
Abstract
Myrmecochorous plants produce seeds with lipid-rich appendages (elaiosomes) which act as a reward for seed-dispersing ants. Seed dispersal is important for exotic species, which often need to establish new mutualistic interactions in order to colonize new non-native habitats. However, little is known about the importance of elaiosomes for seed removal in many of their non-native ranges. We studied ant–seed interactions of elaiosome-bearing and elaiosome-removed seeds of the Australian trees Acacia dealbata and Acacia longifolia in order to assess the relative importance of elaiosomes for seed removal between their native (Australia) and non-native (Portugal) ranges. In Portugal, we also studied the co-occurring native plant species with myrmecochorous seeds, Pterospartum tridentatum and Ulex europaeus, across three contiguous levels of acacia invasion: control (i.e. no acacia), low, and high acacia tree density. Acacia seeds were successfully removed by ants in their non-native region by a diversified assemblage of ant species, even in sites where native plants interacted with only one specialized ant species. In the invaded range, diminishing relative importance of elaiosomes was associated with changes in the ant community due to acacia invasion, and for A. dealbata, ant species richness decreased with increasing acacia tree density. Although seed removal was high for both acacia species, the importance of elaiosomes was proportionally lower for A. dealbata in the non-native region. Native plant species experienced significant reductions in seed removal in areas highly invaded by acacia, identifying another mechanism of displacement of native plants by acacias.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Biotic interactions are essential components of invasive success, either by releasing invasive species from costly interactions in the native range, or by establishing novel beneficial interactions in their non-native ranges (Traveset and Richardson 2014). Mutualisms are essential for the success of numerous invasive exotic plants (Richardson et al. 2000; Traveset and Richardson 2006), and invasive plants can establish many forms of mutualisms that can increase nutrient acquisition, pollination, or seed dispersal, which are crucial for their success in colonizing new areas (Davidson and Morton 1984; Auld 1986; Willson and Traveset 2000; Richardson et al. 2000; Whitney 2002; Jensen and Six 2006). In fact, the introduction of mutualists from the invader’s native range has been reported to trigger invasion success in the non-native ranges of some introduced species (Richardson et al. 2000). Regardless, invasive plants can establish novel mutualisms by taking advantage of native mutualistic networks already present in the invaded regions (Simberloff and Von Holle 1999; Richardson et al. 2000). Exotic plants usually integrate easily into mutualistic networks because these are often be dominated by generalists (Traveset and Richardson 2011), and invasives tend to preferentially interact with generalists both in their native and non-native ranges (Aizen et al. 2008; Bartomeus et al. 2008; Ferrero et al. 2013; Traveset and Richardson 2014; Montesinos et al. 2016). As a result, native generalist mutualists can become more common after invasions, which in turn can have a negative effect on native plants with more specialized interactions (Traveset and Richardson 2006).
Myrmecochory, seed dispersal by ants, is an important type of mutualistic interaction found in more than 11,000 species, or about 4.5% of all plant species (Lengyel et al. 2010). Seeds of numerous plant species possess elaiosomes: protein and lipid rich seed appendages that constitute an important reward for ants in exchange for seed dispersal services (Gorb and Gorb 1999; Willson and Traveset 2000; Giladi 2006; Lengyel et al. 2009; Fokuhl et al. 2012). Up to 90% of myrmecochorous plant species are native to the Southern hemisphere, while only 10% are native to the Northern hemisphere (Gómez and Espadaler 1998; Lorenzo et al. 2010a; Lengyel et al. 2010). The presence of myrmecochorous mutualisms in the northern hemisphere is known to facilitate ant seed dispersal for myrmecochorous plant species introduced from the Southern hemisphere (Pemberton and Irving 1990; Richardson et al. 2000; Jensen and Six 2006), and invasive plants have been found to establish myrmecochorous mutualisms in their non-native ranges with both native and non-native ants (Smith 1989; Bossard 1991; Jensen and Six 2006; Alba-Lynn and Henk 2010). Furthermore, invasive ants sometimes interact preferentially with invasive plants, synergistically strengthening their success in their non-native regions (Prior et al. 2015).
Plant invasions regularly lead to significant changes in the composition and function of the native disperser community (French and Major 2001; Traveset and Richardson 2006), involving cascading effects along the food web which can result in reduced seed dispersal and recruitment of native plant species (Sallabanks 1993; Williams and Karl 2002). In addition to changes in mutualistic interactions, invasive species frequently develop locally adapted traits as a response to the different environmental conditions found in their non-native ranges (Hierro et al. 2005). Plant traits can differ among different areas within a single non-native region, showing, in some cases, differences between the expanding invasion edges and mature invaded areas (Phillips et al. 2006, 2007; Lankau et al. 2009; Montesinos et al. 2012). Recent studies showed that acacias from non-native populations were investing more resources into seed mass than acacias from native populations, but proportionally less resources in elaiosomes than their conspecifics in the native range (Correia et al. 2016). Our study aims to detect potential implications for seed removal of these changes in resource allocation.
Acacia dealbata Link and A. longifolia (Andr.) Willd. are among the 22 species of Australian acacia listed as invaders in different parts of the world (Richardson et al. 2011; Rejmánek and Richardson 2013). In Europe, Portugal is one of the most affected countries by the invasion by Australian acacias with large natural and seminatural areas covered by these species (De Almeida and Freitas 2006; Lorenzo et al. 2010a; Marchante et al. 2010). Factors that are likely to influence success include the establishment of below-ground mutualisms in invaded areas (Rodríguez-Echeverría et al. 2009; Rodríguez-Echeverría 2010); larger seed production than native species, which are able to accumulate in persistent soil seed banks (Marchante et al. 2010; Correia et al. 2014); allelopathy (Lorenzo et al. 2010b, c; Aguilera et al. 2015); changes in soil properties (Rodríguez-Echeverría et al. 2013); enemy release from pre-dispersal seed predators (Correia et al. 2016); and myrmecochory, which is known to be important for acacia invasion success in South Africa (Holmes 1990; French and Major 2001), and has been reported to be present in Southern Europe (Montesinos et al. 2012). Myrmecochory is important because seeds moved away from the parent plant are able to effectively colonize new areas and to avoid seed predation (Willson and Traveset 2000; Giladi 2006; Lach et al. 2010).
