
Abstract
Drought stress encumbers the seed germination and delays seedling establishment in dry direct-seeded rice. Pot and field studies were carried out to ascertain the role of seed-priming on emergence, seedling growth and associated metabolic events in dry-direct seeded rice system. Seeds of two indica rice cultivars were subjected to different priming agents viz., hydropriming (H2O), potassium nitrate (KNO3; 0.5 %), polyethelene glycol-6000 (PEG; 10 %) and spermidine (Spd; 0.5 mmol L−1). A no-priming treatment was kept as control for comparison. Drought stress was imposed by 15 % PEG solution in a pot experiment; whilst in field trial soil moisture contents were maintained between 15 and 18 %. In pot experiment, drought stress severely hampered the germination rate, seedling growth, and starch metabolism, but increased the antioxidant enzymes activity and lipid peroxidation in both rice cultivars as compared with normal conditions. All the seed priming treatments particularly Spd priming, were effective in alleviating the damaging effects of drought stress under controlled as well as field conditions. In field trial, Spd priming recorded 21, 232, 173, 67 and 78 % higher emergence, shoot length, shoot fresh weight, maximum root length and root fresh weight of dry direct-seeded rice, respectively, compared with control. And such increments were associated with better starch metabolism particularly increased α-amylase activity in primed rice seedlings.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Rice is a major staple food crop for nearly 65 % population in China (Zhang et al. 2005). China plays a key role in the global food security as it produces more than 28 % of rice production worldwide. However, burgeoning and ever increasing population necessitates further increase in rice production to ensure future food security and social stability. Traditionally, transplanted flooded rice is the major production system in China accounting nearly 95 % of the total rice grown area (Peng et al. 2009). Nonetheless in recent years, depleting water resources, water-intensive nature of rice cultivation and labor shortage are threatening the sustainability and productivity of transplanted-flooded rice (Sun et al. 2015). Dry direct-seeded rice technology has been proposed to reduce water requirement, save labor demand, increase resource use efficiency and improve environmental sustainability (Liu et al. 2014, 2015). It refers to the process of establishing the crop from dry seeds sown in the field rather than transplanting seedlings from nursery (Liu et al. 2014). Previous researches have recorded similar grain yield in dry direct-seeded rice and traditionally transplanted flooded rice (Qureshi et al. 2006; Liu et al. 2015).
Poor germination and stand establishment of direct-seeded rice is a major deterrent for achieving an optimal crop growth and better productivity especially under drought stress during emergence (Liu et al. 2014). Severe drought stress sometimes may result in complete inhibition of seedling emergence (Kaya et al. 2006). Decreased water uptake during imbibition phase is the major cause for the poor stand establishment (Murillo-Amador et al. 2002). Drought induces various changes in morphological, metabolic and/or physiological functions of plant. It severely hampers both the elongation and expansion in plant growth (Kusaka et al. 2005). Moreover, exposure of plants to drought stress leads to the generation of reactive oxygen species (ROS), which can damage plants by lipid peroxidation, protein degradation, DNA fragmentation and ultimately cell death (Zhang et al. 2015). However, activation of antioxidant system and synthesis of metabolites ensure the tolerance ability of plants in stress prone environments.
The oxidative stress tolerance is genetically controlled for crop improvement using conventional breeding and selection, transgene production or adopting physiological approaches like seed-priming. Seed priming is a controlled hydration technique that allows the pre-germination metabolisms without actual germination (Hussain et al. 2015b). It is one of the most effective, pragmatic and short-term approaches for increasing seed vigor, synchronization of germination, as well as the seedlings growth of many crops under different abiotic stresses including drought (Kaya et al. 2006; Farooq et al. 2009). Higher and synchronized germination of primed seeds (Kaya et al. 2006) primarily occurred due to reduction in the lag time of imbibitions (Brocklehurst and Dearman 2008), build-up of germination enhancing metabolites (Farooq et al. 2007), metabolic repair during imbibition (Bray et al. 1989) and osmotic adjustment (Bradford 1986).
Polyethylene glycol (PEG) and Potassium nitrate (KNO3) are large in molecular size and nontoxic in nature, therefore categorized as osmopriming reagents. Both of the above reagents can lower the water potential without penetrating into seeds on soaking. Osmopriming with PEG has been proposed to improve seed germination, early seedling vigor, antioxidant enzymes activity, and eventually leading to increased stress tolerance in many crop plants (Chen and Arora 2011). Likewise, Kaya et al. (2006) reported that KNO3 priming in sunflower improved germination and stand establishment under drought and salinity stresses. Spermidine (Spd) is a polyamine, which can act as free radical scavengers and protects the membranes from oxidative damages (Besford et al. 1993). Exogenous application of Spd has been reported to improve the drought tolerance in plants (Li et al. 2015; Yin et al. 2014). Recently, it has been found that endogenous production of Spd was associated with improved drought tolerance in rice (Yang et al. 2007; Farooq et al. 2009).
