
Abstract
Purpose
This study investigated the protective effect of probucol on Müller cells exposed to high glucose conditions and examined potential mechanisms of action.
Methods
Primary human retinal Müller cells were incubated with high glucose (HG, 35 mM) in the present or absence of different concentrations of probucol for 24 h. Cell viability was determined using the CCK-8 method. Mitochondrial membrane potential (MMP) was measured using JC-1 staining and cell cycle by flow cytometry. The expression of nuclear factor E2-related factor 2 (Nrf2), glutamate–cysteine ligase catalytic subunit, and p62 was quantified using quantitative polymerase chain reaction and western blot.
Results
We found that HG inhibited cell proliferation, arrested cell cycle, and increased MMP in human Müller cells. Probucol activated the Nrf2/p62 pathway and upregulated the anti-apoptotic protein, Bcl2, and attenuated HG-mediated damage in Müller cells.
Conclusions
Our results suggest that probucol may protect Müller cells from HG-induced damage through enhancing the Nrf2/p62 signaling pathway.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Diabetic retinopathy (DR) is one of the most common and serious complications of diabetes mellitus. Currently, there are limited treatments for DR, and none of them can prevent its progression [1]; the outcome of DR depends on early recognition and early intervention. The pathogenesis of DR remains to be fully elucidated although oxidative stress is known to play a critical role [2]. Chronic hyperglycemia leads to the production of reactive oxygen species (ROS) through various pathways [3]. The retina is rich in mitochondria and has a high respiration rate to meet its metabolic needs; thus, the retina is prone to ROS production and is susceptible to oxidative damage. In DR, an imbalance between ROS production and antioxidant defense systems causes retinal oxidative damage and retinal cell death, which contributes to the pathogenesis of DR [4,5,6].
Müller cell malfunction critically contributes to the development of DR. These cells can release a variety of cytokines and growth factors, some of which are injurious to the retina and are related to vascular dysfunction and generation of new blood vessels, whereas others are beneficial in protecting retinal neurons from various damages; for instance, Nrf2 is mainly expressed in Müller cells in the retina [7,8,9]. Studies have shown that chronic hyperglycemia can first induce apoptosis in Müller cells and then cause changes in the retinal microenvironment and accelerate neuronal apoptosis, finally leading to the occurrence and development of DR [8, 10]. Normalizing Müller cell function is considered a good therapeutic strategy to prevent or treat DR.
Nuclear factor E2-related factor 2 (Nrf2) is one of the most important endogenous antioxidant pathways. Nrf2 can adjust the expression of antioxidant genes and detoxifying enzymes by reacting with antioxidant components (ARE) (including glutamate cysteine ligase catalytic subunit [GCLC], p62, etc.). GCLC can regulate the synthesis of glutathione and exert antioxidant effects [11]. Autophagy is another pathway that maintains cell homeostasis in vivo under oxidative stress; excessive ROS production can activate both autophagy and the Nrf2 pathway [12, 13]. p62 is a substrate for autophagy and may play a role in maintaining protein homeostasis and clearing damaged proteins through autophagy [14]. Further, p62 can activate the Nrf2 signaling pathway by interacting with Keap1. Nrf2 regulates p62 expression, resulting in positive feedback regulation between the two proteins [15, 16]. Autophagy and the Nrf2 pathway are closely connected via the Nrf2/p62 pathway, which plays a crucial role in maintaining homeostasis [17, 18]. Dysregulation of the Nrf2/p62 pathway has been shown to play an important role in the occurrence and development of various diseases, such as diabetic heart disease [19]. Thus, therapeutic approaches targeting the Nrf2/p62 pathway are expected to be beneficial in DR.
Probucol (PB), an anti-hyperlipidemic, anti-oxidative, and anti-inflammatory drug, has two phenolic hydroxyl groups in its molecular structure that are easily oxidized, exerting its strong antioxidant effect [20]. PB can improve cognitive dysfunction caused by oxidative stress in diabetic rats through Nrf2 pathway activation [21]. Further, PB can induce autophagy, reduce nerve cell apoptosis, and promote neurological function recovery after spinal cord injury, also promote neuroregeneration and ameliorate functional deficits in traumatic brain injury [22, 23]. PB has also been demonstrated to inhibit penile cell apoptosis and autophagy in diabetic rats [24]. Previously, we have shown that PB can inhibit intracellular ROS generation, promote proliferation and decrease Müller cell apoptosis under high glucose (HG) conditions [25]. In this study, we further investigated the mechanism of PB in protecting primary human Müller cells from HG-mediated damage, including the Nrf2/p62 signaling pathway.
