
Abstract
The inoculation response of single arbuscular mycorrhiza fungi (AMF) or rhizobia (Rh) in relation to nitrogen (N) acquisition of plants is well established, while the combined effect of both AMF and Rh is poorly known with regard to N assimilation for changes in amino acids of white clover (Trifolium repens). A pot study was carried out to evaluate the effect of single versus dual inoculation of AMF (Rhizophagus intraradices) and rhizobium (Rhizobium trifolii) on plant growth, leaf and root N contents, root amino acid contents, and root N-related enzyme activities in white clover. One hundred days after inoculations, Rh inoculation significantly stimulated the root colonization by R. intraradices. A single inoculation of AMF or Rh improved the plant growth (biomass production, root projected area, and root volume), root N acquisition, and dual inoculation of AMF and Rh further expanded some of these positive effects on root projected area and root N contents than single inoculation. All the inoculations notably increased activities of root asparagines synthase, nitrate reductase, and glutamate synthase, whilst dual inoculation displayed a much stronger effect in asparagine synthase activity than single inoculation. Single Rh treatment increased root glutamate and proline content, single AMF inoculation induced an increase in glutamate, aspartate, arginine, and ornithine content, while dual inoculation stimulated the accumulation of aspartate and proline. These results suggested the cooperation between the AMF and Rh inoculations, which magnified the positive effect on partly N metabolites and N-assimilation relevant enzymes of white clover.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Plant growth is dependent on many external and internal factors, like water, temperature, mineral nutrition, and microbes, whilst plant nitrogen (N) levels are closely associated with growth rates. Changes in stable N-related products such as amino acids have proven to affect the N-assimilation of plants (Wild et al. 2018).
Arbuscular mycorrhizal fungi (AMF), one of the most widely distributed microorganisms in soil ecosystems, can form a beneficial symbiosis with roots of up to 80% of terrestrial plants for improved water and nutrient acquisition (Tuo et al. 2017; Wu et al. 2019; Zhang et al. 2019, 2020; Zou et al. 2020). AMF enables host plants to absorb and transport more N (organic and inorganic forms of N) through greater interception of mycorrhizal mycelium within the mycorrhizosphere, whilst ammonium ion (NH4+) is the preferred form of N for absorption by AMF (Jin et al. 2005; Yang et al. 2021). In addition, mycorrhizal fungi released glomalin into the soil, which also contributes to soil nutrients including N (He et al. 2020; Meng et al. 2020). The increase in N acquisition by mycorrhizas has been reported up to 74% in maize (Tanaka and Yano 2005) and 8.54% in chickpea (Akhtar and Siddiqui 2008).
Rhizobia (Rh) are gram-negative bacteria from soil, which colonize root hairs to form nodules for the N-fixation of atmospheric N by nitrogenase. Rh contributes 40–48 million tons of N annually through various forms of symbiosis with legumes (Neumann et al. 2011). AMF and Rh collectively form two different symbionts with legumes to increase N acquisition, whereas there is less information about the dual inoculations with AMF and Rh on N-assimilation of legumes. The plants co-inoculated with AMF and Rh could fix more N2 and thus have an improved N content, compared with single inoculation (Lakshman and Patil 2004). A symbiotic association of Sesbania rostrata, Azorhizobium caulinodans, and Glomus etunicatum exhibited the highest uranium removal rates than non-inoculation, single Rh inoculation, and single AMF inoculation, indicating an improved phytoremediation efficiency of uranium by the triple symbiosis (Ren et al. 2019). Mycorrhizal symbiosis is, therefore, an established strategy of plants to accelerate symbiotic N acquisition in P-deficit soil (Bulgarelli et al. 2017). In addition to these positive effects on N uptake, no or negative effects of dual inoculation with AMF and Rh have been found in pea and green gram (Saxena et al. 1997; Blilou et al. 1999). Franzini et al. (2010) reported the deleterious effect of co-inoculation with AMF and Rh on the growth of Phaseolus vulgaris, depending on the plant variety and the symbionts. In subtropical grassland soils, AMF inoculation did not change the N content and biological N-fixation of Melilotus alba by Rhizobium meliloti when soil P level was dramatically low (Hack et al. 2019). These results indicated the complex relationships in the triple symbionts of legume-AMF-Rh, and therefore, more work needs to be explored in diverse legume-AMF-Rh systems.