Climate change has been typically summarized by varying precipitation patterns potentially leading to more frequent and severe drought episodes. Approximately one-third of the world land area is prone to drought, and this ratio is higher up to 47 % in China (Wei et al. 1997). Rice, being a high delta crop, is a key target for water saving and developing novel strategies for rice production and is inevitable to face the consequences of future climate change on agriculture and natural ecosystems (Hussain et al. 2015a). Despite the availability of volumetric information on rice seed invigoration techniques, priming agents like Spd, PEG and KNO3 have rarely been tried in dry direct-seeded rice under drought stress. The aims of this study were: (1) to examine the effects of seed priming on emergence and early seedling growth of two rice cultivars in dry direct-seeded rice system under drought stress; (2) to investigate the biochemical changes in rice seedlings induced by seed priming under controlled and open-field conditions.
Materials and methods
Seed source
Seeds of two widely grown indica rice cultivars viz., Huanghuazhan (HHZ, inbred) and Yangliangyou6 (YLY6, hybrid) were obtained from Crop Physiology and Production Center, Huazhong Agricultural University, Wuhan, China. The initial seed moisture contents of HHZ and YLY6 were 10.8 and 11.0 %, respectively. The seeds were placed in nylon mesh bags and dried at ambient temperature in silica gel drier. The dry seeds of HHZ and YLY6 contained moisture contents of 9.07 and 8.98 % (on dry weight basis), respectively. After drying, seeds were sealed in aluminum foil bags for subsequent usage.
Seed priming treatments
Seed priming treatments were pre-optimized in various preliminary studies. Seeds of both rice cultivars were treated with various concentrations of KNO3, Polyethelene glycol-6000 (PEG) and Spd. Effective levels of these treatments were selected based on emergence, early seedling growth and various biochemical attributes under drought (data not shown). Seed priming treatments selected for present studies were: hydropriming (distilled water), KNO3 (0.5 %), PEG (10 %) and Spd (0.5 mmol L−1). Seeds were primed in dark at 25 °C for 24 h, with constant gentle agitation. The ratio of seed weight to solution volume (w/v) was 1:5 and priming solution was changed after every 12 h. The primed seeds were washed with distilled water for 2 min, surface dried and transferred to air dried oven at 25 °C for 48 h to reduce the moisture contents near to 10 %. The seeds were sealed in polythene bags and stored in a refrigerator at 4 °C until use.
Experimentation
Pot experiment
To ascertain the germination and growth responses of rice to different seed priming treatments under PEG-induced drought stress, a pot experiment was conducted in Crop Physiology and Production Center, Huazhong Agricultural University, Wuhan, China. Plastic pots with 23.0 cm × 17.0 cm × 15.0 cm size were filled with 3 l of distilled water for control (CK) and of 15 % PEG solution (w/v) for drought stress treatments. Floating board on the surface of solution contained five separated sections, and thirty seeds of a treatment were sown in each section. The experiment was laid out in a completely randomized design with factorial arrangement replicated thrice. All the pots were placed in growth chamber with 24 h dark period for first 2 days. However, for the rest of study period, growing conditions were kept uniform with 12 h light period, 30 °C day and 25 °C night temperatures.
Germination of seeds was recorded on daily basis according to AOSA (1990) until a constant count was achieved. Seeds were considered to be germinated when radicle and hypocotyl length exceeded 2 mm. Seedlings were harvested at 8 days after sowing (DAS), and shoot length and maximum root length of ten randomly selected seedlings per replication were measured and averaged. Seedlings of each replicate were dissected into roots and shoots and their fresh weights were recorded immediately. Then the seedling samples were put in a disposable ziplock bag and were stored in −80 °C refrigerator for analysis of malondialdehyde (MDA) contents and antioxidant enzymes activities.
To extract antioxidant enzymes, 0.5 g fresh shoot samples in each container were ground using a tissue grinder in 8 ml of 50 mM cool phosphate buffer [pH 7.0, containing 1 % (w/v) polyvinylpyr- rolidone] in tubes that were placed in an ice bath. The homogenate was centrifuged at 15,000×g for 20 min at 4 °C. The supernatant was used for assays of enzyme activity and the level of lipid peroxidation.