Materials and methods
Cell culture and identification
The study protocol was approved by the Ethics Committee of AIER Eye Group. All donated human eyeballs were provided by the AIER Eye Bank of Changsha (Changsha, China). Consent forms were obtained by the Eye Bank. 10 eyes were used in this study. Human primary Müller cells were isolated and cultured as described previously [25]. The phenotype of cultured Müller cells was verified by immunolabeling for glutamine synthase (GS).
Cell proliferation assay
Cell proliferation was assed using the CCK-8 kit (Solarbio, Beijing, China) according to the manufacturer’s instructions. Briefly, cells were cultured in triplicate in 96-well plates (5 × 103 cells/well) in low glucose (LG, 5.5 mM, mimic normal state) or high glucose (HG, 35 mM, mimic diabetic state). Some cells under HG conditions were treated with different concentrations of PB (10 μM, 30 μM, 50 μM, 100 μM, and 150 μM) for 24 h after HG treatment for 24 h. Subsequently, cells were incubated with 10 μl CCK-8 solution at 37 ℃ for 4 h. Optical density values were measured using a multi-functional microplate reader (BioTek, VT, USA) at 490 nm.
Mitochondrial membrane potential (MMP) analysis
The fluorescent, lipophilic, and cationic probe JC-1 (Beyotime, China) was used to measure MMP according to the manufacturer’s directions. Briefly, Müller cells were seeded in a 12-well plate (1 × 105 cells/cell), which were routinely maintained in LG (low glucose—5.5 mM) media, which was the control. When we started the experiment, we first used HG (high glucose—35 mM) treatment for 24-h to mimic diabetic state (hyperglycemia) in vitro, then we added PB into HG cultured cells for additional 24-h as drug intervention (HG + PB group). Subsequently, the medium was removed, the cells were washed twice with cold phosphate-buffered saline (PBS), and then treated with JC-1 (10 mg/mL, 0.5 mL/well) at 37 °C for 30 min. After washing twice with 1 × incubation buffer, fluorescence intensity was immediately measured by fluorescence microscopy. For JC-1 green, Ex = 485 nm and Em = 525 nm; for JC-1 red, Ex = 535 nm and Em = 590 nm.
Cell cycle assay
Cell cycle was analyzed using the Cell Cycle and Apoptosis Analysis Kit (Beyotime, China) according to the manufacturer’s instructions. Briefly, Müller cells were seeded in 6-well plates (2 × 105 cells/well). After treatment with LG, HG and HG + PB (as detailed above), the cells were collected and fixed in ice-cold 70% ethanol at 4 °C overnight. On the next day, the cells were washed with PBS and stained with PI in the dark for 30 min at 37 °C. Samples were analyzed using flow cytometry (BD FACSCelesta, USA), and 30,000–80,000 cells were collected per sample. Data were analyzed using FlowJo 10.6 software.
RNA extraction and quantitative polymerase chain reaction (qPCR)
Total cellular RNA was extracted from different groups of Müller cells using TRIzol Reagent (Thermo Scientific, USA). The cDNA was synthesized using Revert Aid First Strand cDNA Synthesis Kit (Vazyme Biotech, Nanjing, China) according to the manufacturer’s instructions. Gene-specific primers were synthesized by TsingKe Biotech (Wuhan, China). The primer sequences used were as follows: Nrf2 (sense: TCAGCCAGCCCAGCACATCC; antisense: TCTGCGCCAAAAGCTGCATGC), p62 (sense: TCCAGGATCAGGGGTTAGGG; antisense: TAGGCAAGCTATGTGCTGGG), GCLC (sense: ACGGAGGAACAATGTCCGAG; antisense: TACTGAAGCGAGGGTGCTTG), β-actin (sense: CATGTACGTTGCTATCCAGGC, antisense: CTCCTTAATGTCACGCACGAT). Gene expression was then analyzed by real-time PCR (Roche, Switzerland) with 40 cycles of 95 °C for 15 s and 60 °C for 30 s. The β-actin gene was used as an internal control. The mRNA expression levels were calculated using the 2−△△Ct method.