In the process of N-assimilation, nitrate assimilation in the roots occurs by a nitrate reduction to nitrite and then to ammonia, which is incorporated into the glutamine amide-N and then to glutamic acid (Glu) and other amino acids (Yoneyama and Suzuki 2019). Evidence has shown that soil inhabited symbiotic microbes play important roles in the N-assimilation of legumes (Martin et al. 2001), whereas the question of whether or not AMF and Rh have a cooperative effect on N assimilation of a legume plant, white clover, was poorly known. White clover (Trifolium repens L.) is a superior perennial leguminous pasture with high feed quality and is used as green manure to increase soil fertility, a strategy to improve the protein content of the coarse fodder and, thereby, facilitate the reduction of industrial N fertilizers application (Lüscher et al. 2014). The present study was aimed to evaluate the effect of inoculation with AMF and Rh on plant growth, leaf and root N levels, root enzyme activities associated with N-assimilation, and root amino acid levels in white clover, further analyzing the relationship between AMF and Rh in N-assimilation of white clover.
Materials and methods
Microbial materials
The Rhizobium trifolii (Rh) strain was obtained from the Agricultural Culture Collection of China (Beijing; No: ACCC 18,017) and isolated from white clover. The Rh strain was activated on yeast mannitol, and a single colony was cultured on yeast mannitol liquid medium at 28 °C for 18 h to logarithmic growth phase, and centrifuged at 8000×g for 5 min to get the precipitation, which was diluted by ddH2O for the bacterial suspension (approximately 4.29 × 108 cells/mL). An arbuscular mycorrhizal fungus, Rhizophagus intraradices (N.C. Schenck & G.S. Sm.) C. Walker & A. Schüßler, was provided by the Bank of Glomeromycota in China (No: BGC JX04B), and propagated in pot culture with white clover. Mycorrhizal inoculum contained colonized root segments, spores (18 spores/g), and hyphae.
Experimental design
The experiment was carried out in a completely randomized design with four treatments, as shown in Supplementary materials 1. The four treatments included single inoculation with Rhizophagus intraradices (+ AMF−Rh), single inoculation with Rhizobium trifolii (− AMF + Rh), dual inoculation with Rhizophagus intraradices and Rhizobium trifolii (+ AMF + Rh), and inoculation with autoclaved (121 °C, 0.11 MPa, 2 h) mycorrhizal inoculums and ddH2O (− AMF − Rh). Each treatment had six replicates (corresponding to six pots), with a total of 24 pots (12 seedlings per pot).
Plant culture
Seeds of white clover were surface-sterilized with 75% alcohol solution for 5 min and with 0.525% of sodium hypochlorite solution for 5 min, then rinsed with ddH2O three times. Subsequently, the seeds were soaked in the bacterial suspension of Rhizobium trifolii (Rh) for 1 h as the Rh-inoculated treatment, and the non-Rh-inoculated seeds were soaked in ddH2O for 1 h. Afterward, these seeds were sown in plastic pots (15 cm top diameter × 10 cm bottom diameter × 12 cm height) filled with 2.5 kg of autoclaved (121 °C, 0.11 MPa, 2 h) soils and sand (4: 1, v/v) as the growing medium. The soil chemical properties were pH 6.2, Bray-P 16.4 mg/kg, and soil organic carbon 10.5 mg/g.
At the time of sowing, 100 g mycorrhizal inoculum was applied to the growth medium of the pot. Non-AMF treatment also received the same amount of autoclaved (0.11 MPa, 121 °C, 2 h) mycorrhizal inoculums. After 1 week of seed sowing, each pot was thinned to contain only 12 seedlings evenly distributed on the pot. Plants from four different treatments were watered every two days with 100 mL distilled water, and 20 mL full-strength Hoagland solutions (5 mM KNO3, 1 mM NH4H2PO4, 0.5 mM Ca(NO3)2, 0.5 mM MgSO4, 60 μM Fe-citrate, 92 μM H3BO3, 18 μM MnCl2⋅4H2O, 1.6 μM ZnSO4⋅7H2O, 0.6 μM CuSO4⋅5H2O, and 0.7 μM (NH4)6Mo7O24⋅4H2O) were weekly applied into each pot. All the pots were placed in a greenhouse with growing conditions characterized by 768 μmol/m2/s photosynthetic photon flux density, 30 °C/25 °C day/night temperature, and 68% relative humidity for 100 days. All the pots were weekly re-set, to eliminate the effect of pot location.