One way to experimentally assess the relative importance of elaiosomes for seed removal is to compare seed removal rates from elaiosome-bearing seeds with seeds to which the elaiosome has been manually removed. Such experiments invariably show the importance of elaiosomes, but can also be informative about differences in their relative importance across environmental conditions (Hughes and Westoby 1992; Castro et al. 2009; Bas et al. 2009; Montesinos et al. 2012). In this study, we quantified not only seed removal, but also the relative importance of elaiosomes in the establishment of seed–ant interactions along increasing tree densities of the two acacia species in the non-native range of Portugal, and compared it with seed removal in their native range in Australia. Specifically, our goals were: (1) to assess myrmecochory and the relative importance of acacia elaiosomes in the native range of Australia and in the non-native range of Portugal; (2) to quantitatively describe the ant community composition at each site; and (3) to assess the impact of acacia invasion in seed removal by ants for native plant species along a gradient of acacia tree density, from highly dense acacia patches to control areas free of acacia.
Methodology
Study species
Acacia dealbata and A. longifolia are perennial trees native to Australia, and invasive in several regions of the world, including Southern Europe, South Africa, South East Asia, and North and South America (Richardson et al. 2011; Rejmánek and Richardson 2013). Trees have a lifespan of 20–50 years (Richardson et al. 2011). Both species were introduced in Portugal early in the twentieth century as ornamental trees or as forestry species for soil stabilization (De Almeida and Freitas 2006; Lorenzo et al. 2010a) but, after expanding out of their introduction sites during the last decade, are nowadays two of the most invasive species in Portugal (Lorenzo et al. 2010a; Marchante et al. 2010; Gibson et al. 2011). Acacia dealbata inhabits mountain ranges and road sides of continental regions, whereas A. longifolia inhabits coastal sand dunes (De Almeida and Freitas 2006; Rodríguez-Echeverría et al. 2009; Lorenzo et al. 2010a; Buscardo et al. 2010). Both acacia species disperse seed in summer in their native range and non-native range of Portugal (Castroviejo 2001; Pellow et al. 2009).
Study sites
In Portugal A. dealbata was studied in the mountainous area of Lousã (40.102568, − 8.233698); while Acacia longifolia was studied at the secondary dunes of Tocha (40.328420, − 8.807237, datum WSG84). Both sites experience sub-humid Mediterranean climate. The Lousã population is located 70 km inland from Tocha; the mean annual precipitation is 752 mm and mean monthly temperatures range from 3.0 °C in January to 17.6 °C in August, with an annual mean of 9.2 °C. Vegetation is composed mostly by a natural mixed oak forest dominated by Quercus canariensis Willd., Castanea sativa Mill., P. pinaster Aiton, but with large patches of expanding A. dealbata populations; with an understory dominated by Erica australis L., E. arborea L., Pterospartum tridentatum Willk. and Lithodora fruticosa Griseb. Tocha’s mean annual precipitation is 948 mm and mean monthly temperature ranging from 10.2 °C in January to 20.2 °C in June, with an annual mean of 16.2 °C. The population is found on a sand dune system. Vegetation is composed of sown and planted trees of Pinus pinaster and A. longifolia, and characteristic sand dune species of herbs and small shrubs such as Corema album (L.) D. Don ex Steud., Halimium halimifolium Willk., Cistus salviifolius Boiss., Cytisus grandiflorus DC., Cytisus striatus (Hill) Rothm., Euphorbia paralias L., Ulex europaeus L., and Erica australis L.
In Australia, A. dealbata was studied nearby Point Hut Pond District Park, Canberra (− 35.452037, 149.075166); while A. longifolia was studied at Puckeys Estate Reserve, Wollongong (− 34.399254, 150.903614). Point Hut Pond presents a dry continental climate, with an average annual precipitation of 616 mm, and mean monthly temperatures ranging from 5.7 °C in July to 20.6 °C in July, with an annual mean of 13.1 °C. The population is located in a relatively well conserved area. Vegetation is composed by A. dealbata trees and several species of Eucalyptus, and numerous understory species, including Hardenbergia violacea, and species of the genus Banksia, Danthonia, and Themeda. Puckeys State has an oceanic climate, with an average annual precipitation of 1321 mm, and mean monthly temperatures ranging from 12.6 °C in July to 21.7 °C in January, with an annual mean of 17.5 °C. The population occurs in a moderately well-preserved sand dune system. Vegetation is composed by naturally occurring A. longifolia and Eucalyptus botryoides trees, Leptospermum laevigatum shrubs, and by characteristic sand dune species, including species of the genera Spinifex and Banksia.
Experimental set-up
In each of the two non-native sites in Portugal, each dominated by one of the studied acacia species, we randomly selected during 2013 up to ten observation points, separated by at least 20 m from each other, within each of three types of areas: control areas, where no acacia trees were present in a 25 m radius; low acacia densities, where A. dealbata density was 0.30 trees m−2 and A. longifolia was 0.13 trees m−2; and high acacia tree density, with A. dealbata densities of 5.40 trees m−2 and A. longifolia densities of 1.00 tree m−2 (N = 30 observation points per site in Portugal). With this design, we aimed to have a representative sampling of a gradient of invasion within each site, from control uninvaded areas, to the center of highly invaded patches within each site. In the native sites of Australia, during December 2012, we proceeded similarly but with only one kind of area, representing a typical acacia population for each species. Acacia dealbata tree density was 3.31 trees m−2 at Point Hut and A. longifolia tree density was 0.91 trees m−2 at Puckeys (N = 10 observation points in Australia). Thus, for both acacia species, density of Australian populations were just slightly lower than “high density” Portuguese sites. Tree densities were calculated based on a 100 m2 survey (ten 10 × 1 m transects) across each area and site.
We could find at least one plant species with elaiosome-bearing seeds dispersing synchronously with acacia for each Portuguese site. Those species were from the Fabaceae Pterospartum tridentatum at the mountain area of Lousã, invaded by A. dealbata; and Ulex europaeus at the sand dunes of Tocha, invaded by A. longifolia. In Australia, we could not find any other plant species dispersing elaiosome-bearing seeds synchronously with acacias. At each study site, we collected abundant fresh seed both from the acacia and from the co-occurring native species, and manually removed the elaiosome from half of the seeds, to be used immediately in subsequent experiments and observations, coinciding with both species natural seed dispersal phenology.
Seed removal rates
In order to assess the importance of the elaiosome for seed removal by ants we set up a seed removal experiment. We prepared Petri dishes by boring three 5 mm holes in three equidistant lateral points, separated by 5 cm and in direct contact with the ground in order to allow access to the interior of the dish only to ants. In Portugal, we set up two sets of paired Petri dishes, one for each seed species (i.e. acacia, and the native species present at each Portuguese site). Each dish contained either 10 elaiosome-bearing seeds or 10 elaiosome-removed seeds from a single species. Paired Petri dishes were placed in 10 observation points within each density area in Portugal (total of 60 dishes per seed species and site, N = 120), and in each acacia population in Australia (total of 20 dishes). After 48 h we counted the remaining seeds at each Petri dish. Untransformed seed removal rates were analyzed with Generalized Linear Models with Poisson distribution of errors in SPSS 19 (IBM 2010). For each region, seed removal per 48 h was the response variable for each species, and elaiosome presence and acacia density were fixed factors.
