
Abstract
Vegetable oils are important components of the human diet, and are also actively used for industrial and cosmetic purposes and as biofuels. The scope of oil application largely depends on its composition. The fatty acid composition of oils, in its turn, is built upon the presence and activity of the enzymes of their biosynthesis. The presented review is an attempt to summarize and generalize information on the genetic control of fatty acid synthesis and the range of natural variation in their composition in key oilseed crops: peanut (Arachis hypogaea L.), sunflower (Helianthus annuus L.), flax (Linum usitatissimum L.), rapeseed (Brassica napus L.), camelina (Camelina sativa L.), cotton (Gossypium hirsutum L.), safflower (Carhtamus tinctorius L.), castor bean (Ricinus communis L.) etc., as well as approaches to changing the fatty acid percentage in plants, including gene silencing, overexpression of genes for the synthesis of fatty acids, changing the pattern of their expression, and genome editing.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Six annual oilseed crops are mainly used worldwide to produce food oil: soybean (Glycine max (L.) Merr.), rapeseed (Brassica napus (L.) subsp. oleifera Metzg.), cotton (Gossypium hirsutum L.), peanut (Arachis hypogaea L.), sunflower (Helianthus annuus L.), and maize (Zea mays L.) (Seiler and Jan 2010). In Russia, maize and soybean seeds are more used as a source of protein than for oil extraction. For food, the population traditionally prefers sunflower oil, and on a much smaller scale olive, rapeseed, soybean, flax, maize, mustard and camelina oils. Vegetable oils are used for the production of drying oil and paints as well as lubricants for machinery. Requirements for the quality and fatty acid (FA) composition of vegetable oils vary depending on their purpose. An increased oleic acid content impedes oil oxidation, which is important for extending its shelf life and vital for canning industry, fast-food chains (because oleic acid is more stable when fried) and biodiesel production (Gavrilova et al. 2020). Generally, oils with a high content of saturated FAs are resistant to oxidation but solid at ambient temperatures, while oils with a high polyunsaturated fatty acid (PUFA) content remain liquid at low temperatures but suffer from oxidative damage and free radical attack. Oils rich in monounsaturated oleic acid demonstrate a unique combination of good oxidative stability and lubricity at low temperatures (Vanhercke et al. 2013).
An increase in the content of linolenic acid reduces the oil’s resistance to oxidation during storage; however, paint and varnish manufacturers require varieties that produce drying oils with a high iodine number and linolenic acid content of up to 80%. Linolenic acid is the most unsaturated, which determines its high biological activity and the ability to quickly dry, and this makes such oil a leader in the production of paints, but on the other hand, leads to its rapid oxidation or rancidity, thus limiting the period of its use for food to two month (Porokhovinova et al. 2017). The oil in castor bean seeds is used to produce high-quality varnishes and paints, plastics, artificial fibers, printing inks, rubber, insulation materials, and toilet soaps. It is utilized in the textile and leather industries, not to mention the needs of perfumery and medicine (Moshkin 1980). Camelina oil contains eicosenoic acid, which makes it indispensable in cosmetology. Oils with higher levels of erucic and ricinoleic acids are used for the production of polyethylene. Pharmaceutics requires such acids as myristic, linolenic and nervonic. Searching for genotypes with a modified FA composition expands the range of oil applications. Myristic acid is also used on a large scale in the production of ice cream, vegetable milk, baby food, and as a substitute for cocoa butter in confectionery products (https://efko-ingredients.ru/press-centr/relizy/15532 of November 23, 2020; https://efko-ingredients.ru/produkciya/milk_fat/12606/ of November 23, 2020).
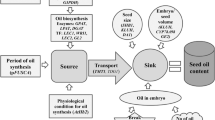
Biosynthesis of fatty acids
The main steps of FA synthesis occur in plastids and the endoplasmic reticulum (ER). The chart (Fig. 1) showing the synthesis stages was drawn up as a result of the analysis of scientific publications produced by various authors (Los 2001, 2014; Ohlrogge and Browse 1995; Dar et al. 2017; Bonaventure et al. 2003; Bao et al. 1998; Haslam and Kunst, 2013).*Footnote 1

FA biosynthesis begins in plastids, where acetyl-CoA carboxylase facilitates the conversion of acetyl-CoA into malonyl-CoA; then two carbon atoms are attached to the growing carbohydrate chain through FA synthase, using acetyl-CoA as a precursor, and malonyl-ACP as an elongator. This enzyme complex includes malonyl-CoA ACP-transacylase, which transfers the two-carbon malonyl unit to an acyl carrier protein (ACP), and the group of 3-ketoacyl-ACP synthase (KAS) enzymes is involved in the lengthening of the FA carbon chain, where malonyl-ACP serves as a C2 donor: KASIII uses acetyl-CoA as an acceptor for the synthesis of acetoacetyl-ACP. KASI acetylates 4:0-ACP to 16:0-ACP, and KAS II lengthens the 16:0 acceptor to 3-keto-18:0-ACP. The end products of KAS are 16:0-ACP and 18:0-ACP, which can be modified with soluble enzymes—palmitoleyl (16:0)-ACP desaturase or stearoyl (18:0)-ACP desaturase (controlled by the Fab2 gene, FA biosynthesis 2, or Sad2, Stearoyl ACP desaturase 2). These enzymes catalyze the formation of double bonds between the C9 and C10 carbon atoms in 16:0-ACP and 18:0-ACP to produce 16:1Δ9-ACP and 18:1Δ9-ACP (Los 2014; Ohlrogge and Browse 1995). The desaturase enzyme removes 2 hydrogen atoms and creates a double bond between carbon atoms. Desaturases are divided into two groups: Δ-desaturases and ω-desaturases. Delta indicates that a double bond evolves at a fixed position from the carboxyl end of the FA chain. Omega means that a double bond develops at a fixed position from the methyl end of the FA chain. Δ9-desaturase generates a double bond between the ninth and tenth carbon atoms from the carboxyl end, while ω3-desaturase (or Δ12-desaturase) creates a double bond between the third and fourth carbon atoms from the methyl end (Berestovoy et al. 2020). It is known that the synthesis of oleic acid occurs under the influence of one of the Δ9-desaturases (also known as stearoyl-CoA-desaturase-1) in chloroplasts, and then can be transported to the ER for subsequent desaturation in a lipid-bound form (Los and Murata 2004; Los 2001; Dar et al. 2017). All desaturases are encoded by the nuclear FAD (FA desaturase) or SAD (Stearate desaturase) (Perez-Vich et al. 2004). Interestingly, only the desaturases that introduce the first double bond in the chain of FAs bound to the acyl-carrying protein are soluble and localized in the chloroplast stroma, while the rest are anchored in the membrane and localized in the membranes of thylakoids or the ER (Los 2014). Palmitic, palmitoleic, stearic and oleic acids are freed from ACP for export to the cytosol using acyl-ACP thioesterases FatA and FatB, most active for C18-ACP and C16-ACP, respectively (Los 2014; Bonaventure et al. 2003; Perez-Vich et al. 2004).
Then, the synthesis of phosphatidic acid takes place mainly in the ER. Acyl-CoA:glycerol-3-phosphate acyltransferase, having membrane localization, transfers the generated acyl group to the sn-1 position on glycerol-3-phosphate. Acyl-CoA:lysophosphatidic acid acyltransferase catalyzes acylation at the sn-2 position. In most edible oils this position is occupied by the unsaturated C18 FAs (Los 2014).
Phosphatidic acid in chloroplasts is dephosphorylated by phosphatase to produce diacylglycerol (DAG), with the subsequent synthesis of glycolipids. In dicotyledons there are two pathways for the biosynthesis of glycerolipids: prokaryotic and eukaryotic. Both pathways begin with the stepwise acetylation of glycerol-3-phosphate and the formation of phosphatidic acid, whose dephosphorylation leads to the formation of DAG, which is used in plastids for assembling photosynthetic membrane lipids via the prokaryotic pathway, while DAGs, mainly phosphatidylcholine synthesized in the ER, are used for the synthesis of nonplastid membrane lipids and triacylglycerols (TAGs). In the prokaryotic pathway 18:1 is attached to glycerol in the sn-1 position, and 16:0 in the sn-2, and in the eukaryotic pathway it is vice versa. The products of both pathways also differ in the third substituent at the sn-3 position. In monocots, only the eukaryotic pathway remains (Ohlrogge and Browse 1995; Los 2014).
