
Abstract
Soil compaction is a major threat to natural resources. However, little information is available on the impacts of soil compaction on arthropod diversity especially relative to different types of vegetation, land use and restoration activities. In response to this dearth of information, we studied soil compaction, as well as percentage soil moisture and mean leaf litter depth, associated with four vegetation types: natural vegetation (fynbos, the historic condition), agricultural land (vineyards), invasive alien trees, and vegetation cleared of invasive alien trees (recovering vegetation). Our study took place in the Cape Floristic Region, South Africa, a biodiversity hotspot, yet also an area of intense viticulture and heavy invasion by alien plants. We sampled soil surface-active arthropods using pitfall traps, and compared species richness and abundance in different vegetation types with various levels of soil compaction and other soil variables. Overall, vineyards had the highest soil compaction while natural fynbos and aliens had low and comparable compaction. For both arthropod species richness and abundance, the order of the four vegetation types was, from highest to lowest: natural fynbos, alien cleared sites, vineyards, and alien infested sites. Level of soil compaction negatively correlated with arthropod species richness but not with abundance. Neither soil moisture nor leaf litter depth on their own significantly affected arthropod species richness or abundance. While alien trees overall had a strong negative effect on both arthropod species richness and abundance, and much more so than vineyards, the situation is reversible, with removal of aliens being associated with rapid recovery of soil structure and of arthropod assemblages. This is an encouraging sign for restoration.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Human-induced soil compaction (dry bulk density) is one of most important factors threatening natural resources (van den Akker and Soane 2005; Eudoxie and Springer 2006; Kirby 2007). It occurs when soil undergoes mechanical stress, mainly through use of heavy machinery or overgrazing, especially during wet soil conditions. The indirect effects of soil compaction may be less clear, and are often the result of highly variable soil characteristics (Boizard et al. 2000).
Soil compaction warrants further attention as it structures vegetation (Roberts 1987; Mitchell 1991), including that of alien plants (Payet et al. 2001) and therefore, may influence temperature of the soil surface directly due to shade, and amount of leaf litter from vegetation (Farji–Brener et al. 2008) necessary for soil fauna. In addition, soil compaction from farming operations also reduces water infiltration (Chan 2001; Mitchell 1991) and increases soil erosion (Chan et al. 2006). Erosion, in turn, reduces vegetation cover (Mitchell 1991). However, the extent of soil compaction and its impact on surface-dwelling arthropods under different land uses has not been investigated to any extent.
Many soil organisms are important members of terrestrial ecosystems (Smit and Van Aarde 2001). While many arthropods spend part or all their lives in the soil (Chan and Barchia 2007), many others feed above ground, using the soil as a pupation or nesting site. The potential benefits from arthropods to agro-ecosystems include improvement of soil structure and nutrient cycling (Edwards and Bohlen 1996).
Plant litter decomposition is an important biological process driven by ground living organisms (Dyer et al. 1990; Lawrence and Samways 2003; Pausas et al. 2004). Knowledge of this process is important for conservation of the soil (Chan 2001). In turn, relationships between soil fauna and soil properties can be used to assess the impact of landscape management (Clapperton et al. 2004). Disturbance of the soil profile can change, directly or indirectly, the composition of arthropod diversity associated with it (Wallwork 1970; Manzer et al. 1984; Wall and Moore 1999; Dunxiao et al. 1999; Holland and Luff 2004; Farji–Brener et al. 2008; Jabin 2008). Moreover, the effects on soil structure may also affect species density through indirect effects mediated by their potential influence on plants (Van Dijck and Van Asch 2002; Coulouma et al. 2006).
In the Cape Floristic Region (CFR), the comparative influence of soil compaction on ground arthropod assemblages under various conditions of land use has not been investigated, despite this region being a global biodiversity hotspot (Mittermeier et al. 2005). There are severe stressors to the biodiversity of the region, especially from agricultural conversion (especially to vineyards) and from the extensive impact of invasive alien plants (Rouget et al. 2003; Gaertner et al. 2009; Fairbanks et al. 2004).