Western blot (WB) analysis
Müller cells were washed with cold PBS, and total proteins were extracted with RIPA buffer (Beyotime, China) supplemented with a Protease Inhibitor Cocktail (Amyjet Scientific Co., Ltd, Wuhan, China). A BCA protein assay kit (Vazyme Biotech) was used to analyze the protein concentrations of the cell lysates. The protein samples (15 μg every sample) were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by transfer to polyvinylidene fluoride (PVDF) films. After blocking for 2 h in 5% BSA (w/v), the PVDF membranes were incubated overnight at 4 °C with the following primary antibodies: rabbit anti-Bcl2 (1:1000, Arigo, China); rabbit anti-GCLC (1:1000, Abcam, UK); rabbit anti-p62(1:1000, Abcam, UK); and mouse anti-GAPDH (1:1000, Abcam, UK). The membranes were then rinsed three times and incubated with anti-rabbit/mouse IgG (1:5000, Molecular Probe, USA) for 2 h. An electrochemiluminescence ECL kit (Thermo, USA) was then used to observe the immunoreactive bands on the Odyssey Fc Imaging System (LI-COR Biosciences, USA) followed by quantification using ImageJ software.
Statistical analysis
SPSS 25.0 and GraphPad Prism 8.0 software were used to analyze the data and plot graphs. Pearson's Chi-square test was used to compare cell cycles between the three groups, other comparisons among the three groups were performed using one-way ANOVA followed by Dunnett’s or Dunnett’s T3 post hoc tests. The results were expressed as the mean ± standard error. All experiments were repeated at least three times. P < 0.05 was used to indicate statistical significance.
Results
Morphological observation and identification of primary Müller cells
The cultured Müller cells at passage 3 were presented as cobblestone-like and some were elongated under phase-contrast light microscope (Fig. 1A). Immunofluorescent investigation showed that > 90% of cells were positive for GS (Fig. 1B–D), indicative of high purity primary human Müller cells. Cells at passages 3–5 were used for this study.

Morphology and identification of human Müller cells. A Morphology of primary Müller cells. Scale bar = 30 μm. B–D Identification of Müller cells by GS immunofluorescence staining. B The nuclei of Müller cells were stained with DAPI. C GS expression in Müller cells. D The merged image of B, C. Scale bar = 15 μm. GS glutamine synthase
Protective effect of PB against HG-induced cytotoxicity in Müller cells
When Müller cells were cultured with 35 mM glucose (HG) for 48 h, the viability was significantly reduced compared to 5.5 mM glucose (LG) treatment (Fig. 2). PB at the concentrations of 100 and 150 μM significantly protected Müller cell from HG-induced cell death. PB at the concentrations of 10–50 μM did not show any protective effect (Fig. 2).

PB improved Müller cell viability under HG conditions. Primary human Müller cells were incubated with 5.5 mM (LG) or 35 mM (HG) glucose for 48 h. The HG treated cells were also treated with or without different concentrations of PB. Cell viability was measured by CCK8 assay. Mean ± SEM, n = 3. *P < 0.05 versus the LG group. &P > 0.05 versus the LG group. One-way ANOVA followed by Dunnett’s post hoc tests. HG high glucose, LG low glucose, PB probucol
PB inhibited HG-induced reduction in mitochondrial potential and Bcl2 expression in Müller cells
To further explore the mechanism of protective effect of PB on HG-induced Müller cell death, we examined the mitochondrial membrane potential (MMP) in Müller cells using JC-1 staining along with the analysis of anti-apoptosis protein, Bcl2, using WB analysis. GH treatment significantly reduced MMP in Müller cells (Fig. 3A, B), but the reduction was prevented by PB treatment (100 μM of PB, Fig. 3A–D). The expression level of Bcl2, as shown in Fig. 3E, F, was significantly reduced under the HG condition compared with that in the control under LG conditions. PB treatment prevented HG-induced Bcl2 reduction.