The synthesis of PUFAs from oleic acid (18:1Δ9) in the ER is catalyzed by microsomal desaturases: the second double bond appears due to the activity of Δ12 (or ω6) oleic acid desaturase (the FAD2 gene) which converts oleic acid into linoleic one (18:1Δ9 → 18:2Δ9,12), and the third double bond can be introduced by the Δ15 (or ω3) linoleic acid desaturase (FAD3) which converts linoleic acid into α-linolenic one (18:2Δ9,12 → 18:3Δ9,12,15). These FAs can also be formed by chloroplast desaturases (controlled by the FAD4 and FAD5 genes) which convert palmitic acid into palmitoleic one (16:0 → 16:1), as well as FAD6 converting oleic acid into linoleic one (18:1 → 18:2), FAD7 and FAD8 converting linoleic acid to α-linolenic one (18:2 → 18:3) (Ohlrogge and Browse 1995; Los 2014).
FAs in plants, functioning as the building blocks of membrane lipids and triacylglycerols, are synthesized almost exclusively in plastids. Lipids of cellular and photosynthetic membranes are represented by DAGs; their sn-1 and sn-2 positions are occupied by saturated or unsaturated C14–C22 FAs, and the sn-3 position by the phosphate group as well as a neutral or charged molecule (Los 2014).
The main lipids of plastid membranes are galactolipids: mono- and digalactosyldiacylglycerols, making up 50 and 25%, respectively, of all lipids in chloroplast membranes. These compounds contain a high proportion of PUFAs, up to 95% of which can be represented by α-linolenic acid located at the sn-1 and sn-2 positions (Los 2014).
In ripening seeds, phosphatidylcholine is the main source of DAGs for the synthesis of TAGs. This synthesis is effectuated via either the acyl-CoA-dependent pathway (Kennedy pathway), catalyzed by diacylglycerol acyl-CoA acyltransferase, or the acyl-CoA-independent pathway, catalyzed by phospholipid:acyl-CoA acyltransferase, and they can duplicate each other. The synthesized TAGs are packed into oleosomes consisting of TAGs surrounded by a monolayer of phospholipids with numerous specific proteins, mainly oleozymes. These proteins prevent oleosomes from sticking together (Los, 2014; Sidorov and Tsydendambaev 2014).
It should be noted that a majority of subsequent FA modifications, such as elongation after the formation of stearic or oleic acid and desaturation, most likely occur after the development of phosphatidylcholine or other glycerolipids of FAs (Ohlrogge and Browse 1995; Dar et al. 2017; Los 2001, 2014). Most of these reactions involve FA elongase (the FAE1 gene, or FA Elongase 1) (Fig. 1), when stearic acid is the first to be formed from arachidic acid, with behenic and lignoceric acids coming next. Further elongation of unsaturated FAs occurs already after the formation of the first double bond: eicosenoic, erucic and nervonic acids are developed in turn from oleic acid (Bao et al. 1998; Haslam and Kunst 2013).
Hereditary variability of fatty acid composition
There is a natural diversity in the composition of FAs among different cultivars, accessions and lines of each oilseed crop (Supplementary).
Peanut (Arachis hypogaea L.)
Peanut, being one of the most important oil crops the world production, contains oleic (39–70% of the total FAs), linoleic (26–42%), linolenic (0.5–1.3%), eicosenoic (1.1–4.0%) acids as well as such saturated FAs as palmitic (8.6–13.4%), stearic (2–6%), arachidic (0.8–2.3%), behenic (0.5–4.0%) and lignoceric (0.5–2.1%), minor amounts of myristic acids (Ermakov et al. 1982; Wang et al. 2015a). The first spontaneous mutant line of peanut with high (80%) oleic acid content (F435) was identified in 1987 (Norden et al. 1987). Molecular analysis of the high-oleate peanut line F435 made it possible to identify mutations in both homeologous ahFAD2: the substitution of “G” for “A” at 448 bp from the start codon (G448A) in the ahFAD2A gene, resulting in the replacement of aspartic acid with asparagine (D150N), as well as the “A” insertion between 441 and 442 bp (441_442insA) in the ahFAD2B gene, which led to a reading frame shift and generated a premature stop codon (Lopez et al. 2000; Jung et al. 2000). Mutant alleles of both genes, being present in the homozygous state concurrently in one genotype, lead to higher oleic acid content in it (Chu et al. 2009).
Later, as a result of mass screening, a new genotype with high oleic acid content (PI342666) was identified (Wang et al. 2015b). The mutation resulted from one substitution of “C” for “G” at 301 bp from the start codon (C301G) in ahFAD2B and led to the substitution of the amino acid. These reports demonstrated that mutations in the coding region of the ahFAD2A and ahFAD2B genes can alter enzymatic activity, increasing the oleic acid content in mutant genotypes (Lopez et al. 2000).
Safflower (Carhtamus tinctorius L.)
The oil of safflower is characterized by a higher content of linoleic acid (68–79%) compared to most other oil crops. Standard safflower oil contains about 6–8% of palmitic acid, 2–3% of stearic acid, 16–20% of oleic acid, and 71–75% of linoleic acid (Velasco and Fernandez-Martinez 2001). Safflower has a specific advantage over rapeseed or soybean in that it does not contain ɑ-linolenic acid in seed oil, a trait that simplifies genetic interventions needed to improve overall oxidative stability (Wood et al. 2018). Safflower is characterized by great variability in its FA composition. Modern safflower cultivars are of two types: either with low oleic acid (13% of oleic and 78% of linoleic acid) or high oleic acid (78% of oleic and 13% of linoleic acid). Such significant discrepancy in the ratio of oleic and linoleic acids is induced by a mutation in the coding region of CtFAD2-1, where a single-nucleotide deletion was found in the ol allele (Liu et al. 2013). This leads to premature termination of translation (Guan et al. 2012). Mutant types of safflower with very high oleic acid content (> 85%) were obtained (Fernandez-Martinez et al. 1993). Safflower accessions from Portugal demonstrated high levels of linoleic acid (87–89%) and very low amounts of oleic acid (3–7%) (Futehally and Knowles 1981). The OLOL genotype determined the synthesis of 72–80% of linoleic acid in oil, while olol was characterized by an increased content of oleic acid (72–80%) (Knowles 1989). Safflower was found to incorporate a large family of FAD2 genes with various manifestations of their expression in different plant tissues. The ol allele was later introduced into safflower breeding programs, and the first high-oleate safflower cultivar ‘UC-1’ was developed in 1965 in the United States. Subsequently, based on this material, a series of Saffola was released, including Saffola 317 (S-317), S-517 and S-518 (Knowles et al. 1965). The ol allele was also used in breeding programs all over the world as genetic material to further enhance oleic acid content in safflower seeds (Weisker 1997).
Brassicaceae Burnett, nom. cons. (1835)
Almost all oilseed of the genus Brassica L., with the exception of Camelina sativa, are characterized by high amounts of erucic acid from 20 to 60%. The highest erucic acid content in oil is reported for crambe (Crambe abyssinica L.) and rocket (Eruca sativa L.) (about 60%). For all species of mustard (Brassica junceae L.), the level of erucic acid in oil is 20–40%. Ethiopian mustard (Brassica carinata Braun), like other mustard species, is characterized by a high content of erucic acid (35–48%), while its oleic acid level is about 10%, and linoleic and linolenic acids are present in almost equal portions (15–17%) (Scharafi et al. 2015). Ethiopian mustard genotypes with reduced erucic acid content are unknown.
