
Abstract
Salvia sinaloensis Fern. (sage) is a medicinal plant containing plant secondary metabolites (PSMs) with antioxidant properties. The current study investigated the effects of drought stress on S. sinaloensis morphological and ecophysiological traits, and active constituent production. Sage plants were cultivated in controlled conditions for 34 days and exposed to full irrigation as control, half irrigation, or no irrigation. Changes in growth index (G.I.), dry biomass, leaf water potential (LWP), physiological parameters, active compounds, volatilome (BVOCs) and essential oils (EOs) were determined. Not irrigated plants showed a decrease in total chlorophyll content (~ − 14.7%) and growth (G.I., ~ − 59.4%) from day 18, and dry biomass at day 21 (− 56%), when the complete leaf withering occurred (LWP, − 1.10 MPa). Moderate drought stressed plants showed similar trends for chlorophyll content and growth but kept a constant LWP (− 0.35 MPa) and dry biomass throughout the experiment, as control plants. Carotenoids were not affected by water regimes. The photosynthetic apparatus tolerated mild to severe water deficits, without a complete stomatal closure. Plants under both stress conditions increased the percentage of phenols and flavonoids and showed altered BVOC and EO chemical profiles. Interestingly Camphor, the main EO oxygenated monoterpene, increased in moderate stressed plants while the sesquiterpene hydrocarbon Germacrene D decreased. The same trend was seen in the headspace under stress severity. The data evidenced a possible role of the active molecules in the response of S. sinaloensis plants to drought stress. Taking together, these findings point at S. sinaloensis as a potential drought adaptive species, which could be used in breeding strategies to obtain sages with high quality PSMs, saving irrigation water.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Biogenic volatile organic compounds (BVOCs) and essential oils (EOs) are plant secondary metabolites (PSMs), with important ecological functions within defence, protection, and signalling mechanisms (Loreto and Schnitzler 2010; Raut and Karuppayil 2014). Plants modulate their concentration to withstand stress-related conditions (Fleta-Soriano and Munné-Bosch 2016), such as those provoked by drought (Selmar and Kleinwachter 2013; Kleinwächter et al. 2015). Water deficit affects many aspects of plant physiology and biochemistry, activating root to shoot signalling hydraulic and hormones-mediated processes (Gulen and Eris 2004; Jaleel et al. 2008; Schachtman and Goodger 2008; Pirbalouti et al. 2014; Fleta-Soriano and Munné-Bosch 2016; Ali et al. 2017; Caser et al. 2017). Plants respond by closing their stomata and accumulating compatible solutes to maintain a low water potential and avoid dehydration (Skirycz and Inzé 2010). The increased diffusion barrier impairs influx of carbon dioxide (CO2) into leaves. In consequence, lower amounts of CO2 are fixed via the Calvin Cycle, and then less reduction equivalents (NADP+) are available as electron acceptors (Kleinwächter et al. 2015). In this state, all reactions must be promoted to consume NADPH + H+ such as the biosynthesis of highly reduced secondary compounds, i.e. phenols, terpenoids and alkaloids.
BVOCs and EOs are also sources for pharmaceutical, food additives, flavours, and fine chemicals (Zhao et al. 2005). In the last years the global EO production was around 50–100 tonnes per annum (Lubbe and Verpoorte 2011; Rauter et al. 2012; Ben Farhat et al. 2013). In medicinal and aromatic plants (MAPs), drought stress enhanced total phenolics and flavonoids production in Labisia pumila Benth. & Hook. (Jaafar et al. 2012) and monoterpene hydrocarbons and oxygenated sesquiterpenes in Helichrysum petiolare Hilliard & B.L. (Caser et al. 2016).
Salvia species is well known for possessing numerous secondary metabolites with significant benefits to human nutrition and health, used in folk medicine for a long time and therefore many species were listed in the official Pharmacopoeias. Antiinflammatory and antioxidant properties were observed in leaf extracts of S. officinalis L. (Munné-Bosch et al. 2001), anticonvulsant and sedative properties in leaves and seed extracts of S. leriifolia Benth (Hosseinzadeh and Arabsanavi 2001), and analgesic, neuroprotective and antiparkinson properties in roots of S. miltiorrhiza Bunge (Du and Zhang 2004). However, other Salvia species, such as in S. sinaloensis Fern, had beneficial properties (Abreu et al. 2008) but little information is available.
Despite the enormous richness of MAP species, breeding activities are advanced only for those plants with high demand and cultivation area (e.g. Origanum vulgare L. and Matricaria recutita L.). Wild collection and simple domestication are therefore still the main production strategies (Novak 2017). The need of support and speed up MAPs breeding also for minor crops impelled us to deepen the knowledge about S. sinaolensis characteristics in relation to cultivation practices.
In sage, imposed drought stress already proved to increase EOs, total polyphenols content and monoterpenes in S. officinalis (Abreu and Munné-Bosch 2008; Bettaieb et al. 2009, 2011; Radwan et al. 2017) and active constituents in S. miltiorrhiza (Liu et al. 2011).
Here we investigated S. sinaloensis dynamics in response to water limitation through morphological, ecophysiological and phytochemical analyses, contributing to our understanding of the mechanisms underlying plant adaptation and PSMs production.
Materials and methods
Plant material and treatments
Three year old plants from the CREA-FSO collection (Code database RGV FAO Salv095, Sanremo, Imperia, Italy − 43°81′60.28″N Lat, 7°76′67.38″E Long) were cloned by cuttings in the glasshouse of Dept. of Agricultural, Forest and Food Sciences of the University of Torino (Italy, 45°06′23.21″N Lat, 7°57′82.83″E Long), and cultivated in vases (9 cm in diameter—one plant per vase) containing peat (Silver Torf, Agrochimica, Bolzano, Italy) and Agriperlite® (70:30), and fertilized with 4 g L−1, Osmocote (15:11:13; Scotts Europe, The Netherland). When plants reached 20 cm in height, 120 pots were transferred in a growth chamber at 20 °C, 60% relative humidity and 16 h photoperiod (constant average PAR of 300 μmol m−2 s−1).
The experimental design was a split-plot design with three treatments and four replications per treatment. Plants were randomly divided in three groups and subjected to irrigation at 100% of container capacity (CC, control), half of the irrigation volume provided to CC controls (50% CC—moderate drought stress), or no irrigation (0% CC). The value of 50% CC was set to determine moderate drought stress in S. sinaloensis plants in a previous study (Caser et al. 2012). All the water contents were kept constant throughout the experiment. Gravimetric determinations of water contents were made by weighing soil samples before and after oven-drying to constant weight at 80 °C for 1 week. These values were used to calibrate all measurements of the moisture content of the substrate in the container. Container capacity was determined 48 h after irrigation and was calculated according to the equation of Paquin and Mehuys (1980). The soil moisture levels were maintained by manual irrigation and checked by weighting individual container every 2 days. The experiment lasted for a total of 34 days.