Observations on plant growth responses
The treated white clover plants were harvested 100 days after inoculation. To avoid plant enzyme inactivation, we randomly selected six plants per pot to analyze changes in plant growth, root morphology, root mycorrhizal colonization, and N contents, and the other six plants per pot were frozen in liquid nitrogen and stored at -80 °C for later analysis of amino acid content and N-assimilation relevant enzyme activities. Plants were divided into shoots and roots, whose fresh biomass was weighed and recorded. The root growth responses in terms of root total length, projected area, and volume were measured by the Epson Flatbed Scanner (J221A, Jakarta Selatan, Indonesia) and professional WinRHIZO software (Regent Instruments Inc., Quebec, Canada).
Determination of root mycorrhizal colonization
Five 1-cm long root segments per seedling were used for the analysis of root mycorrhizal colonization, resulting in a total of 180 root segments per treatment. The root samples from lateral roots were cleared by 10% of KOH solution for 10 min, bleached by 10% H2O2 for 10 min, incubated by 0.2 mol/L HCl for 8 min, and stained by 0.05% (w/v) of trypan blue for 5 min (Phillips and Hayman 1970). The mycorrhiza in the root was observed by a light microscope. Root mycorrhizal colonization was calculated as the percentage of AMF-colonized root lengths versus total root lengths.
Determination of leaf chlorophyll concentrations
A 0.2-g leaf sample was ground with 80% acetone solutions, filtered, and metered a constant volume of 25 mL. The absorbance of the filtrate was determined by a spectrophotometer at 663 nm and 646 nm. Chlorophyll a and b concentrations were calculated as per the formula described by Lichtenthaler and Wellburn (1983). Total chlorophyll concentration was the sum of chlorophyll a and chlorophyll b.
Determination of N and amino acid content
A 0.1-g fresh root sample was extracted with 1 mL 5.7 mol/L HCl for 22 h and centrifuged at 10,000×g for 5 min. The sample injected volume was 1 μL for the analysis of the LC–MS system (A Shimadzu LC-20ADXR HPLC system with an applied biosystems sciex Q-trap 5500 mass spectrometer). The sample vials were maintained at 4 °C, and the chromatographic separation was achieved using a gradient elution at 40 °C on an ACQUITY UPLC® BEH Amide column (100 mm × 2.1 mm, 1.7 μm, Waters, USA) with a VanGuard column (5 mm × 2.1 mm, 1.7 μm, Waters, USA). Mobile phase A was water containing 0.1% formic acid, and phase B was acetonitrile/water (95: 5, v/v) containing 2.5 mmol/L ammonium formate and 0.1% formic acid. The mobile phase gradient was followed by a flow rate of 0.3 mL/min. The process of the gradient elution included 100% B for 1 min, 100% B to 50% B for 5 min, 40% B for 3 min, and an equilibration time was 3 min (Liyanaarachchi et al. 2018).
Leaf and root N contents were determined following the method suggested by Zhao et al. (2012) with an electrochemical analyzer (Smartchem 200, Scientific Instruments Limited, Weston, USA).
Determination of N-assimilation relevant enzyme activities
The 0.2-g fresh root samples were extracted with 3 mL 50 mmol/L Tris–HCl buffer solutions (pH 7.5) containing 0.1 mol/L Tris, 2 mmol/L MgSO4, 2 mmol/L dithiothreitol, and 40 mmol/L sucrose, and then centrifuged at 10,000×g for 15 min at 4 °C. The supernatant was used for glutamine synthase (GS) and glutamate synthase (GOGAT) activity assays. The GS activity was assayed according to Husted et al. (2002) with minor modification, based on measuring the formation of γ-glutamyl hydroxamate. The 0.7 mL supernatants were added in 1.6 mL of reaction solution containing 80 mmol/L MgSO4, 20 mmol/L l -Na-glutamate, 20 mmol/L l-cysteine, and ATP at 37 °C for 30 min. The 1 mL of chromogenic reagent containing 0.2 mol/L trichloroacetic acid, 0.37 mol/L FeCl3 and 0. 6 mol/L HCl was added, and the absorbance was measured at 540 nm after 10 min. The GOGAT activity was measured as per the protocol described by Singh and Srivastava (1986) with minor modifications. The 0.3 mL supernatants were incubated with 2.7 mL mixed solutions containing 50 mmol/L Tris–HCl, 20 mmol/L l-glutamine, 20 mmol/L 2-oxoglutarate, and 0.3 mmol/L NADH at 30 °C for 30 min. The absorbance was measured at 340 nm.
Asparagine synthetase (AS) activity in roots was monitored as per the method as described by Joy et al. (1983). Nitrate reductase (NR) activity in roots was analyzed according to Foyer et al. (1998).