Old landraces of rapeseed (B. napus (L.) subsp. oleifera Metzg.) and turnip rape (Brassica campestris (L.) DC. = Brassica rapa subsp. oleifera Metzg.) contain from 12 to 49% of erucic acid (Nizova et al. 1999). However, if rapeseed and turnip rape were found to have natural erucic acid-free genotypes, it is extremely difficult to obtain non-erucic forms of brown mustard. Continuous efforts were undertaken by plant breeders to cross brown mustard with non-erucic rapeseed lines, but in the resulting hybrids, despite all long-term selection procedures, only forms with the erucic acid content from 10 to 17% were obtained (Shpota and Podkolzina 1980). Australian scientists reported the release of two non-erucic lines Zem1 and Zem2 (Kirk and Oram 1981), but according to the VIR Catalogue, Zem1 contains from 2 to 6% of erucic acid (Nizova et al. 1999), which may be the result of cross-pollination among free-flowering mustard accessions.
Camelina oil contains from 1.8 to 4.4% of erucic acid, with approximately 14% of eicosenoic acid. Probably this property of camelina oil makes it indispensable in cosmetology. In addition to the content of these long-chain acids, the oil of this cruciferous crop is balanced in the composition of unsaturated acids: 15–17% of oleic, about 20% of linoleic, and about 30% of linolenic acid (Nizova et al. 1999), and is used for food.
Rapeseed oil contains FAs containing 12–24 carbon atoms. Saturated FAs account for about 5% and are represented by palmitic, stearic, arachidic, behenic, and lignoceric acids. Unsaturated acids are palmitoleic (0.01–0.5%), oleic (20–30%), linoleic (10–20%), linolenic (1.8–3.0%), and erucic. The content of erucic acid in the absence of selection for this trait can vary within wide limits, for example, from 13% in cv. ‘Fedorovsky’ (k-4680) to 51% in cv. ‘Snitynsky’ (k-4688) (Nizova et al. 1999). Widespread use of rapeseed oil for food became possible due to pollination of cv. ‘Liho’ with high erucic acid content, which resulted in identifying genotypes with different levels of erucic acid, including the spontaneous erucic acid-free mutant (Stefansson et al. 1961). The synthesis of erucic acid in rapeseed was found to be controlled by two genes with a series of multiple alleles that determine different ratios of erucic acid in oil: the e (less than 1% in the homozygote), Ea (10%), Eb (15%), Ec (30%) and Ed (3.5%) alleles (Krzymanski and Downey 1969). Studying the nature of these genes showed that they represent FAE genes. The control of erucic acid in the natural rapeseed amphidiploid is determined by the additive interaction between the homologous genes FAE1.1 (the A genome from B. rapa) and FAE1.2 (the C genome from B. oleracea) (Harvey and Downey 1964), which are 99.4% identical in the nucleotide sequence. A decrease in the content of erucic acid down to its almost complete absence occurs when the functional activity of these genes is lost. For the FAE1.2 gene, it happens as a result of a double-nucleotide deletion at position 1422 or a four-nucleotide deletion at position 1366, while the reading frame is shifted and the translation of the 3-ketoacyl-CoA synthase enzyme is prematurely terminated (Fourmann et al. 1998). For the FAE1.1 gene, it is due to a single-nucleotide substitution at position 282, resulting in the substitution of the amino acid serine with phenylalanine in the synthesized protein (Katavic et al. 2002). Specific DNA markers for the FAE1.1 mutant allele have been developed (Michaels and Amasino 1998; Rahman et al. 2008; Grushetskaya et al. 2011). Non-erucic mutants were obtained in Canada for both rapeseed and turnip rape. The absence of erucic acid in oil was a trait so important for Canadian researchers that they considered it possible to merge the two species. The newly developed cultivars of B. napus and B. campestris (and later B. junceae) with an improved FA composition, short growing season, and compact habitus were combined into one crop called ‘Canola’ (Harvey and Downey 1964).
In erucic acid-free oil, the percentages of oleic (from 12–20% to 65–70%) and linoleic (from 10 to 18–20%) FAs are significantly increased (Gavrilova et al. 2020) due to the termination of the hydrocarbon chain lengthening process. Four alleles of the FAD2 gene were identified. Combining the properties of each individual FAD2 allele makes it possible to produce an optimal combination of alleles for obtaining new rapeseed lines with oleic acid content of up to 86% (Nath et al. 2016). It was established that the synthesis of linolenic acid could proceed in two different ways: direct desaturation of oleic acid to linoleic and then to ɑ-linolenic acid, or desaturation of the short-chain acid with 12 carbon atoms followed by subsequent prolongation (C16:3 and then C18:3). The ratio of linoleic acid to linolenic one is controlled by the FAD3 gene (Bocianowski et al. 2012).
Castor bean (Ricinus communis L.)
Castor bean seeds contain a very special castor oil. It comprises ricinoleic acid (C18H34O3, or C18:1 1,12-OH), synthesized based on oleic acid (Fig. 1). Ricinoleic acid is characterized by one unsaturated bond, high molecular weight, and a low melting point (5 °C). The content of ricinoleic acid depends on the genotype and ranges from 78 to 91%. Due to the high content of this acid, castor oil is viscous and does not solidify under negative temperatures (down to − 18… − 20 °C), which makes it a lubricant of unsurpassed quality, especially for engines operating in boreal environments or in the aircraft and space sectors (Moshkin 1980).
Sunflower (Helianthus annuus L.)
Sunflower oil for food contains on average: 1.0% of stearic, 1.2% of palmitic, 8.0% of oleic, 70.0% of linoleic, and 1.0% of linolenic acid (Gavrilova et al. 2020). The first mutants with a modified FA composition of sunflower oil were obtained by K.I. Soldatov (1976) using the mutagen diethyl sulfate. The content of oleic acid in the original cultivar ‘VNIIMK 8931’ was about 30%. In the resulting mutants, the oleic acid content averaged 72%. Based on mutant lines cv. ‘Pervenets’ was developed. Later, the lines containing 88.2% of oleic acid were released (Demurin 2003). The increased oleic acid content is inherited as a dominant trait. From one to five genes were assumed to determine this feature (Berville 2010). Subsequently, one gene was identified, the Ol gene, which was mapped to linkage group 14 (LG14) (Lacome and Berville 2001; Perez-Vich et al. 2002). It was shown that the increased content of oleic acid correlates with the expression of the FAD2-1 gene, and this gene is identical to the Ol gene (Lacome and Berville 2001). The FAD2-1 gene is expressed only in developing seeds (Hongtrakul et al. 1998). Genes FAD2-2 and FAD2-3 control the synthesis of linoleic acid from oleic acid (Hongtrakul et al. 1998; Schuppert et al. 2006). After simultaneous treatment of sunflower with two chemical mutagens NMU and EMS, two lines with low palmitic acid content were obtained (Miller and Vick 1999). X-ray treatment helped to produce a high-palmitic mutant with palmitic acid content of about 25%, while a normal level of this acid in sunflower is about 7% (Perez-Vich 2002), as well as mutants CAS-3 CAS-4 and CAS- 8 with high stearic acid content (about 26%) (Fernandez-Martinez et al. 2004). The recessive allele of the es1 gene, localized in the first linkage group (LG1), is responsible for increased stearic acid content in the CAS3 line (Perez-Vich et al. 2002), and the recessive allele of the es3 gene, belonging to the 8th linkage group (LG8), in the CAS14 line (Perez-Vich et al. 2006). The first linkage group also includes the genes responsible for the FA synthesis: SAD17 locus A (stearate desaturase); SAD6 locus B (stearate desaturase), FatA (thioesterase locus A), and oleate desaturase, identified when sunflower lines HA89 and CAS20 were crossed (Perez-Vich et al. 2004). In the same crossing, the following genes were identified: SAD6 locus A (LG11), SAD6 locus C (LG4), FatA (thioesterase locus B–LG2), FatB (thioesterase locus A–LG7). Using chemical mutagenesis, the CAS-7 mutant with 14% of palmitic acid was produced. (Perez-Vich et al. 2004).
Flax (Linum usitatissimum L.)