Ecophysiological measurements
Leaf water potential (LWP, Ψw), internal CO2 concentration (C i ), transpiration rate (E), stomatal conductance (g s ) and net photosynthetic rate (A) were measured twice a week (10–12 a.m.). In each treatment, LWP was counted in three leaves of five plants with a Scholander chamber (Soil Moisture Equipment, Santa Barbara, CA, USA) (Scholander et al. 1965).
C i E, g s and A were measured through a gas analyzer ADC-LCPro+ (The Analytical Development Company Ltd, Hoddesdon, UK). For each treatment three apical leaves of five plants were clamped in the leaf chamber, and routines of measurements were performed according to Caser et al. (2016). The CO2 concentration (450–470 ppm) and vapour pressure deficit (19.46 ± 1.67 Pa/KPa) were kept constant during the experiment. The ratio between A and E was used to calculate the instantaneous water use efficiency (WUE).
Determination of pigment content
Fifty mg from four fresh fully formed leaves per treatment were used to determine chlorophyll and carotenoids twice a week. After an over-night extraction in 5 ml of methanol at 4 °C in the dark, pigments were spectrophotometrically determined at 665, 652, and 470 nm using a Ultrospec 2100 pro (Amersham Biosciences, UK) as described by Lichtenthaler (1987).
The Chlorophyll Meter SPAD-502 (Konica Minolta Sensing Inc., Osaka, Japan) was used to measure the relative quantity of chlorophyll present in five randomly selected leaves per treatment.
Determination of growth index and biomass
Height and diameter of each plant per treatment were measured to calculate the growth index (G.I.; Π*{[(W’+W’’)/2]/2}2*H, W’ is the broadest diameter, W’’ is the perpendicular diameter and H is the height; Hidalgo and Harkess 2002). At day 34, roots and aerial parts of ten plants per treatment were separated and weighed. After recording their fresh biomass, they were oven-dried at 65 °C for 1 week and dry biomass was measured.
Determination of phenol and flavonoid content, and antioxidant activity
Fresh leaves (100 mg per treatment) were pulverized and homogenized in a mortar with 1 ml of 70% (v/v) methanol to facilitate the extraction. After 30 min of incubation on ice, the extracts were centrifuged at 10.000 g for 10 min at room temperature to collect the supernatant (methanol extract) to be used for the determination of phenol and flavonoid content, and the antioxidant activity. The content of total phenols was measured by using the Folin–Ciocalteau’s phenolic method and determined as reported by Singleton and Rossi (1965). Twenty µl of methanol extracted samples were added and mixed with 0.5 ml of Folin–Ciocalteau’s reagent and 0.45 ml of 7.5% (w/v) of saturated sodium carbonate solution. After the incubation at room temperature for 2 h, the absorbance at 765 nm of the samples was detected in UV–Vis spectrophotometer (Cintra 101, GBC Instruments, Australia).
Total flavonoid content was determined by applying the colorimetric method of Kim et al. (2003). 25 µl of methanolic extract were added to 225 μl of distilled water and to 75 μl of 5% (w/v) sodium nitrite (NaNO2). After 5 min of incubation were added 75 µl of 10% (w/v) of aluminum trichloride (AlCl3) and after 5 min were added 500 μl of 1M sodium hydroxide (NaOH). The absorbance of the samples was detected at the UV–Vis spectrophotometer (Cintra 101, GBC Instruments, Australia) after 15 min at 415 nm. The quantitative determination was made using a calibration curve with, as standard, quercetin 1:1 (w/v) dissolved in absolute methanol.
The antioxidant activity was determined by using the ferric reducing antioxidant power (FRAP) method with minor modification (Szőllősi and SzőllősiVarga 2002). The FRAP procedure is based on the reduction of a ferric-tripyridyl triazine complex to its ferrous colored form in the presence of antioxidants. An intense blue color complex was formed when ferric tripyridyl triazine (Fe3+-TPTZ) complex was reduced to the ferrous (Fe2+) form and the absorption at 593 nm was recorded. The calibration curve was plotted with absorbance at 593 nm versus concentration of Fe2+ solution which in turn plotted against concentrations of standard antioxidant. A total of 50 μl samples extract were added to 1.5 ml of the FRAP reagent and mixed well. The absorbance is measured at 593 nm using UV–Vis spectrophotometer (Cintra 101, GBC Instruments, Australia). Samples are measured in three replicates.
At the end of the experiment, the total amount of total phenols, flavonoids and antioxidant activity per plant (mg/plant FW) was estimated on the basis of the plant fresh biomass.
BVOC analysis
The evaluation of BVOCs was conducted at day 14. The headspace of 3 g of twig was measured by applying a Supelco Solid Phase Micro Extraction (SPME) (Supelco, Bellefonte, PA, USA) with polydimethylsiloxane (PDMS, 100 µm). Each sample was introduced into a 100 ml glass conical flask and equilibrated for 30 min. After the equilibration time, the fiber was exposed to the headspace for 15 min at room temperature; once sampling was finished the fiber was withdrawn into the needle and transferred to the injection port of the gas chromatography–electron impact mass spectrometry (GC-EIMS) system where the fiber was desorbed. GC-EIMS analysis was performed with a Varian CP 3800 gas chromatograph (Varian, Inc., Palo Alto, CA) equipped with a DB-5 capillary column (30 m × 0.25 mm; coating thickness 0.25 μm) and a Varian Saturn 2000 ion trap mass detector chromatograph (Varian, Inc., Palo Alto, CA). Analytical conditions were as follows: injector and transfer line temperature were 250 and 240 °C, respectively; oven temperature was programmed from 60 to 240 °C at 3 °C min−1; helium as carrier gas was set at 1 mL min−1; and the injection was in splitless mode. Identification of the constituents was based on comparison of their retention times with those of authentic samples, and on computer matching against commercial (NIST 98 and ADAMS) and home-made library mass spectra built from pure substances and MS literature data (Swigar and Silvestein 1981; Davies 1990; Adams 1995).
The relative proportions of the volatile constituents were percentages obtained by peak-area normalisation, and all relative response factors were taken as one.
EO analysis
Twenty g of dried leaves were used for the distillation of essential oils (EOs) by using a Clevenger-type apparatus (2 h) (Farmacopea Ufficiale della Repubblica Italiana, vol I. IXth edn. Zecca dello Stato: Rome, 1991). The yields of distillation were not determined due to the low amount of the starting plant material. The EOs obtained were solubilized in n-hexane, filtered over anhydrous sodium sulphate and stored in a vial at 4 °C in the dark until use. GC-EIMS was used to analyse all EOs (injection of 0.2 μl, 10% hexane solution) as reported above.
Statistical analysis
Data were previously tested for the variance homogeneity. Ryan-Einot-Gabriel-Welsch-F post-hoc test (REGW-F) (P < 0.05) was applied by means of the SPSS statistical package (version 19.0; SPSS Inc., Chicago. Illinois).