Relative importance of elaiosomes for seed removal
Ant interactions with seeds can be defined as either granivorous or myrmecochorous, and although it is difficult to assess their relative importance in natural conditions, it is commonly accepted that the difference in seed removal rates between elaiosome-bearing and elaiosome-removed seeds is a good indicator of the relative importance of myrmecochory (Willson and Traveset 2000). This can be expressed as:
where Mri is the relative importance of myrmecochory and Ie and Ine represent the number of observed interactions with elaiosome bearing or elaiosome-removed seeds, respectively. However, this indicator is based on counts and would impede adequate comparison among different experiments or species, like the ones used in this work.
Consequently, we used a proportional term in order to produce a normalized index which would be fully comparable, and that overcomes the poor statistical properties of ratios (Hedges et al. 1999). Specifically, we adapted the Relative Interaction Indexes (RII) (Armas et al. 2004) to assess the relative importance of these appendages across regions and environmental conditions for seed dispersal. The index was originally conceived to assess the sign (competition/facilitation) of plant–plant interactions based on continuous biomass variables, whereas ant–seed interactions are typically recorded as counts. We adapted RII to necessarily take into account the maximum number of possible ant–seed interactions, i.e. the number of seeds with or without elaiosome offered to ants. Therefore, we used a Relative Interaction Index adapted to elaiosome-seed removal (eRII) as:
in which Ieo and Ineo represent the number of elaiosome-bearing or elaiosome-removed seeds offered in each observational unit. Mathematically, the index can range from − 1 to 1, with negative and positive values indicating lower or higher relative importance of myrmecochory, respectively. However, in real cases we should never expect a higher number of interactions for elaiosome-removed than for elaiosome-bearing seeds, since granivorous ants should not distinguish between them. Consequently, the index will range from zero to one in real field studies. Values nearing zero will indicate a small relative importance of elaiosomes (e.g. because of a predominance of granivorous ants), and values nearing one will indicate a high importance of myrmecochory (e.g. because of the absence of granivorous ants, and the almost exclusive activity of myrmecochorous ants).
We used seed removal rates from the experiment described above to calculate eRII. Each pair of Petri dishes with either elaiosome-bearing or elaiosome-removed seeds (N = 10 pairs), within each area (N = 3 densities for Portugal; N = 1 for Australia) and region (N = 2 countries), was used to obtain an individual value for the eRII for each seed species, which was later analyzed statistically with R 3.1.2 (R Development Core Team 2010) by means of Generalized Linear Models with Gaussian distribution, with eRII as the dependent variable, calculated from seed removal from each individual pair of Petri dishes. We compared differences between native and invaded regions, using region and plant species as fixed factors. We also compared differences between native and introduced species in Portugal, using acacia density as a fixed factor.
Ant species richness
We set up one pitfall trap at each observation point filled with a 1:2 mix of 90% alcohol and glycerin. Pitfall traps were removed after 48 h and all ant individuals found within the traps were identified taxonomically (Shattuck 1999; www.hormigas.org). We used elaiosome-bearing and elaiosome-removed seeds to perform cafeteria experiments at the observation points, in which we offered simultaneously four seeds of each plant species; two with their natural elaiosome, and two with the elaiosome-removed. Each group of four seeds was placed on the natural ground randomly, and visual observations were made in 10-min intervals at different locations within each site. We observed ants visiting the Petri dishes and counted an interaction when the ant species picked up a seed and tried to carry it away. We collected these ants for taxonomical identification and replaced the removed seed until the 10-min interval ended. We made observations from dawn to dusk on three different days for each site, and accumulated a total observation time of 293 and 525 min for A. dealbata sites in Portugal and Australia, respectively; and of 400 and 420 min for A. longifolia sites in Portugal and Australia, respectively. In order to assess that we had a sufficient and representative number of observations we created rarefaction curves using EstimateS 9.1.0 (Colwell 2013) with 500 randomizations with replacement. Curves showed saturation or near-saturation for all sites (Appendix 1: Fig. 3).
To describe the ant community for each area and site we calculated ant species richness, Shannon–Wiener diversity index (H′), Pielou’s evenness index (J′), and α and βW diversity. Shannon–Wiener diversity index was calculated as: H′ = ∑ pi (ln pi), were pi was the proportional abundance of each species; and Pielou’s evenness index as: J′ = H′/H′max, with H′max being the maximum value of H′ for a given community, calculated as the natural logarithm of the total number of species present in that site. Pielou’s J′ ranges from zero to one, with one indicating the highest possible evenness and zero the lowest. In order to account for abundance effects, evenness was also assessed with Hulbert’s probability of specific encounter, PIE (Hurlbert 1971), which measures the chance that two individuals randomly sampled pertain to two different species. β diversity was used to assess the variation in species composition among sites, and provides an interesting link between ant diversity (α diversity) and the total species pool within the area (γ diversity) (Anderson et al. 2011). We calculated the commonly used Whittaker’s β diversity index Bw (Whittaker 1972) in which βW = (γ/α − 1), being γ the total number of ant species at the area and α the mean number of ant species interacting with seeds of each plant species. Standardized Sørensen’s β diversity was also assessed with the procedure beta.multi of R’s library betapart (Baselga and Orme 2012).
Results
Seed removal rates
In the A. dealbata site in Portugal, elaiosome-bearing seeds experienced significantly higher removal rates both for A. dealbata (Wald χ2 = 14.06, df = 1, p ≤ 0.001), and for P. tridentatum (Wald χ2 = 15.79, df = 1, p ≤ 0.001). Seed removal rates were highest in areas with no acacias and lowest in areas with higher acacia densities both for A. dealbata (Wald χ2 = 78.19, df = 2, p ≤ 0.001) and for the native P. tridentatum (Wald χ2 = 30.48, df = 2, p ≤ 0.001). Significant interactions between the effects of elaiosome presence and acacia tree density were found both for the invasive (Wald χ2 = 6.42, df = 2, p = 0.040), and for the native seeds (Wald χ2 = 12.13, df = 2, p ≤ 0.002), indicating that removal rates of elaiosome-bearing and elaiosome-removed seeds varied differently among different acacia densities. In Australia, the presence of elaiosomes had a similarly positive and significant effect on seed removal rates, but of a much larger magnitude that it did in the invaded Portugal (Wald χ2 = 39.46, df = 1, p ≤ 0.001) (Fig. 1).