Flax seeds contain 25–45% of oil, which includes palmitic (5–7%), stearic (3–4%), oleic (16–20%), linoleic (14–17%) and linolenic (50–60%) acids (Porokhovinova et al. 2019).
The FA biosynthesis in flax is controlled by the genes of ketoacyl-CoA synthase KASIII (Acetyl-CoA + Malonil-CoA → 4:0), KASI (4:0 → 16:0) and KASII (16:0 → 18:0), sad1 and sad2 genes of stearoyl-ACP desaturase (18:0 → 18:1), fad2a and fad2a genes of desaturase 2 (18:1 → 18:2). The genes fad3a and fad3b (also known as ln1 and ln2) encode desaturases that convert linoleic acid into linolenic one, needed for industrial oils. The genes differ in size due to indels from 1 to 29 bp, located in introns. For Fad3a, 6 isoforms are known, 4 of which do not lead to inactivation of the enzyme. The first of them, A, is the most common, while isoforms D and E have lost functionality due to a nonsense mutation. For Fad3b, 7 isoforms are known; only two of them do not yield a full-fledged product: B due to a nonsense mutation in the first exon, and C due to the substitution of histidine with tyrosine in the first His box of the active center of desaturase. All flax accessions with genes lacking a functional product were obtained using EMS mutagenesis (Thambugala et al. 2013).
The first low-linolenic flax (solin) cultivars (Linola™) were developed in the late 1970s in Canada. They were the products of hybridization between two medium-linolenic lines obtained by EMS mutagenesis based on cv. ‘Glenelg’. Solin cultivars contain about 2% of linolenic acid are double-recessive homozygotes for the complementary ln1 and ln2 genes (Green 1990). Later, other lines and cultivars were released, either descending from cv. ‘Linola’ or obtained independently (Dmitriev et al. 2020).
The VIR collection holds several medium-linolenic lines developed from local plant accessions collected in Russia and India. Interestingly, the lower share of linolenic acid in them is compensated by an increased proportion of oleic acid, not linoleic one (Porokhovinova et al. 2019).
Cotton (Gossypium L.)
Cotton oil is used for food and, to a lesser extent, for industrial purposes. It is used mostly for the production of salad oil and frying oil, margarine, mayonnaise, lard substitute and, on a smaller scale, for storage and transportation of meat and fish (Salunkhe and Desai 1986). Cottonseed oil belongs to the linoleic type of oils with a predominance of unsaturated linoleic acid up to about 50%, while 18–30% falls on oleic acid and 20–25% on saturated palmitic acid. The remaining 10% may include up to 1.2% of palmitoleic, 2–7% of stearic and minor amounts of 7–9 other FAs. A distinctive feature of the Malvaceae is the presence of cyclopropane FAs in the oil that have three-carbon-atom rings in the middle of their structures. Cottonseed oil contains from 0.5 to 2% of malvalic (cpe18:1), sterculic (cpe19:1) and dihydrosterculic (cpa19:0) acids. After heating their content is less than 0.4%.
All types of cotton contain the fat-soluble pigment gossypol. that is toxic to humans, pigs and chickens, but harmless to cattle, so the oil intended for food purposes is necessarily refined and deodorized (Dowd 2015).
Several accessions of wild G. barbadense L. with an increased content of oleic acid (38–42%) were identified in the U.S. National Cotton Germplasm Collection (NCGC) (Shockey et al. 2017).
Interspecific hybridization between G. barbadense accessions with a high content of oleic acid and G. hirsutum L. lines resistant to reniform nematodes (Rotylenchulus reniformis Linford and Oliveira), and subsequent selection resulted in producing resistant lines with an increased content of oleic acid (38–42%), which was twice higher than the normal level (Dowd et al. 2020). Four pairs of FAD2 genes (4 from each of the A and D genomes) were identified in allotetraploid cotton plants (Chapman and Pirtle 2001). Expressions of a different form of this enzyme were found in GB713 (Shockey et al. 2017) and GB331 (Sturtevant et al. 2017).
Modification variability and value of desaturases
The synthesis of FAs is affected by environmental conditions. Low temperatures increase the content of PUFAs in the oil. The effect of exposure to temperatures is expressed in different species to varying degrees. In safflower ol1ol1 and OLol1 genotypes the content of linoleic acid slightly decreased under high temperatures, while the content of oleic acid increased. The OLOL and olol genotypes were more stable under temperature changes (Knowles 1989). In flax a wet and cold summer formed seeds with a higher percentage of linolenic and linoleic acids than a hot and dry one (Porokhovinova et al. 2017). The synthesis of linoleic acid in cottonseed, sunflower and camelina oils was found to occur more intensively at lower temperatures (Podolnaya et al. 2019). The CAS14 mutant sunflower line showed very high levels of stearic acid in oil (up to 37%), but only under higher temperatures during seed maturation (Fernandes-Moya et al. 2002).
The FA composition of peanut oil is influenced by several factors: genotype, seasonal variation, growing area and/or temperature conditions, diseases, and insect damage (Sanders 1982). All components of peanut oil quality are highly dependent on the genotype (Bovi 1982; Norden 1987). Seeds that develop at lower temperatures contain more unsaturated FAs in their oil due to an increased activity of oleate desaturase (Slack and Browse 1984).
It was established for cotton that under hotter and drier conditions the content of saturated FAs increased and the content of unsaturated ones decreased (Pettigrew et al. 2011; Podolnaya et al. 2019). On saline soils the level of linoleic acid decreases and that of stearic and oleic acids increases (Ahmad et al. 2007). In cotton, the synthesis of linoleic acid from oleic one occurs under the influence of Δ12 desaturase. The activity of Δ12 desaturase was shown to be affected by temperature. The expression of desaturase-encoding genes was proved to increase at low temperatures (Berestovoy et al. 2020). Desaturases are supposed to play an important role in changing membrane fluidity. Under the impact of various stressors (cold, drought, salinity etc.,) the fluidity of cell membranes decreases. Membrane fluidity parameters are decisive for the functioning of membrane-bound enzyme systems; with a decrease in membrane fluidity, the vital activity of cells is disrupted, which leads to the death of the entire plant (Los 2001). The lower are the content of unsaturated FAs in membrane lipids and the fluidity of membranes, the faster is the activation of desaturase genes. To maintain a certain level of membrane fluidity, the expression of desaturase genes increases and plant defense mechanisms are provided (Los 2001, 2014; Berestovoy 2020). D.A. Los supposed that with the changing environmental temperature and mobility of membrane lipids (their fluidity), their state is perceived by the membrane-bound sensor histidine, kinase Hik33, and the signal is transmitted further to the desaturase gene promoters through the phosphorylation/dephosphorylation cascade with the participation of soluble histidine kinase Hik19 and Rer1 response regulator (Los 2004). Thus, it can be assumed that desaturases contribute to cold and drought resistance, which is important for cultivation of southern crops (peanut, soybean or cotton) at higher northern latitudes.
Genetic engineering for creating oilseeds with new properties
The development of genetic engineering methods has opened new opportunities for changing the FA composition of oil crops. Over the past couple of decades, extensive material has been accumulated for the development of plant forms with a modified composition of FAs (Khan et al. 2017).
Various approaches have been used to achieve this goal, such as gene silencing, overexpression of genes for the synthesis of FAs, changing the pattern of their expression, and genome editing (Table 1). As a result, new genotypes have been obtained. They were characterized by new properties due to the modified balance of FAs or the appearance of fundamentally new compounds for a particular plant species.
Initially, two main approaches were proposed to alter the balance of FAs. In the first case, attempts were made to enhance the expression of genes whose products were supposed to control the production of a particular compound in larger amounts than in the original form.
The second approach, was aimed at silencing the gene expression in order to shift the balance of FAs towards the precursor of the acid whose production was suppressed.
Overexpression
For overexpression, plant genes are placed under strong constitutive or tissue-specific promoters.