Results and discussion
Changes in ecophysiological parameters
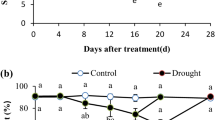
In this study, moderate drought stress did not affect LWP compared to control (Fig. 1). In both cases it was constant throughout the experiment, with a mean value of − 0.35 MPa. On the opposite, no irrigation significantly reduced LWP to − 0.64 MPa at day 7. Then, a decreasing trend was observed till day 21 (− 1.10 MPa) when the complete leaf withering occurred. Within the genus, similar results were found by Eakes et al. (1991) in S. splendens Sellow ‘Bonfire’ whose leaves reached a LWP ranging between − 1.10 and − 1.40 MPa when exposed to episodic drought (moisture stress conditioned). In other xerophytic sage species, drought induced much lower LWP. S. officinalis plants subjected to 50% CC and 25% CC reached LWP equal to − 3.00 and − 4.80 MPa, respectively (Bettaieb et al. 2011), and S. mellifera Greene under severe drought condition reached LWP equal to − 8.0 MPa, even if severely affected by embolism (Hargrave et al. 1994).

Dynamics of leaf water potential (LWP) of Salvia sinaloensis plants treated with full irrigation (100% container capacity, CC—black line), half irrigation (50% CC—dark grey line), or no irrigation (0% CC—light grey line). Means superscripted by the same letter do not differ significantly, according to REGW-F post-hoc test (ns non significant)
Stomatal closure is the primary mechanism by which plants regulate water loss. It is widely observed that plants reduce stomatal conductance of their leaves in response to declining water potential (Oren et al. 1999). Here, drought did not affect internal CO2 (C i) concentration with the exception for the 0% CC treatments that led to increase C i at day 21(Fig. 2a). More differences were found for E, gs and A, showing similar trends (Fig. 2b–d). Not irrigated plants showed lower values from day 14, shifted to day 21 (as regarding gs and A) and from day 25 (E). At the end of the experiment no differences were observed between control and half irrigation for all measured traits. Thus, drought stress led to partial stomata closure (Fig. 2b). As result, the influx of CO2 is diminished, visible by the strongly decreased rate of photosynthesis in the stressed plants (Fig. 2c) and the lower growth rate (Fig. 3). However, as the stomata closure is not very distinct, the actual CO2 concentration in the stressed plants is not significantly lower than in the controls. Obviously, the steady state concentration, at least in part, is determined by the photosynthetic action (i.e. the affinity of RuBisCO). This is confirmed by the fact that in the severely stressed plants, the internal CO2 concentration increased, when CO2 consumption is stopped due to the lacking photosynthetic activity after 21 days. Results suggested that the photosynthetic apparatus of S. sinaloensis could tolerate mild to severe water deficits and decreasing in photosynthesis couple with reduced stomatal function (Eakes et al. 1991; Galmes et al. 2007).

Gas exchange (internal CO2 concentration, C i—(a); transpiration rate, E—(b); stomatal conductance, g s—(c); net photosynthetic rate, A—(d) and dynamic of the instantaneous water use efficiency (WUE, A/E)—(e) measured on Salvia sinaloensis plants treated with full irrigation (100% container capacity, 100% CC—black line), half irrigation (50% CC—dark grey line), or no irrigation (0% CC—light grey line). Mean values showing the same letter are not statistically different at P ≤ 0.05 according to the REGW-F post-hoc test. The statistical relevance of ‘Between-Subjects Effects’ tests (ns non significant, *P < 0.05) was evaluated

Average values of growth index (G.I., cm3) during the experiment. S. sinaloensis plants were treated with three irrigation regimes: full irrigation (100% container capacity, 100% CC, black line), half irrigation (50% CC, dark grey line), or no irrigation (0% CC, light grey line). Means superscripted by the same letter do not differ significantly, according to REGW-F test (ns non significant; *P < 0.05; **P < 0.001)
Instantaneous water use efficiency (WUE, A/E) is used as an indicator of the water amount applied for growing by plants. Different authors indicated that stressed plants showing higher A/E are more able to utilize energy obtained by photosynthesis per unit of water transpired (Liu and Stützel 2004; Monclus et al. 2006). In this study, drought did not affect WUE except for the 0% CC treatment at day 21 (Fig. 2e). This finding indicates that S. sinaloensis could efficiently use water resources under moderate and severe drought stress condition at least under control conditions. In many species WUE was improved under water limitation and low gs (Liu et al. 2005). Among these, Trifolium alexandrinum L. (Lazaridou and Koutroubas 2004), Spartina alterniflora Loisel (Hessini et al. 2009), Callistemon (Alvarez et al. 2011), Hybamthus floribundus Lindl. (Kachenko et al. 2011), Rosa hybrida L. (Cai et al. 2012), Vitis vinifera L. ‘Grenache’ and ‘Tempranillo’ (Lovisolo et al. 2010; Medrano et al. 2015), and Helichrysum petiolare (Caser et al. 2016). In sage, Lambrecht et al. (2011) observed that leaf physiological activity of S. mellifera was not limited by imposed water deficits as well as indicated in our study. The photosynthetic apparatus was not completely affected by drought stress also in S. splendens ‘Bonfire’ (Eakes et al. 1991) and in S. pitcheri L. (Hamerlynck et al. 1997), highlighting the positive water use efficiency attitude of different species within the genus Salvia.
Changes in chlorophyll, carotenoids and plant growth
The decrease of chlorophyll and carotenoids in drought stressed plants is reported by several authors (Kaminska-Rozek and Pukacki 2004; Pastenes et al. 2005; Guerfel et al. 2009). Here, moderate and severe drought stress reduced the total chlorophyll content starting from day 18 up to the end of the experiment (Supplementary Table 1), while carotenoids and Chl:Car ratios were not affected by the imposed irrigation regimes. Plants of S. officinalis treated with 50% CC and 25% CC presented similar values and trends (Bettaieb et al. 2011). Similarly, a chlorophyll decrease was observed also in plants of Catharanthus roseus L. (Jaleel et al. 2008) and Helichrysum petiolare (Caser et al. 2016). Jaleel et al. (2009) indicated that carotenoids play an important role in protecting different processes from reactive oxygen species damages. Here, the presence of constant amount of carotenoid during the experiment suggests a possible role to cope with oxidative damages. Similar results were observed by Caser et al. (2016) in H. petiolare plants subjected to 50% CC water regime.
The evaluation of SPAD units by the use of the chlorophyll meter is commonly used to associate leaf damages and abiotic stresses (Caser et al. 2013) and leaf photosynthesis (Castelli et al. 1996). Here, no significant results were highlighted (data not shown). Similarly, Caser et al. (2012) showed no differences of SPAD values in S. dolomitica Codd and S. sinaloensis plants under drought conditions with the exception for those irrigated with 20% CC in which was observed an increase.
