
Abstract
Fish assemblages in tropical lowland rivers are characterized by a high richness of species that feed on a diverse array of food resources. Although closely related species often have similar feeding ecology, species within the family Cichlidae display a broad spectrum of trophic niches, and resource partitioning has been inferred from studies conducted in Neotropical rivers. We investigated interspecific variation in food resource use and its relationship to morphological variation among cichlid fishes within the Pantanos de Centla Biosphere Reserve, a coastal area encompassing the delta of the Grijalva-Usumacinta River in Tabasco, Mexico. Most species consumed benthic crustaceans, aquatic insect larvae, and detritus, but some were more herbivorous, and one species was a specialized piscivore. Dietary niche overlap among species was higher than expected for one assemblage, and similar to random expectations for another, suggesting a lesser role for resource partitioning than has been shown for some cichlid assemblages, perhaps due to availability of abundant resources, even in low-water conditions. Canonical correspondence analysis revealed that greatest morphological differences am7ong species involved functional traits directly associated with resource use. Relationships between feeding ecology and morphology were similar to those described for other riverine cichlids. Strong ecomorphological relationships facilitate inferences about the ecology of cichlid species, including species that currently lack data from field studies. Knowledge of ecological relationships will be important for conservation in the Pantanos de Centla, an ecosystem of global significance for biodiversity and ecosystem services.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In species-rich freshwater fish assemblages, co-occurring species occupy a diverse suite of trophic niches, and some have been found to partition resources, presumably to reduce interspecific competition (Ross 1986; Winemiller and Pianka 1990; Herder and Freyhof 2006). Species within the same family are often ecologically similar due to phylogenetic niche conservatism (e.g., McNyset 2009), but closely related species may occupy diverse trophic niches in regions where ecological adaptive radiations have occurred (Schluter 2000). In the incredibly diverse cichlid communities of Lake Malawi in Africa, Genner et al. (1999a, 1999b) found that cichlids coexisting along rocky shores showed significant differences in food resource use, but many species had considerable dietary overlap. They suggested that ecological segregation may not be necessary to support coexistence in these diverse cichlid assemblages. In more dynamic fluvial habitats, environmental variation often causes shifts in resource availability (Grossman et al. 1998), and trophic segregation may occur during periods when fish densities are high while habitat and food availability are low (Winemiller 1990, 1991b; Winemiller and Kelso-Winemiller 2003).
Cichlids in the Neotropics consume a broad array of resources, from aquatic macrophytes to other fishes, with some being trophic specialists while retaining an ability to exploit a variety of food items (Lowe-McConnell 1991; Winemiller et al. 1995; Montaña and Winemiller 2013). Differences in dietary preferences are likely to be reflected in morphological traits related to foraging and consuming food. Prior studies of riverine cichlids have revealed strong relationships between morphological traits and diet (e.g., Winemiller et al. 1995; López-Fernández et al. 2012; Montaña and Winemiller 2013). For example, Winemiller et al. (1995) found that across continents, riverine cichlids showed convergent relationships between traits such as gut length, gape size, and head length and the proportion of fishes, invertebrates, vegetation, and detritus consumed.
Cichlids comprise a major component of the freshwater fish fauna of Mesoamerica, and the evolution and zoogeography of the group (subfamily Cichlinae) has been studied extensively (e.g., Martin and Bermingham 1998; Hulsey et al. 2004; López-Fernández et al. 2013). Centers of Mesoamerican cichlid diversity, such as the Río Grijalva-Usumacinta region of southern México, contain many coexisting cichlid species that display considerable morphological diversity that likely reflects niche diversification (Myers 1966; Bussing 1985; Pease et al. 2012). Studies in other Mesoamerican regions (e.g., Winemiller et al. 1995; Cochran-Biederman and Winemiller 2010) suggest that coexisting cichlid species partition trophic niches. Relatively little ecological information is available for cichlids of the Lower Grijalva-Usumacinta (Miller 2005) despite the high diversity and importance of many species for small-scale fisheries (Mendoza-Carranza et al. 2013).
In this study we examined diets, niche breadth, and niche overlap of coexisting cichlid species in the Grijalva-Usumacinta River delta within the Pantanos de Centla Biosphere Reserve in Tabasco, México. We also sought to identify relationships between morphological features and the feeding ecology of cichlids in these assemblages. We carried out the study during the dry season, a period when dietary niche partitioning has been shown for other fluvial cichlid assemblages (e.g., Winemiller 1991b; Jepsen et al. 1997). During low-water periods, availability of food resources declines and densities of most fish species increases, which increases the potential for competition. We also hypothesized that morphological variation among cichlids would involve traits directly related to trophic resource use in a manner similar to ecomorphological relationships documented for other fluvial cichlids (e.g., Winemiller et al. 1995; Cochran-Biederman and Winemiller 2010; Montaña and Winemiller 2013). Information on the trophic ecology of cichlid fishes in the Lower Grijalva-Usumacinta should enhance management of fisheries resources and conservation of this region’s unique biodiversity.