PB reduced HG-induced apoptosis in Müller cells. A–C Representative images of JC-1 staining of Müller cells in the LG, HG, and HG + PB groups, Green: JC-1 monomer (green), Red: JC-1 polymer. Scale bar = 15 μm. D Analysis of fluorescence intensity in Müller cells. Mean ± SEM, n = 3. ***P < 0.001 versus the LG group. #P < 0.05 versus the HG group. One-way ANOVA followed by Dunnett’s post hoc tests. E Representative WB showing the expression of Bcl2 and housekeeping protein GAPDH. F Bar graph showing the relative expression of Bcl2. Mean ± SEM, n = 3. **P < 0.01 versus the LG group. One-way ANOVA followed by Dunnett’s post hoc tests. #P < 0.05 versus the HG group. One-way ANOVA followed by Dunnett’s T3 post hoc tests. HG high glucose, LG low glucose, PB probucol
PB inhibited the HG-mediated S phase arrest in Müller cells
We next investigated the effect of PB on HG-induced on the cell cycle arrest. In cells treated with HG, the percentage of cells in G1 phase was reduced and the percentage of cells in the S phase was increased (Fig. 4). PB treatment (100 μM) prevented HG-induced S phase arrest (P < 0.05, Fig. 4A, B).

PB prevented HG-induced S phase arrest in Müller cells. A Representative plots of cell cycle distribution in the LG, HG, and HG + PB groups. B Statistical chart of the proportion of cell cycle phases in each intervention group. Mean ± SEM, n = 420,463. *P < 0.05 versus the LG group. #P < 0.05 versus the HG group. Pearson's Chi-square test was used. HG high glucose, LG low glucose, PB probucol
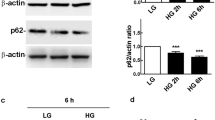
PB activated the Nrf2/p62 pathway in HG-treated Müller cells
To explore the effect of PB on the Nrf2/p62 pathway, the mRNA expression levels of Nrf2, GCLC, and p62 were determined by qPCR, and the protein expression of GCLC and p62 was analyzed by WB. HG treatment significantly reduced the mRNA expression of Nrf2, GCLC and p62 genes (Fig. 5A–C), and this was prevented by PB treatment (Fig. 5A–C). This effect was further confirmed at the protein levels by WB (Fig. 5D–F).

PB activated the Nrf2/p62 antioxidant signaling pathway in Müller cells. A–C Nrf2, GCLC, and p62 mRNA expression levels. Mean ± SEM, n = 12. *P < 0.05 versus the LG group. ***P < 0.0001 versus the LG group. #P < 0.05 versus the HG group. One-way ANOVA followed by Dunnett’s T3 post hoc tests. D–F GCLC and p62 protein expression levels. Mean ± SEM, n = 3. *P < 0.05 versus the LG group. **P < 0.01 versus the LG group. ####P < 0.00001 versus the HG group. One-way ANOVA followed by Dunnett’s post hoc tests. HG high glucose, LG low glucose, PB probucol, Nrf2 Nuclear factor E2-related factor 2, GCLC glutamate cysteine ligase catalytic subunit
Discussion
Diabetic retinopathy shows increasing prevalence and incidence rates worldwide, and the risk of vision loss is very high. Currently, various therapeutic methods, including laser therapy, antibody injection, and vitrectomy, have been developed for treatment, which play an important role in preventing vision loss [3]. However, since treatments to restore retinas to pre-diabetic retinopathy states have not yet been developed, inhibiting progression from non-proliferative diabetic retinopathy diabetic retinopathy to proliferative diabetic retinopathy is more important in preventing vision loss and is the subject of intensive research [1]. We discovered a lipid-lowering drug that could prevent the occurrence of diabetic retinopathy in previous studies [19, 25], and we conducted the current study to further evaluate the efficacy of the natural product extracts and their underlying mechanisms.