Statistical analysis
Data were analyzed by one-way analysis of variation (ANOVA) with SAS® software (9.1.3v) (SAS Institute Inc., Cary, NC, USA). The significant difference between the treatments was analyzed by the Tukey’s Honestly Significant difference at the 5% level. The Pearson’s correlation coefficients between N-assimilation relevant enzyme activities and amino acid contents in the root of white clover were performed using the Proc Corr’s procedure in SAS.
Results
Changes in root mycorrhizal colonization and plant growth responses
No mycorrhizal structures were observed in the non-AMF-inoculated white clover plants. The root mycorrhizal colonization of + AMF–Rh and + AMF + Rh treated plants was observed as 85.75% and 94.25%, respectively (Table 1). Therefore, Rh significantly increased root AMF colonization. Compared with the non-inoculated plant, single and dual inoculations displayed significantly higher shoot, root and total biomass, total root length, root projected area, and root volume (Table 1). The positive effect on the root projected area was considerably higher with + AMF + Rh treatment conditions than with either AMF or Rh inoculation treatment.
Changes in leaf chlorophyll levels
Compared with non-inoculated treatment, leaf chlorophyll a, chlorophyll b, and total chlorophyll levels were increased by sole AMF inoculation, single Rh inoculation, and dual inoculation, respectively (Supplementary materials 2). Dual inoculation with both AMF and Rh represented a significantly higher concentration of leaf chlorophyll a and total chlorophyll than sole inoculation with either AMF or Rh.
Changes in leaf and root N content and root amino acid contents
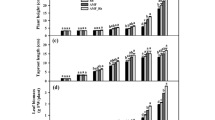
In white clover, root N content was considerably higher in roots than in leaves (Fig. 1a). Compared to – AMF − Rh treatment, treatments with – AMF + Rh, + AMF–Rh, and + AMF + Rh significantly increased leaf N content (Fig. 1a). Single Rh treatment also showed significantly higher leaf N content than single AMF inoculation and co-inoculation. Compared to – AMF − Rh treatment, + AMF − Rh, and − AMF + Rh treatments dramatically increased root Glu contents by 32.41% and 18.42%, respectively, without any significant change with + AMF + Rh treatment (Fig. 1b). The seedlings treated with + AMF − Rh and + AMF + Rh recorded 17.96% and 37.16% significantly higher Asp, 14.53% and 20.72% significantly higher Arg, and 14.94% and 33.15% significantly higher Orn in root, as compared with the seedlings with – AMF − Rh (Fig. 1b). However, no significant change in root Asp, Arg, and Orn levels was observed between − AMF + Rh treatment and – AMF − Rh treatment. The treatment with − AMF + Rh and + AMF + Rh observed an increase in root Pro contents by 22.94% and 42.56%, respectively, as compared with – AMF − Rh treatment.

Effects of arbuscular mycorrhizal fungi (Rhizophagus intraradices) and rhizobia (Rhizobium trifolii) on N contents in leaves and roots (a) and glutamate, aspartate, arginine, proline, and ornithine contents (b) in roots of white clover. Data (means ± SD, n = 6) followed by different letters above bars showed significant (P < 0.05) differences between treatments in the same tissue. * indicates the significant difference of N contents between leaf and root
Changes in enzyme activities relevant for N-assimilation in roots
Compared to the non-inoculated seedlings, the seedlings treated by single Rh, single AMF, and dual AMF and Rh inoculation represented 41.64%, 48.40%, and 43.45% significantly higher root GOGAT activities; 49.52%, 11.09%, and 61.23% significantly higher root AS activities; and 33.41%, 52.96%, and 66.67% significantly higher root NR activities, respectively (Table 2). However, different inoculation treatments displayed diverse responses on root GS activities: increase with dual inoculation; reduction with single Rh inoculation; and no changes with single AMF treatment.
Pearson's correlation studies
Root Glu levels were significantly positively correlated with GOGAT activity and negatively correlated with GS activity (Table 2). Likewise, root Asp, Pro, and Orn contents were significantly and positively correlated with GS, GOGAT, AS, and NR activity. Additionally, root Arg content was significantly and positively correlated with individual root GS activity.
Discussion
In this study, we evaluated the single or combined effect of AMF and Rh on plant growth and root N acquisition and assimilation in a leguminous plant, white clover. The results showed that single inoculation with AMF or Rh exhibited a positive effect on root N acquisition, amino acid contents, and N-related enzyme activities, and dual inoculation of AMF and Rh further magnified some of these responses, as evidenced by increased part N-assimilation enzyme activities and accumulation of some amino acids in roots. As a consequence, the two microbes had a cooperative effect on N acquisition and N assimilation, to some extent.