Materials and methods
Study sites and data collection
The Grijalva and Usumacinta join together in Tabasco, Mexico to form the largest river of Mesoamerica and a major center of fish diversity (Myers 1966; Bussing 1985; Miller 2005). Approximately 115 fish species from 31 families have been documented in the Grijalva-Usumacinta region, and an estimated 36% of these species are endemic (Miller 2005). Cichlids are the most species-rich family in this region, with many species occurring together in local assemblages (Rodiles-Hernández et al. 1999; Miller 2005; Soria-Barreto and Rodiles-Hernández 2008).
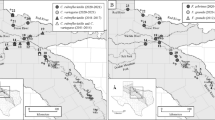
Sampling locations were within the Pantanos de Centla Biosphere Reserve (Fig. 1), a vast protected area within the Lower Grijalva-Usumacinta Basin which was designated by the RAMSAR convention of 1995 as a wetland of international significance. San Pedrito Lagoon (18°20′36”N 92°33′50”W) is a permanent freshwater lagoon with expansive beds of aquatic macrophytes (chiefly Vallisneria americana) and shorelines dominated by reeds (Phragmites australis). Polo Stream (18°29′21”N 92°38′23”W) is a slow-moving, mangrove-dominated freshwater tributary of the Río Grijalva below its confluence with the Río Usumacinta. Along the shorelines of the stream, structural cover is provided by riparian plants (mangrove trees and terrestrial grasses), and the sandy substrate is covered with coarse particulate organic matter. A detailed description of these study sites is given in Mendoza-Carranza et al. (2010) and Sepúlveda-Lozada et al. (2015). Intensive surveys were carried out in February and March 2007 during the dry season (Espinal et al. 2007; Yáñez-Arancibia et al. 2009). In Pantanos de Centla, the dry season is characterized by low water levels (< 2 m in Polo Stream and <1 m in San Pedrito Lagoon), low turbidity (0.90–1.5 m Secchi depth), and low salinity (4–5 UPS) (Arévalo Frías and Carranza 2012; Sepúlveda-Lozada et al. 2015). Precipitation in Tabasco during February and March 2007 was 127.0 and 55.8 mm, respectively, corresponding to the lowest values of the year (Servicio Meteorológico Nacional 2017).

Map of the study area within Pantanos de Centla Biosphere Reserve, Tabasco, Mexico
Specimens were collected using seine nets, gill nets, hook and line, and a boat-mounted trawl net. Though all main mesohabitat types were sampled at the sites, the specimens used for this study were captured among the Vallisneria beds in San Pedrito (depths of 0.5–1 m), and along the vegetated shorelines of Polo Stream (depths of 0.1–1.2 m), areas where cichlids were abundant and diverse. Specimens were identified following Miller (2005) and deposited in the fish collection at El Colegio de la Frontera Sur in Villahermosa, Tabasco.
Given that the objective of this study was to evaluate interspecific patterns of morphological and dietary diversity within local species assemblages, juvenile size classes, which tend to show relatively low interspecific divergence in morphology and diet (i.e., small size classes of all species consume mostly microcrustacea and other small aquatic invertebrates), were excluded from analyses. Species that were rarely captured during surveys of the mesohabitats (N < 5) also were excluded from analysis. Volumetric proportions of stomach contents were estimated following the methods of Winemiller (1990). Fishes consumed were identified to species when possible, and invertebrates were identified to order. Prey items were later grouped into broader categories for statistical analyses. These categories were: fishes, mollusks, decapod crustaceans, aquatic insect larvae, benthic meiofauna, aquatic macrophytes, algae, and detritus.
Morphological measurements were made on five specimens of each species included in the assemblage dataset. Morphological traits were measured to the nearest 0.1 mm using vernier calipers. Following methods described by Winemiller (1991a), the following 24 morphological features were measured (Table 1): maximum standard length, gut length, head length, head depth, oral gape, eye position, eye diameter, snout length, jaw protrusion length, body depth, body width, caudal peduncle depth, caudal peduncle width, body depth below midline, dorsal fin length, dorsal fin height, anal fin length, anal fin height, caudal fin depth, caudal fin length, pectoral fin length, pelvic fin length, gill raker number, and gill raker length. Prior to statistical analysis, morphological measurements were standardized for size by converting to proportions of standard length, body width, or head length as described by Winemiller (1991a).
