vector pAdhVEGF121 [52]. Briefly, a synthetic 500 bp sequence encoding human VEGF plus a 6xHis tag was cloned into XhoI/EcoRV sites of the shuttle vector pAdtrack-CMV. The resulting plasmid was transformed into the E. coli BJ5183 strain along with the pAdEasy vector to generate a defective adenoviral vector by in vivo recombination. The resulted plasmid (pAdhVEGF121) has two cytomegaloviruses (CMV) promoters, one for the green fluorescent protein (GFP) and the other for VEGF. The initial viral pool was prepared by transiently transfecting the pAdhVEGF121 plasmid into HEK-293 cells. Adenovirus stocks were further amplified in HEK-293 cells and harvested by a freeze-thaw lysis protocol. Virus concentration was calculated as gene transfer units (GTU) by GFP expression in HEK-293 cells.
2.2 VEGF121 Production in SiHa Cells
SiHa cells were grown in a 24-well plate in DMEM medium supplemented with 10% fetal bovine serum (FBS). After reaching 80% of confluence, cells were infected with the replication-defective adenovirus at the following multiplicity of infection (MOI): 0.5, 10, 20, 40, 80, 160 GTU/cell. Six hours later, media was removed, cells were washed twice with PBS, and 2 ml of DMEM serum- free were added per well. After 72 h, supernatants were harvested, and VEGF expression was analyzed by SDS-PAGE, Western blot with an anti-Histidine antibody (Sigma, USA). VEGF121 were quantified by the NovexR Human VEGF solid-phase sandwich ELISA (ThermoFisher Scientific, EEUU).
2.3 VEGF121 Purification from the Culture Medium
The expressed protein was purified by immobilized metal affinity chromatography (IMAC) using an ÄktaPurifier liquid chromatography system in the following manner. Cell culture supernatant was centrifuged and filtrated through a 0.2 μm filter to remove debris and floating cells. Clarified medium was applied to a 5 ml Ni-Sepharose fast flow column (GE, USA), previously equilibrated with 5 volumes of equilibrium buffer (0.5 mM PMSF, 100 mM NaH2PO4, imidazole 5 mM pH 7.4). After washing with equilibrium buffer containing 100 mM imidazole, the VEGF121 protein was eluted by raising the imidazole concentration to 250 mM. Protein purification was monitored by UV absorption at A280 nm. Each collected fraction was analyzed by 12% SDS-PAGE/Coomassie staining. VEGF121 was detected by Western blot with an anti-His antibody (Sigma, USA). The eluted fraction was dialyzed in 10 mM NaH2PO4, pH 7.4.
2.4 N-Deglycosylation of VEGF121
The purified VEGF121 was deglycosylated by PNGase F (New England Biolabs). VEGF121 samples were mixed with denaturing glycoprotein buffer and incubated 5 min at 100 °C. The denatured VEGF121 were incubated at 37 °C overnight with PNGase F in the buffer provided by the manufacturer and 1% NP40. VEGF was analyzed by SDS-PAGE/Coomassie blue staining and Western blot.
2.5 Anti-VEGF121 Antibody Binding Affinity Measurement
The Monolith NT.115 device (NanoTemper Technologies GmbH, Germany) was used for analyzing binding affinity by microscale thermophoresis (MST). Bevacizumab antibody was labeled with NT-647 probe and used at a concentration of 6 nM. VEGF121 was titrated from 3 pM to 100 nM. The experiments were performed in binding buffer (Tris–HCl 50 mM, pH 7.4, NaCl 150 mM, MgCl2 10 mM and 0.05% Tween-20). Samples were incubated for 1 h at room temperature before loaded into MST hydrophobic capillaries. The data were analyzed with NT Affinity Analysis software.
2.6 In Vitro Proliferation Assay
VEGF121 biological activity was analyzed by MTT proliferation assays in Human umbilical vein endothelial cells (HUVEC) and ECV-304 cells. HUVEC cells were cultured in M-199 medium supplemented with 10% FBS and ECV-304 cells in DMEM medium supplemented with 10% FBS. HUVEC and ECV-304 cells were seeded in 96-well plates at 1 × 104 cells/well and 2 × 104 cells/well, respectively. After 24 h, cells were washed once with PBS and incubated for 6 h in serum-free medium before adding (0, 10, 20, 40, 80, 160, 320, 640 and 1280 ng/ml) of VEGF121 for 48 h. Then, 10 μl of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide at 5 mg/ml was added to each well and placed at 37 ° C for 4 h. Media were removed and formazan crystals were solubilized by adding 100 μl of DMSO. The plate was stirred for 15 min at 37 °C and measured the absorbance at 570 nm. EC50 value was calculated by the GraphPad Prism 6 software.
2.7 In Vivo Neovascularization Assay
VEGF121 was mixed with Matrigel (BD Bioscience, USA) at 30 ng/ml and injected subcutaneously into the laterodorsal abdominal region of female CF1-BALB/c mice of 4 weeks of age. After 6 days, PEG IRDye 800CW contrast agent (LI-COR Biosciences, USA) was injected through the tail vein of Matrigel-VEGF121 carrier mice. 24 h post-injection, the leakage of dye from capillaries was quantified by near-infrared fluorescent imaging (NIRF) using the Pearl® Impulse Small Animal Imaging System (LI-COR Biosciences, USA).