In our study, inoculation with Rhizobium trifolii considerably stimulated the colonization of Rhizophagus intraradices in roots of white clover, which is in agreement with the observations made earlier in Acacia senegal (Ndoye et al. 2015), It is well documented that AMF maintained a comparatively higher P-supply level in the mycorrhizosphere to meet the demand for elevated N2 fixation by Rh, which eventually aided in promoting higher mycorrhizal colonization of roots (Xavier and Germida 2003). Single Rh inoculation promoted the N level of white clover. Meanwhile, to maintain a stoichiometric balance between N and P in the plant, P becomes the more important nutrient element in the plant. As a result, more carbon sources would be assigned to AMF for exchanging more P, thereby, potentially increasing root mycorrhizal colonization (Liu et al. 2020). High AMF colonization also means high fungal biomass and carbon influx towards the fungus (Bulgarelli et al. 2017). Besides, the growth performance of white clover was observably promoted by inoculation with Rh, AMF, and dual Rh + AMF treatments. The earlier studies carried out in Pisum sativum (Yaseen 2016) and Amorpha canescens (Larimer et al. 2014) also reported similar findings. The improvement in plant biomass by co-inoculation is associated with changes in root morphology. Interestingly, co-inoculation did not show a dramatical advantage over single inoculation in improving biomass, which may be due to the fact that more energy materials (e.g. carbon) were consumed by the two microorganisms than by single microorganism, as seen with high root mycorrhizal colonization in co-inoculated white clover plants in our study and co-inoculated soybean plants with Bradyrhizobium elkanii and Glomus macrocarpum (Bulgarelli et al. 2017). More studies could be done around the change in biomass by single or dual inoculation.
The present study indicated increases in chlorophyll a, chlorophyll b, and total chlorophyll levels by different inoculation treatments, whereas dual AMF and Rh treatment further strengthened the magnitude of the positive effect on chlorophyll a and total chlorophyll levels. This suggested that co-inoculated plants produce more photosynthates in leaves than single inoculated plants and transfer them to the root, providing the substrate for the growth of mycorrhizal fungi and rhizobia. Thus, there was no obvious difference in biomass between the double and single inoculation treatments, as a result of changes in carbohydrate expenditure.
Inoculation of AMF or Rh improved the N accumulation in leaves and roots of white clover, but without any significant difference between single AMF treatment and single Rh inoculation on root N content. In leaves, single Rh treatment supported a significantly higher N content than single AMF inoculation. These observations suggested that mycorrhizal contribution to N accumulation is equivalent to the Rh contribution in roots, while Rh contribution to N accumulation is relatively higher than AMF in leaves. Dual inoculation of AMF and Rh showed a similar leaf and root N content, compared with single AMF treatment, while recorded a proportionately higher root N content and a lower leaf N content than single Rh treatment, implying that dual inoculation and AMF only altered N distribution in leaf and root. Analogous results were earlier reported by Hack et al (2019), who found that Melilotus alba plants co-inoculated with AMF and Rh represented lower leaf N content and higher root N content than single inoculation. Mycorrhizal hyphae are said to contribute as much as 30% N accumulated in plants. Moreover, the enhancement in N accumulation upon mycorrhization is strongly linked with an improvement of root architecture (Guo et al. 2017), mycorrhizal hyphal network (Clark and Zeto 2000), and changes in enzyme activities related to N metabolism (Hajong et al. 2013).
The enzyme NR plays a major role in N-assimilation by participating in the transformation of NO3− into NH4+ for plant’s utilization. In this study, Rh, AMF, and their combination collectively increased root NR activities of white clover, whereas dual inoculation of AMF and Rh had relatively greater effects than single Rh treatment. Mycorrhizal plants usually require higher NR activities to meet the requirement of accelerated N assimilation (Bago et al. 1996). GS/GOGAT can utilize ammonia as a substrate to synthesize glutamine and Glu for ammonia assimilation in plants. In our study, only dual inoculation of AMF and Rh stimulated GS activities in roots. Mycorrhizal extraradical hyphae can absorb and transport NH4+ for ammonium assimilation (Jin et al. 2005), a rather mandatory step to enhance the activity of GS/GOGAT. Our study also indicated the trend of the increase in root AS activities listed as + AMF + Rh treatment > − AMF + Rh treatment > + AMF-Rh treatment > − AMF − Rh treatment in the decreasing order, further suggesting a synergistic role of both AMF and Rh.