The lipid peroxidation level was determined in terms of malondialdehyde (MDA) content by the method of Dhindsa et al. (1981) and Zhang and Kirkham (1994). A 2-ml aliquot of enzyme solution was added to a tube containing 1 ml 20 % (v/v) trichloroacetic acid and 0.5 % (v/v) thiobarbituric acid. The mixture was heated in a water bath at 95 °C for 20 min, cooled to room temperature and then centrifuged at 10,000×g for 10 min. The absorbance of supernatant at 532 nm was determined and the nonspecific absorbance at 600 nm was subtracted. The MDA content was calculated by the extinction coefficient of 155 mM−1 cm−1 (Heath and Packer 1968).
The CAT activity determination was performed according to the method of Beers and Sizer (1952) with slight modifications. The reaction mixture consisted of 2 ml of sodium phosphate buffer (50 mM, pH 7.0), 0.5 ml H2O2 (40 mM) and 0.5 ml enzyme extract in a total volume of 3.0 ml. The decomposition of H2O2 was measured by decline in absorbance at 240 nm (Tecan infinite M200, Swit). One unit CAT activity was defined as an absorbance change of 0.01 units per min and demonstrated as U g−1 FW.
The POD activity was determined by the method of guaiacol oxidation according to description of Chance and Maehly (1995) with modifications. The POD reaction solution (3 ml) contained 50 mM sodium acetate buffer (pH 5.0), 20 mM guaiacol, 40 mM H2O2, and 0.1 ml enzyme extract. The reaction mixture in which enzyme solution was replased with sodium acetate buffer served as a blank. Readings at 470 nm (Tecan infinite M200, Swit) were recorded within 3 min after the start of the reaction at 1 min intervals. One unit POD activity was defined as an absorbance change of 0.01 units per min and was expressed as U g−1 FW.
The activity of SOD was determined by measuring its ability to inhibit the photoreduction of nitro blue tetrazolium (NBT) following the method of Giannopolitis and Ries (1977) with slight modifications. The reaction mixture consisted of 1.5 ml phosphate buffer (50 mM, pH 7.8), 0.3 ml methionine (130 mM), 0.3 ml NBT (750 μM), 0.3 ml EDTA (100 μM), 0.3 ml riboflavin (20 μM) and 0.3 ml enzyme extract in a total volume of 3.0 ml. Test tubes containing the reaction solution were irradiated under a light bank (15 fluorescent lamps) at 78 μmol m−2 s−1 for 15 min. The absorbance of the irradiated and nonirradiated solution at 560 nm was determined with a spectrophotometer (Tecan infinite M200, Swit). One unit of SOD activity was defined as the amount of enzyme that gives 50 % inhibition of NBT photoreduction and was presented as U g−1 FW.
A separate set of each treatment was maintained for recording α-amylase activity and total soluble sugar contents at 5 DAS. For α-amylase activity, 1.0 g seedling sample including shoot and root was ground and mixed with 100 ml distilled water, and left for 24 h at 4 °C. The enzyme activity was determined from supernatant liquid by dinitrosalicyclic acid (DNS) method (Bernfeld 1955). In order to determine total soluble sugar contents, ground seedling sample (1.0 g) was mixed with 10 ml distilled water, and left for 24 h at 25 °C (Lee and Kim 2000). Mixture was filtered with Whatman No. 42 and the final volume was made to 10 ml with distilled water. Total soluble sugar contents were determined by the phenol sulfuric method (Dubois et al. 1956).
Field experiment
In order to assess the influence of seed priming on dry direct-seeded rice under field conditions, a trial was carried out at the experimental station of Zhangbang Village, Dajin Town, Wuxue Country, Hubei Province, China (29°51′N 115°33′E). The soil of experimental field was silt loam with the proportion of sand, silt and clay as 26, 64 and 10 %, respectively. The organic matter, total nitrogen, available phosphorus, and potassium contents of upper 20 cm soil were 30.7, 1.98 g kg−1, 23.5, and 161.5 mg kg−1, respectively.
The experiment was randomized in a split plot design with four replications. Two rice cultivars (HHZ and YLY6) were assigned to main plot, while various seed priming treatments (control, hydropriming, 0.5 % KNO3 priming, 10 % PEG priming, 0.5 mmol L−1 Spd priming) were kept in subplots. Before sowing, the soil was dry ploughed and harrowed without puddling. Dry seeds were manually sown at 20 cm row to row and 5 cm plant to plant distance on 29th June 2014. The seeds were covered with soil immediately after sowing. The soil moisture contents were in the range of 15–18 % during the course of study.
Emergence of seeds was recorded on daily basis according to AOSA (1990) and was expressed as percentage. At 8 DAS, the seedlings of 1.0 m row were sampled carefully, washed and kept in a disposable ziplock bag for subsequent analysis. After measuring shoot length and maximum root length of 10 randomly selected plants, all seedlings were dissected into root and shoot for determination of their fresh weights. All the samples were stored in liquid N2 and were kept in −80 °C refrigerator for further analysis of α-amylase activity and total soluble sugar contents.