3 Results
3.1 VEGF Expression in Adenoviral-Transduced SiHa Cells
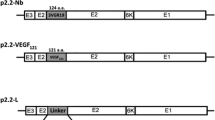
The replication-defective adenoviral vector pAdhVEGF121 was constructed by cloning the synthetic VEGF121 gene in pAdtrack followed by recombination with the pAdEasy vector in E. coli BJ5183 strain (Fig. 1a). pAdTrack-hVEGF and pAdEasy-hVEGF plasmids were digested with PacI to check cloning and recombination events. A single band of 4.5 kbp in pAdEasy-hVEGF digested with PacI indicated correct recombination between pAdTrack-hVEGF and pAdEasy (Fig. 1b). The recombinant viral genomes were transfected into the complementing cell line HEK-293. Replication-defective viruses manifest by comet-like fluorescent plaques around 7 days after transfection (Fig. 1c).

Generation of replication-defective adenoviral vectors. a Construction of the recombinant adenoviral expression vector pAdTrack-hVEGF-A121. A synthetic 500 bp band encoding human VEGF and a 6xHis tag was cloned in pAdtrack-CMV. b Restriction enzyme analysis of pAdTrack-hVEGF-A121 (Lane 1), pAdEasy (Lane 2), and homologous recombination product (Lane 3) with PacI. c pAdhVEGF121 was transfected in HEK-293 cells and GFP expression was visualized by fluorescence microscopy. Comet-like fluorescent plaques became apparent at 7 days (red circle). The microphotographs were acquired with an Olympus IX81 DSU microscope (× 40 magnification)
To determine VEGF121 expression conditions, SiHa cells were infected at MOI of 5, 10, 20, 40, 80 and 160 GTU/ml. Transduction efficiency and cytopathic effect were evaluated by fluorescence microscopy (GFP expression) and phase contrast microscopy, respectively. Seventy-two hours post infection, the number, and fluorescent intensity of cells increased in a GTU dose-dependent manner; however, cytotoxicity and cell death also increased (Fig. 2a). For each MOI tested, we analyzed VEGF121 expression by SDS-PAGE and Western blot, detecting two protein bands, at 17 kDa and at 19 kDa. A linear relationship was also observed between the amount of VEGF121 secreted in the supernatants and the adenovirus load used to infect SiHa cells (Fig. 2b, c). Among 10–80 GTU/cell, the expression levels of VEGF121 underwent an eightfold increase. Therefore, we select 80 UFC/cell for next experiments as the optimal MOI that allows a balance between the fewer cytopathic effect and increased VEGF121 expression.

Viral load effects on cytotoxicity and VEGF121 expression. a SiHa cells were infected at MOI of 0, 10, 20, 40, 80 and 160 GTU/ml. Adenovirus infection resulted in a dose-dependent increase of cytopathic effect and GFP expression. b VEGF121 expression in SiHa cells infected at different MOI. Proteins were separated in 12% SDS-PAGE gel under reducing conditions and immunodetected with an anti-Histidine antibody. At 20 MOI, two bands of VEGF121 were detected, at 17 kDa and 19 kDa. However, at higher viral load VEGF121 overexpression also increases giving rise to a saturation effect that affects the visualization of these 2 bands. c VEGF121 expression levels at different viral load were quantified by NovexR Human VEGF solid-phase sandwich ELISA
3.2 VEGF121 Purification and Biochemical Characterization
The recombinant VEGF121 was purified by IMAC. The chromatogram illustrates the elution profile of VEGF121 on the IMAC column (Fig. 3a). Initial sample, non-binding fraction, washed fraction and eluted fraction were analyzed by 12% SDS-PAGE/Coomassie staining (Fig. 3b). VEGF121 was purified as a homodimer of approximately 35 kDa with an estimated purity of 98% by densitometry with the Image J software. The protein was recognized by anti-poly-histidine antibody (Fig. 3c). 12 mg of purified VEGF121 were obtained from 100 ml of culture medium using this purification procedure.

Purification and characterization of VEGF121. a Chromatogram of immobilized metal affinity chromatography using a Ni-Sepharose Fast Flow column. b SDS-PAGE analysis of samples from the affinity chromatography steps. Lane 1: Culture medium of SiHa cells (starting material); Lane 2: Flow-through; Lane 3: Proteins from the washing step at 100 mM Imidazole; Lane 4: VEGF121 elution at 250 mM Imidazole. Bands were quantified by densitometry to estimate purity. c SDS-PAGE and Western blotting analysis of VEGF121 produced in SiHa cells, respectively. Lane 1: Non-reducing conditions. Lane 2: Reducing conditions. dN-Deglycosylation assay of VEGF121 by treatment with PNGase F enzyme. VEGF121 (90 μg) was untreated (lane 1) or treated with PNGase F overnight (lane 2) at 37°C and analyzed by SDS-PAGE on a 12% gel under reducing conditions followed by Coomassie blue staining and Western blot with an anti-Histidine antibody
The glycosylation state of VEGF121 was analyzed with PNGase F enzyme. We demonstrated that the upper band of 19 kDa detected on Western blots correspond to N-glycosylated forms of VEGF121 monomers, whereas the lower band of 17 kDa contains non-N-glycosylated VEGF121. Additionally, a third immunoreactive band of around 15 kDa was identified (Fig. 3d). Furthermore, it was demonstrated by MST that VEGF121 can bind Bevacizumab antibody with a KD of 7 ± 1 nM (Fig. 4).