Ant seed removal rates for elaiosome-bearing and elaiosome-removed seeds of each acacia species and accompanying elaiosome-bearing native species at both Australia and Portugal (mean ± SE), across a gradient of acacia tree density, with controls representing areas with no acacia presence. Different uppercase or lowercase letters indicate statistically significant differences among densities, or between elaiosome removal treatments, respectively
Acacia longifolia presented a seed removal pattern resembling that of A. dealbata sites both in Portugal and in Australia, however, the importance of the presence of elaiosomes was of a higher magnitude both for the invasive acacia in the non-native range and for the co-occurring native U. europaeus. Elaiosome-bearing seeds experienced significantly higher removal rates both for A. longifolia (Wald χ2 = 258.93, df = 1, p ≤ 0.001) and for the native U. europaeus (Wald χ2 = 627.85, df = 1, p ≤ 0.001). Seed removal rates of acacia were significantly higher in areas with no acacias (control), and lowest in areas with higher acacia densities (Wald χ2 = 425.03, df = 2, p ≤ 0.001). For the native U. europaeus removal rates were also lower in high acacia density areas (Wald χ2 = 25.35, df = 2, p ≤ 0.001) whereas control and low density areas experienced similar removal rates (Wald χ2 = 0.97, df = 2, p = 0.325). Significant interactions between elaiosome presence and acacia density both for the invasive (Wald χ2 = 62.23, df = 2, p ≤ 0.001), and for the native (Wald χ2 = 64.26, df = 2, p ≤ 0.001), signposted for unequal effect of elaiosomes among environments, driven by the large differences in removal rates of elaiosome-bearing and elaiosome-removed seeds found in the high-density site. In Australia, elaiosome presence on A. dealbata seeds had a highly significant and important effect on seed removal rates, with a higher relative importance in Australia than in high density acacia stands in Portugal, but lower proportional importance than in control and low density Portuguese acacia stands (Wald χ2 = 36.88, df = 1, p ≤ 0.001) (Fig. 1).
Relative importance of elaiosomes for seed removal
For sites with A. dealbata presence, eRII’s significantly differed among sites (t = 3.474; df = 62; p ≤ 0.001). Tukey post hoc tests showed that the relative importance of elaiosomes was highest for Australian acacias, and lowest for Portuguese control sites and for high density acacia stands (p = 0.003; p = 0.025; p = 0.009; Fig. 2).

Elaiosome removal relative interaction index (eRII; mean ± SE) for sites with A. dealbata or A. longifolia presence. PT stands for Portugal. Different letters indicate statistically significant differences among groups
For sites with A. longifolia presence, elaiosomes relative importance also presented significant differences among groups (t = − 2.371; df = 68; p = 0.021). However, Tukey post hoc test indicated that the only clearly different group was high density A. longifolia in Portugal, which presented the lowest value of all (p = 0.006; p < 0.001; p = 0.062; p = 0.054; Fig. 2).
Ant species richness
The combination of pitfall traps and direct observations in field cafeteria experiments allowed us to identify 20 different ant species in Portugal and 13 ant species in Australia. Of those, the highest proportion of seed disperser ant taxa was found in Australia (63–78%), whereas myrmecochorous taxa in Portugal was similar among environments for A. longifolia (34–50%), but slightly higher in control areas (67%) for A. dealbata (vs. 50% in low and high density stands) (Tables 1, 2).
Within the invaded A. dealbata mountain site at Lousã, species richness, diversity, and evenness presented a descending cline of ant species richness, with a richer (γ), more diverse (H′) and even (J′, PIE) ant community at control sites, where no acacia trees were present, and a poorer, less diverse and less even ant species community at high acacia density locations (Table 2). In terms of beta diversity (βW) the pattern was the reverse to that of diversity (H′), with more diverse environments presenting a more stable composition of ant species along the control areas, and a highly variable ant community among heavily invaded areas. However, Sørensen’s standardized dissimilarity index showed that species turnover was very similar across environments, albeit with a tendency to increase in control and native areas. The Australian site of Point Hut presented values for all indices more similar to the intermediate values found for low acacia density areas in Portugal, even though Australian sites presented tree densities closer to those categorized as “high density” in Portugal.
Within the A. longifolia invaded sand dunes of Tocha, in Portugal, ants had similar species richness (γ), diversity index (H′), and evenness (J′, PIE), among acacia tree densities. Beta diversity (βW) increased slightly as acacia tree density decreased, in a pattern contrary to that found for A. dealbata, although of a much lower magnitude. The Australian site of Puckeys presented a higher ant richness than the site in Portugal, with medium low diversity index (H′) and evenness (J′, PIE) due to the extraordinary importance of one ant species (Rhytidoponera metallica), which accounted for most of the ant observations across sampled areas within the population; this also resulted in a slightly higher beta diversity (βW). Sørensen’s standardized dissimilarity index βSOR showed that species turnover was slightly lower in high and low density areas and slightly higher in control and native areas.
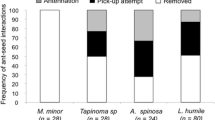
Overall, the number of interactions per minute were many-fold higher in Australia than in Portugal for both acacia species. In the cafeteria experiments, numerous ant species interacted with seeds of both acacia species in Australia, but Pheidole spp. ants proved to be the most important seed dispersing genus of both acacia species, summing up to 80% of interactions for A. dealbata, and 42% for A. longifolia (Appendix 2, Table 3, Fig. 4). In the Portuguese site with A. dealbata all five ant species interacted with seeds from both native and invasive plant species, although seeds of the native plant P. tridentatum were removed mainly by the invasive ant Linepithema humile, which presented a species strength of 0.75, and a species specificity index of 0.966 (Appendix 2, Table 4). Quite differently, in the site with A. longifolia in Tocha, seeds of the native U. europaeus were removed by just a single ant native species, Themnothorax pardoi, whereas A. longifolia seeds were removed by four different ant species, including T. pardoi which thus presented the highest species strength (Appendix 2, Table 5). Interestingly, we did not directly observe T. pardoi ants removing U. europaeus seeds from high density areas, but this or some other ant species must be removing those seeds, according to the seed removal data shown in Fig. 1. Comparing the interaction between native species and acacias in Portugal, it is interesting to note that the native P. tridentatum was able to establish two times more interactions with seed dispersers than A. dealbata in Lousã, while in Tocha the opposite pattern was observed, with A. longifolia presenting more than twice as many interactions as U. europaeus (Appendix 2).