Müller cells, which are pivotal for maintaining retinal homeostasis, exposure to high glucose levels can lead to retinal damage and dysfunction. Müller cell dysfunction is considered to being a major cause of DR, including diabetic retinal vasculopathy and neuropathy [10, 26], although the underlying mechanism remains poorly defined. In the present study, we found that HG could reduce Müller cell viability through multiple mechanisms, including affecting mitochondrial membrane potential, inducing cell cycle arrest, and powering the expression of anti-apoptotic molecule Bcl2.
PB, a lipid-lowering drug, has strong antioxidant effects due to its unique molecular structure of the phenolic hydroxyl group. The beneficial effects of PB on various chronic diseases have been described previously [21, 22]. Recent evidence indicated that PB has a great therapeutic potential in diabetes and its complications, such as diabetic nephropathy [25]. We found that PB at the concentration of 100 μM and above could protect Müller cells from HG-mediated toxicity. PB treatment increased MMP and Bcl2 expression, normalized cell cycle in Müller cells cultured under HG conditions.
The Nrf2 pathway is the most important endogenous antioxidant pathway and significantly contributes to the regulation of oxidative stress and apoptosis in various cells [27]. Malfunction of the Nrf2 pathway is known to contribute to many neurological diseases, such as Parkinson’s disease and Alzheimer’s disease [28,29,30,31]. Notably, this pathway has been reported to be associated with diabetic complications, including diabetic nephropathy [32] and DR [29]. Autophagy has been shown to result in prolonged Nrf2 activation in a p62-dependent manner. p62, an autophagy adaptor and acceptor, has a Keap1-interacting region domain, which allows p62 to sequester Keap1 into autophagosomes and impairs Nrf2 ubiquitylation [33], thus leading to further activation of the Nrf2 signaling pathway [34]. Under normal conditions, the Nrf2-Keap1-p62 loop is in a dynamic steady state to maintain cellular redox homeostasis [15]. In diabetes, the increased ROS production leads to autophagy activation, which prompts p62 to bind with Keap1 and subsequently inactivates Nrf2 [35]. In this study, we found that PB could activate the Nrf2/p62 signaling pathway in Müller cells under HG conditions. The expression of its downstream antioxidant reaction elements GCLC was also normalized by PB treatment. These data suggest that PB might protect Müller cells from HG by activating the Nrf2/p62 pathway.
Given the detrimental impact of high glucose levels on Müller cell function and retinal health, identifying therapeutic agents that can mitigate this damage is essential. Probucol, with its antioxidant and potential autophagy-inducing properties, offers promise as a potential therapeutic option for managing retinal complications associated with high glucose levels. However, at present, it is challenging to foresee the potential clinical application of the findings due to the absence of animal or human data in this study. Nevertheless, ongoing tests are in progress to investigate the in vivo effects of PB treatment.
Conclusions
In conclusion, this study highlights the beneficial effects of probucol in attenuating high glucose-induced damage to Müller cells, potentially through enhancing the Nrf2/p62 signaling pathway. The findings provide valuable insights into the molecular mechanisms underlying these protective effects and suggest probucol as a potential therapeutic candidate for managing retinal complications associated with high glucose levels. Further research is needed to uncover the precise mechanisms and determine the clinical viability of probucol as a treatment option.
Data availability
All data generated or analyzed during this study are included in this published article.