Glu is generally required for ammonium assimilation in AMF and bacteria by the GS/GOGAT pathway (Martin et al. 2001; Dunn 2015). In this study, single AMF or Rh treatment was beneficial for Glu accumulation in leaves and roots, while dual treatment of both AMF and Rh significantly decreased the Glu content, because Glu is considered as a precursor to synthesize other amino acids, as evidenced for higher Arg and Pro levels in dual-inoculated plants. Besides, Glu was significantly negatively correlated with GS activity while positively correlated with GOGAT activity in our study, since GS catalyzes Glu into Gln utilizing ammonia as a substrate, and GOGAT catalyses Gln into Glu (Yoneyama and Suzuki 2019). On the other hand, mycorrhizal fungal GS participated in ammonia assimilation into amino acids (Martin et al. 2001), and thus, legume plants do not need more GS involvement in N assimilation. The relationship between Glu and GS/GOGAT further indicated that in white clover, symbionts trigger the GOGAT activity to produce Glu in the process of ammonium assimilation.
AMF-inoculated plants, but not Rh-inoculated plants, had higher root Asp, Arg, and Orn contents than – AMF − Rh-treated plants, and dual inoculation of AMF and Rh further strengthened the accumulation of Asp. Asp has been proposed as an amino acid to link micro- and macro-symbiont C and N metabolism (Dunn 2015). Arg is the principal amino acid for N transfer into mycelia of AMF (Cruz et al. 2007), and as much as 50.4% N-assimilation in roots was reported as a contribution from the AMF (Jin et al. 2005). Arg was significantly and positively correlated with only root GS activity, as reported by Cruz et al. (2007) in the mycorrhizal mycelium. Hence, the relatively higher root mycorrhizal colonization as a result of dual inoculation of AMF and Rh further accelerated the rate of N absorption and their onwards assimilation. On the other hand, Orn is generated by Arg degradation in the intraradical mycelium of AMF and transferred into the glucose as a donor of carbon for mycorrhizal development (Wagemaker et al. 2007). The increase in Orn is, therefore, considered beneficial to N remobilization in Quercus ilex (Majumdar et al. 2016). Our study also revealed the significant positive correlation of root Asp, Pro, and Orn contents with NR, GS, GOGAT, and AS activities. Asp serves as a N-metabolic precursor and a N-transport compound in plants. Pro is derived from the deamination of Orn, and the Pro accumulation is responded to the developmental processes, photosynthesis, and protein synthesis (Armengaud et al. 2004). It was, therefore, concluded that dual inoculation with AMF and Rh operated through a cooperative/synergistic relationship to stimulate root Asp and Pro accumulation for essential metabolites of symbiosis in white clover, as well as an accelerated N-assimilation.
References
Akhtar MS, Siddiqui ZA (2008) Biocontrol of a root-rot disease complex of chickpea by Glomus intraradices, Rhizobium sp. and Pseudomonas straita. Crop Prot 27:410–417
Armengaud P, Thiery L, Buhot N, March GG, Savouré A (2004) Transcriptional regulation of proline biosynthesis in Medicago truncatula reveals developmental and environmental specific features. Physiol Plant 120:442–450
Bago B, Vierheilig H, Piché Y, Aguilar CA (1996) Nitrate depletion and pH changes induced by the extraradical mycelium of the arbuscular mycorrhizal fungus Glomus intraradices grown in monoxenic culture. New Phytol 133:273–280
Blilou J, Ocampo J, Garcia-Garrido J (1999) Resistance of pea roots to endomycorrhiza fungus or Rhizobium correlates with enhanced levels of endogenous salicylic acid. J Exp Bot 50:1663–1668
Bulgarelli RG, Marcos FCC, Ribeiro RV, Andrade SAL (2017) Mycorrhizae enhance nitrogen fixation and photosynthesis in phosphorus-starved soybean (Glycine max, L. Merrill). Environ Exp Bot 140:26–33
Clark RB, Zeto SK (2000) Mineral acquisition by arbuscular mycorrhizl plants. J Plant Nutr 23:867–902