Discussion
Our observations support the generalized presence of ant–seed interactions for these acacia species in the non-native range, even in sites where native European species showed specialized interactions. Our results also showed a reduction in the relative importance of elaiosomes for seed removal of A. dealbata seeds in the non-native region and, for A. longifolia, only on densely invaded sites in Portugal. Seed removal rates by ants were many-fold higher in Australia than they were in Portugal for both acacia species, however, they were still an important asset to guarantee seed dispersal of acacia species in the non-native region.
Myrmecochorous ant species prefer larger elaiosomes, and selective forces often favor increases in elaiosome size in order to attract high quality dispersers (Hughes and Westoby 1992; Mark and Olesen 1996; Leal et al. 2014). However, increased investment of resources in larger elaiosomes usually involves compensatory trade-offs, typically in the form of smaller seeds or in a reduction in seed production (Mark and Olesen 1996; Leal et al. 2014). A recent study comparing seed and elaiosome size of A. longifolia and A. dealbata from Portugal and Australia showed a significant increase in seed size in Portugal, and a trend towards reducing the proportion of biomass allocated to elaiosomes (Correia et al. 2016). Another study comparing A. longifolia from native and non-native ranges within Australia found that this species allocated proportionally less resources to elaiosomes in the non-native ranges than in the native ones (Harris et al. 2016). The present study sheds light on one of the potential selective forces that might be responsible for a shift in resource allocation trade-offs, since our observations of seed removal showed that the difference in removal rates of elaiosome-bearing and elaiosome-removed seeds was proportionally lower in Portugal than in Australia, particularly in the high-density acacia stands in Portugal. Consequently, we could expect that selective forces will favor further shifts in resource allocation trade-offs towards reduced proportional investments in elaiosome size in the non-native region of these acacia species, where increased seed size could offer a more beneficial competitive advantage by increasing the success of seedling recruitment (Correia et al. 2016).
Seed removal rates for elaiosome-bearing seeds were higher than removal rates for elaiosome-removed seeds for all species and regions. We cannot confirm the fate of removed seeds, since ants can be either seed dispersers or granivorous seed eaters (Willson and Traveset 2000). Elaiosome-removed seeds are more likely to be removed by granivorous ants, while elaiosome-bearing seeds should attract both granivorous ants and mutualistic ant seed dispersers looking for an elaiosome reward (e.g. Bas et al. 2009). Furthermore, myrmecochorous interactions are sometimes dominated by one or few highly effective ant species (Gove et al. 2007; Manzaneda and Rey 2008, 2009). However, there are no clear boundaries between myrmecochorous and non-myrmecochorous ant species, and while some high quality seed disperser ant species consistently and effectively disperse seeds across long distances, other seed disperser ants eat the elaiosomes at point of discovery (Lach et al. 2010). Even strictly granivorous ants can play a beneficial role for seed dispersal, since seeds stored in their nests are protected from other predators and wildfires, and can be abandoned and able to germinate (Bas et al. 2007; Lach et al. 2010). Thus, most granivorous ant species often play a dual role as both dispersers and predators (Gómez and Espadaler 2013). The important increase in the observed seed removal rates of elaiosome-bearing seeds, and our direct observations of ant–seed interactions with known seed disperser ant species, suggest that most of the elaiosome-bearing seeds were effectively dispersed. In Australia, manual removal of elaiosomes resulted in large and significant reductions in acacia seed removal rates by ants. In non-native Portugal, elaiosomes were also important, but the effect of elaiosome removal varied between acacia species and among acacia tree densities. Interestingly, seed removal rates on native Australian sites were similar to seed removal rates of control and low density non-native areas, even though acacia tree densities found in Australia were closer to tree densities of highly dense invaded areas. Thus, in the non-native region, seed removal was always higher in non-invaded areas, revealing a potential mechanism for the dispersal of acacia seeds into uninvaded areas.
Portuguese native plant species fared similarly to their co-occurring acacia species in terms of seed-removal rates, even though acacia seeds and their elaiosomes are more than twice as large as those of the studied native species (Castroviejo 2012). Differences in removal rates along the acacia density gradient can be explained by differences in the ant community composition, which is frequently driven by changes in the plant community due to invasions (Traveset and Richardson 2006). Preliminary studies showed differences in seed removal rates along environmental gradients for the same study system (Montesinos et al. 2012), but there was no information about the ant community responsible for that pattern. In the current study, we found that the ant community of the sand dunes invaded by A. longifolia was resilient to differences in acacia tree density, but that there was a clear reduction in ant species richness with increasing A. dealbata tree density. Similar results had been previously reported for A. saligna invaded Fynbos in South Africa (French and Major 2001). However, species richness should not always be regarded as beneficial per se, since some of the sites not invaded by acacias accounted with the presence of invasive ant species, and the presence of invasive ant species also has a deep impact on local ant community compositions which could act synergistically with the effect of plant invasions in still unpredictable ways.
Myrmecochory has been regarded as a generalized form of interaction, although evidence is contradictory. In numerous cases one single ant species can be responsible for most interactions, most likely due to intrinsic traits of the seeds more than the specific ecological context, which tends to be highly variable at temporal and spatial scales (Gove et al. 2007; Manzaneda and Rey 2009). Australian sites presented many-fold more interactions per minute than Portuguese sites. Acacia dealbata showed a specialized pattern of interaction in the study site of Australia, where one single ant species was responsible for two-thirds of the observed interactions. In Portugal, the same acacia species was served by the same array of ant species as the native P. tridentatum. In contrast, A. longifolia was served by a diverse group of ant species both in the sites of Australia and in Portugal, however, the native U. europaeus was served by only one species in this site, demonstrating a very specialized pattern in contrast with the generalist pattern showed by the invasive tree in Portugal. Although these patterns are interesting, further investigation would be needed at a range of native and non-native sites to confirm the generality of our findings.
Plants introduced into new geographical ranges usually encounter fewer dispersal barriers than those that arrive naturally (Traveset and Richardson 2006) and, certainly, seed dispersal mutualisms established by acacias in their non-native range guarantee essential dispersal services. Many factors may significantly contribute to the enormous success of these species in Portugal. For instance, acacias have been found to be highly competitive under low resource supply (Werner et al. 2009), to successfully and advantageously establish belowground mutualisms in the invaded areas (Rodríguez-Echeverría et al. 2009, 2012; Rodríguez-Echeverría 2010), to successfully use allelopathic compounds (Lorenzo et al. 2010b, c), and to produce massive amounts of seeds in the introduced range which are larger and escape pre-dispersal predation (Correia et al. 2014). This study reveals that myrmecochory is important in the non-native regions, and could contribute to the expansion of the invaders.