Statistical analysis
Data were analyzed to confirm its variability following analysis of variance using Statistix 9.0. The differences between treatments were separated using least significance difference (LSD) test at 0.05 probability level.
Results
Pot experiment
Significant (p ≤ 0.05) variations in germination of rice were observed under the influence of PEG-induced drought stress, seed priming treatments and cultivars (Fig. 1). Drought stress drastically reduced germination speed and rate in both rice cultivars. Nevertheless, all the seed priming treatments significantly (p ≤ 0.05) enhanced the rice germination compared with control under drought as well as normal conditions (Fig. 1). Under normal conditions, the maximum germination of HHZ (98.9 %) and YLY6 (96.7 %) was recorded by Spd priming. Seed priming with PEG and hydropriming were the 2nd best treatments for HHZ and YLY6, respectively (Fig. 1). Under drought stress, HHZ primed with KNO3, PEG, and Spd recorded 88–93 % germination at 6 DAS, which were significantly (p ≤ 0.05) higher than that recorded in control (62 %). Similar was the case for YLY6 and all seed priming treatments recorded more than 79 % germination under drought conditions. Seed priming of rice also enhanced the speed of germination, therefore germination of primed seeds at 2 DAS under drought stress was in the range of 13.3–24.4 %, as compared to no germination for control treatments of both cultivars (Fig. 1).

Germination dynamics in primed and non-primed seeds of HHZ and YLY6 under normal (a, b) and PEG-induced drought stress (c, d) in a pot experiment. Error bars represent the standard error
Drought stress markedly inhibited the early seedling growth of rice, and considerable reductions in shoot and root length as well as in their fresh weights were recorded for both cultivars (Fig. 2). However, different seed priming treatments assuaged the damaging effects of PEG-induced drought stress. Averaged across cultivars and seed priming reagents; shoot length, maximum root length, shoot fresh weight and root fresh weight of primed rice seeds grown under drought stress, were increased by 54.1, 32.3, 35.5 and 45.9 %, respectively compared with control. Seed priming with Spd performed better than all other seed priming treatments by recording 64.9, 51.8, 36.8 and 76.6 % increase in shoot length, maximum root length, shoot fresh weight and root fresh weight, respectively (Fig. 2). Among all the seed priming treatments, PEG priming was least effective under drought stress, but it still presented a better growth than control (Fig. 2). Variations between cultivars were also apparent regarding their response to different seed priming treatments under drought stress, and the effects of seed priming treatments on growth of YLY6 were higher than that of HHZ. For YLY6, all the priming treatments significantly (p ≤ 0.05) improved the growth performance under drought stress as compared with control, while this was not in the case of HHZ, where only hydropriming, KNO3 and Spd could significantly (p ≤ 0.05) enhance rice growth under drought stress (Fig. 2).

The maximum root length, shoot length, root fresh weight and shoot fresh weight of primed and non-primed seedlings of HHZ and YLY6 under normal condition (a–d)and PEG-induced drought stress (e–h) in a pot experiment. Error bars represent the standard error. The parameters were measured at 8 days after sowing. The means for each cultivar under each growing medium not sharing a common letter are significantly different at 0.05 probability level according to least significant difference (LSD)
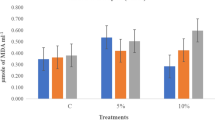
Lipid peroxidation was assessed in terms of MDA contents in rice seedlings. The highest MDA contents in the seedlings of HHZ (155.5 nmol g−1 FW) and YLY6 (142.3 nmol g−1 FW) were observed in control treatments, grown under drought stress (Fig. 3). Variations among seed priming treatments for MDA contents were also observed even under normal condition and primed seedlings recorded significantly (p ≤ 0.05) lower MDA contents. Regardless of growing media and cultivars, Spd priming recorded 25 % lower MDA contents and was the most effective treatment for reducing lipid peroxidation compared with control (Fig. 3).

The Malondialdehyde (MDA) content, Catalase (CAT) activity, Peroxidase (POD) activity and Superoxide dismutase (SOD) activity in primed and non-primed seedlings of HHZ and YLY6 under normal condition (a–d)and PEG-induced drought stress (e–h) in a pot experiment. Error bars represent the standard error. The parameters were measured at 8 days after sowing. The means for each cultivar under each growing medium not sharing a common letter are significantly different at 0.05 probability level according to least significant difference (LSD)
Antioxidant enzymes (SOD, POD and CAT) activities in rice seedlings varied significantly (p ≤ 0.05) in response to different seed priming treatments and drought stress in both cultivars (Fig. 3). Drought stress considerably increased the antioxidant enzymes activities in both rice cultivars predominately in HHZ. Moreover, seed priming of both rice cultivars also increased the activities of SOD, POD, and CAT compared with control. The maximum activities of SOD (989.5 U g−1 FW), POD (1069.0 U g−1 FW), and CAT (903.4 U g−1 FW) were recorded by Spd priming in HHZ cultivar. Rice cultivars showed different responses to various seed priming treatments, for example, POD activities of HHZ seedlings primed with distilled water and KNO3 were similar (p > 0.05) with control under normal conditions. In contrast, all seed priming treatments significantly (p ≤ 0.05) enhanced the POD activity in YLY6 (Fig. 3).