Much effort has been focused on transgenic overexpression of FAD2 that catalyzes the conversion of oleic acid into linoleic acid and is the main determinant of the levels of essential PUFAs in seed oils. However, since the discovery of the FAD2 gene, almost none have shown an increase in 18:2 in FAD2 overexpression lines. In contrast, FAD2 co-suppression was reported for Arabidopsis thaliana L. by Cartea et al. (1998), soybean by Kinney (1998), flax, Ethiopian mustard and camelina (Du et al. 2019). There is little evidence at present to indicate that overexpression of FAD2 is a reliable strategy to improve PUFA content in oil crops. A solution to the problem may lie in the use of mutant plant lines with impaired silencing. This approach was implemented using Arabidopsis forms where FAD2 co-suppression was overcomed in the rdr6 mutant, which is impaired in post-transcriptional gene silencing (PTGS). So, Arabidopsis lines with high PUFA content were stable through four generations. Thus, overexpression of desaturase genes does not necessarily lead to an increase of the PUFA level. Implementation of this trait depends on the genetic background, in particular, mutations in genes whose products are involved in PTGS (Du et al. 2019). In addition, the likelihood of success is higher with tissue-specific (Yeom et al. 2020) or inducible expression of the target gene. For example, the use of cold-induced promoters makes it possible to change the balance of FAs in response to cold weather. In the Nicotiana tabacum L. model system, the FAD7 gene was introduced under the control of a cold-inducible promoter (cor15a) from A. thaliana, resulting in a linolenic acid increase by 18.5% after a long cold temperature exposure, while linoleic acid declined to non-detectable levels, and the content of oleic acid decreased by 55% (Khodakovskaya et al. 2006).
Post-transcriptional gene silencing
Various PTGS strategies were exploited to engineer plant genotypes (Kamthan et al. 2015). The traditional hairpin-based RNA interference (RNAi) technique was successfully used to change the FA composition in rapeseed (Peng et al. 2010), brown mustard (Sivaraman et al. 2004), peanut (Yin et al. 2007), and safflower (Wood et al. 2018).
The artificial microRNA (amiRNA) technique of gene silencing is becoming a powerful tool of genetic engineering. The amiRNA technology could be employed to down-regulate gene expression without affecting the expression of other members of gene families, which reduces the risk of cross-silencing (Tang et al. 2007). This approach started to be intensively used in genetic engineering of oilseed crops. In the Arabidopsis model object, three important genes, i.e., FAD2, FAE1 and FATB involved in seed oil metabolism, were efficiently silenced. The efficiency of FAD2 and FATB silencing was matched with the well-characterized mutant alleles (Belide et al. 2012). There is a known safflower accession with superhigh oleic acid content: 93% of oleic acid, 4% of saturated acids, and only 1.5% of PUFAs (linoleic and linolenic). Such content of oleic acid resulted from a specific RNA interference of FAD2.2 and FATB genes (Wood et. al. 2018).
Hairpin RNA-mediated gene silencing was used in cotton to suppress the expression of ghSAD-1 encoding stearoyl-ACP Δ9-desaturase and ghFAD2-1 encoding oleoyl-phosphatidylcholine Δ6-desaturase. As a result of a decrease in the expression of the ghSAD-1 gene, the amount of stearic acid in oil significantly increased (from 2–3 to 40%). Blocking the ghFAD2-1 gene led to a significant increase in the oleic acid content—up to 77%, compared to 15% in the original cotton cultivar (Liu et al. 2002). In all variants, the content of palmitic acid was reduced.
Thus, methods based on RNA interference are very popular and quite predictable in terms of their results.
Genome editing
Different methods have historically been used to edit genomes. Initially, editing systems were based on the DNA–protein interactions to introduce targeted mutations. This ideology, implemented in the TALLEN method, was applied to editing the FAD2 and FAD3 genes in soybean and peanuts (Demorest et al. 2016; Wen et al. 2018), leading to an increase in the content of oleic acid in the seeds of transformed plants up to 70% (Yin et al. 2007).
The possibilities of genome editing have greatly increased with the use of the CRISPR/Cas technology, since nucleic acid interactions based on the principle of complementarity have been used to find the editing site, greatly facilitating the procedure of creating vectors for genome editing. As a result, novel applications of this system are being developed rapidly. Table 1 contains CRISPR/Cas9 application examples to target the FAD2 gene in Camelina seeds, where oleic acid content was raised from 16% to over 50%. These increases were associated with significant decreases in the less desirable PUFAs, linoleic acid (from ~ 16 to < 4%) and linolenic acid (from ~ 35 to < 10%) (Jiang et al. 2017).
Metabolic engineering for the synthesis of new compounds
Methods of metabolic engineering make it possible to introduce new compounds into the composition of a vegetable oil, thus enriching it with previously uncharacteristic valuable nutritional properties. In some cases, it is sufficient to introduce 1 or 2 heterologous genes into plants to obtain a new FA in the composition of its oil. Some plants, such as primula, are known to be able to produce stearidonic acid in their seeds (Sayanova et al. 1997). However, these plants are not part of the human diet and agronomically not amenable to cultivation. Oilseeds can therefore be engineered to produce this nutritionally important FA by transferring the 6-desaturase gene to produce SDA, which has been successfully done for soybeans (Eckert et al. 2006).
Safflower, canola and brown mustard oils were enriched with γ-linolenic acid (Nykiforuk et al. 2012; Liu et al. 2001; Hong et al. 2002), while cottonseed oil was enriched with vernolic acid (Zhou et al. 2006). An important alternative source of fish oil substitutes are metabolically engineered plants. To produce eicosapentaenoic acid (EPA, 20:5n-3), five heterologous genes driven by the constitutive 35S promoter, namely ∆9-elongase (Isochrysis galbana Parke), ∆8-desaturase (Euglena gracilis Klebs) ∆5-desaturase (Mortierella alpine Peyronel), ∆15-desaturase (A. thaliana) and ∆17-desaturase (Phytophthora infestans (Mont.)de Bary) (Fig. 2), were successfully transferred into peanut through Agrobacterium-mediated transformation (Wang et al. 2019). Many desired oil modifications require an even larger number of transgenes to produce target phenotypes. For example, the production of the high-value nutritional FA docosahexaenoic acid (DHA) was achieved in B. juncea by seed-specific co-expression of nine transgenes (Wu et al. 2005), and the ISAAA database contains a canola line containing a cassette of 12 genes, the products of which are involved in the synthesis of DHA. These achievements are encouraging, suggesting that the composition of FAs in other plants can be altered.

A flowchart of fatty acid synthesis in plants exposed to metabolic engineering
Conclusion
Changes in the FAs composition of oil crop seeds can be achieved in several ways. This goal can be reached by studying the natural diversity of a crop and disclosing the potential for variability through selection, self-pollination or interspecific hybridization. The use of chemical mutagenesis or X-ray treatment makes it possible to obtain forms with a high content of oleic acid in sunflower, rapeseed and cotton, with a low content of linolenic acid in flax, and with an increased content of palmitic or stearic acid in sunflower. The third way is to study in detail the genetic control of FA biosynthesis to provide the means for transgenesis and editing the genome and obtaining the desired plant form by turning on or inactivating certain genes, or engineering new metabolic pathways. This is important because the genes that control desaturase are involved not only in the synthesis of FAs s in seeds, but also in ensuring the vital activity of cell membranes by changing their fluidity and, consequently, the plant organism as a whole, especially under unfavorable conditions, such as low or high temperatures, drought, soil salinity, etc.
Notes
2* Hereinafter, the gene names are given in accordance with the authors’ notation.