The decrease of chlorophyll content might cause a reduction in growth parameters of plants under water stress conditions (Viera et al. 1991). In this study, all the treated plants grew similarly till day 11 (Fig. 3). Then, drought stressed plants significantly reduced their growth. Not irrigated plants reached the lowest G.I. at day 21 (2654.00 cm3), while plants under 50% CC kept constant G.I. from day 18 onward, ranging between 6908.73 and 5686.40 cm3. These results are in agreement with a preliminary study on S. sinaloensis plants treated with different irrigation regimes where irrigation with 20% CC and 40% CC significantly reduced the plant growth compared to 80% CC and controls (Caser et al. 2012). However, as shown in Table 1, here only severe drought stress reduced drastically the total dry mass of plants (− 56%), affecting particularly the roots (− 67%). Thus, the R:A ratio (− 55%), favoring the shoot as showed in other species under severe drought stress conditions (Comas et al. 2013; Caser et al. 2016). A reduction of shoot dry weight, leaf area, stem length and root length was similarly seen in S. splendens (Eakes et al. 1991; Burnett et al. 2005). On the opposite, half irrigated plants showed an increase in root dry biomass (+ 14%) and consequently in R:A ratio (+ 37%) compared to control. Since roots are the only source to acquire water from soil, the root growth, its density, proliferation and size are key responses of plants to drought stress (Kavar et al. 2007). The drought tolerance of tea, onion and cotton was increased by improved root growth and root functioning (Farooq et al. 2009).
Changes in phenols and flavonoids and antioxidant activity
Polyphenols and flavonoids are among the most adaptable PSMs, helping plants to cope with different stress conditions (Di Ferdinando et al. 2014). In this study the total content of phenols in full-irrigated plants decreased during the whole experiment, reaching the lowest value between days 21 and 25 (0.98 mg/g FW) (Fig. 4a). Conversely, at the same data point moderate stressed plants exhibited the highest amounts (14.26 mg/g FW and 13.16 mg/g FW at the days 21 and 25, respectively). A different behavior was also observed in severe stressed plants, that showed higher content of phenols from day 11 to day 18 with the highest value at day 11 (15.74 mg/g FW). A similar dynamic was observed for the total flavonoids accumulation (Fig. 4b). In control plants, flavonoids increased during the first 7 days of cultivation. Subsequently, they dramatically decreased reaching minimum values between days 21 (1.19 mg/g FW) and 25 (1.16 mg/g FW). By contrast, in moderate stress condition flavonoid content kept constant until day 21, later increased with higher values from day 18 till the end of the experiment, reaching the maximum content at day 25 (2.89 mg/g FW). In the not irrigated plants, the total content of flavonoids peaked at day 11 (3.73 mg/g FW). In literature, different works showed that in drought stressed plants was observed an increase in secondary metabolites content. Plants of Prunus persica L., Echinacea purpurea L., Hypericum brasiliense Choisy, Trachyspermum ammi L. and Labisia pumila Benth. & Hook. subjected to drought stress showed significant increase in total phenols (Kubota et al. 1988; Gray et al. 2003; de Abreu and Mazzafera 2005; Azhar et al. 2011; Jaafar et al. 2012) and Pisum sativum in flavonoids (Nogués et al. 1998). In Salvia officinalis, an increase in phenols was observed in half irrigated plants (Bettaieb et al. 2011). Here, a mid to late increase in total phenols and flavonoids occurred in plants subjected to 50% CC. According with Paulsen and Selmar (2016), imposed drought stress generally results in decrease of aerial biomass gain and the corresponding dry or fresh weight are strongly influencing the content of natural products in MAPs. Here, as indicated in Table 2, moderate drought conditions resulted in a significant increase of the total content of phenols in comparison to severe drought stressed plants and controls (16.44, 22.85 and 10.43 mg/plant in 100% CC, 50%CC and 0% CC, respectively). While, no differences were noted in flavonoid content between moderate drought stress and control conditions (6.42, 6.47 and 2.73 mg/plant FW of flavonoids in 100% CC, 50% CC and 0% CC, respectively). These findings should be taken in consideration for future industrial purposes.

Leaf total phenols (a) and flavonoids (b), and antioxidant activity (c) of Salvia sinaloensis plants under control condition (black line, 100% container capacity, CC), moderate drought stress (dark grey line, 50% CC), and severe drought stress (light grey, 0% CC). Means superscripted by the same letter do not differ significantly, according to REGW-F test (ns not significant; *P < 0.05; **P < 0.001)
Drought induces oxidative stress in plants, in which reactive oxygen species (ROS) are produced (Munné-Bosch and Peñuelas 2003). Plant resistance to ROS is associated with an increase in antioxidant activity to prevent stress damage (Bor et al. 2003). In this study, only plants under 50% CC showed a significant increase of antioxidant activity at the end of the experiment (Fig. 4c). Similarly, at the end of the experiment the total amount of antioxidant activity per plant was much superior in moderate drought stressed plants than the others (106.37, 125.98 and 20.86 mg/plant FW in 100% CC, 50% CC and 0% CC, respectively) (Table 2). These findings confirm the ability of this plant to encounter the water limitation (50% CC) activating stress defense signaling and pathways respect to severe drought stress (0% CC) where metabolic and physiological constraints are not able to allow plant survival.
Changes in emitted BVOCs
Different authors observed that any stress factor can potentially change the rate of volatile release and alter the bouquet of BVOCs (Dicke and Baldwin 2010; Holopainen and Gershenzon 2010; Niinemets et al. 2013). Their emission can vary drastically depending on the species, organ, developmental stage and environmental conditions (Holopainen and Gershenzon 2010). Abiotic stress can enhances the emission of BVOC rates and patterns by the alteration of the communication with other organisms and photochemical cycles (Loreto and Schnitzler 2010). In the present study, the volatiles emitted and identified from the analyzed twigs are reported in Supplementary Table 2. In total a number of 20, 32 and 38 compounds were recognized in plants treated with full irrigation, half irrigation and no irrigation, respectively, accounting for 92.1, 96.6 and 95.4% of the total compositions, respectively. The volatile fractions were characterized mainly by sesquiterpene hydrocarbons (sh) although a mild reduction by increasing stress conditions was observed (57.5% < 42.8% < 42.7% in full irrigation, half irrigation and no irrigation, respectively). No irrigation affected the production of four of the six reported volatile molecule classes. In particular, an increase in oxygenated monoterpenes (om) (9.7, 16.2 and 23.6% in full irrigation, half irrigation and no irrigation, respectively), was observed. Among the other categories not-terpene derivates (nt), oxygenated sesquiterpene (os) and apocarotenoids (ac) were observed only in few percentage. However, plants under 50% CC exhibited the highest content in monoterpene hydrocarbons (mh) (24.2, 37.1 and 22.0% in full irrigation, half irrigation and no irrigation, respectively). All investigated headspaces showed different array of the main constituents. The chemical profile in fully irrigated plants was characterized by Germacrene D > β-Caryophyllene > β-Pinene, in moderate stressed plants by β-Pinene > Camphor > β-Caryophyllene and in severe stressed plants by β-Caryophyllene > Camphor > β-Pinene. Among the various constituents, a very sharp decrease (4.4 times) was observed for the Germacrene D (from 22.0 to 5.0% in full irrigation and no irrigation, respectively). On the opposite the oxygenated monoterpene Camphor increased 2.5 times when plants were subjected to 0% CC.