It is unknown how infestation by alien trees and from agricultural practices (specifically viticulture) influence soil compaction and associated arthropod diversity. Apart from Magoba and Samways (2012), there is little knowledge on how these assemblages recover when the alien trees are removed, a significant mitigation measure in the region which has been assessed using aerial taxa (Samways and Sharrat 2010). To date there is no information focusing on the role of soil compaction in this process of recovery. In view of this dearth of information, we pose two hypotheses. Firstly, we hypothesize that a change in vegetation structure will alter soil compaction. Secondly, we hypothesize that there are significant correlations between arthropod diversity, soil compaction, leaf litter, site location or soil moisture content under the four main vegetation types (natural vegetation, agricultural production, invasive alien trees, and recovering vegetation).
Study area and methods
Study sites
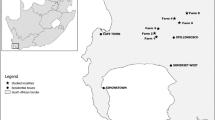
Vegetation of the study area (between Stellenbosch and Somerset West, South Africa: 18°15′E, 34°00′S) included many species-rich communities, occurring on highly infertile soils derived from sandstone of the Table Mountain Group. These communities range from upland study sites where soils are well drained and rocky to sand-loamy soil on the foothills and in the valleys. Intrusion of shale in some vineyards (i.e. Rustenberg) is evident. Boucher and Moll (1981) recognize two main soil types in the area. Lithosols, are generally shallow (<30 cm), grey sandy soils associated with the Table Mountain Group (TMG) sandstone, have a weak profile differentiation and contain coarse fragments and solid rock, and are largely the soil type studied here. Selected sites had similar edaphic conditions derived from the same parent material, TMG sandstone. For each site, the topographical conditions of adjacent distinct vegetation were similar.
At each of ten localities (three protected areas (PAs): Jonkershoek, Helderberg and Hottentots Holland; and seven wine estates: Vergelegen, Bilton, Stellenzicht, Driekoppen, Waterford, Rustenberg and Dornier) four vegetation types were selected: natural fynbos, alien invasive trees (IATs), cleared of invasive alien trees (CIATs), and vineyards. Transects (see details below) were established, so that the four vegetation types were adjacent to each other in their various combinations. This resulted in six different pairs of vegetation types (Natural fynbos-IATs; Natural fynbos-CIATs; IATs-CIATs; Natural fynbos-vineyards; vineyards-IATs; and vineyards-CIATs). In total, there were 36 transects.
The natural fynbos sites were untransformed by human activity and selected from the PAs and wine farms with <10 % alien tree cover. Natural fynbos was predominantly mountain fynbos, with common plant species being Aspalathus forbesii, A. aspalathoides, Lebeckia sepiaria, Lotononis prostrata, Cyphia phyteuma, Chasmanthe aethiopica, Watsonia borbonica, Gymnodiscus capillaris, Dimorphotheca pluvialis, Hymenolepis crithmoides Protea compacta, P. repens, P. neriifolia, and Salix species, as well as various ericas. The IAT sites had >90 % alien tree cover, represented mainly by Acacia mearnsii, A. longifolia, A. saligna, Hakea sericea, H. drupacea, Pinus pinaster, P. radiata, Eucalyptus lehmannii, E. diversicolor and Populus trees, with an understorey of grasses and forbs. The CIAT sites had been cleared of the alien trees in 2000, by felling and herbicide treating of stumps, and allowing the indigenous fynbos to recover of its own accord. After 6 years of recovery, the vegetation at these cleared sites was comparable in dominant species to that in natural fynbos (see above). The fourth vegetation type was organic vineyards, where there was no application of artificial fertilizers as the soils are relatively fertile through permanent crop cover (i.e. wheat), and pesticides were only applied as a last resort when necessary. One application of the chemical agent chlorpyrifos was applied during early August to control the mealybug Planococcus ficus which is the vector of a viral disease of the vines.