Abbreviations
- HG:
-
High glucose
- LG:
-
Low glucose
- PB:
-
Probucol
- MMP:
-
Mitochondrial membrane potential
- Nrf2:
-
Nuclear factor E2-related factor 2
- GCLC:
-
Glutamate cysteine ligase catalytic subunit
- qPCR:
-
Quantitative polymerase chain reaction
- DR:
-
Diabetic retinopathy
- ROS:
-
Reactive oxygen species
- ARE:
-
Antioxidant components
- GS:
-
Glutamine synthase
- PBS:
-
Phosphate-buffered saline
- WB:
-
Western blot
- SDS-PAGE:
-
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
- PVDF:
-
Polyvinylidene fluoride
References
Kayama Y, Raaz U, Jagger A, Adam M, Schellinger IN, Sakamoto M, Suzuki H, Toyama K, Spin JM, Tsao PS (2015) Diabetic cardiovascular disease induced by oxidative stress. Int J Mol Sci 16(10):25234–25263. https://doi.org/10.3390/ijms161025234
Ahsan H (2015) Diabetic retinopathy–biomolecules and multiple pathophysiology. Diabetes Metab Syndr 9(1):51–54. https://doi.org/10.1016/j.dsx.2014.09.011
Kang Q, Yang C (2020) Oxidative stress and diabetic retinopathy: molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol 37:101799. https://doi.org/10.1016/j.redox.2020.101799
Iwona BS (2016) Growth factors in the pathogenesis of retinal neurodegeneration in diabetes mellitus. Curr Neuropharmacol 14(8):792–804. https://doi.org/10.2174/1570159x14666160813182009
Ola MS, Al-Dosari D, Alhomida AS (2018) Role of oxidative stress in diabetic retinopathy and the beneficial effects of flavonoids. Curr Pharm Des 24(19):2180–2187. https://doi.org/10.2174/1381612824666180515151043
Rodriguez ML, Perez S, Mena-Molla S, Desco MC, Ortega AL (2019) Oxidative stress and microvascular alterations in diabetic retinopathy: future therapies. Oxidative Med Cell Longev 2019:4940825. https://doi.org/10.1155/2019/4940825
Navneet S, Zhao J, Wang J, Mysona B, Barwick S, Ammal Kaidery N, Saul A, Kaddour-Djebbar I, Bollag WB, Thomas B, Bollinger KE, Smith SB (2019) Hyperhomocysteinemia-induced death of retinal ganglion cells: the role of Müller glial cells and NRF2. Redox Biol 24:101199. https://doi.org/10.1016/j.redox.2019.101199
Coughlin BA, Feenstra DJ, Mohr S (2017) Müller cells and diabetic retinopathy. Vis Res 139:93–100. https://doi.org/10.1016/j.visres.2017.03.013
Ghaseminejad F, Kaplan L, Pfaller AM, Hauck SM, Grosche A (2020) The role of Müller cell glucocorticoid signaling in diabetic retinopathy. Graefes Arch Clin Exp Ophthalmol 258(2):221–230. https://doi.org/10.1007/s00417-019-04521-w
Govetto A, Hubschman JP, Sarraf D, Figueroa MS, Bottoni F, dell’Omo R, Curcio CA, Seidenari P, Delledonne G, Gunzenhauser R, Ferrara M, Au A, Virgili G, Scialdone A, Repetto R, Romano MR (2020) The role of Müller cells in tractional macular disorders: an optical coherence tomography study and physical model of mechanical force transmission. Br J Ophthalmol 104(4):466–472. https://doi.org/10.1136/bjophthalmol-2019-314245
Qin S, Chen Z, Wen Y, Yi Y, Lv C, Zeng C, Chen L, Shi M (2022) Phytochemical activators of Nrf2: a review of therapeutic strategies in diabetes. Acta Biochim Biophys Sin 55(1):11–22. https://doi.org/10.3724/abbs.2022192
Jain A, Lamark T, Sjottem E, Larsen KB, Awuh JA, Overvatn A, McMahon M, Hayes JD, Johansen T (2010) p62/SQSTM1 is a target gene for transcription factor NRF2 and creates a positive feedback loop by inducing antioxidant response element-driven gene transcription. J Biol Chem 285(29):22576–22591. https://doi.org/10.1074/jbc.M110.118976
Dehdashtian E, Mehrzadi S, Yousefi B, Hosseinzadeh A, Reiter RJ, Safa M, Ghaznavi H, Naseripour M (2018) Diabetic retinopathy pathogenesis and the ameliorating effects of melatonin; involvement of autophagy, inflammation and oxidative stress. Life Sci 193:20–33. https://doi.org/10.1016/j.lfs.2017.12.001