The VEGF121/Bevacizumab binding analyze by MST. a MST data for VEGF121 interaction with Bevacizumab. b Binding curve between VEGF121 and Bevacizumab. VEGF121 at concentrations from 3 pM to 100 nM was titrated against 6 nM of Bevacizumab labeled with the fluorescent probe NT-650. MST experiments were performed at a LED power of 60%. Plotting of the change in thermophoresis and concomitant fitting of the data yielded a Kd of 7+ 1 nM. Error bars = s.d.; n = 2
3.3 VEGF121 Induces In Vitro Cell Proliferation in HUVEC and ECV-304 Cells
The biological activity of VEGF121 was studied by its growth-stimulating effect on cells by MTT assays. The effect of recombinant VEGF121 on HUVEC and ECV-304 cells proliferation was dose-dependent (Fig. 5a, b). An increase in VEGF121 concentration caused HUVEC and ECV-304 cells increased proliferation. The half maximal effective concentration (EC50) of VEGF121 to induce cell proliferation was estimated at 11.02 ng/ml and 13.03 ng/ml for HUVEC and ECV-304 cells, respectively.

Cell growth stimulation of HUVEC and ECV-304 cells by VEGF121. HUVEC and ECV-304 cells were treated with increasing amounts of purified VEGF121 (0, 10, 20, 40, 80, 160, 320, 640 and 1280 ng/ml), and cell proliferation was quantified by MTT assay. Error bars = s.d.; n = 3
3.4 VEGF121 Increase In Vivo Neovascularization in Matrigel-Bearing Mice
VEGF121 induced neovascularization was demonstrated by a matrigel plug assay in mice using IRDye® 800CW PEG as a contrast agent to visualize and quantify vascular permeability. NIRF-800 images showed that mice treated with VEGF121 exhibited greater probe accumulation outside the vasculature than the placebo group, as a result of dye extravasation out of the matrigel plug (Fig. 6a, b).

VEGF121-induces vascular permeability by matrigel plug assays.VEGF121 (30 ng/ml) was mixed with matrigel and implanted on the back of CF1-BALB/c mice. The IRDye 800CW PEG probe was i.v injected 6 days later. Vascular permeability was measured by near-infrared fluorescence. a Representative NIRF-800 images of matrigel alone (upper panel) and matrigel-VEGF121-bearing mice (lower panel) 24 h after NIRF dye inoculation. Extensive leakage of the fluorescent probe outside the matrigel plugs was observed on VEG121-bearing mice. b Relative quantification of NIRF fluorescence of matrigel-VEGF121-bearing mice. The statistical analysis was performed according to a t-test. Error bars = s.d; n = 3. Error bars = s.d; n = 3
4 Discussion
VEGF is a glycosylated protein that forms homodimers by interchain disulfide bonds. VEGF121 homodimeric form is essential for its biological activity. VEGF is an asymmetric antiparallel homodimer with two receptor binding interfaces found at each pole of the homodimer. Each of the two binding interfaces must be able to contact a receptor monomer, triggering receptor dimerization and activation [1]. In the present work, we reported the expression of biologically active VEGF121 by SiHa cells transduced with replication-defective adenovirus. We select this cell line because they express the Coxsackievirus/Adenovirus receptor (CAR) needed for effective adenoviral transduction [53,54,55]. Non-reducing SDS-PAGE/Western blot results demonstrated that VEGF121 was secreted to the culture medium mainly as a homodimer with a near molecular weight of 35–37 kDa (58% of total protein, Fig. 3c, Lane 1). This result agrees with the molecular weight of VEGF121 obtained from other sources, including yeast and bacteria [37, 40]. We did not detect monomer forms but higher-molecular-mass complexes (Fig. 3c, Lane 1). Expressions of VEGF121 in Pichia pastoris and baculovirus/insect cells has also resulted in multiple VEGF bands, which was interpreted as covalently linked high-molecular-mass molecules [56, 57]. VEGF isoforms have been obtained in various conformations even in E. coli, with roughly equal proportions of monomeric and dimeric VEGF121 forms [58].
Additionally, we demonstrated that VEGF121 was secreted as a glycosylated protein. After N-deglycosylation with PNGase F enzyme, VEGF121 migrated in SDS-PAGE as a band of around 17 kDa corresponding to the non-N-glycosylated VEGF121 form. However, we detected a second immunoreactive band of approximately 15 kDa. We hypothesize that the 15 kDa band represent the N- and O-non-glycosylated form of VEGF121 and the 17 kDa band represents the N-deglycosylated VEGF121 that still contains O-linked oligosaccharides in two solvent-exposed serine residues (S74 and S95). Therefore, we believe that VEGF121 is secreted as an N- and O-linked glycoprotein. While this hypothesis was not demonstrated, the presence of two accessible serine residues at position 74 and 95 in the molecular surface of VEGF121 homodimer supports this hypothesis.