References
Ahmad S, Anwar F, Hussain A et al (2007) Does soil salinity affect yield and composition of cottonseed oil? J Am Oil Chem Soc 84:845–851. https://doi.org/10.1007/s11746-007-1115-8
Bao X, Pollard M, Ohlrogge J (1998) The biosynthesis of erucic acid in developing embryos of Brassica rapa. Plant Physiol 118:183–190
Belide S, Petrie J, Shrestha P, Singh S (2012) Modification of seed oil composition in Arabidopsis by artificial microRNA-mediated gene silencing. Front Plant Sci 3:168. https://doi.org/10.3389/fpls.2012.00168
Berestovoy M, Pavlenko O, Goldenkova-Pavlova I (2020) Plant fatty acid desaturases: role in the life of plants and biotechnological potential. Biol Bull Rev 10(2):127–139. https://doi.org/10.1134/s2079086420020024
Berville A (2010) Oil composition variations. In: Hu J, Seiler G, Kole C (eds) Genetics, Genomics and Breeding of Sunflower. Science Publisher, Boca Raton, pp 253–277
Bocianowski J, Mikołajczyk K, Bartkowiak-Broda I (2012) Determination of fatty acid composition in seed oil of rapeseed (Brassica napus L.) by mutated alleles of the FAD3 desaturase genes. J Appl Genet 53(1):27–30. https://doi.org/10.1007/s13353-011-0062-0
Bonaventure G, Salas J, Pollard M, Ohlrogge J (2003) Disruption of the FATB gene in Arabidopsis demonstrates an essential role of saturated fatty acids in plant growth. Plant Cell 15:1020–1033. https://doi.org/10.2307/3871906
Cartea M, Migdal M, Galle A et al (1998) Comparison of sense and antisense methodologies for modifying the fatty acid composition of Arabidopsis thaliana oilseed. Plant Sci 136:181–194
Chapman K, Dand R, Pirtle C (2001) Molecular cloning and functional expression of the gene for a cotton Delta-12 fatty acid desaturase (FAD2). Biochim Biophys Acta 1522:122–129. https://doi.org/10.1016/s0167-4781(01)00312-8
Chu Y, Holbrook C, Ozias-Akins P (2009) Two alleles of ahFAD2B control the high oleic acid trait in cultivated peanut. Crop Sci. 49:2029–2036. https://doi.org/10.2135/cropsci2009.01.0021
Bovi MLA (1982) Genotypic and environmental effects on fatty acid composition, iodine value and oil content of peanut (Arachis hypogaea L.). PhD Dissertation, University of Florida
Dar A, Choudhury A, Kancharla P, Arumugam N (2017) The FAD2 Gene in plants: occurrence, regulation, and role. Front Plant Sci 8:1–16. https://doi.org/10.3389/fpls.2017.01789
Demorest ZL, Coffman A, Baltes NJ et al (2016) Direct stacking of sequence-specific nuclease-induced mutations to produce high oleic and low linolenic soybean oil. BMC Plant Biol 16:225. https://doi.org/10.1186/s12870-016-0906-1
Demurin Y (2003) Up-to-date results on biochemical genetics of sunflower in VNIIMK. Helia 26(38):137–142. https://doi.org/10.2298/HEL0338137D
Dmitriev A, Kezimana P, Rozhmina T et al (2020) Genetic diversity of SAD and FAD genes responsible for the fatty acid composition in flax cultivars and lines. BMC Plant Biol 20:301. https://doi.org/10.1186/s12870-020-02499-w
Dowd M (2015) Seed. In: Fang D, Percy R (ed) Cotton. Agronomy Monographs, 57:745–782. https://doi.org/10.2134/agronmonogr57.2013.0032
Dowd M, McCarty J Jr, McCarty J, Jenkins J (2020) Registration of four upland cotton germplasm lines with elevated levels of seed oil oleic acid. J Plant Regist 14(1):64–71. https://doi.org/10.1002/plr2.20017
Du C, Chen Y, Wang K et al (2019) Strong co-suppression impedes an increase in polyunsaturated fatty acids in seeds overexpressing FAD2. J Exp Bot 70(3):985–994. https://doi.org/10.1093/jxb/ery378
Eckert H, La Vallee B, Schweiger B et al (2006) Co-expression of the borage Δ6-desaturase and the Arabidopsis Δ15-desaturase results in high accumulation of stearidonic acid in the seeds of transgenic soybean. Planta 224(5):1050–1057. https://doi.org/10.1007/s00425-006-0291-3
Ermakov A, Davidyan G, Yarosh N et al (1982) Characterization of oil quality for fatty acid content. Catalogue of the VIR Global Collection. Oil Crops. VIR, Leningrad (in Russian)
Fernandes-Moya V, Martines-Force E, Garces R (2002) Temperature effect on a high stearic acid sunflower mutant. Phytochem 59(1):33–37. https://doi.org/10.1016/s0031-9422(01)00406-x
Fernandez-Martinez J, Rio M, Haro A (1993) Survey of safflower (Carthamus tinctorius L.) germplasm for variants in fatty acid composition and other seed characters. Euphytica 69:115–122. https://doi.org/10.1007/BF00021734
Fernandez-Martinez J, Velasco L, Perez-Vich B (2004) Progress in the genetic modification of sunflower oil quality. In: Proceedings of the16th international sunflower conference. Fargo, North Dakota, USA, 1: 1–14
Fourmann M, Barret P, Renard M et al (1998) The two genes homologous to Arabidopsis FAE1 co-segregate with the two loci governing erucic acid content in Brassica napus. Theor Appl Genet 96:852–858. https://doi.org/10.1007/s001220050812
Futehally S, Knowles P (1981) Inheritance of very high levels of linoleic acid in an introduction of safflower (Carthamus tinctorius L.) from Portugal. In: Proceedings of the first international safflower conference, Davis, California, pp 56–61
Gavrilova V, Shelenga T, Porokhovinova E et al (2020) The diversity of fatty acids composition in traditional and rare oil crops cultivated in Russia. Biol Commun 65(1):68–81. https://doi.org/10.21638/spbu03.2020.106
Green A (1990) Low linolenic acid oil producing flax cultivars. European patent application: AP0431833A11990
Grushetskaya Z, Mozgova G, Bakanovskaya A et al (2011) Development of genome-specific markers to identify FAE1 genes controlling erucic acid synthesis in Brassica napus. In: Scientific priorities for innovative development of plant production sector: results and prospects. Proceedings of the international scientific and practical conference. Zhodino, Belarus, 271–273 (in Russian)
Guan L, Wang Y, Shen H et al (2012) Molecular cloning and expression analysis of genes encoding two microsomal oleate desaturases (FAD2) from safflower (Carthamus tinctorius L.). Plant Mol Biol Rep 30:139–148. https://doi.org/10.1007/s11105-011-0322-5
Harvey B, Downey R (1964) The inheritance of erucic acid content in rapeseed (Brassica napus). Can J Plant Sci 44(11):104–111. https://doi.org/10.4141/cjps64-019
Haslam T, Kunst L (2013) Extending the story of very-long-chain fatty acid elongation. Plant Sci 210:93–107. https://doi.org/10.1016/j.plantsci.2013.05.008
Hong H, Datla N, Reed D et al (2002) High-level production of γ-linolenic acid in Brassica juncea using a Δ6 desaturase from Pythium irregulare. Plant Physiol 129:354–362
Hongtrakul V, Slabaugh M, Knapp S (1998) A seed specific Δ12-oleate desaturase gene is duplicated, and weakly expressed in high oleic acid sunflower lines. Crop Sci 38:1245–1249. https://doi.org/10.2135/cropsci1998.0011183X003800050022x
https://efko-ingredients.ru/press-centr/relizy/15532, Nov 23, 2020
https://efko-ingredients.ru/produkciya/milk_fat/12606/, Nov 23, 2020
https://isaaa.org, Nov 10, 2020
Hu Z, Wu Q, Dalal J et al (2017) Accumulation of medium-chain, saturated fatty acyl moieties in seed oils of transgenic Camelina sativa. PLoS ONE 12(2):e0172296. https://doi.org/10.1371/journal.pone.0172296