The literature is ambiguous concerning BVOC emission in relation to water availability. Several recent reviews addressed the roles of BVOCs in enhancing the tolerance of plants to various general abiotic stressors (Holopainen and Gershenzon 2010; Loreto and Schnitzler 2010; Possell and Loreto 2013; Rinnan et al. 2014). In particular, under drought Mediterranean climatic condition, the emission of oxygenated monoterpene is common (Loreto et al. 2014) and is thought to promote direct and indirect defense by modulating the signaling that biochemically activates defense pathways in response to stressful conditions (Loreto and Schnitzler 2010; Rinnan et al. 2014). Pistelli et al. (2013) reported an increase in oxygenated monoterpenes in Salvia officinalis plants under in vitro growing conditions. Moreover, there is evidence that the strength of the emission of BVOCs can be quantitatively related to the severity of abiotic stresses (Niinemets et al. 2013). As reported by Niinemets et al. (2004) the emission of oxygenated BVOCs depends on stomatal behavior. The stomatal closure in response to drought stress is therefore expected to drastically affect the emission of these compounds as showed in Quercus ilex L. (Bertin and Staudt 1996; Llusia and Peñuelas 1998; Loreto et al. 2001). Here, on the contrary we observed an increase in oxygenated monoterpenes under mild and severe drought stress associated with a not completely stomatal closure. Progressing soil water deficiency enhanced monoterpene emissions also in Pinus halepensis Mill. and Cistus albidus L., whereas did not affect it in Rosmarinus officinalis L. and Quercus coccifera L. (Ormeño et al. 2007). By contrast, overall emissions of sesquiterpenes were reduced by water deficiency in all the four species (Ormeño et al. 2007). Moreover, Hansen et al. (1997) and Ormeño et al. (2007) noticed that monoterpene emission in R. officinalis was not dependent by photosynthesis, but may originate from “de novo” synthesis in the photosynthetic tissues (Steinbrecher et al. 1999; Ormeño et al. 2009; Nogués et al. 2015). Taking together, all these results on BVOCs could represent important findings for studying the plant-environment interactions and can be used in breeding strategies to improve volatile compounds yield and quality.
Changes on essential oil
In a wide range of experiments was highlighted that plants under drought stress showed an increase in secondary metabolites content (Selmar and Kleinwachter 2013). Here, a total of 78 constituents were detected in the investigated EOs of which 59, 41 and 62 in full, half and not irrigated plants, respectively (Supplementary Table 3). Drought stress conditions only slightly increased the total number of constituents identified in the EOs (94.0, 99.6 and 98.8% at full irrigation, half irrigation and no irrigation, respectively). Similar results were observed in Satureja hortensis L. (Baher et al. 2002), Lippia berlandieri Schauer (Dunford and Vazquez 2005), Petroselinum crispum (Mill.) (Petropoulos et al. 2008), S. officinalis (Bettaieb et al. 2009), and Laurus nobilis L. (Maatallah et al. 2016) plants treated with similar drought stressed conditions. Regarding the type of constituents, moderate drought stress increased the total amount of monoterpenes (both oxygenated monoterpenes and monoterpene hydrocarbons: 51.5, 77.1 and 54.9% and 1.4, 6.0 and 1.9% in full irrigation, half irrigation and no irrigation, respectively) and deeply reduced the oxygenated sesquiterpenes (29.2, 6.3 and 26.9% in full irrigation, half irrigation and no irrigation, respectively). Similarly, imposed moderate drought stress strongly increased (two to fourfold) the content of Camphor, a specific compound in the essential oil, as also reported in S. officinalis plants (Bettaieb et al. 2009; Nowak et al. 2010) and in Picea abies L. and Pinus silvestris L. (Turtola et al. 2003). Nowak et al. (2010) reported that moderate stressed condition resulted in a massive increase of concentration of monoterpenes compensating the reduction in biomass. In fact, the stimulation to produce high terpene content under drought stress could be due to the low allocation of carbon to the growth, suggesting a trade-off between growth and defense (Turtola et al. 2003).
The main constituent of EOs in all the studied plants was Camphor (an oxygenated monoterpene). This constituent strongly increased under 50% CC (36.5, 62.8, 38.7% in full irrigation, half irrigation and no irrigation, respectively). Our results showed a chemical profile composed by Camphor > Caryophyllene oxide > α-Cadinol in 100% CC; Camphor > β-Caryophyllene > Borneol in 50% CC; and by Camphor > Caryophyllene oxide > β-Caryophyllene in 0% CC. Abreu et al. (2008) reported interesting antioxidant properties of S. sinaloensis due to the presence of phenolic diterpenes in comparison with S. officinalis extracts. In the present study, moderate drought stress application induced an increase in oxygenated monoterpenes that are generally the most active components in many EOs (Ruberto and Baratta 2000; Imanshahidi and Hosseinzadeh 2006), such as in S. cinnabarina L. with hypotensive effect (Alfieri et al. 2007), S. dolomitica with anti-bacterial activity (Kamatou et al. 2007) and in S. officinalis with antifungal and anti-inflammatory activity on mammalian cells (Abu-Darwish et al. 2013). These results may contribute to a good utilization of S. sinaloensis EOs for plant breeding and pharmaceutical applications.
Conclusions
Drought is one of the abiotic stresses that most severely affects plant growth and development. Consequently, plants adjust their structure, metabolism and function to withstand it. Generally, it is characterized by reduction of water content and leaf water potential, turgor loss, closure of stomata and decrease in cell enlargement and growth. However, with respect to MAPs, the water shortage can lead to an enhancement of the content of PSMs, Our results showed that in S. sinaloensis drought affects morphological, ecophysiological and chemical traits depending on the stress severity. Plant growth parameters and chlorophyll were generally reduced but the photosynthetic apparatus demonstrated to tolerate moderate to severe drought conditions, without a complete stomatal closure. Interestingly, under moderate stressed conditions, monoterpene hydrocarbons increased in both BVOCs and Eos (e.g. camphene), while volatile oxygenated monoterpenes increased still with increasing stress severity (e.g. camphor). Thus, drought stress did not inhibit per se the biosynthesis of monoterpenes, even better the highest BVOCs and EOs contents were obtained in moderate drought stress condition. Furthermore, this stress condition induced a mid to late increase in phenols, flavonoids and antioxidant activity, suggesting that active molecule accumulation is associated with drought tolerance.
In conclusion, here an integrated approach, combining metabolomic and physiological studies, allowed us to get new insights in mechanisms and processes involved in S. sinaloensis adaptation to drought stress and PSMs production. Coupling this information, breeders and industries may optimize MAPs breeding and cultivation, in order to produce high quality plant materials adopting sustainable cultivation techniques.