Sampling
Sampling of arthropods was on three occasions (August–October 2006, May–July 2007 and November 2007–January 2008) using pitfall traps (Samways et al. 2010). Each sampling occasion was for 5 days. Soil compaction and percentage (%) soil moisture content measurements at each sampling station were undertaken using a Radioactive moisture-density gauge instrument (Troxler 3411-B). Leaf litter depth was determined by inserting a steel rod, 4 mm in diameter, into the leaf-litter until the harder soil layer was reached (Lawes et al. 2005). Mean leaf-litter depth was then estimated from three random measurements in each 2 m2 quadrat created at each sampling station around the trap set.
The 256 m transects consisted of a trap-set of two individual pitfall traps, 1 m apart, placed at log 2 intervals: 2, 4, 8, 16, 32, 64 and 128 m on either side of the boundary between two adjoining vegetation types. Every patch on either side of the boundary was >100 m × 100 m, and all but one >250 m × 250 m. This spacing gave equal weighting to edges and interiors of patches. However, two transects, between alien vegetation and fynbos, were each four traps short, owing to unavailability of extensive sites. The total was 1000 pitfall traps (two per set, fourteen sets per transect, six transects per vegetation type pair and six vegetation pairs from four vegetation types, minus eight traps).
Pitfall traps for sampling arthropods were 500 ml plastic honey jars, each containing a replaceable paper cup, 8 cm diameter, 12 cm deep. Each trap was one-third filled with 70 % ethanediol. Traps remained closed during non-sampling periods, and opened for 5 consecutive days without rain during sampling periods (Borgelt and New 2006). Samples were then washed in water, and later transferred to 70 % ethanol for later identification in the laboratory. The collected surface-active arthropods were sorted and allocated to families. Where possible, they were identified to species level. Voucher specimens are in the Entomology Museum, Stellenbosch University, although spiders are in the National Collection of Arachnida, National Museum, Pretoria. Identification was by keys and expert opinion.
Data analyses
One-way analysis of variance (ANOVA) using SPSS v17 software (SPSS Inc. 2006), was performed on the selected soil factors comparing the different vegetation types. Classification trees for all the vegetation types in terms of soil compaction, leaf litter depth, percentage soil moisture and species richness were produced separately using CHAID growth limits incorporated in SPSS v17 software (SPSS Inc. 2006). Significance level for splitting nodes and merging categories was 0.05 and the significance values adjusted using the Bonferroni method. A variety of non-parametric species estimators were used to provide the best overall arthropod species estimates for all the vegetation types (Hortal et al. 2006). Incidence-based Coverage Estimator (ICE) is a robust and accurate estimator of species richness (Chazdon et al. 1998), whereas Chao2 and Jackknife estimators provide the least biased estimates for insufficient sampling (Colwell and Coddington 1994). Therefore, we calculated these estimators using EstimateS (Colwell 2006) for all the vegetation types separately and for a combination of these. One-way ANOVA was performed on the species and the log transformed abundance data comparing the different vegetation types with multiple comparisons of the means, using Tamahane’s post hoc. We used SpatialPack (version 0.2–3) in R (R Core Team 2013) to calculate the corrected Pearson’s correlations for spatial autocorrelation between assemblage composition and soil factors.
Results
Soil factors among the four vegetation types
Soil compaction was highest in vineyards, followed by cleared sites and fynbos, and lowest in aliens (Table 1). In turn, percentage moisture content was lowest in vineyards, followed by cleared sites, aliens, and finally fynbos with the highest. Litter depth was greatest by far in aliens, followed by fynbos and closely by cleared sites, with vineyards with by far the least. ANOVA among the four vegetation types showed that there were significant differences in soil compaction (df = 3, f = 19.36, p < 0.001), litter depth (df = 3, f = 296.6, p < 0.001) and percentage soil moisture (df = 3, f = 15.8, p < 0.001).