In this work, we did not perform a comparative analysis between glycosylated VEGF derived from SiHa cells and non-glycosylated variants. There is no evidence that VEGF glycosylation is required for dimerization neither binding to its receptor [59]. However, VEGF glycosylation is essential for molecule secretion; demonstrated by reduced secretion when glycosylation sites were mutated in mouse VEGF164 and human VEGF165 [59, 60]. Regarding its biological activity, it was demonstrated that VEGF glycosylation is not required for mitogenic activity [61]. Besides, in vessel permeability assays, there were no differences between VEGF with mutated glycosylation sites and wild-type VEGF [62].
VEGF121 expressed in the supernatant of SiHa cells was purified by a single step of IMAC chromatography. We obtained 12 mg of VEGF121 from 100 ml of clarified culture media. The single-step purification method reported in this paper is simple, fast, and does not require difficult renaturation methods. VEGF121 was expressed and purified from inclusion bodies in E. coli with high yield and purity [37, 63]. However, this procedure involves denaturation/renaturation steps. Conditions for refolding the denatured proteins to its native functional form are cumbersome and have to be standardized by trial-and-error methods, being time-consuming and expensive for scaling-up [64].
Bevacizumab is a therapeutic humanized antibody that blocks angiogenesis by inhibiting VEGF. This antibody recognizes a conformational epitope [1, 65], and bind to VEGF with a KD of 4.45 nM [66]. By MST, we determine that Bevacizumab binds to the purified VEGF121 with a KD of 7 nM. The slight difference between the KD determined by MST and reported KD for Bevacizumab, seem to be a consequence of different experimental conditions. However, this result confirms that the VEGF121 expressed in SiHa cells transduced with adenoviral vector keep the conformational epitope recognized by Bevacizumab, and suggest that VEGF121 is secreted properly folded.
VEGF has two major biological activities: (a) it is mitogenic to vascular endothelial cells and (b) can induce capillary leakage (vascular permeability) [67]. VEGF stimulates HUVEC proliferation and promotes its migration. Therefore, HUVEC cells are generally used for assaying VEGF biological activity [68, 69]. VEGF121 produced in SiHa cells was able to induce mitogenic responses in HUVEC in a dose-dependent manner. The EC50 of VEGF121 was 11.02 ng/ml, similar to the EC50 reported to VEGF purified from bacteria and yeast [37, 40]. Although HUVEC is the most extensively used endothelial cell type to study endothelial functions [68] and responds to VEGF, these cells are isolated as primary cell culture. Working with primary cultures is complex, and it is only possible to keep them in vitro for short periods of time, besides there are increased risks of microorganisms’ contamination. Thus, working with a cell line [70] that has endothelial properties could replace the use of HUVEC.
ECV-304 cell line was first described as a spontaneously transformed cell line originated from a Japanese human umbilical vein endothelial cell culture. Later, it was found to be derived from the human urinary bladder carcinoma T24 cell line [71]. However, ECV-304 cells have many features of endothelial cells [72], such as increasing cell proliferation in the presence of VEGF [73], a characteristic only observed in endothelial cells but not found in T24 cells [74]. Hence, ECV-304 cells would serve as an angiogenesis model allowing us to characterize endothelial functions [75]. Based on this information, we evaluated the mitogenic activity of VEGF121 on ECV-304 cells. VEGF121 produced in SiHa cells was able to enhance ECV-304 cell proliferation in a cell dose-dependent manner, with an EC50 of 13.03 ng/ml. This result suggests that ECV-304 cell can be used to evaluate the biological activity of VEGF121, in replace of HUVEC primary cell culture.
VEGF induced neovascularization is characterized by immature and highly fenestrated blood vessels. There is a direct relationship between neovascularization and vascular permeability. Classically, neovascularization has been measured using the Miles Assay [76]. This test uses a spectrophotometer to quantify the leakage of a dye from the vasculature to the surrounding tissue, with the limitation of the analysis of a single time point, which must be selected empirically. In this work, we present a modified method to visualize and quantify in real-time the vascular leakage using a fluorescent dextran-based dye. VEGF121 mixed in the matrigel plug injected into mice induced the formation of new and highly fenestrated blood vessels, which trigger the leakage of the fluorescent probe from the vasculature to the plugs. Thus, VEGF121 caused an increase in fluorescence accumulation in matrigel plugs.