Starch metabolism in rice seedlings was assessed in terms of α-amylase activity and total soluble sugar contents (Fig. 4). Exposure of drought stress reduced the total soluble sugar contents in both cultivars, while α-amylase activity in HHZ. Nevertheless, all seed priming treatments significantly (p ≤ 0.05) improved the α-amylase activity (Fig. 4a, c) and soluble sugar contents (Fig. 4b, d) in both cultivars under drought as well as normal conditions. Averaged across cultivars and seed priming reagents, α-amylase activity and total soluble sugar contents of primed rice seedlings were increased by 103.6 and 44.4 % compared with control. Both cultivars responded differentially to drought stress and seed priming treatments. Drought stress significantly (p ≤ 0.05) reduced α-amylase activity (Fig. 4a, c) and total soluble sugar contents (Fig. 4b, d) in HHZ and Spd priming was the most effective treatment in improving these two traits under drought as well as normal conditions. In contrast, increase in α-amylase activity expect for PEG primed seedlings and decrease in total soluble sugars under drought were observed in YLY6. Although, all the seed priming treatments increased the α-amylase activity and total soluble sugars in YLY6 as compared with control, yet hydropriming and KNO3 were relatively more effective for these attributes.

The α-amylase activity and total soluble sugars in primed and non-primed seedlings of HHZ and YLY6 under normal condition (a, b) and PEG-induced drought stress (c, d) in a pot experiment. Error bars represent the standard error. The α-amylase activity and soluble sugars were measured at 5 days after sowing. a FW fresh weight. The means for each cultivar not sharing a common letter are significantly different at 0.05 probability level according to least significant difference (LSD)
Field experiment
Temporal data regarding emergence of dry direct-seeded rice under drought stress were presented in Fig. 5. Consistent with pot experiment, all seed priming treatments notably enhanced emergence of both rice cultivars. When averaged across cultivars, the emergence in control plots at 6 DAS was only 4 %, while average emergence of primed seeds was 31 %. The Spd priming showed the highest emergence in both cultivars followed by PEG priming. Whilst hydropriming and KNO3 priming were moderately effective regarding emergence of dry direct-seeded rice (Fig. 5).

The emergence of primed and non-primed seeds of HHZ (a) and YLY6 (b) in an open-field experiment under dry direct-seeded rice system. Error bars represent the standard error
Seed priming treatments depicted significant (p ≤ 0.05) improvement in early seedling growth of dry direct-seeded rice in both cultivars (Fig. 6). Furthermore, response of two cultivars to seed priming treatments also varied for early seedling growth attributes. In HHZ, the highest values for shoot length (4.8 cm), maximum root length (6.0 cm), shoot fresh weight (29.2 mg seedling−1) and root fresh weight (13.4 mg seedling−1) were observed by Spd priming (Fig. 6). Seed priming with KNO3 was least effective in HHZ for these attributes. In YLY6, Spd priming was more beneficial in terms of seedling growth but it was similar (p > 0.05) with PEG and KNO3 priming for root growth attributes (Fig. 6).

The maximum root length (a), root fresh weight (b), shoot length (c), and shoot fresh weight (d) of primed and non-primed seedlings of HHZ and YLY6 in an open-field experiment under dry direct-seeded rice system. The parameters were recorded at 8 days after sowing. Error bars represent the standard error. The means for each cultivar not sharing a common letter are significantly different at 0.05 probability level according to least significant difference (LSD)
Different seed priming treatments also significantly (p ≤ 0.05) enhanced α-amylase activity and total soluble sugar contents in seedlings of both cultivars under dry direct-seeded rice system (Fig. 7). Averaged across cultivars, the α-amylase activity in control was only 5.27 mg g−1 min−1, while in different seed priming treatments; it ranged from 10.18 to 19.17 mg g−1 min−1. Likewise, total soluble sugar contents in different seed priming treatments were 55–79 % higher than that of control. Seed priming with Spd recorded higher α-amylase activity and total soluble sugar contents than all other seed priming treatments (Fig. 7).