The classification tree of significant soil compaction values indicated similarity between fynbos and cleared sites (Table 1; online Figure 1). Aliens and vineyards were significantly different from each other, but neither was comparable to either fynbos or cleared sites. Classification of site locations based on soil compaction resulted in three nodes (Fig. 1; online Figure 2). All three nature reserves had similar low soil compaction (Node 1). Some vineyards (Node 2) had significantly higher soil compaction than any other location. Rustenberg, Bilton, and Dornier vineyards had more comparable soil compaction (Node 3).

Boxplots comparing means among fynbos (natural), invasive alien trees (IATs), cleared invasive alien trees and vineyard sites in terms of species richness, species abundance, soil compaction (kg/m3), percentage soil moisture and leaf litter depth (mm). Letters at the boxplots (a–d) indicate significant differences between means, based on the results of a Tamahane’s post hoc test after an analysis of variance test. The dark middle line of the box is the median. Whiskers (vertical lines) represent maximum and minimum values that are not statistical outliers. Black dots and stars are statistical outliers
Classification of the different vegetation types in terms of litter depth resulted in four separate categories (Table 1, online Figure 3). Vineyard clustering (Node 4) showed low leaf litter depth. Moreover, there were significant differences between the ten site locations (df = 506, f = 35.36, p < 0.001) in terms of litter depth, with the highest leaf litter depths recorded from Jonkershoek and Hottentots Holland nature reserves respectively (online Figure 4). However, Helderberg nature reserve was associated with relatively lower litter depth, which was comparable to those in some vineyards. Classification of different vegetation types using percentage soil moisture resulted in two vegetation categories: cleared sites were comparable to vineyards, and fynbos to aliens (online Figure 5).
Arthropod species richness among the four vegetation types
A total of 22,255 individuals were sampled, and assigned to 198 morphospecies/species. In terms of both arthropod species and abundance, the four vegetation types followed the same sequence, from highest to lowest: fynbos, cleared sites, vineyards, and aliens (Table 2). ANOVA among the four vegetation types gave significant differences among (df = 3, f = 41.65, p < 0.0001), and within (df = 509, f = 41.65, p < 0.0001) them in terms of species richness. Any one vegetation type shared at least 77 % of all sampled species, with fynbos having 90 % and cleared sites 85 % (Table 2). A Tamhane’s post hoc test after ANOVA among the four vegetation types (Fig. 1) gave significant differences in species richness between fynbos and aliens (p < 0.001); fynbos and vineyards (p < 0.001); cleared sites and alien sites (p < 0.001); and cleared sites and vineyard (p < 0.001). However, there were no statistically significant differences between cleared sites and fynbos (p = 1.00) in terms of species richness (online Figure 6). Although vineyards had relatively higher mean arthropod abundance than aliens (Table 2), the difference was not significant (p = 1.00). All other combinations of abundance were significant.
Correlation between soil factors and arthropod species richness and abundance among the four vegetation types
There was a significant negative correlation between soil compaction and species richness, but not abundance (Table 3). High soil compaction in vineyards did not correlate with reduced species richness. However, percentage soil moisture content and leaf litter depths were significantly negatively correlated with the soil compaction, and leaf litter depth was positively correlated to the percentage soil moisture, although neither soil moisture or litter depth on their own significantly affected arthropod species richness or abundance (Table 3).
Discussion
Soil factors associated with the vegetation types
We found high soil compaction in the vineyards, which comes about from many activities associated with grape production (Ferrero et al. 2005). At the other end of the spectrum, we found alien trees had significantly lower soil compaction compared to either fynbos or the sites cleared of alien trees, possibly due to a combination of root activity and accumulation of litter. As well as having highest compaction, vineyards also had lowest percentage moisture content, probably due to high evaporation rates associated with the structurally open production system, in comparison with the dense fynbos or alien vegetation. Overall, soil moisture was highly responsive to the type and structure of associated vegetation. While fynbos and alien vegetation had highest soil moisture, clearing of alien trees resulted in slightly reduced soil moisture. In addition to relatively higher soil moisture content, alien vegetation and fynbos both had deep litter layers, while vineyards and cleared sites had the shallowest layers, apparently due to increased exposure leading to increased decomposition.