Cruz C, Egsgaard H, Trujillo C, Ambus P, Requena N, Martins-Loução MA, Jakobsen I (2007) Enzymatic evidence for the key role of arginine in nitrogen translocation by arbuscular mycorrhizal fungi. Plant Physiol 144:782–792
Dunn MF (2015) Key roles of microsymbiont amino acid metabolism in rhizobia-legume interactions. Crit Rev Microbiol 41:411–451
Foyer CH, Valadier MH, Migge A, Becker TW (1998) Drought-induced effects on nitrate reductase activity and mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Physiol 117:283–292
Franzini VI, Azcon R, Mendes FL, Aroca R (2010) Interactions between Glomus species and Rhizobium strains affect the nutritional physiology of drought-stressed legume hosts. J Plant Physiol 167:614–619
Guo Q, Love J, Roche J, Song JC, Turnbull MH, Jameson PE (2017) A RootNav analysis of morphological changes in Brassica napus L. roots in response to different nitrogen forms. Plant Growth Regul 83:83–92
Hack CM, Porta M, Schäufele R, Grimoldi AA (2019) Arbuscular mycorrhiza mediated effects on growth, mineral nutrition and biological nitrogen fixation of Melilotus alba Med. in a subtropical grassland soil. Appl Soil Ecol 134:38–44
Hajong S, Kumaria S, Tandon P (2013) Comparative study of key phosphorus and nitrogen metabolizing enzymes in mycorrhizal and non-mycorrhizal plants of Dendrobium chrysanthum Wall. ex Lindl. Acta Physiol Plant 35:2311–2322
He JD, Chi GG, Zou YN, Shu B, Wu QS, Srivastava AK, Kuča K (2020) Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl Soil Ecol 154:103592
Husted S, Mattsson M, Mollers C, Wallbraun M, Schjoerring JK (2002) Photorespiratory NH4+ production in leaves of wild-type and glutamine synthetase 2 antisense oilseed rape. Plant Physiol 130:989–998
Jin HR, Pfeffer PE, Douds DD, Piotrowski E, Lammers PJ, Shachar-Hill Y (2005) The uptake, metabolism, transport and transfer of nitrogen in an arbuscular mycorrhizal symbiosis. New Phytol 168:301–310
Joy KW, Ireland RJ, Lea PJ (1983) Asparagine synthesis in pea leaves, and the occurrence of an asparagine synthetase inhibitor. Plant Physiol 73:165–168
Lakshman HC, Patil GB (2004) Dual inoculation of VA-mycorrhiza Rhizobium beneficial to Acacia nilotica wild. Ecol Environ Conserv 10:461–464
Larimer AL, Clay K, Bever JD (2014) Synergism and context dependency of interactions between arbuscular mycorrhizal fungi and rhizobia with a prairie legume. Ecology 95:1045–1054
Lichtenthaler HK, Wellburn RR (1983) Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans 11:591–592
Liyanaarachchi GV, Mahanama KR, Somasiri HP, Punyasiri PAN (2018) Development and validation of a method for direct, underivatized analysis of free amino acids in rice using liquid chromatography-tandem mass spectrometry. J Chromatogr A 1568:131–139
Liu CY, Zhang F, Zhang DJ, Zou YN, Shu B, Wu QS (2020) Transcriptome analysis reveals improved root hair growth in trifoliate orange seedlings by arbuscular mycorrhizal fungi. Plant Growth Regul 92:195–203
Lüscher A, Mueller-Harvey I, Soussana JF, Rees RM, Peyraud JL (2014) Potential of legume-based grassland–livestock systems in Europe: a review. Grass Forage Sci 69:206–228
Majumdar R, Barchi B, Turlapati SA, Gagne M, Minocha R, Long S, Minocha SC (2016) Glutamate, ornithine, arginine, proline, and polyamine metabolic interactions: the pathway is regulated at the post-transcriptional level. Front Plant Sci 7:1–17
Martin F, Cliquet JB, Stewart G (2001) Nitrogen acquisition and assimilation in mycorrhizal symbioses. In: Lea PJ, Morot-Gaudry JF (eds) Plant nitrogen. Springer, Berlin, pp 147–166
Meng LL, He JD, Zou YN, Wu QS, Kuča K (2020) Mycorrhiza-released glomalin-related soil protein fractions contribute to soil total nitrogen in trifoliate orange. Plant Soil Environ 66:183–189
Ndoye F, Kane A, Diedhiou AG, Bakhoum N, Fall D, Sadio O, Sy MO, Noba K, Diouf D (2015) Effects of dual inoculation with arbuscular mycorrhizal fungi and rhizobia on Acacia senegal (L.) Willd. seedling growth and soil enzyme activities in senegal. Int J Biosci 6:36–48