Overall, our study confirms the importance of elaiosomes for seed removal by ants both in the native and in the non-native regions. However, it also identifies differences in its relative importance between Australia and Portugal and along gradients of acacia density. Our data also shows that invasion affects ant communities and dispersal of native species. Specifically, ant communities change along acacia densities and are impoverished in some highly invaded areas, when compared to uninvaded areas, likely a consequence of abiotic and biotic modifications derived from acacia invasion. Importantly, native plant species experience a significant reduction in seed dispersal services in highly invaded areas, signposting another mechanism of native plant displacement, and exemplifying the difficulty of natives to re-colonize already invaded areas.
References
Aguilera N, Becerra J, Villaseñor-Parada C et al (2015) Effects and identification of chemical compounds released from the invasive Acacia dealbata Link. Chem Ecol 31:479–493. https://doi.org/10.1080/02757540.2015.1050004
Aizen MA, Morales CL, Morales JM (2008) Invasive mutualists erode native pollination webs. PLoS Biol 6:e31. https://doi.org/10.1371/journal.pbio.0060031
Alba-Lynn C, Henk S (2010) Potential for ants and vertebrate predators to shape seed-dispersal dynamics of the invasive thistles Cirsium arvense and Carduus nutans in their introduced range (North America). Plant Ecol 210:291–301. https://doi.org/10.1007/s11258-010-9757-2
Anderson MJ, Crist TO, Chase JM et al (2011) Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecol Lett 14:19–28. https://doi.org/10.1111/j.1461-0248.2010.01552.x
Armas C, Ordiales R, Pugnaire FI (2004) Measuring plant interactions: a new comparative index. Ecology 85:2682–2686. https://doi.org/10.1890/03-0650
Auld TD (1986) Population dynamics of the shrub Acacia suaveolens (Sm.) Willd.: fire and the transition to seedlings. Aust J Ecol 11:373–385. https://doi.org/10.1111/j.1442-9993.1986.tb01407.x
Bartomeus I, Vilà M, Santamaría L (2008) Contrasting effects of invasive plants in plant-pollinator networks. Oecologia 155:761–770. https://doi.org/10.1007/s00442-007-0946-1
Bas JM, Oliveras J, Gómez C (2007) Final seed fate and seedling emergence in myrmecochorous Plants: effects of ants and plant species. Sociobiology 50:101–111. www.csuchico.edu/biol/Sociobiology/sociobiologyindex.html
Bas JM, Oliveras J, Gómez C (2009) Myrmecochory and short-term seed fate in Rhamnus alaternus: ant species and seed characteristics. Acta Oecol 35:380–384. https://doi.org/10.1016/j.actao.2009.02.003
Baselga A, Orme CDL (2012) Betapart: an R package for the study of beta diversity. Methods Ecol Evol 3:808–812. https://doi.org/10.1111/j.2041-210X.2012.00224.x
Blüthgen N, Menzel F, Blüthgen N (2006) Measuring specialization in species interaction networks. BMC Ecol 6:9. https://doi.org/10.1186/1472-6785-6-9
Bossard CC (1991) the role of habitat disturbance, seed predation and ant dispersal on establishment of the exotic shrub cytisus scoparius in California. Am Midl Nat 126:1–13. https://doi.org/10.2307/2426145
Burgos E, Ceva H, Perazzo RPJ et al (2007) Why nestedness in mutualistic networks? J Theor Biol 249:307–313. https://doi.org/10.1016/j.jtbi.2007.07.030
Buscardo E, Rodríguez-Echeverría S, Martín MP et al (2010) Impact of wildfire return interval on the ectomycorrhizal resistant propagules communities of a Mediterranean open forest. Fungal Biol 114:628–636. https://doi.org/10.1016/j.funbio.2010.05.004
Castro S, Ferrero V, Loureiro J et al (2009) Dispersal mechanisms of the narrow endemic Polygala vayredae: dispersal syndromes and spatio-temporal variations in ant dispersal assemblages. Plant Ecol 207:359–372. https://doi.org/10.1007/s11258-009-9679-z
Castroviejo S (2001) Claves de Flora Iberica: plantas vasculares de la Península Ibérica e Islas Baleares. Editorial CSIC—CSIC Press, Madrid
Castroviejo S (2012) Flora ibérica: plantas vasculares de la Península Ibérica e Islas Baleares. Real Jard¡n Botánico, C.S.I.C., Madrid
Colwell RK (2013) EstimateS: statistical estimation of species richness and shared species from samples. Version 9.1.0
Correia M, Castro S, Ferrero V et al (2014) Reproductive biology and success of invasive Australian acacias in Portugal. Bot J Linn Soc 174:574–588. https://doi.org/10.1111/boj.12155
Correia M, Montesinos D, French K, Rodríguez-Echeverría S (2016) Evidence for enemy release and increased seed production and size for two invasive Australian acacias. J Ecol 104:1391–1399. https://doi.org/10.1111/1365-2745.12612
Davidson DW, Morton SR (1984) Dispersal adaptations of some Acacia species in the Australian arid zone. Ecology 65:1038–1051
De Almeida JD, Freitas H (2006) Exotic naturalized flora of continental Portugal—a reassessment. Bot Complut 30:117–130
Dormann CF, Fruend J, Bluethgen N, Gruber B (2009) Indices, graphs and null models: analyzing bipartite ecological networks. Open Ecol J 2:7–24
Dunne JA, Williams RJ, Martinez ND (2002) Food-web structure and network theory: the role of connectance and size. Proc Natl Acad Sci USA 99:12917–12922. https://doi.org/10.1073/pnas.192407699
Ferrero V, Castro S, Costa J et al (2013) Effect of invader removal: pollinators stay but some native plants miss their new friend. Biol Invasions 15:2347–2358. https://doi.org/10.1007/s10530-013-0457-4
Fokuhl G, Heinze J, Poschlod P (2012) Myrmecochory by small ants—beneficial effects through elaiosome nutrition and seed dispersal. Acta Oecol 38:71–76. https://doi.org/10.1016/j.actao.2011.09.007
French K, Major RE (2001) Effect of an exotic Acacia (Fabaceae) on ant assemblages in South African fynbos. Austral Ecol 26:303–310. https://doi.org/10.1046/j.1442-9993.2001.01115.x
Gibson MR, Richardson DM, Marchante E et al (2011) Reproductive biology of Australian acacias: Important mediator of invasiveness? Divers Distrib 17:911–933. https://doi.org/10.1111/j.1472-4642.2011.00808.x
Giladi I (2006) Choosing benefits or partners: a review of the evidence for the evolution of myrmecochory. Oikos 112:481–492. https://doi.org/10.1111/j.0030-1299.2006.14258.x