Abbreviations
- BVOCs:
-
Biogenic volatile organic compounds
- EOs:
-
Essential pils
- PSMs:
-
Plant secondary metabolites
- MAPs:
-
Medicinal and aromatic plants
- CC:
-
Container capacity
- LWP:
-
Leaf water potential
- Ci :
-
Internal CO2 concentration
- E :
-
Transpiration rate
- g s :
-
Stomatal conductance
- A :
-
Net photosynthetic rate
- G.I.:
-
Growth index
- FRAP:
-
Ferric reducing antioxidant power
- Fe3+-TPTZ:
-
Ferric tripyridyl triazine
- SPME:
-
Solid phase micro extraction
- GC-EIMS:
-
Gas chromatography–electron impact mass spectrometry
- WUE:
-
Water use efficiency
- FW:
-
Fresh weight
- DW:
-
Dry weight
References
Abreu ME, Munné-Bosch S (2008) Salicylic acid may be involved in the regulation of drought-induced leaf senescence in perennials: a case study in field-grown Salvia officinalis L. plants. Env Exp Bot 64:105–112
Abreu ME, Müller M, Alegre L, Munné-Bosch S (2008) Phenolic diterpene and α-tocopherol contents in leaf extracts of 60 Salvia species. J Sci Food Agric 88:2648–2653
Abu-Darwish MS, Cabral C, Ferreira IV, Gonçalves MJ, Cavaleiro C, Cruz MT, Al-bdour TH, Salgueiro L (2013) Essential oil of common sage (Salvia officinalis L.) from Jordan: assessment of safety in mammalian cells and its antifungal and anti-inflammatory potential. Biomed Res Int 2013:538940
Adams RP (1995) Identification of essential oil components by gas chromatography-mass spectroscopy. Allured, Carol Stream
Alfieri A, Maione F, Bisio A, Romussi G, Mascolo N, Cicala C (2007) Effect of a diterpenoid from Salvia cinnabarina on arterial blood pressure in rats. Phytother Res 21:690–692
Ali F, Bano A, Fazal A (2017) Recent methods of drought stress tolerance in plants. Plant Growth Regul 82:363–375
Alvarez S, Navarro A, Nicolas E, Sanchez-Blanco MJ (2011) Transpiration, photosynthetic responses, tissue water relations and dry mass partitioning in Callistemon plants during drought conditions. Sci Hortic 129(2):306–312
Azhar N, Hussain B, Ashraf MY, Abbasi KY (2011) Water stress mediated changes in growth, physiology and secondary metabolites of desi ajwain (Trachyspermum ammi L.). Pak J Bot 43:15–19
Baher ZF, Mirza M, Ghorbanli M, Rezaii MB (2002) The influence of water stress on plant height, herbal and essential oil yield and composition in Satureja hortensis L. Flavour Frag J 17:275–277
Ben Farhat M, Landoulsi A, Chaouch-Hamada R, Sotomayor JA, Jordán MJ (2013) Characterization and quantification of phenolic compounds and antioxidant properties of Salvia species growing in different habitats. Ind Crop Prod 49:904–914
Ben Taarit M, Msaada K, Hosni K, Hammami M, Kchouk ME, Marzouk B (2009) Plant growth, essential oil yield and composition of sage (Salvia officinalis L.) fruits cultivated under salt stress conditions. Ind Crops Prod 30:333–337
Bertin N, Staudt M (1996) Effect of water stress on monoterpene emissions from young potted holm oak (Quercus ilex L.) trees. Oecologia 107:456–462
Bettaieb I, Zakhama N, Aidi Wannes W, Kchouk ME, Marzouk B (2009) Water deficit effects on Salvia officinalis fatty acids and essential oils composition. Sci Hortic 120:271–275
Bettaieb I, Hamrouni-Sellami I, Bourgou S, Limam F, Marzouk B (2011) Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol Plant 33:1103–1111
Bor M, Özdemir F, Türkan I (2003) The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci 164:77–84
Bretzel F, Benvenuti S, Pistelli L (2014) Metal contamination in urban street sediment in Pisa (Italy) can affect the production of antioxidant metabolites in Taraxacum officinale Weber. Environ Sci Pollut Res 21:2325–2333
Burnett SE, Pennisi SV, Thomas PA, van Iersel MW (2005) Controlled drought affects morphology and anatomy of Salvia splendens. J Am Soc Hortic Sci 130:775–781
Cai X, Starman T, Niu G, Hall C, Lombardini L (2012) Response of selected garden rose to drought stress. HortScience 47:1050–1055
Caser M, Ruffoni B, Scariot V (2012) Screening for drought tolerance in Salvia spp. and Helichrysum petiolare: a way to select low maintenance ornamental plants. Acta Hortic 953:239–246
Caser M, Scariot V, Gaino W, Larcher F, Devecchi M (2013) The effects of sodium chloride on the aesthetic value of Buxus spp. Eur J Hortic Sci 78:153–159
Caser M, D’Angiolillo F, Chitarra W, Lovisolo C, Ruffoni B, Pistelli L, Pistelli L, Scariot V (2016) Water deficit trigger changes in valuable physiological and phytochemical parameters in Helichrysum petiolare Hilliard & B.L. Burtt. Ind Crops Prod 83:680–692
Caser M, Lovisolo C, Scariot V (2017) The influence of water stress on growth, ecophysiology and ornamental quality of potted Primula vulgaris ‘Heidy’ plants. New insights to increase water use efficiency in plant production. Plant Growth Regul 83:361–373
Castelli F, Contillo R, Miceli F (1996) Non-destructive determination of leaf chlorophyll content in four crop species. J Agric Crop Sci 4:275–283
Comas L, Becker S, Cruz VMV, Byrne PF, Dierig DA (2013) Root traits contributing to plant productivity under drought. Front Plant Sci 4:442
Davies NW (1990) Gas chromatographic retention indexes of monoterpenes and sesquiterpenes on methyl silicone and carbowx20M phases. J Chrom 503:1–24
de Abreu IN, Mazzafera P (2005) Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol Biochem 43(3):241–248
di Ferdinando M, Brunetti C, Agati G, Tattini M (2014) Multiple functions of polyphenols in plants inhabiting unfavorable Mediterranean areas. Environ Exp Bot 103:107–116
Dicke M, Baldwin IT (2010) The evolutionary context for herbivore-induced plant volatiles: beyond the ‘cry for help’. Trends Plant Sci 15:167–175
Du GH, Zhang JT (2004) The general situation and progress of the modern research of red sage root (Radix Salviae miltiorrhizae). II. Yiyao Daobao 23:435–440
Dunford NT, Vazquez RS (2005) Effect of water stress on plant growth and thymol and carvacrol concentrations in Mexican oregano grown under controlled conditions. J Appl Hortic 7:20–22
Eakes DJ, Wright RD, Seiler JR (1991) Moisture stress conditioning effects on Salvia splendens ‘Bonfire’. J Am Soc Hortic Sci 116:716–719
Farmacopea Ufficiale della Repubblica Italiana (1991) 9th edn. Istituto Poligrafico Zecca dello Stato, Roma
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009) Plant drought stress: effects, mechanisms and management. Agric Sustain Dev 29:185–212
Fleta-Soriano E, Munné-Bosch S (2016) Stress memory and the inevitable effects of drought: a physiological perspective. Front Plant Sci 7:143
Galmés J, Medrano H, Flexas J (2007) Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms. New Phytol 175:81–93