5 Conclusions
In this study, we demonstrated functional expression of VEGF121 in the culture media of SiHa cells transduced with a replication-defective adenoviral vector (pAdhVEGF121). VEGF121 expression levels were strictly depended on the viral load used, reaching the maximum expression level at 80 GTU/cell. Expression in SiHa cells resulted in a soluble N-glycosylated dimer of 35–37 kDa, and partial formation of high molecular weight oligomers covalently linked. The purified VEGF121 was recognized by the anti-VEGF antibody Bevacizumab, demonstrating that VEGF121 produced in SiHa cells keep the conformational epitope recognized by Bevacizumab. VEGF121 induced cell proliferation in both HUVEC and ECV-304 cell line. Therefore, ECV-304 cells can replace the primary culture model HUVEC for assaying VEGF bioactivity. Finally, a vascular permeability test with mice carrying matrigel plugs and a fluorescent probe made it possible to determine that purified VEGF121 promotes capillary leakage. In conclusion, we have successfully developed an easy and efficient procedure for VEGF121 expression and purification. This molecule could be used for the evaluation of therapies against angiogenic tumors and wound healing.
References
Muller YA, Li B, Christinger HW, Wells JA, Cunningham BC, de Vos AM (1997) Vascular endothelial growth factor: crystal structure and functional mapping of the kinase domain receptor binding site. Proc Natl Acad Sci USA 94(14):7192–7197
Kowanetz M, Ferrara N (2006) Vascular endothelial growth factor signaling pathways: therapeutic perspective. Clin Cancer Res 12(17):5018–5022. https://doi.org/10.1158/1078-0432.CCR-06-1520
Ferrara N (2000) Vascular endothelial growth factor and the regulation of angiogenesis. Recent Prog Horm Res 55:15–35 (discussion 35–16)
Leung DW, Cachianes G, Kuang WJ, Goeddel DV, Ferrara N (1989) Vascular endothelial growth factor is a secreted angiogenic mitogen. Science 246(4935):1306–1309
Houck KA, Leung DW, Rowland AM, Winer J, Ferrara N (1992) Dual regulation of vascular endothelial growth factor bioavailability by genetic and proteolytic mechanisms. J Biol Chem 267(36):26031–26037
Tischer E, Mitchell R, Hartman T, Silva M, Gospodarowicz D, Fiddes JC, Abraham JA (1991) The human gene for vascular endothelial growth factor. Multiple protein forms are encoded through alternative exon splicing. J Biol Chem 266(18):11947–11954
Vempati P, Popel AS, Mac Gabhann F (2014) Extracellular regulation of VEGF: isoforms, proteolysis, and vascular patterning. Cytokine Growth Factor Rev 25(1):1–19. https://doi.org/10.1016/j.cytogfr.2013.11.002
Ferrara N, Davis-Smyth T (1997) The biology of vascular endothelial growth factor. Endocr Rev 18(1):4–25. https://doi.org/10.1210/edrv.18.1.0287
Yancopoulos GD, Davis S, Gale NW, Rudge JS, Wiegand SJ, Holash J (2000) Vascular-specific growth factors and blood vessel formation. Nature 407(6801):242–248. https://doi.org/10.1038/35025215
Torry DS, Torry RJ (1997) Angiogenesis and the expression of vascular endothelial growth factor in endometrium and placenta. Am J Reprod Immunol 37(1):21–29
Demir R, Yaba A, Huppertz B (2010) Vasculogenesis and angiogenesis in the endometrium during menstrual cycle and implantation. Acta Histochem 112(3):203–214. https://doi.org/10.1016/j.acthis.2009.04.004
Bassett JR, Cairncross KD (1977) Changes in the coronary vascular system following prolonged exposure to stress. Pharmacol Biochem Behav 6(3):311–318
Koch AE (1998) Review: angiogenesis: implications for rheumatoid arthritis. Arthritis Rheum 41(6):951–962
Chua RA, Arbiser JL (2009) The role of angiogenesis in the pathogenesis of psoriasis. Autoimmunity 42(7):574–579
Sluimer JC, Daemen MJ (2009) Novel concepts in atherogenesis: angiogenesis and hypoxia in atherosclerosis. J Pathol 218(1):7–29. https://doi.org/10.1002/path.2518
Ferrara N (2010) Vascular endothelial growth factor and age-related macular degeneration: from basic science to therapy. Nat Med 16(10):1107–1111. https://doi.org/10.1038/nm1010-1107
Folkman J (1971) Tumor angiogenesis: therapeutic implications. N Engl J Med 285(21):1182–1186. https://doi.org/10.1056/NEJM197111182852108
Zetter BR (1998) Angiogenesis and tumor metastasis. Annu Rev Med 49:407–424. https://doi.org/10.1146/annurev.med.49.1.407
Folkman J (2002) Role of angiogenesis in tumor growth and metastasis. Semin Oncol 29(6 Suppl 16):15–18. https://doi.org/10.1053/sonc.2002.37263
Folkman J (1990) What is the evidence that tumors are angiogenesis dependent? J Natl Cancer Inst 82(1):4–6