All vineyards had similar and high levels of soil compaction, suggesting that compaction levels are not restricted to a particular local area but are relative to land use, with farming operations often occurring when the soils are moist and prone to compaction (Chan et al. 2006). This increased compaction can potentially reduce root penetration, water extraction and plant growth (Kirkegaard et al. 1992; Passioura 2002), leading to reduced crop production (McGarry and Chan 1984; McGarry 1990; Radford et al. 2001; Hamza and Anderson 2003).
The thick litter layer under alien vegetation may have lacked essential detritivores. This is suggested from the fact that the dense fynbos vegetation had much less litter than the aliens, probably because it supported the indigenous detritivores leading to the highly decomposed materials under fynbos compared to aliens and even vineyards.
Arthropod diversity in the different vegetation types
Arthropod species richness in cleared sites was high and similar to that in fynbos, and supported about 90 and 85 % of the total sampled species respectively. On the other hand, vineyards and alien sites had the lowest number of species respectively. Farm activities cause high soil compaction in vineyards, with the intensity of this compaction aggravated by relatively higher soil water content associated with vineyards (Usowicz 1992). Interestingly, species richness in vineyards was higher than in aliens, with vineyards being far from species poor. This suggests that the local surface-active fauna are well adapted to drier conditions, probably by being selected through the regular appearance of drought conditions in summer in this Mediterranean-type climate. Associated with this, is location, which also plays an important role, with Vergelegen Estate, for example, supporting more species than the Jonkershoek PA, suggesting that some vineyards can have value in maintaining an important component of arthropod diversity.
The aliens here supported fewer arthropod species than did those of indigenous vegetation, supporting the findings of Watts (1951) and Samways et al. (1996). The alien sites were drier than native vegetation but also had a dense (canopy giving deep shade) and a litter cover physically and chemically different from the historic condition. This meant that the alien tree cover was functionally different from the native vegetation, as in Michigan (Leege and Murphy 2001). Furthermore, arthropod species richness was relatively high in the native fynbos, with its low and varied soil compaction levels, compared to aliens, which had relatively uniform and low compaction levels.
Correlation between soil factors and arthropod species richness and abundance in different vegetation types
Density of soil fauna in agricultural areas is closely linked to soil structure (Usher 1975). Furthermore, increased soil compaction leads to a reduction in arthropod species diversity (Heisler and Kaiser 1995; Schrader and Lingnau 1997; Dittmer and Schrader 2000), with vineyard activities increasing soil compaction and altering the characteristics of the soil, leading to a reduction in habitats for the soil mesofauna (Larsen et al. 2004; Watson and Kelsey 2006). However, our study does not confirm these findings. This implies that disturbance of soil profile alone does not necessarily have as great an impact on the associated surface-active arthropods, as other studies might suggest. Nevertheless, soil has an effect on the amount of water held to enhance plant growth (Thomas and Squires 1991), and so provision of habitat for arthropods, which is reduced when the soil is compacted.
The differences in compaction levels between natural fynbos and cleared sites were small. It is interesting to view this in terms of restoration, with natural fynbos and cleared sites having similar soil compaction, and comparable arthropod species richness. This implies that, in terms of compaction at least, that alien trees leave the ground in a reduced state of compaction which, from our results, suggests that this enables good ground-living arthropod species recovery. In short, clearing alien trees sets the stage for recovery of arthropod assemblages to levels similar to those in natural vegetation, bearing in mind however, that aliens can change soil chemistry (Payet et al. 2001).