Jadhav A, Katavic V, Marillia EF, Michael Giblin E, Barton DL, Kumar A, Sonntag C, Babic V, Keller WA, Taylor DC (2005) Increased levels of erucic acid in Brassica carinata by co-suppression and antisense repression of the endogenous FAD2 gene. Metab Eng 7(3):215–220. https://doi.org/10.1016/j.ymben.2005.02.003
Jiang W, Henry I, Lynagh P et al (2017) Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol 15:648–657. https://doi.org/10.1111/pbi.12663
Jung S, Powell G, Moore K, Abbott A (2000) The high oleate trait in the cultivated peanut (Arachis hypogaea L.). II. Molecular basis and genetics of the trait. Mol Gen Genet 263:806–811. https://doi.org/10.1007/s004380000243
Kamthan A, Chaudhuri A, Kamthan M, Datta A (2015) Small RNAs in plants: recent development and application for crop improvement. Front Plant Sci 6:208. https://doi.org/10.3389/fpls.2015.00208
Kanrar S, Venkateswari J, Dureja P, Kirti PB, Chopra VL (2006) Modification of erucic acid content in Indian mustard (Brassica juncea) by up-regulation and down-regulation of the Brassica juncea fatty acid elongation1 (BjFAE1) gene. Plant Cell Rep 25:148–155
Katavic V, Mietkievska E, Barton D et al (2002) Restoring enzyme activity in nonfunctional low erucic acid Brassica napus fatty acid elongase 1 by a single amino acid substitution. Eur J Biochem 269:5625–5631. https://doi.org/10.1046/j.1432-1033.2002.03270.x
Khan A, Chun-Mei H, Khan N et al (2017) Bioengineered plants can be a useful source of omega-3 fatty acids. BioMed Res Int 2017:7348919. https://doi.org/10.1155/2017/7348919
Khodakovskaya M, McAvoy R, Peters J et al (2006) Enhanced cold tolerance in transgenic tobacco expressing a chloroplast ω-3 fatty acid desaturase gene under the control of a cold-inducible promoter. Planta 223:1090–1100. https://doi.org/10.1007/s00425-005-0161-4
Kinney A (1998) Plants as industrial chemical factories – new oils from genetically engineered soybeans. Lipid–Fett 100(4–5):173–176. https://doi.org/10.1002/(sici)1521-4133(19985)100:4/5%3c173:aid-lipi173%3e3.0.co;2-d
Kirk J, Oram R (1981) Isolation of erucic acid-free lines of Brassica juncea: Indian mustard now a potential oilseed crop in Australia. J Aust Inst Agric Sci 47:51–52
Knowles P, Hill A, Ruckman F (1965) High oleic acid content in new safflower. Calif Agric 19:12–15
Knowles P (1989) Safflower. In: Robbelen G, Downey R, Ashri A (eds) Oil crops of the world. McGraw-Hill, New York, pp 363–374
Krzymanski I, Doney R (1969) Inheritance of fatty acid composition in winter forms of rape-seed, Brassica napus. Can J Plant Sci 49:313–319. https://doi.org/10.4141/cjps69-053
Lacome S, Berville A (2001) A dominant mutation for high oleic acid content of sunflower (Helianthus annuus L.) oil is genetically linked to a single oleate-desaturase RFLP locus. Mol Breed 8:129–137
Liu J, DeMichele S, Bergana M et al (2001) Characterization of oil exhibiting high γ-linolenic acid from a genetically transformed canola strain. J Amer Oil Chem Soc 78:489–493. https://doi.org/10.1007/s11746-001-0291-2
Liu Q, Cao S, Zhou X et al (2013) Nonsense-mediated mRNA degradation of CtFAD2-1 and development of a perfect molecular marker for olol mutation in high oleic safflower (Carthamus tinctorius L.). Theor Appl Genet 126(9):2219. https://doi.org/10.1007/s00122-013-2129-2
Liu Q, Singh S, Green A (2002) High-stearic and high-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing. Plant Physiol 129:1732–1743. https://doi.org/10.1104/pp.001933
Lopez Y, Nadaf H, Smith O et al (2000) Isolation and characterization of the Δ12-fatty acid desaturase in peanut (Arachis hypogaea L.) and search for polymorphisms for the high oleate trait in Spanish market-type lines. Theor Appl Genet 101:1131–1138. https://doi.org/10.1007/s001220051589
Los D (2014) Fatty acid desaturases. Nauchny Mir, Moscow (in Russian)
Los D, Murata N (2004) Membrane fluidity and its roles in the perception of environmental signals. Biochim Biophys Acta 1666:142–157. https://doi.org/10.1016/j.bbamem.2004.08.002
Los D (2001) Structure, expression regulation and functioning of fatty acid desaturases. Biolo Chem Rev 41:163–198 (in Russian)
Michaels S, Amasino R (1998) A robust method for detecting single-nucleotide changes as polymorphic markers by PCR. Plant J 14:381–385. https://doi.org/10.1046/j.1365-313x.1998.00123.x
Miller JF, Vick DA (1999) Inheritance of Reduced Stearic and Palmitic Acid Content in Sunflower Seed Oil. Crop Sci 39(2):364–367. https://doi.org/10.2135/cropsci1999.0011183X0039000200010x
Moshkin V (ed) (1980) Castor bean. Kolos, Moscow (in Russian)
Nath K, Kim H, Khatun K et al (2016) Modification of fatty acid profiles of rapeseed (Brassica napus L.) oil for using as food, industrial feed-stock and biodiesel. Plant Breed Biotechnol 4(2):123–134. https://doi.org/10.9787/PBB.2016.4.2.123
Nizova G, Dubovskaya A, Konkova N, et al (1999) Cruciferous crops: rapeseed, turnip rape, brown mustard, and camelina (characterization of accessions for oil, fatty acid and protein content). Catalogue of the VIR Global Collection, Issue 700. VIR, St. Petersburg (in Russian)
Norden A, Gorbet D, Knauft D, Young C (1987) Variability in oil quality among peanut genotypes in the Florida breeding program. Peanut Sci 14:7–11. https://doi.org/10.3146/i0095-3679-14-1-3
Nykiforuk C, Shewmaker C, Harry I et al (2012) High level accumulation of gamma linolenic acid (C18:3Δ6.9,12 cis) in transgenic safflower (Carthamus tinctorius) seeds. Transgenic Res 21:367–381. https://doi.org/10.1007/s11248-011-9543-5
Ohlrogge J, Browse G (1995) Lipid biosynthesis. Plant Cell 7:957–970. https://doi.org/10.1105/tpc.7.7.957
Okuzaki A, Ogawa T, Koizuka C, Kaneko K, Inaba M, Imamura J, Koizuka N (2018) CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol Biochem 131:63–69. https://doi.org/10.1016/j.plaphy.2018.04.025
Peng Q, Hu Y, Wei R et al (2010) Simultaneous silencing of FAD2 and FAE1 genes affects both oleic acid and erucic acid contents in Brassica napus seeds. Plant Cell Rep 29:317–325. https://doi.org/10.1007/s00299-010-0823-y
Perez-Vich B, Fernandez J, Garces R, Fernandez-Martinez J (2002) Inheritance of high palmitic acid content in the sunflower mutant CAS-12 and its relationship with high oleic content. Plant Breed 121:49–56. https://doi.org/10.1046/j.1439-0523.2002.00655.x
Perez-Vich B, Knapp S, Leon A et al (2004) Mapping minor QTL for increased stearic acid content in the sunflower seed oil. Mol Breed 13:313–322. https://doi.org/10.1023/b:molb.0000034081.40930.60
Perez-Vich B, Velasco L, Muñoz-Rus J, Fernandez-Martinez J (2006) Inheritance of high stearic acid content in the sunflower mutant CAS-14. Crop Sci 46(1):49–56. https://doi.org/10.2135/cropsci2004.0723
Pettigrew W, Dowd M (2011) Varying planting dates or irrigation regimes alters cottonseed composition. Crop Sci 51(5):2155–2164. https://doi.org/10.2135/cropsci2011.02.0085
Podolnaya L, Asfandiarova M, Tuz R et al (2019) Variability of fatty acids composition of cotton oil of VIR collection accessions. Oil crops 4(180):29–35. https://doi.org/10.25230/2412-608X-2019-4-180-29-35 (in Russian)