Zhang W, Geng X, Dong Q, Li X, Ye P, Lin M, Xu B, Jiang H (2023) Crosstalk between autophagy and the Keap1-Nrf2-ARE pathway regulates realgar-induced neurotoxicity. J Ethnopharmacol 301:115776. https://doi.org/10.1016/j.jep.2022.115776
Komatsu M, Kurokawa H, Waguri S, Taguchi K, Kobayashi A, Ichimura Y, Sou YS, Ueno I, Sakamoto A, Tong KI, Kim M, Nishito Y, Iemura S, Natsume T, Ueno T, Kominami E, Motohashi H, Tanaka K, Yamamoto M (2010) The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat Cell Biol 12(3):213–223. https://doi.org/10.1038/ncb2021
Kageyama S, Gudmundsson SR, Sou YS, Ichimura Y, Tamura N, Kazuno S, Ueno T, Miura Y, Noshiro D, Abe M, Mizushima T, Miura N, Okuda S, Motohashi H, Lee JA, Sakimura K, Ohe T, Noda NN, Waguri S, Eskelinen EL, Komatsu M (2021) p62/SQSTM1-droplet serves as a platform for autophagosome formation and anti-oxidative stress response. Nat Commun 12(1):16. https://doi.org/10.1038/s41467-020-20185-1
Suzuki T, Yamamoto M (2017) Stress-sensing mechanisms and the physiological roles of the Keap1-Nrf2 system during cellular stress. J Biol Chem 292(41):16817–16824. https://doi.org/10.1074/jbc.R117.800169
Kaushal GP, Chandrashekar K, Juncos LA (2019) Molecular interactions between reactive oxygen species and autophagy in kidney disease. Int J Mol Sci 20(15):3791. https://doi.org/10.3390/ijms20153791
Liu HW, Luo Y, Zhou YF, Chen ZP (2020) Probucol prevents diabetes-induced retinal neuronal degeneration through upregulating Nrf2. Biomed Res Int 2020:3862509. https://doi.org/10.1155/2020/3862509
Moskal N, McQuibban GA (2023) From jeopardy champion to drug discovery; semantic similarity artificial intelligence. Autophagy. https://doi.org/10.1080/15548627.2023.2210995
Huang JL, Yu C, Su M, Yang SM, Zhang F, Chen YY, Liu JY, Jiang YF, Zhong ZG, Wu DP (2019) Probucol, a “non-statin” cholesterol-lowering drug, ameliorates D-galactose induced cognitive deficits by alleviating oxidative stress via Keap1/Nrf2 signaling pathway in mice. Aging 11(19):8542–8555. https://doi.org/10.18632/aging.102337
Zhou Z, Chen S, Zhao H, Wang C, Gao K, Guo Y, Shen Z, Wang Y, Wang H, Mei X (2016) Probucol inhibits neural cell apoptosis via inhibition of mTOR signaling pathway after spinal cord injury. Neuroscience 329:193–200. https://doi.org/10.1016/j.neuroscience.2016.05.019
Chen CM, Gung PY, Ho YC, Hamdin CD, Yet SF (2023) Probucol treatment after traumatic brain injury activates BDNF/TrkB pathway, promotes neuroregeneration and ameliorates functional deficits in mice. Br J Pharmacol. https://doi.org/10.1111/bph.16157
Zhang KQ, Tian T, Hu LL, Wang HR, Fu Q (2020) Effect of probucol on autophagy and apoptosis in the penile tissue of streptozotocin-induced diabetic rats. Asian J Androl 22(4):409–413. https://doi.org/10.4103/aja.aja_89_19
Zhou X, Ai S, Chen Z, Li C (2019) Probucol promotes high glucose-induced proliferation and inhibits apoptosis by reducing reactive oxygen species generation in Müller cells. Int Ophthalmol 39(12):2833–2842. https://doi.org/10.1007/s10792-019-01130-8
Cuenca N, Fernandez-Sanchez L, Campello L, Maneu V, De la Villa P, Lax P, Pinilla I (2014) Cellular responses following retinal injuries and therapeutic approaches for neurodegenerative diseases. Prog Retin Eye Res 43:17–75. https://doi.org/10.1016/j.preteyeres.2014.07.001
Yin X, Nie J, Nie X, Nie Y (2022) The relationship between nuclear factor-erythrocyte-related factor 2 and antioxidant enzymes in the placenta of patients with gestational diabetes and the metabolism of umbilical cord endothelial cells. Cell Mol Biol 68(12):97–103. https://doi.org/10.14715/cmb/2022.68.12.18