Neumann U, Kosier B, Jahnke J, Priefer UB, Al-Halbouni D (2011) Soil factors exhibit greater influence than bacterial inoculation on alfalfa growth and nitrogen fixation. FEMS Microbiol Ecol 77:590–599
Phillips JM, Hayman DS (1970) Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc 55:158–161
Ren CG, Kong CC, Wang SX, Xie ZH (2019) Enhanced phytoremediation of uranium-contaminated soils by arbuscular mycorrhiza and rhizobium. Chemosphere 217:773–779
Saxena AK, Rathi SK, Tilak KVBR (1997) Differential effect of various endomycorrhizal fungi on nodulating ability of green gram by Bradyrhizobium sp. (Vigna) strain S24. Biol Fert Soils 24:175–178
Singh RP, Srivastava HS (1986) Increase in glutamate synthase (NADH) activity in maize seedlings in response to nitrate and ammonium nitrogen. Physiol Plant 66:413–416
Tanaka Y, Yano K (2005) Nitrogen delivery to maize via mycorrhizal hyphae depends on the form of N supplied. Plant Cell Environ 28:1247–1254
Tuo XQ, Li H, Zou YN (2017) Alleviation of drought stress in white clover after inoculation with arbuscular mycorrhizal fungi. Not Bot Horti Agrobo 45:220–224
Wagemaker MJM, Eastwood DC, Drift CVD, Jetten MSM, Burton K, Griensven LJV, Camp HJOD (2007) Argininosuccinate synthetase and argininosuccinate lyase: two ornithine cycle enzymes from Agaricus bisporus. Mycol Res 111:493–502
Wild B, Alves RJE, Barta J, Capek P, Gentsch N, Guggenberger G, Hugelius G, Knoltsch A, Kuhry P, Lashchinskiy N, Mikutta R, Palmtag J, Prommer J, Schnecker J, Shibistova O, Takriti M, Urich T, Richte A (2018) Amino acid production exceeds plant nitrogen demand in Siberian tundra. Environ Res Lett 13:034002
Wu QS, He JD, Srivastava AK, Zou YN, Kuča K (2019) Mycorrhizas enhance drought tolerance of citrus by altering root fatty acid compositions and their saturation levels. Tree Physiol 39:1149–1158
Xavier LJC, Germida JJ (2003) Selective interactions between arbuscular mycorrhizal fungi and Rhizobium leguminosarum bv. viceae enhance pea yield and nutrition. Biol Fertil Soils 37:262–267
Yaseen T (2016) Effect of rhizobium, VAM and rock phosphate inoculation on growth and productivity and isolation of VAMF spores of Pisum sativum. Pesqui Agropecu Bras 5:564–572
Yang L, Zou YN, Tian ZH, Wu QS, Kuča K (2021) Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci Hortic 277:109815
Yoneyama T, Suzuki A (2019) Exploration of nitrate-to-glutamate assimilation in non-photosynthetic roots of higher plants by studies of 15N-tracing, enzymes involved, reductant supply, and nitrate signaling: a review and synthesis. Plant Physiol Bioch 136:245–254
Zhang F, Wang P, Zou YN, Wu QS, Kuča K (2019) Effects of mycorrhizal fungi on root-hair growth and hormone levels of taproot and lateral roots in trifoliate orange under drought stress. Arch Agron Soil Sci 65:1316–1330
Zhang F, Zou YN, Wu QS, Kuča K (2020) Arbuscular mycorrhizas modulate root polyamine metabolism to enhance drought tolerance of trifoliate orange. Environ Exp Bot 171:103962
Zhao XY, Zhou S, Wang G, Xing G, Shi W, Xu RK, Zhu ZL (2012) Nitrogen balance in a highly fertilized rice-wheat double-cropping system in Southern China. Soil Sci Soc Am J 76:1068–1078
Zou YN, Wu QS, Kuča K (2020) Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. https://doi.org/10.1111/plb.13161
Author information
Authors and Affiliations
Contributions
QSW designed the study; SMC measured experiment indexes; YNZ contributed to reagents, materials, analysis software, and experience equipment; MMX analyzed the data and draft the manuscript; and AKS, MMR and KK devoted to language modification. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Communicated by Hang-Wei Hu.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Xie, MM., Chen, SM., Zou, YN. et al. Effects of Rhizophagus intraradices and Rhizobium trifolii on growth and N assimilation of white clover. Plant Growth Regul 93, 311–318 (2021). https://doi.org/10.1007/s10725-020-00689-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-020-00689-y