The α-amylase activity (a) and total soluble sugars (b) in primed and non-primed seedlings of HHZ and YLY6 in an open-field experiment under dry direct-seeded rice system. The α-amylase activity and soluble sugars were measured at 5 days after sowing. Error bars represent the standard error. The means for each cultivar not sharing a common letter are significantly different at 0.05 probability level according to least significant difference (LSD)
Discussion
Present study demonstrated that seed priming efficiently improved emergence and seedling growth performance of dry direct-seeded rice under water deficit conditions. Exposure of drought stress in field conditions or in PEG solution culture resulted in impaired and unsynchronized germination as well as poor seedling establishment (Figs. 1, 5). Previously, Kaya et al. (2006) reported that reduced germination and poor seedling stand is the first and foremost response of plants to water shortage. While studying on pea, Okcu et al. (2005) reported hampered germination and early seedling growth of five tested cultivars under drought stress. Moreover in alfalfa crop, reduced germination potential, lower hypocotyl length, and less shoot and root fresh weights were recorded by PEG-induced drought stress (Zeid and Shedeed 2006).
In both studies (pot and field), seed priming treatments improved germination and early seedling growth performance of rice particularly under drought stress. Better ability of primed seeds to complete the germination process in a short time and cope with water-limited conditions might be attributed to readily available food for germinating seedlings. Wahid et al. (2007) stated that seed priming enhances the germination rate, speed and uniformity even under less than optimum conditions thus enabling the uniform and vigorous crop stand. Furthermore, early and uniformed stand establishment enables the crop to complete the other phonological events in crop ontogeny well in time (Farooq et al. 2009). Harris et al. (2002) observed that in drought-prone areas, primed rice seeds recorded better and uniform germination as well as faster seedlings growth resulting in higher yield.
Our results depicted that Spd priming was more effective for most of the studied characteristics than all other seed priming treatments. Our results are in agreement with Tang and Newton (2005), who indicated that seed priming with Spd improves cell membrane stability, increases antioxidant enzymes activities, decreases stress-induced growth inhibition, and enhances stress tolerance ability in plants. Yang et al. (2007) also pointed out that endogenous production of Spd was associated with improved drought tolerance in rice. While studying on hybrid sunflower, Farooq et al. (2007) observed that seed priming with Spd was effective in improving the emergence and seedling growth.
In pot study, PEG-induced drought stress led to higher MDA accumulation, whilst seed priming treatments effectively reduced it in both rice cultivars compared with control (Fig. 3). Gill and Tuteja (2010) noted that plants exposed to various abiotic stresses exhibited higher lipid peroxidation due to the generation of ROS. In present study, primed seedlings showed lower lipid peroxidation in normal as well as drought conditions (Fig. 3). The lowest MDA levels in Spd primed seedlings may justify the highest growth of these seedlings among all seed priming treatments. Therefore, lowering of ROS levels and consequently lipid peroxidation might be a major factor in the drought tolerance attained by primed seedlings. Decreased integrity of biological membranes, principally due to the oxidative damage is another intricate effect of drought stress (Farooq et al. 2009) and production of ROS, especially H2O2 is a major cause of such effects (Munné-Bosch and Penuelas 2003). The ROS in plants are scavenged by a variety of antioxidant enzymes including SOD, POD, and CAT (Khaliq et al. 2015). Antioxidant activity was increased in both rice cultivars in response to PEG-induced drought stress (Fig. 3), which could be involved in scavenging ROS produced under stress conditions. The SOD plays a key role in catalysing the dismutation of O2 −, while CAT and POD mainly scavenge H2O2 (Foyer et al. 1994). Ostrovskaya et al. (2009) argued that increased SOD activity under stress conditions is considered a protective mechanism against the formation of superoxide. Furthermore, increase in POD (Liu and Huang 2000) and CAT (Demiral and Turkan 2004) in response to abiotic stresses has also been documented. All the seed priming treatments enhanced the activities of SOD, CAT, and POD under normal as well as drought conditions (Fig. 3), which is in line with the results of Goswami et al. (2013).
The ability of plants to degrade starch into soluble sugars under water limited conditions probably plays a key role in their ability to survive and grow faster under stress. In rice, amylase activity is highly induced during germination (Hussain et al. 2015b); however, this activity begins to appear even earlier, during seed maturation. Present study highlighted the importance of α-amylase activity and total soluble sugar contents during germination and early seedling growth under drought conditions. Drought stress considerably decreased the total soluble sugars in both cultivars and α-amylase activity in HHZ, which could be explained by the fact that decrease in water availability slowed down the starch degradation under drought stress. Therefore, delay in germination/emergence and slow seedling growth was observed under drought stress.