Conclusions and recommendations
We show here that vineyard soils are subject to some compactive degradation. This means that proficiency of soil to support higher arthropod diversity within the vineyards can be increased when operations are scheduled at appropriate moisture content (i.e. in dry soils). This is not to say that vineyards were poor in species, with 80 % of the total sampled accounted for. Even the alien plants, probably because of their ability to reduce compaction and increase leaf litter and moisture, had 77 % of the total arthropod fauna. However, arthropods were not so numerous or evenly distributed in the soil under alien trees than in natural vegetation, suggesting perhaps there could be ongoing impoverishment of the fauna should the alien trees not be removed. Overall, these results indicate the negative impact of alien invasive vegetation, more so than soil compaction in vineyards. Yet, this negative situation is reversible, with removal of invasive alien trees being associated with rapid recovery of certain soil conditions, enough to improve arthropod diversity.
References
Boizard H, Richard G, Brancourt-Hulmel M, Guérif J (2000) Effect of cropping systems on change in bulk density, penetration resistance and hydraulic conductivity in subsoil. In: Horn R, van den Akker JJH, Arvidsson J (eds) Advances in Geoecology, vol 32., CatenaReiskirchen, Germany, pp 233–241
Borgelt A, New TR (2006) Pitfall trapping for ants (Hymenoptera, Formicidae) in mesic Australia: what is the best trapping period? J Insect Conserv 10:75–77
Boucher C, Moll EJ (1981) South African Mediterranean shrublands. In: Di Castri F, Goodall DW, Specht RL (eds) Mediterranean-type shrublands. Elsevier Scientific, New York, pp 233–267
Chan KY (2001) An overview of some tillage impacts on earthworm population and diversity—implications for functioning in soils. Soil Tillage Res 57:179–191
Chan KY, Barchia I (2007) Soil compaction controls the abundance, biomass and distribution of earthworms in a single dairy farm in south-eastern Australia. Soil Tillage Res 94:75–82
Chan KY, Oates A, Swan A, Hayes RC, Dear BS, Peoples MB (2006) Agronomic consequences of tractor wheel compaction on a clay soil. Soil Tillage Res 89:13–21
Chazdon RL, Colwell RK, Denslow JS, Guariguata MR (1998) Species richness of woody regeneration in primary and secondary rainforests of north eastern Costa Rica. In: Dallmeier F, Comiskey JA (eds) Forest biodiversity research, monitoring and modeling: conceptual background and old world case studies. Pathenon, Paris, pp 285–309
Clapperton J, Chan KY, Larney FJ (2004) Managing the soil habitat for enhanced biological fertility. In: Abbott L, Murphy D (eds) Soil biological fertility—a key to sustainable land use in agriculture. Kluwer Academic Publishers, Dordrecht, pp 203–224
Colwell RK (2006) EstimateS 8.0.0. http://viceroy.eeb.uconn.edu/EstimateS
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity through extrapolation. Philos Trans R Soc Lond 345:101–118
Coulouma G, Boizard H, Trotoux G, Lagacherie P, Richard G (2006) Effect of deep tillage for vineyard establishment on soil structure: a case study in Southern France. Soil Tillage Res 88:132–143
Dittmer S, Schrader S (2000) Longterm effects of soil compaction and tillage on Collembola and straw decomposition in arable soil. Pedobiologia 44:527–538
Dunxiao H, Chunru H, Yaling X, Banwang H, Liyuan H, Paoletti MG (1999) Relationship between soil arthropods and soil properties in a suburb of Qianjiang City, Hubei, China. Crit Rev Plant Sci 18:467–473
Dyer ML, Meentemeyer V, Berg B (1990) Apparent controls of mass loss rate of leaf litter on a regional scale: litter quality vs. climate. Scand J For Res 5:311–323
Edwards CA, Bohlen PJ (1996) Biology and ecology of earthworms. Chapman and Hall, London, pp 142–145