Shweiki D, Itin A, Soffer D, Keshet E (1992) Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature 359(6398):843–845. https://doi.org/10.1038/359843a0
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144(5):646–674. https://doi.org/10.1016/j.cell.2011.02.013
Viallard C, Larrivee B (2017) Tumor angiogenesis and vascular normalization: alternative therapeutic targets. Angiogenesis 20(4):409–426. https://doi.org/10.1007/s10456-017-9562-9
Kazemi M, Carrer A, Moimas S, Zandona L, Bussani R, Casagranda B, Palmisano S, Prelazzi P, Giacca M, Zentilin L, De Manzini N, Giacca M, Zacchigna S (2016) VEGF121 and VEGF165 differentially promote vessel maturation and tumor growth in mice and humans. Cancer Gene Ther 23(5):125–132. https://doi.org/10.1038/cgt.2016.12
Ferrara N, Damico L, Shams N, Lowman H, Kim R (2006) Development of ranibizumab, an anti-vascular endothelial growth factor antigen binding fragment, as therapy for neovascular age-related macular degeneration. Retina 26(8):859–870. https://doi.org/10.1097/01.iae.0000242842.14624.e7
Kieran MW, Kalluri R, Cho YJ (2012) The VEGF pathway in cancer and disease: responses, resistance, and the path forward. Cold Spring Harbor Perspect Med 2(12):a006593. https://doi.org/10.1101/cshperspect.a006593
Kong DH, Kim MR, Jang JH, Na HJ, Lee S (2017) A review of anti-angiogenic targets for monoclonal antibody cancer therapy. Int J Mol Sci. https://doi.org/10.3390/ijms18081786
Ciombor KK, Berlin J, Chan E (2013) Aflibercept. Clin Cancer Res 19(8):1920–1925. https://doi.org/10.1158/1078-0432.CCR-12-2911
Ebos JM, Kerbel RS (2011) Antiangiogenic therapy: impact on invasion, disease progression, and metastasis. Nat Rev Clin Oncol 8(4):210–221. https://doi.org/10.1038/nrclinonc.2011.21
Adeghate J, Nurulain S, Tekes K, Feher E, Kalasz H, Adeghate E (2017) Novel biological therapies for the treatment of diabetic foot ulcers. Expert Opin Biol Ther 17(8):979–987. https://doi.org/10.1080/14712598.2017.1333596
Shi R, Lian W, Han S, Cao C, Jin Y, Yuan Y, Zhao H, Li M (2018) Nanosphere-mediated co-delivery of VEGF-A and PDGF-B genes for accelerating diabetic foot ulcers healing in rats. Gene Ther 25(6):425–438. https://doi.org/10.1038/s41434-018-0027-6
Tsui HY, Liu YC, Yan X, Lin Y, Xu Y, Tan Q (2017) Combined effects of artificial dermis and vascular endothelial growth factor concentration gradient on wound healing in diabetic porcine model. Growth Factors 35(6):216–224. https://doi.org/10.1080/08977194.2018.1435532
Rico T, Green J, Kirsner RS (2009) Vascular endothelial growth factor delivery via gene therapy for diabetic wounds: first steps. J Invest Dermatol 129(9):2084. https://doi.org/10.1038/jid.2009.235
Koransky ML, Robbins RC, Blau HM (2002) VEGF gene delivery for treatment of ischemic cardiovascular disease. Trends Cardiovasc Med 12(3):108–114
Qiu F, Cao H, Wu Y (2017) A new analytical model for the changes of vascular endothelial growth factor in ischemic diseases. Med Hypotheses 109:73–76. https://doi.org/10.1016/j.mehy.2017.09.024
Crafts TD, Jensen AR, Blocher-Smith EC, Markel TA (2015) Vascular endothelial growth factor: therapeutic possibilities and challenges for the treatment of ischemia. Cytokine 71(2):385–393. https://doi.org/10.1016/j.cyto.2014.08.005
Siemeister G, Schnurr B, Mohrs K, Schachtele C, Marme D, Martiny-Baron G (1996) Expression of biologically active isoforms of the tumor angiogenesis factor VEGF in Escherichia coli. Biochem Biophys Res Commun 222(2):249–255. https://doi.org/10.1006/bbrc.1996.0730
Pizarro SA, Gunson J, Field MJ, Dinges R, Khoo S, Dalal M, Lee M, Kaleas KA, Moiseff K, Garnick S, Reilly DE, Laird MW, Schmelzer CH (2010) High-yield expression of human vascular endothelial growth factor VEGF(165) in Escherichia coli and purification for therapeutic applications. Protein Expr Purif 72(2):184–193. https://doi.org/10.1016/j.pep.2010.03.007
Lee IL, Li PS, Yu WL, Shen HH (2011) Prokaryotic expression, refolding, and purification of functional human vascular endothelial growth factor isoform 165: purification procedures and refolding conditions revisited. Protein Expr Purif 76(1):54–58. https://doi.org/10.1016/j.pep.2010.08.014
Kondo S, Matsumoto T, Yokoyama Y, Ohmori I, Suzuki H (1995) The shortest isoform of human vascular endothelial growth factor/vascular permeability factor (VEGF/VPF121) produced by Saccharomyces cerevisiae promotes both angiogenesis and vascular permeability. Biochem Biophys Acta 1243(2):195–202. https://doi.org/10.1016/0304-4165(94)00150-v