Gray DE, Pallardy SG, Garrett HE, Rottinghaus G (2003) Acute drought stress and plant age effects on alkamide and phenolic acid content in purple coneflower roots. Planta Med 69:50–55
Guerfel M, Baccouri O, Boujnah D, Chaibi W, Zarrouk M (2009) Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci Hortic 119:257–263
Gulen H, Eris A (2004) Effect of heat stress on peroxidase activity and total protein content in strawberry plants. Plant Sci 166:739–744
Hamerlynck EP, McAllister CA, Knapp AK, Ham JM, Owensby CE (1997) Photosynthetic gas exchange and water relation responses of three tallgrass prairie species to elevated carbon dioxide and moderate drought. Int J Plant Sci 158:608–616
Hansen U, van Eijk J, Bertin N, Staudt M, Kotzias D, Seufert G, Fugit JL, Torres L, Cecinato A, Brancaleoni E, Ciccioli P, Bomboi T (1997) Biogenic emissions and CO2 gas exchange investigated on four Mediterranean shrubs. Atmos Environ 31:157–166
Hargrave KR, Kolb KJ, Ewers FW, Davis SD (1994) Conduit diameter and drought-induced embolism in Salvia mellifera Greene (Labiatae). New Phytol 126:695–705
Hessini K, Martinez JP, Gandour M, Albouchi A, Soltani A, Abdelly C (2009) Effect of water stress on growth, osmotic adjustment, cell wall elasticity and water-use efficiency in Spartina alterniflora. Environ Exp Bot 67:312–319
Hidalgo PR, Harkess RL (2002) Earthworm castings as a substrate for poinsettia production. HortScience 37:304–308
Holopainen JK, Gershenzon J (2010) Multiple stress factors and the emission of plant VOCs. Trends Plant Sci 15:176–184
Hosseinzadeh H, Arabsanavi J (2001) Anticonvulsant effect of Salvia leriifolia Benth. Seed and leaf extracts in mice. Iran J Basic Med Sci 3:163–170
Imanshahidi M, Hosseinzadeh H (2006) The pharmacological effects of Salvia species on the central nervous system. Phytother Res 20:427–437
Jaafar HZE, Ibrahim MH, Fakri NFM (2012) Impact of soil field water capacity in secondary metabolites, phenylalanine ammonia-lyase (PAL), maliondialdehyde (MDA) and photosynthetic responses of Malaysian kacip Fatimah (Labisia pumila Benth). Molecules 17:7305–7322
Jaleel CA, Gopi R, Sankar B, Gomathinayagam M, Panneerselvam R (2008) Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. C R Biologies 331:42–47
Jaleel CA, Manivannan P, Wahid A, Farooq M, Somasundaram R, Panneerselvam R (2009) Drought stress in plants: a review on morphological characteristics and pigments composition. Int J Agric Biol 11:100–105
Kachenko AG, Bhatia N, Singh B (2011) Influence of drought stress on the nickel-hyperaccumulating shrub Hybanthus floribundus (Lindl.) F. Muell. Subsp. floribundus. Int J Plant Sci 172(3):315–322
Kamatou GPP, Van Vuuren SF, Van Heerden FR, Seaman T, Viljoen AM (2007) Antibacterial and antimycobacterial activities of South African Salvia species and isolated compounds from S. chamelaegnea. South Afr J Bot 73:552–557
Kaminska-Rozek E, Pukacki PM (2004) Effect of water deficit on oxidative stress and degradation of cell membranes in needles of Norway spruce (Picea abies (L.) Karst.). Acta Physiol Plant 26:431–442
Kavar T, Maras M, Kidric M, Sustar-Vozlic J, Meglic V (2007) Identification of genes involved in the response of leaves of Phaseolus vulgaris to drought stress. Mol Breed 21:159–172
Kim DO, Jeong SW, Lee CY (2003) Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem 81(3):321–326
Kleinwächter M, Paulsen J, Bloem E, Schnug E, Selmar D (2015) Moderate drought and signal transducer induced biosynthesis of relevant secondary metabolite in thyme (Thymus vulgaris), greater celandine (Chelidonium majus) and parsley (Petroselinum crispum). Ind Crops Prod 64:158–166
Kramer PJ, Boyer JS (1995) Water relations of plants and soils. Academic Press, New York
Kubota N, Mimura H, Shimamura K (1988) The effects of drought and flooding on the phenolic compounds in peach fruits. Sci Rep Fac Agric Okayama Univ 171:17–21
Lambrecht SC, Santiago LS, DeVan CM, Cervera JC, Stripe CM, Buckingham LA, Pasquini SC (2011) Plant water status and hydraulic conductance during flowering in the southern California coastal sage shrub Salvia mellifera (Lamiaceae). Am J Bot 98:1286–1292
Lazaridou M, Koutroubas SD (2004) Drought effect on water use efficiency of berseem clover at various growth stages. In: New directions for a diverse planet: Proceedings of the 4th International Crop Science Congress Brisbane, Australia, vol. 26
Lichtenthaler HK (1987) Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Meth Enzymol 148:350–382
Liu F, Stützel H (2004) Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Sci Hortic 102:15–27
Liu F, Andersen MN, Jacobsen SE, Jensen CR (2005) Stomatal control and water use efficiency of soybean (Glycine max L. Merr.) during progressive soil drying. Environ Exp Bot 54:33–40
Liu H, Wang X, Wang D, Zou Z, Liang Z (2011) Effect of drought stress on growth and accumulation of active constituents in Salvia miltiorrhiza Bunge. Ind Crops Prod 33:84–88
Llusia J, Peñuelas J (1998) Changes in terpene content and emission in potted Mediterranean woody plants under severe drought. Can J Bot 76:1366–1373
Loreto F, Schnitzler JP (2010) Abiotic stresses and induced BVOCs. Trends Plant Sci 15:154–166
Loreto F, Fischbach RJ, Schnitzler JP, Ciccioli P, Brancaleoni E, Calfapietra C, Seufer G (2001) Monoterpene emission and monoterpene synthase activities in the Mediterranean evergreen oak Quercus ilex L. grown at elevated CO2 concentrations. Glob Change Biol 7:709–717
Loreto F, Dicke M, Schnitzler JP, Turlings TCJ (2014) Plant volatiles and the environment. Plant Cell Environ 37:1905–1908
Lovisolo C, Perrone I, Carra A, Ferrandino A, Flexas J, Medrano H, Schubert A (2010) Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non hydraulic interactions at the whole plant level: a physiological and molecular update. Funct Plant Biol 37:98–116
Lubbe A, Verpoorte R (2011) Cultivation of medicinal and aromatic plants for specialty industrial materials. Ind Crops Prod 34:785–801
Maatallah S, Nasri N, Hajlaoui H, Albouchi A, Elaissi A (2016) Evaluation changing of essential oil of laurel (Laurus nobilis L.) under water deficit stress conditions. Ind Crops Prod 91:170–178
Medrano H, Tomas M, Martorell S, Flexas J, Hernandez E, Rossello J, Pou A, Escalona JM, Bota J (2015) From leaf to whole-plant water use efficiency (WUE) in complex canopies: limitations of leaf WUE as a selection target. Crop J 3(3):220–228
Monclus R, Dreyer E, Villar M, Delmotte FM, Delay D, Petit JM, Barbaroux C, Thiec D, Bréchet C, Brignolas F (2006) Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides × Populus nigra. New Phytol 169:765–777