Seed priming treatments recorded pronounced increase in α-amylase activity and total soluble sugar contents under normal and drought conditions (Figs. 4, 7). Higher α-amylase activity in primed seedlings was consistent with the better germination/emergence and faster growth compared with that in control (Figs. 1, 5, 6). Furthermore, greater α-amylase activity in primed seeds was also reflected through higher soluble sugar concentrations (Figs. 4, 7) and faster rate of starch breakdown in germinating primed seeds, which presumably provided the substrates necessary for generating the energy required for growth and maintenance processes (Hussain et al. 2015b).
Conclusively, studies conducted under controlled and field conditions revealed that seed priming can alleviate the negative effects of drought stress on emergence as well as seedling establishment and growth of dry-direct seeded rice. Seed priming with Spd was found to be more effective among all treatments to thrive under drought. Better emergence and vigorous seedling growth due to seed priming was associated with increased α-amylase activity in primed seedlings.
References
Association of Official Seed Analysis (AOSA) (1990) Rules for testing seeds. J Seed Technol 12:1–112
Beers RF, Sizer IW (1952) A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J Biol Chem 95:133–140
Bernfeld P (1955) Methods in enzymology. Academic Press, New York, pp 149–158
Besford RT, Richardson CM, Campos JL, Tiburcio AF (1993) Effects of polyamines on stabilization of molecular complexes in thylakoid membranes of osmotically stressed oat leaves. Planta 189:201–206
Bradford KJ (1986) Manipulation of seed water relations via osmotic priming to improve germination under stress conditions. Hort Sci 21:1105–1112
Bray CM, Davision PA, Ashraf M, Taylor RM (1989) Biochemical changes during osmopring of leak seeds. Ann Bot 6:185–193
Brocklehurst PA, Dearman J (2008) Interaction between seed priming treatments and nine seed lots of carrot, celery and onion II. Seedling emergence and plant growth. Ann Appl Biol 102:583–593
Chance B, Maehly AC (1995) Assay of catalase and peroxidase. Meth Enzymol 2:764–775
Chen K, Arora R (2011) Dynamics of the antioxidant system during seed osmopriming, post-priming germination, and seedling establishment in spinach (Spinacia oleracea). Plant Sci 180:212–220
Demiral T, Turkan I (2004) Does exogenous glycine betaine affect antioxidative system of rice seedlings under NaCl treatment? J Plant Physiol 161:1089–1100
Dhindsa RS, Dhindsa PP, Thorpe TA (1981) Leaf senescence: correlated with increased level of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J Exp Bot 32:93–101
Dubois M, Giles KA, Hamilton JJ, Roberes PA, Smith F (1956) Colorometric method for determination of sugars and related substances. Ann Chem 28:350–356
Farooq M, Basra SMA, Hussain M, Rehman H, Saleem BA (2007) Incorporation of polyamines in the priming media enhances the germination and early seedling growth in hybrid sunflower (Helianthus annuus L.). Int J Agric Biol 9:868–872
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009) Plant drought stress: effects, mechanisms and management. Agron Sustain Dev 29:185–212
Foyer CH, Descourvieres P, Kunert KJ (1994) Protection against oxygen radicals: an important defense mechanism studied in transgenic plants. Plant, Cell Environ 17:507–523
Giannopolitis CN, Ries SK (1977) Superoxide dismutase. I. Occurrence in higher plants. Plant Physiol 59:309–314
Gill SS, Tuteja N (2010) Reative oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Goswami A, Banerjee R, Raha S (2013) Drought resistance in seedlings conferred by seed priming. Protoplasma 250:1115–1129
Harris D, Tripathi RS, Joshi A (2002) On-farm seed priming to improve crop establishment and yield in dry direct-seeded rice. In: Pandey S (ed) Direct seeding: research strategies and opportunities. International Research Institute, Manila, pp 231–240
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Hussain S, Peng S, Fahad S, Khaliq A, Huang J, Cui K, Nie L (2015a) Rice management interventions to mitigate greenhouse gas emissions: a review. Environ Sci Pollut Res 22:3342–3360
Hussain S, Zheng M, Khan F, Khaliq A, Fahad S, Peng S, Huang J, Cui K, Nie L (2015b) Benefits of rice seed priming are offset permanently by prolonged storage and the storage conditions. Sci Rep 5:8101. doi:10.1038/srep08101
Kaya MD, Okçu G, Atak M, Çıkılı Y, Kolsarıcı Ö (2006) Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annus L.). Eur J Agron 24:291–295