Ma L, Wang XN, Zhang ZQ, Chen AJ, Yao LH (2001) Expression of human vascular endothelial growth factor (VEGF165) in Pichia pastoris and its biological activity. Biomed Environ Sci 14(4):302–311
Lee GY, Jung WW, Kang CS, Bang IS (2006) Expression and characterization of human vascular endothelial growth factor (VEGF165) in insect cells. Protein Expr Purif 46(2):503–509. https://doi.org/10.1016/j.pep.2005.09.023
Lee SB, Park JS, Lee S, Park J, Yu S, Kim H, Kim D, Byun TH, Baek K, Ahn YJ, Yoon J (2008) Overproduction of recombinant human VEGF (vascular endothelial growth factor) in Chinese hamster ovary cells. J Microbiol Biotechnol 18(1):183–187
Grabenhorst E, Schlenke P, Pohl S, Nimtz M, Conradt HS (1999) Genetic engineering of recombinant glycoproteins and the glycosylation pathway in mammalian host cells. Glycoconj J 16(2):81–97
Sanchez O, Barrera M, Rodriguez MP, Frias MT, Figueroa NE, Naranjo P, Montesino R, Farnos O, Castell S, Venereo A, Ganges L, Borroto C, Toledo JR (2008) Classical swine fever virus E2 glycoprotein antigen produced in adenovirally transduced PK-15 cells confers complete protection in pigs upon viral challenge. Vaccine 26(7):988–997. https://doi.org/10.1016/j.vaccine.2007.11.014
Sanchez O, Toledo JR, Rodriguez MP, Castro FO (2004) Adenoviral vector mediates high expression levels of human growth hormone in the milk of mice and goats. J Biotechnol 114(1–2):89–97. https://doi.org/10.1016/j.jbiotec.2004.06.009
Benihoud K, Yeh P, Perricaudet M (1999) Adenovirus vectors for gene delivery. Curr Opin Biotechnol 10(5):440–447
Han ZS, Li QW, Zhang ZY, Yu YS, Xiao B, Wu SY, Jiang ZL, Zhao HW, Zhao R, Li J (2008) Adenoviral vector mediates high expression levels of human lactoferrin in the milk of rabbits. J Microbiol Biotechnol 18(1):153–159
Sanchez O, Barrera M, Farnos O, Parra NC, Salgado ER, Saavedra PA, Meza CD, Rivas CI, Cortez-San Martin M, Toledo JR (2014) Effectiveness of the E2-classical swine fever virus recombinant vaccine produced and formulated within whey from genetically transformed goats. Clin Vaccine Immunol 21(12):1628–1634. https://doi.org/10.1128/CVI.00416-14
Gaillet B, Gilbert R, Amziani R, Guilbault C, Gadoury C, Caron AW, Mullick A, Garnier A, Massie B (2007) High-level recombinant protein production in CHO cells using an adenoviral vector and the cumate gene-switch. Biotechnol Prog 23(1):200–209. https://doi.org/10.1021/bp060187j
Garnier A, Cote J, Nadeau I, Kamen A, Massie B (1994) Scale-up of the adenovirus expression system for the production of recombinant protein in human 293S cells. Cytotechnology 15(1–3):145–155. https://doi.org/10.1007/bf00762389
He TC, Zhou S, da Costa LT, Yu J, Kinzler KW, Vogelstein B (1998) A simplified system for generating recombinant adenoviruses. Proc Natl Acad Sci USA 95(5):2509–2514. https://doi.org/10.1073/pnas.95.5.2509
Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, Hong JS, Horwitz MS, Crowell RL, Finberg RW (1997) Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 275(5304):1320–1323. https://doi.org/10.1126/science.275.5304.1320
Fuxe J, Liu L, Malin S, Philipson L, Collins VP, Pettersson RF (2003) Expression of the coxsackie and adenovirus receptor in human astrocytic tumors and xenografts. Int J Cancer 103(6):723–729. https://doi.org/10.1002/ijc.10891
Pose AG, Gomez JN, Sanchez AV, Redondo AV, Rodriguez ER, Segui RM, Ramos EM, Molto MP, Rodriguez ES, Cordero LR, Mallon AR, Nordelo CB (2011) Subunit influenza vaccine candidate based on CD154 fused to HAH5 increases the antibody titers and cellular immune response in chickens. Vet Microbiol 152(3–4):328–337. https://doi.org/10.1016/j.vetmic.2011.05.033
Fiebich BL, Jager B, Schollmann C, Weindel K, Wilting J, Kochs G, Marme D, Hug H, Weich HA (1993) Synthesis and assembly of functionally active human vascular endothelial growth factor homodimers in insect cells. Eur J Biochem 211(1–2):19–26
Cohen T, Gitay-Goren H, Neufeld G, Levi BZ (1992) High levels of biologically active vascular endothelial growth factor (VEGF) are produced by the baculovirus expression system. Growth Factors 7(2):131–138
Backer MV, Backer JM (2001) Functionally active VEGF fusion proteins. Protein Expr Purif 23(1):1–7. https://doi.org/10.1006/prep.2001.1472
Claffey KP, Senger DR, Spiegelman BM (1995) Structural requirements for dimerization, glycosylation, secretion, and biological function of VPF/VEGF. Biochem Biophys Acta 1246(1):1–9. https://doi.org/10.1016/0167-4838(94)00144-6