Eudoxie GD, Springer RO (2006) Assessing and predicting compaction on agriculturally important soils in Trinidad. Biosyst Eng 95:119–126
Fairbanks D, Hughes C, Turpie J (2004) Potential impact of viticulture expansion on habitat types in the Cape Floristic Region, South Africa. Biodivers Conserv 13:1075–1100
Farji-Brener AG, Carvaja LD, Gei MG, Olano J, Sánchez JD (2008) Direct and indirect effects of soil structure on the density of an antlion larva in a tropical dry forest. Ecol Entomol 33:183–188
Ferrero A, Usowicz B, Lipiec J (2005) Effects of tractor traffic on spatial variability of soil strength and water content in grass covered and cultivated sloping vineyard. Soil Tillage Res 84:127–138
Gaertner M, DenBreeyen A, Hui C, Richardson DM (2009) Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: a meta-analysis. Prog Phys Geogr 33:319–338
Hamza MA, Anderson WK (2003) Responses of soil properties and grain yields to deep ripping and gypsum application in a compacted loamy sand soil contrasted with a sandy clay loam soil in Western Australia. Aust J Agric Res 54:273–282
Heisler C, Kaiser EA (1995) Influence of agricultural traffic and crop management on Collembola and microbial biomass in arable soil. Biol Fertil Soils 19:159–165
Holland JM, Luff ML (2004) The effects of agricultural practices on carabidae in temperate agroecosystems. Integr Pest Manag Rev 5:109–129
Hortal J, Borges PAV, Gaspar C (2006) Evaluating the performance of species richness estimators: sensitivity to sample grain size. J Anim Ecol 75:274–287
Jabin M (2008) Influence of environmental factors on the distribution pattern of centipedes (Chilopoda) and other soil arthropods in temperate deciduous forests. Cuvillier Verlag, Göttingen
Kirby M (2007) Letter to the editor: whither soil compaction research? Soil Tillage Res 93:472–475
Kirkegaard JA, So HB, Troedson RJ, Wallis ES (1992) The effect of compaction on the growth of pigeonpea on clay soils. I. Mechanisms of crop response and seasonal effects on a vertisol in a sub-humid environment. Soil Tillage Res 24:107–127
Larsen T, Schjønning P, Axelsen J (2004) The impact of soil compaction on euedaphic Collembola. Appl Soil Ecol 26:273–281
Lawes MJ, Kotze DJ, Bourquin SL, Morris C (2005) Surface-active invertebrates as potential ecological indicators of Afromontane forest condition in South Africa. Biotropica 37:109–118
Lawrence JM, Samways MJ (2003) Litter breakdown by the Seychelles giant millipede and the conservation of soil processes on Cousine Island, Seychelles. Biol Conserv 113:125–132
Leege LM, Murphy PG (2001) Ecological effects of the non-native Pinus nigra on sand dune communities. Can J Bot 79:429–437
Magoba RNN, Samways MJ (2012) Comparative footprint of alien, agricultural and restored vegetation on surface-active arthropods. Biol Invasions 14:165–177
Manzer FE, Storch RH, Sewell GH (1984) Evidence for a relationship between certain soil arthropods and acid scab development. Am J Potato Res 61:741–746
McGarry D (1990) Soil compaction and cotton growth on a vertisol. Aust J Soil Res 28:869–877
McGarry D, Chan KY (1984) Preliminary investigation of clay soil’s behaviour under furrow irrigated cotton. Aust J Soil Res 22:99–108
Mitchell PB (1991) Historical perspectives on some vegetation and soil changes in semi-arid New South Wales. Vegetatio 91:169–182
Mittermeier RA, Gil PR, Hoffman M, Pilgrim J, Brooks T, Mittermeier CG, Lamoreux J, da Fonseca GAB (2005) Hotspots revisited: earth‘s biologically richest and most endangered terrestrial ecoregions. Cemex, Conservation International and Agrupacion Sierra Madre, Monterrey
Passioura JB (2002) Soil conditions and plant growth. Plant Cell Environ 25:311–318
Pausas JG, Casals P, Romanyà J (2004) Litter decomposition and faunal activity in Mediterranean forest soils: effects of N content and the moss layer. Soil Biol Biochem 36:989–997