Khaliq A, Aslam F, Matloob A, Hussain S, Geng M, Wahid A, Rehman H (2015) Seed priming with selenium: consequences for emergence, seedling growth, and biochemical attributes of rice. Biol Trace Elem Res. doi:10.1007/s12011-015-0260-4
Kusaka M, Ohta M, Fujimura T (2005) Contribution of inorganic components to osmotic adjustment and leaf folding for drought tolerance in pearl millet. Physiol Plant 125:474–489
Lee SS, Kim JH (2000) Total sugar, α-amylase activity, and emergence after priming of normal and aged rice seeds. Korean J Crop Sci 45:108–111
Li Z, Zhou H, Peng Y, Zhang XQ, Ma X, Huang LK, Yan Y (2015) Exogenously applied spermidine improves drought tolerance in creeping bentgrass associated with changes in antioxidant defense, endogenous polyamines and phytohormones. Plant Growth Regul 76:71–82
Liu XZ, Huang BR (2000) Heat stress injury in relation to membrane lipid peroxidation in creeping bentgrass. Crop Sci 40:503–510
Liu H, Hussain S, Zheng M, Sun L, Fahad S, Huang J, Cui K, Nie L (2014) Progress and constraints of dry direct-seeded rice in China. J Food Agric Environ 12:465–472
Liu H, Hussain S, Zheng M, Peng S, Huang J, Cui K, Nie L (2015) Dry direct-seeded rice as an alternative to transplanted-flooded rice in Central China. Agron Sustain Dev 35:285–294
Munné-Bosch S, Penuelas J (2003) Photo and antioxidative protection, and a role for salicylic acid during drought and recovery in fieldgrown Phillyrea angustifolia plants. Planta 217:758–766
Murillo-Amador B, Lopez-Aguilar R, Kaya C, Larrinaga-Mayoral J, Flores-Hernandez A (2002) Comparative effects of NaCl and polyethylene glycol on germination, emergence and seedling growth of cowpea. J Agron Crop Sci 188:235–247
Okcu G, Kaya MD, Atak M (2005) Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turk J Agric For 29:237–242
Ostrovskaya LK, Truch VV, Mikhailik OM (2009) Superoxide dismutase activation in response to lime-induced chlorosis. New Phytol 114:39–45
Peng SB, Tang QY, Zou YB (2009) Current status and challenges of rice production in China. Plant Prod Sci 12:3–8
Qureshi AS, Masih I, Turral H (2006) Comparing land and water productivities of transplanted and direct dry seeded rice for Pakistani Punjab. J Appl Irrig Sci 41:47–60
Sun L, Hussain S, Liu H, Peng S, Huang J, Cui K, Nie L (2015) Implications of low sowing rate for hybrid rice varieties under dry direct-seeded rice system in Central China. Field Crop Res 175:87–95
Tang W, Newton RJ (2005) Polyamines reduce salt-induced oxidative damage by increasing the activities of antioxidant enzymes and decreasing lipid peroxidation in Virginia pine. Plant Growth Regul 46:31–43
Wahid A, Perveen M, Gelani S, Basra SMA (2007) Pretreatment of seed with H2O2 improves salt tolerance of wheat seedlings by alleviation of oxidatice damage and expression of stress proteins. J Plant Physiol 164:283–294
Wei L, Jia L, Hu X, Zhao F (1997) Advances in studies on the physiology and biochemistry of maize drought resistance. Agric Res Arid Areas 15:66–71
Yang JC, Zhang JH, Liu K, Wang ZQ, Liu LJ (2007) Involvement of polyamines in the drought resistance of rice. J Exp Bot 58:1545–1555
Yin ZP, Li S, Ren J, Song XS (2014) Role of spermidine and spermine in alleviation of drought-induced oxidative stress and photosynthetic inhibition in China dwarf cherry (Cerasus humilis) seedlings. Plant Growth Regul 74:209–218
Zeid IM, Shedeed ZA (2006) Response of alfalfa to putrescine treatment under drought stress. Biol Plant 50:635–640
Zhang JX, Kirkham MB (1994) Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol 35:785–791
Zhang XF, Wang DY, Fang FP, Zhen YK, Liao XY (2005) Food safety and rice production in China. Res Agric Modern 26:85–88
Zhang M, Jin ZQ, Zhao J, Zhang GP, Wu FB (2015) Physiological and biochemical responses to drought stress in cultivated and Tibetant wild barley. Plant Growth Regul 75:567–574
Acknowledgments
This work is supported by the National Natural Science Foundation of China (Project No. 31371571), the Open Project Program of Key Laboratory of Crop Ecophysiology and Farming System, Ministry of Agriculture (Project No. 201301), the National Science & Technology Pillar Program (2013BAD20B06), and the Fundamental Research Funds for the Central Universities (Project No. 2013PY109).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Zheng, M., Tao, Y., Hussain, S. et al. Seed priming in dry direct-seeded rice: consequences for emergence, seedling growth and associated metabolic events under drought stress. Plant Growth Regul 78, 167–178 (2016). https://doi.org/10.1007/s10725-015-0083-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-015-0083-5