Abd El Aziz AE, Sayed RH, Sallam NA, El Sayed NS (2021) Neuroprotective effects of telmisartan and nifedipine against cuprizone-induced demyelination and behavioral dysfunction in mice: roles of NF-kappaB and Nrf2. Inflammation. https://doi.org/10.1007/s10753-021-01447-6
Foresti R, Bucolo C, Platania CM, Drago F, Dubois-Rande JL, Motterlini R (2015) Nrf2 activators modulate oxidative stress responses and bioenergetic profiles of human retinal epithelial cells cultured in normal or high glucose conditions. Pharmacol Res 99:296–307. https://doi.org/10.1016/j.phrs.2015.07.006
Zhou Y, Jiang Z, Lu H, Xu Z, Tong R, Shi J, Jia G (2019) Recent advances of natural polyphenols activators for Keap1-Nrf2 signaling pathway. Chem Biodivers. 16(11):e1900400. https://doi.org/10.1002/cbdv.201900400
Osama A, Zhang J, Yao J, Yao X, Fang J (2020) Nrf2: a dark horse in Alzheimer’s disease treatment. Ageing Res Rev 64:101206. https://doi.org/10.1016/j.arr.2020.101206
Chen YJ, Kong L, Tang ZZ, Zhang YM, Liu Y, Wang TY, Liu YW (2019) Hesperetin ameliorates diabetic nephropathy in rats by activating Nrf2/ARE/glyoxalase 1 pathway. Biomed Pharmacother 111:1166–1175. https://doi.org/10.1016/j.biopha.2019.01.030
Tang Z, Hu B, Zang F, Wang J, Zhang X, Chen H (2019) Nrf2 drives oxidative stress-induced autophagy in nucleus pulposus cells via a Keap1/Nrf2/p62 feedback loop to protect intervertebral disc from degeneration. Cell Death Dis 10(7):510. https://doi.org/10.1038/s41419-019-1701-3
Jiang T, Harder B, Rojo de la Vega M, Wong PK, Chapman E, Zhang DD (2015) p62 links autophagy and Nrf2 signaling. Free Radic Biol Med 88(Pt B):199–204. https://doi.org/10.1016/j.freeradbiomed.2015.06.014
Luo J, Yan D, Li S, Liu S, Zeng F, Cheung CW, Liu H, Irwin MG, Huang H, Xia Z (2020) Allopurinol reduces oxidative stress and activates Nrf2/p62 to attenuate diabetic cardiomyopathy in rats. J Cell Mol Med 24(2):1760–1773. https://doi.org/10.1111/jcmm.14870
Acknowledgements
The authors thank the staff at the Research Institute of AIER Eye for providing technical advice in this study.
Funding
This research supported by Research and Development Plan of Key Fields in Hunan Province (2020SKC2007), Projects of research and development in key areas of Hunan Province (2020SK2133), the Natural Science Foundation of Hunan Province (2021JJ30046), the Science Research Fund of AIER Eye Hospital Group (AF2201D12, AM2001D4).
Author information
Authors and Affiliations
Contributions
YFZ and ZPC designed the study. YFZ, HWL, CXL, and XY conducted the experiments. YL, YFZ, and ZPC analyzed the data. YFZ wrote the manuscript. ZPZ and JSC revised the manuscript. All authors read the final version of the manuscript. All authors approve and agree to be responsible for all aspects of this work.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical approval and consent to participate
The experimental approach was approved by the Ethics Committee of AIER Eye Group. All donated human eyeballs were provided by the AIER Eye Bank of Changsha (Changsha, China). Donors and their legally authorized representatives agreed to the donation of their eyeballs after death for scientific research. Consent forms were also signed.
Consent for publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhou, YF., Liu, HW., Yang, X. et al. Probucol attenuates high glucose-induced Müller cell damage through enhancing the Nrf2/p62 signaling pathway. Int Ophthalmol 43, 4595–4604 (2023). https://doi.org/10.1007/s10792-023-02859-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10792-023-02859-z