Guzman-Hernandez ML, Potter G, Egervari K, Kiss JZ, Balla T (2014) Secretion of VEGF-165 has unique characteristics, including shedding from the plasma membrane. Mol Biol Cell 25(7):1061–1072. https://doi.org/10.1091/mbc.E13-07-0418
Peretz D, Gitay-Goren H, Safran M, Kimmel N, Gospodarowicz D, Neufeld G (1992) Glycosylation of vascular endothelial growth factor is not required for its mitogenic activity. Biochem Biophys Res Commun 182(3):1340–1347. https://doi.org/10.1016/0006-291x(92)91880-y
Walter DH, Hink U, Asahara T, Van Belle E, Horowitz J, Tsurumi Y, Vandlen R, Heinsohn H, Keyt B, Ferrara N, Symes JF, Isner JM (1996) The in vivo bioactivity of vascular endothelial growth factor/vascular permeability factor is independent of N-linked glycosylation. Lab Invest 74(2):546–556
Morera Y, Lamdan H, Bequet M, Ayala M, Rojas G, Munoz Y, Gavilondo JV (2006) Biologically active vascular endothelial growth factor as a bacterial recombinant glutathione S-transferase fusion protein. Biotechnol Appl Biochem 44(Pt 1):45–53. https://doi.org/10.1042/BA20050169
Lindwall G, Chau M, Gardner SR, Kohlstaedt LA (2000) A sparse matrix approach to the solubilization of overexpressed proteins. Protein Eng 13(1):67–71
Hudson EP, Uhlen M, Rockberg J (2012) Multiplex epitope mapping using bacterial surface display reveals both linear and conformational epitopes. Sci Rep 2:706. https://doi.org/10.1038/srep00706
Yang J, Wang X, Fuh G, Yu L, Wakshull E, Khosraviani M, Day ES, Demeule B, Liu J, Shire SJ, Ferrara N, Yadav S (2014) Comparison of binding characteristics and in vitro activities of three inhibitors of vascular endothelial growth factor A. Mol Pharm 11(10):3421–3430. https://doi.org/10.1021/mp500160v
Favot L, Keravis T, Holl V, Le Bec A, Lugnier C (2003) VEGF-induced HUVEC migration and proliferation are decreased by PDE2 and PDE4 inhibitors. Thromb Haemost 90(2):334–343. https://doi.org/10.1160/TH03-02-0084
Marin V, Kaplanski G, Gres S, Farnarier C, Bongrand P (2001) Endothelial cell culture: protocol to obtain and cultivate human umbilical endothelial cells. J Immunol Methods 254(1–2):183–190
Baudin B, Bruneel A, Bosselut N, Vaubourdolle M (2007) A protocol for isolation and culture of human umbilical vein endothelial cells. Nat Protoc 2(3):481–485. https://doi.org/10.1038/nprot.2007.54
Geraghty RJ, Capes-Davis A, Davis JM, Downward J, Freshney RI, Knezevic I, Lovell-Badge R, Masters JR, Meredith J, Stacey GN, Thraves P, Vias M, Cancer Research UK (2014) Guidelines for the use of cell lines in biomedical research. Br J Cancer 111(6):1021–1046. https://doi.org/10.1038/bjc.2014.166
Dirks WG, MacLeod RA, Drexler HG (1999) ECV304 (endothelial) is really T24 (bladder carcinoma): cell line cross-contamination at source. In Vitro Cell Dev Biol Anim 35(10):558–559. https://doi.org/10.1007/s11626-999-0091-8
Kiessling F, Kartenbeck J, Haller C (1999) Cell-cell contacts in the human cell line ECV304 exhibit both endothelial and epithelial characteristics. Cell Tissue Res 297(1):131–140
Park JS, Hong GR, Baek SW, Shin DG, Kim YJ, Shim BS (2002) Expression and regulation of endothelial nitric oxide synthase by vascular endothelial growth factor in ECV 304 cells. J Korean Med Sci 17(2):161–167. https://doi.org/10.3346/jkms.2002.17.2.161
Suda K, Rothen-Rutishauser B, Gunthert M, Wunderli-Allenspach H (2001) Phenotypic characterization of human umbilical vein endothelial (ECV304) and urinary carcinoma (T24) cells: endothelial versus epithelial features. In Vitro Cell Dev Biol Anim 37(8):505–514
Hughes SE (1996) Functional characterization of the spontaneously transformed human umbilical vein endothelial cell line ECV304: use in an in vitro model of angiogenesis. Exp Cell Res 225(1):171–185. https://doi.org/10.1006/excr.1996.0168
Miles AA, Miles EM (1952) Vascular reactions to histamine, histamine-liberator and leukotaxine in the skin of guinea-pigs. J Physiol 118(2):228–257. https://doi.org/10.1113/jphysiol.1952.sp004789
Acknowledgements
The authors acknowledge financial support from FONDEF Grant D10i1282.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Parra, N.C., Mansilla, R., Aedo, G. et al. Expression and Characterization of Human Vascular Endothelial Growth Factor Produced in SiHa Cells Transduced with Adenoviral Vector. Protein J 38, 693–703 (2019). https://doi.org/10.1007/s10930-019-09867-y
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10930-019-09867-y