Munné-Bosch S, Peñuelas J (2003) Photo- and antioxidative protection, and a role for salicyclic acid during drought and recovery in field-grown Phillyrea angustifolia plants. Planta 217:758–766
Munné-Bosch S, Mueller M, Schwarz K, Alegre L (2001) Diterpenes and antioxidative protection in drought-stressed Salvia officinalis plants. J Plant Physiol 11:1431–1437
Niinemets Ü, Loreto F, Reichstein M (2004) Physiological and physicochemical controls on foliar volatile organic compound emissions. Trends Plant Sci 9:180–186
Niinemets Ü, Kännaste A, Copolovici L (2013) Quantitative patterns between plant volatile emissions induced by biotic stresses and the degree of damage. Front Plant Sci 4:262
Nogués S, Allen DJ, Morison JIL, Baker NR (1998) Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants. Plant Physiol 117:173–181
Nogués I, Muzzini V, Loreto F, Bustamante MA (2015) Drought and soil amendment effects on monoterpene emission in rosemary plants. Sci Total Environ 538:768–778
Novak J (2017) Letter to the Editor on “How to implement GACP of MAPs? A practical implementation guide to Good Agricultural and Wild Collection Practices (GACP)”. J Appl Res Med Aromat Plants. https://doi.org/10.1016/j.jarmap.2016.12.003
Nowak M, Kleinwächter M, Manderscheid R, Weigel HJ, Selmar D (2010) Drought stress increases the accumulation of monoterpenes in sage (Salvia officinalis), an effect that is compensated by elevated carbon dioxide concentration. J Appl Bot Food Qual 83:133–136
Oren R, Sperry JS, Katul GG, Pataki DE, Ewers BE, Phillips N, Schäfer KVR (1999) Survey and synthesis of intra- and interspecific variation in stomatal sensitivity to vapour pressure deficit. Plant Cell Env 22:1515–1526
Ormeño E, Mévy JP, Vila B, Bousquet Mélou A, Greff S, Bonin G, Fernandez C (2007) Water deficit stress induces different monoterpene and sesquiterpene emission changes in Mediterranean species. Relationship between terpene emissions and plant water potential. Chemosphere 67:276–284
Ormeño E, Romain O, Mevy JP, Baldy V, Fernandez C (2009) Compost may affect volatile and semi-volatile plant emissions through nitrogen supply and chlorophyll fluorescence. Chemosphere 77:94–104
Paquin R, Mehuys GR (1980) Influence of soil moisture on cold tolerance of alfalfa. Can J Plant Sci 60:139–147
Pastenes C, Pimentel P, Lillo J (2005) Leaf movements and photoinhibition in relation to water stress in field-grown beans. J Exp Bot 56:425–433
Paulsen J, Selmar D (2016) The difficulty of correct reference values when evaluating the effects of drought stress: a case study with Thymus vulgaris. J Appl Bot Food Qual 89:287–289
Petropoulos SA, Daferera D, Polissiou MG, Passam HC (2008) The effect of water deficit stress on the growth, yield and composition of essential oils of parsley. Sci Hortic 115:393–397
Pirbalouti AG, Samani MR, Hashemi M, Zeinali H (2014) Salicylic acid affects growth, essential oil and chemical compositions of thyme (Thymus daenensis Celak.) under reduced irrigation. Plant Growth Regul 72:289–301
Pistelli L, Noccioli C, D’Angiolillo F, Pistelli L (2013) Composition of volatile in micropropagated and field grown aromatic plants from Tuscany Islands. Acta Biochim Pol 60:43–50
Possell M, Loreto F (2013) The role of volatile organic compounds in plant resistance to abiotic stresses: responses and mechanisms. In: Biology, controls and models of tree volatile organic compound emissions (Eds Ü. Niinemets, RK Monson). Springer, Berlin, pp 209–235
Radwan A, Kleinwachter M, Selmar D (2017) Impact of drought stress on specialised metabolism: biosynthesis and the expression of monoterpene synthases in sage (Salvia officinalis). Phytochemistry 141:20–26
Raut JS, Karuppayil SM (2014) A status review on the medicinal properties of essential oils. Ind Crops Prod 62:250–264
Rauter AP, Dias C, Martins A, Branco I, Neng NR, Nogueira JM, Goulart M, Silva FVM, Justino J, Trevitt C, Waltho JP (2012) Non-toxic Salvia sclareoides Brot. extracts as a source of functional food ingredients: phenolic profile, antioxidant activity and prion binding properties. Food Chem 132:1930–1935
Rinnan R, Steinke M, McGenity T, Loreto F (2014) Plant volatiles in extreme terrestrial and marine environments. Plant Cell Environ 37:1776–1789
Ruberto G, Baratta MT (2000) Antioxidant activity of selected essential oil components in two lipid model systems. Food Chem 69:167–174
Schachtman DP, Goodger JQD (2008) Chemical root to shoot signaling under drought. Trends Plant Sci 13:281–287
Scholander PF, Hammel HT, Bradstreet ED, Hemmingsen EA (1965) Sap pressure in vascular plants. Negative hydrostatic pressure can be measured in plants. Science 148:339–346
Selmar D, Kleinwachter M (2013) Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind Crops Prod 42:558–666
Singleton VL, Rossi JAJ (1965) Colorimetry of total phenolics with phosphomolybdic–phosphotungstic acid reagent. Am J Enol Vitic 16:144–158
Skirycz A, Inzé D (2010) More from less: plant growth under limited water. Curr Opin Biotechnol 21:197–203
Steinbrecher F, Hauff K, Hakola H, Rössler J (1999) A revised parameterization for emission modeling of isoprenoids for boreal plants. In: Laurilla T, Lindfors V (eds) Biogenic VOC emissions and phytochemistry in the boreal regions of Europe air pollution research report 70. Commission of European Communities, Luxembourg, pp 29–43
Swigar AA, Silverstein RM (1981) Monoterpenes. Aldrich, Milwaukee
Szôllôsi R, Szôllôsi Varga I (2002) Total antioxidant power in some species of Labiatae (adaptation of FRAP method). Acta Biol Szegediensis 46:125–127
Turtola S, Manninen A, Rikal R, Kainulainen P (2003) Drought stress alters the concentration of wood terpenoids in scots pine and Norway spruce seedling. J Chem Ecol 29:1981–1995
Viera HJ, Bergamaschi H, Angelocci LR, Libardi PL (1991) Performance of two bean cultivars under two water availability regimes. II. Stomatal resistance to vapour diffusion, transpiration flux density and water potential in the plant (in Portugal). Pesqui Agropecu Bras 9:1035–1045
Zhao J, Davis LC, Verpoorte R (2005) Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol Adv 23:283–333
Acknowledgements
This research was partially funded by the INTERREG-ALCOTRA 2007–2013 Project “AROMA” (n. 68). Authors acknowledged Claudio Cervelli and Paolo Lo Turco for plant furnishing and multiplication.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Caser, M., D’Angiolillo, F., Chitarra, W. et al. Ecophysiological and phytochemical responses of Salvia sinaloensis Fern. to drought stress. Plant Growth Regul 84, 383–394 (2018). https://doi.org/10.1007/s10725-017-0349-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-017-0349-1