Payet C, Scholes MC, Balkwill K (2001) Some effects of the cultivation of pine on the chemistry of ultramafic soils. S Afr J Sci 97:603–608
R Core Team (2013) R: A language and environment for statistical computing. R foundation for statistical computing. Available: http://www.R-project.org. Accessed 2015 Feb. 02
Radford BJ, Yule DF, McGarry D, Playford C (2001) Crop response to applied soil compaction and to compaction repair treatments. Soil Tillage Res 61:157–166
Roberts DW (1987) A dynamical systems perspective on vegetation theory. Vegetatio 69:27–33
Rouget M, Richardson DM, Cowling RM, Lloyd JW, Lombard AT (2003) Current patterns of habitat transformation and future threats to biodiversity in terrestrial ecosystems of the Cape Floristic Region, South Africa. Biol Conserv 112:63–85
Samways MJ, Sharrat NJ (2010) Recovery of endemic dragonflies after removal of invasive alien trees. Conserv Biol 24:267–277
Samways MJ, Caldwell PM, Osborn RM (1996) Ground-living invertebrate assemblages in native, planted and invasive vegetation in South Africa. Agric Ecosyst Environ 59:19–32
Samways MJ, McGeoch MA, New TR (2010) Insect conservation: a handbook of approaches and methods. Oxford University Press, Oxford
Schrader S, Lingnau M (1997) Influence of soil tillage and soil compaction on microarthropods in agricultural land. Pedobiologia 41:202–209
Smit AM, Van Aarde RJ (2001) The influence of millipedes on selected soil elements: a microcosm study on three species occurring on coastal sand dunes. Funct Ecol 15:51–59
SPSS Inc. (2006) SPSS version 15.0 for windows. SPSS Inc., Chicago
Thomas DA, Squires VR (1991) Available soil moisture as a basis for land capability assessment in semi arid regions. Vegetatio 91:183–189
Usher MB (1975) Aggregation responses of soil arthropods in relation to the soil environment. In: Anderson JM, MacFayden A (eds.) The role of terrestrial and aquatic organisms in decomposition processes, p. 61–93
Usowicz B (1992) Statistical-physical model of thermal conductivity in soil. Pol J Soil Sci 25:25–34
van den Akker JJH, Soane B (2005) Compaction. In: Rattan L (ed) Encyclopaedia of soil science. Marcel Dekker, New York, pp 285–293
Van Dijck SJE, Van Asch TWJ (2002) Compaction of loamy soils due to tractor traffic in vineyards and orchards and its effect on infiltration in Southern France. Soil Tillage Res 63:141–153
Wall DH, Moore JC (1999) Interactions underground: soil biodiversity, mutualism, and ecosystem processes. Bioscience 49:109–117
Wallwork JA (1970) Ecology of soil animals. McGraw Hill Publisher, London, p 2883
Watson GW, Kelsey P (2006) The impact of soil compaction on soil aeration and fine root density of Quercus palustris. Urban Forestry and Urban Greening 4:69–74
Watts JCD (1951) Some comparative studies of the fauna in soils developed under natural forest, Pine and Blue Gum. Unpublished M.Sc. thesis, Grahamstown, Rhodes University, p. 72–115
Acknowledgments
We thank Cape Nature and farmers for their support, particularly G. Wright. The Department of Civil Engineering, Stellenbosch University kindly provided a radioactive moisture-density gauge (Troxler 3411-B). Special thanks to R. Gavhi, R. Magoba, M. Tshikalange, A. Johnson, T. Muofhe and S. Mchunu for their field assistance. Financial support was from the Centre for Invasion Biology (CIB) (RM), the Andrew Mellon Foundation, the National Research Foundation (MS), and the Claude Leon Foundation (JS).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Magoba, R.N., Samways, M.J. & Simaika, J.P. Soil compaction and surface-active arthropods in historic, agricultural, alien, and recovering vegetation. J Insect Conserv 19, 501–508 (2015). https://doi.org/10.1007/s10841-015-9771-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-015-9771-8