Gómez C, Espadaler X (1998) Myrmecochorous dispersal distances: a world survey. J Biogeogr 25:573–580
Gómez C, Espadaler X (2013) An update of the world survey of myrmecochorous dispersal distances. Ecography (Cop) 36:1193–1201. https://doi.org/10.1111/j.1600-0587.2013.00289.x
Gorb EV, Gorb SN (1999) Dropping rates of elaiosome-bearing seeds during transport by ants (Formica polyctena Foerst.): implications for distance dispersal. Acta Oecol 20:509–518
Gove AD, Majer JD, Dunn RR (2007) A keystone ant species promotes seed dispersal in a “diffuse” mutualism. Oecologia 153:687–697. https://doi.org/10.1007/s00442-007-0756-5
Harris CJ, Manea A, Moles AT et al (2016) Differences in life-cycle stage components between native and introduced ranges of five woody Fabaceae species. Austral Ecol. https://doi.org/10.1111/aec.12456
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecology 80:1150–1156. https://doi.org/10.1890/0012-9658(1999)080%5b1150:TMAORR%5d2.0.CO;2
Hierro JL, Maron JL, Callaway RM (2005) A biogeographical approach to plant invasions: the importance of studying exotics in their introduced and native range. J Ecol 93:5–15. https://doi.org/10.1111/j.1365-2745.2004.00953.x
Holmes PM (1990) Dispersal and predation in alien Acacia. Oecologia 83:288–290
Hughes L, Westoby M (1992) Effect of diaspore characteristics on removal of seeds adapted for dispersal by ants. Ecology 73:1300–1312. https://doi.org/10.2307/1940677
Hurlbert SH (1971) The nonconcept of species diversity: a critique and alternative parameters. Ecology 52:577–586. https://doi.org/10.2307/1934145
IBM (2010) IBM SPSS Statistics for Windows
Jensen JM, Six DL (2006) Myrmecochory of the exotic plant, Centaurea maculosa: a potential mechanism enhancing invasiveness. Environ Entomol 35:326–331. https://doi.org/10.1603/0046-225X-35.2.326
Lach L, Parr CL, Abbott KL (2010) Ant ecology. Oxford University Press, Oxford
Lankau RA, Nuzzo V, Spyreas G, Davis AS (2009) Evolutionary limits ameliorate the negative impact of an invasive plant. Proc Natl Acad Sci USA 106:15362–15367. https://doi.org/10.1073/pnas.0905446106
Leal LC, Neto MCL, de Oliveira AFM et al (2014) Myrmecochores can target high-quality disperser ants: variation in elaiosome traits and ant preferences for myrmecochorous Euphorbiaceae in Brazilian Caatinga. Oecologia 174:493–500. https://doi.org/10.1007/s00442-013-2789-2
Lengyel S, Gove AD, Latimer AM et al (2009) Ants sow the seeds of global diversification in flowering plants. PLoS ONE 4:e5480. https://doi.org/10.1371/journal.pone.0005480
Lengyel S, Gove AD, Latimer AM et al (2010) Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey. Perspect Plant Ecol Evol Syst 12:43–55. https://doi.org/10.1016/j.ppees.2009.08.001
Lorenzo P, González L, Reigosa MJ (2010a) The genus Acacia as invader: the characteristic case of Acacia dealbata Link in Europe. Ann For Sci 67:101. https://doi.org/10.1051/forest/2009082
Lorenzo P, Palomera-Pérez A, Reigosa MJ, González L (2010b) Allelopathic interference of invasive Acacia dealbata Link on the physiological parameters of native understory species. Plant Ecol 212:403–412. https://doi.org/10.1007/s11258-010-9831-9
Lorenzo P, Pazos-Malvido E, Reigosa MJ, González L (2010c) Differential responses to allelopathic compounds released by the invasive Acacia dealbata Link (Mimosaceae) indicate stimulation of its own seed. Aust J Bot 58:546–553
Manzaneda AJ, Rey PJ (2008) Geographic variation in seed removal of a myrmecochorous herb: influence of variation in functional guild and species composition of the disperser assemblage through spatial and temporal scales. Ecography (Cop) 31:583–591. https://doi.org/10.1111/j.0906-7590.2008.05345.x
Manzaneda AJ, Rey PJ (2009) Assessing ecological specialization of an ant–seed dispersal mutualism through a wide geographic range. Ecology 90:3009–3022. https://doi.org/10.2307/25592842
Marchante H, Freitas H, Hoffmann JH (2010) Seed ecology of an invasive alien species, Acacia longifolia (Fabaceae), in Portuguese dune ecosystems. Am J Bot 97:1780–1790. https://doi.org/10.3732/ajb.1000091
Mark S, Olesen JM (1996) Importance of elaiosome size to removal of ant-dispersed seeds. Oecologia 107:95–101. https://doi.org/10.1007/BF00582239
Memmott J, Waser NM, Price MV (2004) Tolerance of pollination networks to species extinctions. Proc Biol Sci 271:2605–2611. https://doi.org/10.1098/rspb.2004.2909
Montesinos D, Castro S, Rodríguez-Echeverría S (2012) Invasive acacias experience higher ant seed removal rates at the invasion edges. Web Ecol 12:33–37. https://doi.org/10.5194/we-12-33-2012
Montesinos D, Castro S, Rodríguez-Echeverría S (2016) Two invasive acacia species secure generalist pollinators in invaded communities. Acta Oecol 74:46–55. https://doi.org/10.1016/j.actao.2016.06.002
Pellow BJ, Henwood MJ, Carolin RC (2009) Flora of the Sydney region: a complete revision, 5th edn. Sydney University Press, Sydney
Pemberton RW, Irving DW (1990) Elaiosomes on weed seeds and the potential for myrmecochory in naturalized plants. Weed Sci 38:615–619
Phillips BL, Brown GP, Webb JK, Shine R (2006) Invasion and the evolution of speed in toads. Nature 439:803. https://doi.org/10.1038/439803a
Phillips BL, Brown GP, Greenlees M et al (2007) Rapid expansion of the cane toad (Bufo marinus) invasion front in tropical Australia. Austral Ecol 32:169–176. https://doi.org/10.1111/j.1442-9993.2007.01664.x
Prior KM, Robinson JM, Dunphy SA, Frederickson ME (2015) Mutualism between co-introduced species facilitates invasion and alters plant community structure. Proc Biol Sci 282:20142846. https://doi.org/10.1098/rspb.2014.2846
R Development Core Team (2010) R: a language and environment for statistical computing. R Found Stat Comput Vienna Austria 0. ISBN 3-900051-07-0
Rejmánek M, Richardson DM (2013) Trees and shrubs as invasive alien species—2013 update of the global database. Divers Distrib 19:1093–1094. https://doi.org/10.1111/ddi.12075
Richardson DM, Allsopp N, D’Antonio CM et al (2000) Plant invasions—the role of mutualisms. Biol Rev Camb Philos Soc 75:65–93