Porokhovinova E, Shelenga T, Kosykh L et al (2017) Biochemical diversity of fatty acid composition in flax from VIR’s genetic collection and effect of environment on its development. Russ J Genet 7:626–639. https://doi.org/10.1134/S2079059717060107
Porokhovinova E, Shelenga T, Matveeva T et al (2019) Polymorphism of genes controlling low level of linolenic acid in lines from VIR flax genetic collection. Ecol Genet 17(2):5–19. https://doi.org/10.17816/ecogen1725-19
Rahman M, Sun Zh, McVetty P, Li G (2008) High throughput genome-specific and gene-specific molecular markers for erucic acid genes in Brassica napus (L.) for marker-assisted selection in plant breeding. Theor Appl Genet 117(6):895–904. https://doi.org/10.1007/s00122-008-0829-9
Salunkhe D, Desai B (1986) Postharvest biotechnology of oilseed. CRC Press, Boca Raton
Sanders T (1982) Peanut triacylglycerols: effect of season and production location. J Am Oil Chem Soc 59:346–351
Sayanova O, Smith M, Lapinskas P et al (1997) Expression of a B. officinalis desaturase cDNA containing an N-terminal cytochrome b5 domain results in the accumulation of high levels of D6-desaturated fatty acids in transgenic tobacco. PNAS 94(8):4211–4216. https://doi.org/10.1073/pnas.94.8.4211
Scharafi Y, Majidi M, Goli S et al (2015) Oil content and fatty acids composition in Brassica species. Inter J Food Prop 18(10):2145–2154. https://doi.org/10.1080/10942912.2014.968284
Schuppert G, Tang S, Slabaugh M, Knapp S (2006) The sunflower high-oleic mutant Ol carries variable tandem repeats of FAD2-1, a seed-specific oleoyl-phosphatidyl choline desaturase. Mol Breed 17(3):241–256. https://doi.org/10.1007/s11032-005-5680-y
Seiler G, Jan C (2010) Chapter 1: Basic information. In: Hu J, Seiler G, Kole C (eds) Genetics, genomics and breeding of sunflower. Science Publishers, Enfield, pp 1–50
Shockey J, Dowd M, Mack B et al (2017) Naturally occurring high oleic acid cottonseed oil: identification and functional analysis of a mutant allele of Gossypium barbadense fatty acid desaturate-2. Planta 245:611–622. https://doi.org/10.1007/s00425-016-2633-0
Shpota V, Podkolzina V (1980) Effectiveness of interspecific hybridization in cruciferous crop breeding for oil quality. Oil Crop Sci Tech Bull VNIIMK 4:18–22 (in Russian)
Sidorov R, Tsydendambaev V (2014) Biosynthesis of fatty oils in higher plants. Russ J Plant Physiol 61(1):3–22 (in Russian)
Sinha S, Jha JK, Maiti MK et al (2007) Metabolic engineering of fatty acid biosynthesis in Indian mustard (Brassica juncea) improves nutritional quality of seed oil. Plant Biotechnol Rep 1:185–197. https://doi.org/10.1007/s11816-007-0032-5
Sivaraman I, Arumugam N, Sodhi Y et al (2004) Development of high oleic and low linoleic acid transgenics in a zero erucic acid Brassica juncea L. (Indian mustard) line by antisense suppression of the fad2 gene. Mol Breed 13(4):365–375. https://doi.org/10.1023/B:MOLB.0000034092.47934.d6
Slack S, Browse J (1984) Synthesis of storage lipids in developing seeds. In: Murray DR (ed) Seed Physiology, vol 1. Academic Press Inc, Orlando Florida, pp 209–244
Soldatov K (1976) Chemical mutagenesis in sunflower breeding. In: Proceeding of the 7th international sunflower conference. International sunflower association, Vlaardingen, Netherlands, pp 352–357
Stefansson B, Hougen F, Downey R (1961) Note on the isolation of rape plants with seed oil free from erucic acid. Can J Plant Sci 41:218–225. https://doi.org/10.4141/cjps61-028
Sturtevant D, Horn P, Kennedy C et al (2017) Lipid metabolites in seeds of diverse Gossypium accessions: molecular identification of a high oleic mutant allele. Planta 245:595–610. https://doi.org/10.1007/s00425-016-2630-3
Tang G, Galili G, Zhuang X (2007) RNAi, microRNA: breakthrough technologies for the improvement of plant nutritional value and metabolic engineering. Metabolomics 3:357–369. https://doi.org/10.1007/s11306-007-0073-3
Thambugala D, Duguid S, Loewen E et al (2013) Genetic variation of six desaturase genes in flax and their impact on fatty acid composition. Theor Appl Genet. 126(10):2627–41. https://doi.org/10.1007/s00122-013-2161-2
Vanhercke T, El Tahchy A, Liu Q et al (2013) Metabolic engineering of biomass for high energy density: oilseed-like triacylglycerol yields from plant leaves. Plant Biotechnol J 12(2):231–239. https://doi.org/10.1111/pbi.12131
Velasco L, Fernandez-Martinez J (2001) Breeding for oil quality in safflower. In: Bergman, JW, Mündel HH (ed) Proceedings of the 5th international safflower conference. Williston, ND, Sidney, MT, USA, pp 133–137
Wang C, Qing X, Yu M et al (2019) Production of eicosapentaenoic acid (EPA, 20:5n–3) in transgenic peanut (Arachis hypogaea L.) through the alternative Δ8-desaturase pathway. Mol Biol Rep 46(1):333–342. https://doi.org/10.1007/s11033-018-4476-1
Wang M, Khera P, Pandey M et al (2015) Genetic mapping of QTLs controlling fatty acids provided insights into the genetic control of fatty acid synthesis pathway in peanut (Arachis hypogaea L.). PLoS One 10(4):e0119454. https://doi.org/10.1371/journal.pone.0119454
Wang M, Tonnis B, Charles Y et al (2015) Newly identified natural high-oleate mutant from Arachis hypogaea L. subsp. hypogaea. Mol Breed 35(9):186. https://doi.org/10.1007/s11032-015-0377-3
Weisker A (1997) Safflower products with very high levels of unsaturated fatty acids. Patent (5912416), California Oils Corporation, USA
Wen S, Liu H, Li X et al (2018) TALEN-mediated targeted mutagenesis of fatty acid desaturase 2 (FAD2) in peanut (Arachis hypogaea L.) promotes the accumulation of oleic acid. Plant Mol Biol 97(1–2):177–185. https://doi.org/10.1007/s11103-018-0731-z
Wood C, Okada S, Taylor M et al (2018) Seed-specific RNAi in safflower generates a superhigh oleic oil with extended oxidative stability. Plant Biotechnol J 16(10):1788–1796. https://doi.org/10.1111/pbi.12915
Wu G, Truksa M, Datla N et al (2005) Stepwise engineering to produce high yields of very long-chain polyunsaturated fatty acids in plants. Nat Biotechnol 23:1013–1017. https://doi.org/10.1038/nbt1107
Yeom W, Kim H, Lee K et al (2020) Increased production of α-linolenic acid in soybean seeds by overexpression of Lesquerella FAD3-1. Front Plant Sci 10:1812. https://doi.org/10.3389/fpls.2019.01812
Yin D, Deng S, Zhan K, Cui D (2007) High-oleic peanut oils produced by HpRNA-mediated gene silencing of oleate desaturase. Plant Mol Biol Rep 25:154–163. https://doi.org/10.1007/s11105-007-0017-0
Zhou X, Singh S, Liu Q, Green A (2006) Combined transgenic expression of Δ12-desaturase and Δ12-epoxygenase in high linoleic acid seeds leads to increased accumulation of vernolic acid. Funct Plant Biol 33:585–592
Funding
This work was supported by the Russian Science Foundation [Grant Number 21-14-00050].
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Porokhovinova, E.A., Matveeva, T.V., Khafizova, G.V. et al. Fatty acid composition of oil crops: genetics and genetic engineering. Genet Resour Crop Evol 69, 2029–2045 (2022). https://doi.org/10.1007/s10722-022-01391-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-022-01391-w