Richardson DM, Carruthers J, Hui C et al (2011) Human-mediated introductions of Australian acacias—a global experiment in biogeography. Divers Distrib 17:771–787. https://doi.org/10.1111/j.1472-4642.2011.00824.x
Rodríguez-Echeverría S (2010) Rhizobial hitchhikers from down under: Invasional meltdown in a plant-bacteria mutualism? J Biogeogr 37:1611–1622. https://doi.org/10.1111/j.1365-2699.2010.02284.x
Rodríguez-Echeverría S, Crisóstomo JA, Nabais C, Freitas H (2009) Belowground mutualists and the invasive ability of Acacia longifolia in coastal dunes of Portugal. Biol Invasions 11:651–661. https://doi.org/10.1007/s10530-008-9280-8
Rodríguez-Echeverría S, Fajardo S, Ruiz-Díez B, Fernández-Pascual M (2012) Differential effectiveness of novel and old legume–rhizobia mutualisms: implications for invasion by exotic legumes. Oecologia 170:253–261. https://doi.org/10.1007/s00442-012-2299-7
Rodríguez-Echeverría S, Afonso C, Correia M et al (2013) The effect of soil legacy on competition and invasion by Acacia dealbata Link. Plant Ecol 214:1139–1146. https://doi.org/10.1007/s11258-013-0238-2
Rodriguez-Girones MA, Santamaria L (2006) A new algorithm to calculate the nestedness temperature of presence-absence matrices. J Biogeogr 33:924–935. https://doi.org/10.1111/j.1365-2699.2006.01444.x
Sallabanks S (1993) Fruiting plant attractiveness to avian seed dispersers: native versus invasive Crataegus in Western Oregon. Madrono 40:108–116
Shattuck S (1999) Australian ants. Their biology and identification. CSIRO Publishing, Sydney
Simberloff D, Von Holle B (1999) Positive interactions of nonindigenous species: Invasional meltdown? Biol Invasions 1:21–32
Smith JMB (1989) An example of ant-assisted plant invasion. Aust J Ecol 14:247–250. https://doi.org/10.1111/j.1442-9993.1989.tb01433.x
Traveset A, Richardson D (2006) Biological invasions as disruptors of plant reproductive mutualisms. Trends Ecol Evol 21:208–216. https://doi.org/10.1016/j.tree.2006.01.006
Traveset A, Richardson DM (2011) Mutualisms : Key drivers of invasions … key casualties of invasions. Fifty years invasion Ecol. Leg. Charles Elt. Blackwell Publishing Ltd, pp 143–160
Traveset A, Richardson DM (2014) Mutualistic interactions and biological invasions. Annu Rev Ecol Evol Syst 45:89–113. https://doi.org/10.1146/annurev-ecolsys-120213-091857
Werner C, Zumkier U, Beyschlag W, Máguas C (2009) High competitiveness of a resource demanding invasive acacia under low resource supply. Plant Ecol 206:83–96. https://doi.org/10.1007/s11258-009-9625-0
Whitney KD (2002) Dispersal for distance? Acacia ligulata seeds and meat ants Iridomyrmex viridiaeneus. Austral Ecol 27:589–595. https://doi.org/10.1046/j.1442-9993.2002.01216.x
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21:213–251
Williams PA, Karl BJ (2002) Birds and small mammals in kanuka (Kunzea ericoides) and gorse (Ulex europaeus) scrub and the resulting seed rain and seedling dynamics. N Z J Ecol 26:31–41
Willson MF, Traveset A (2000) The ecology of seed dispersal. In: Fenner M (ed) Seeds the ecology of regeneration plant communities, 2nd edn. CAB International, Wallingford, pp 85–110
Acknowledgements
Thanks to Andreia Jorge for help in the lab and to Steve Shattuck and Xavier Espadaler for confirmation of ant identification. Research was Funded by the Portuguese Fundação para a Ciência e a Tecnologia (FCT) via Project Mutualnet (PCT/BIA-BEC/103507/2008), co-founding by the EU via QREN, COMPETE and FEDER (UID/BIA/04004/2013), and scholarships and Grants to DM (SFRH/BPD/72595/2010; Starting grant IF/00066/2013), SC (Starting Grant IF/01267/2013), and SRE (Development Grant IF/00462/2013).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Appendices
Appendix 1: Rarefaction curves
See Fig. 3.

Rarefaction curves for ant–seed interactions observed at each of the study sites
Appendix 2: Interaction networks
See Fig. 4 and Tables 3, 4 and 5.

Ant–seed mutualistic networks based on the observed interactions rates (seed removal) for sites with A. dealbata (top), or A. longifolia (below), on either Portugal (left), or Australia (right)
To visualize the relative strength of interactions on each site, we calculated ant–seed interaction networks for each of the two Portuguese sites with the statistical package “bipartite” on R 3.1.2 (Dormann et al. 2009). For each network, we calculated the following indexes: connectance, indicative of the realized proportion of possible links, obtained by the total sum of links divided by the number of cells in the interaction matrix (Dunne et al. 2002); nestedness (weighted NODF) indicating how the system is organized, with values closest to zero indicating high nestedness and values nearing 100 low nestedness (Rodriguez-Girones and Santamaria 2006); and network specialization index H2′, describing the level of specialization of the network, and ranging from zero (no specialization) to 1 (complete specialization) (Blüthgen et al. 2006); animal robustness, measuring the sensitivity of the system to the loss of plant species; and plant robustness, measuring the sensitivity of the system to the loss of animal species (Memmott et al. 2004; Burgos et al. 2007). For detailed descriptions of the different indexes used, see Dormann et al. (2009) and references therein. The study of interaction networks requires at least two species at each interaction level. Since we only had one plant species in Australia, we could only calculate interaction indexes for the Portuguese populations, for which two plant species were considered. However, we plotted interaction graphs for the Australian observations to quantitatively compare their interaction strength to those of Portugal.
Rights and permissions
About this article

Cite this article
Montesinos, D., Correia, M., Castro, S. et al. Diminishing importance of elaiosomes for acacia seed removal in non-native ranges. Evol Ecol 32, 601–621 (2018). https://doi.org/10.1007/s10682-018-9959-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10682-018-9959-y