Statistical analyses
For species at each sampling site, dietary niche breadth was estimated using Levins’s (1968) standardized index. Pianka’s (1973) symmetrical index of niche overlap was calculated as a measure of dietary similarity between species. Both indices were calculated using volumetric proportions of the nine aggregated prey categories above. For both measures, values may range from 0 to 1, with higher values indicating greater diet diversity and more complete overlap. To test whether dietary niche overlap was higher or lower than random expectations, a randomization test (1000 iterations) was also performed using the niche overlap module of EcoSim Professional (Entsminger 2014). We used the “conserved-zeroes” algorithm of Winemiller and Pianka (1990) in the randomization, retaining the niche breadths of species and the zero structure of the food resource use matrix (i.e., resources not used by a species remained unused in the randomization). Mean observed and randomized niche overlaps by nearest neighbor rank were also plotted.
To ordinate species within assemblage morphological space, principal components analysis (PCA) was performed based on the correlation matrix of log-transformed morphological data (species average trait values). Relationships between diet (using data from both survey sites) and morphology were examined using canonical correspondence analysis (CCA). This allowed for measurement of the amount of variation in dietary resource use that could be explained by axes of morphological variables. The statistical significance of the diet-morphology relationship from CCA was assessed with a permutation test (1000 simulations). Both ordination analyses were performed using the Vegan package in R version 2.11.1 (R Foundation for Statistical Computing 2010).
To examine phylogenetic signal in the diet composition of cichlid species, we used the test for phylogenetic serial independence (TFSI, Abouheif 1999; Pavoine et al. 2008) with mean proportions of the eight prey categories above as traits. The phylogeny of Mesoamerican cichlids from Rican et al. (2016) was used, and the analysis was performed using Phylogenetic Independence v.2.0 (http://biology.mcgill.ca/faculty/abouheif/). For each dietary category, a C statistic was calculated for phylogenetic autocorrelation, and topology of the original data was randomized 1000 times to generate a null distribution for assessing statistical significance.
Results
Twelve species of native cichlids were captured during surveys of Pantanos de Centla Biosphere Reserve. The eight most abundant species (Petenia splendida, Vieja melanura, Vieja bifasciata, Mayaheros urophthalmus, Trichromis salvini, Cribroheros robertsoni, Thorichthys pasionis, and Thorichthys helleri) were used for dietary and morphological analyses (total of 323 individuals). Four additional species were collected in small numbers: Cincelicthys pearsei, Maskaheros argenteus, Thorichthys meeki, and Thorichthys socolofi. Thorichthys helleri was by far the most abundant species at both sites (n = 99). All species were present at both sites; more C. robertsoni were collected at the Polo Stream site, and adult size classes of P. splendida were only captured from San Pedrito Lagoon.
Examination of stomach contents revealed that most species consumed benthic meiofauna, aquatic insect larvae, and detritus (Table 2). The two Vieja species were largely herbivorous/detritivorous, consuming large proportions of coarse vegetative detritus and aquatic macrophytes. Petenia splendida was a piscivore with a diet restricted to fishes (mostly juvenile cichlids). The two Thorichthys species and C. robertsoni consumed small benthic invertebrates (benthic meiofauna, insect larvae, and gastropods) along with coarse detritus. Thorichthys helleri consumed a higher proportion of snails than the other invertebrate feeders. Trichromis salvini and M. urophthalmus had more generalist, omnivorous diets composed of shrimp and smaller invertebrates as well as detritus and plant matter. In Polo Stream, all cichlid species consumed substantial proportions of detritus. In general, cichlid species consumed more aquatic plants and mollusks (bivalves and gastropods) in San Pedrito Lagoon. The diet of M. uropthalmus differed considerably between the two sites, with a more carnivorous diet (including high volumes of fish and decapods as well as plants) in San Pedrito Lagoon, and a diet of mostly detritus and plants in Polo Stream. The TFSI tests revealed phylogenetic constraints on the consumption of aquatic insects (p < 0.01) and algae (p = 0.04).
Dietary niche breadths were widest for the Thorichthys species, C. robertsoni, and T. salvini, and these species consumed relatively even proportions of various invertebrate categories and detritus as well as small volumes of several other categories (Table 2). Both Vieja species (herbivore-detritivores) and P. splendida (piscivore) had much narrower dietary niches. For the Polo Stream cichlid assemblage, mean dietary niche overlap was 0.61, a value significantly greater (p = 0.001) than expected based on 1000 randomized simulations. Dietary overlaps in Polo Stream were higher than expected for all nearest neighbor ranks (Fig. 2). Mean dietary niche overlap in the San Pedrito Lagoon assemblage (0.34) was neither higher (p = 0.43) nor lower (p = 0.57) than expected based on randomized simulations. Across sites, the highest interspecific niche overlap was between the two Vieja species and among the two Thorichthys species, C. robertsoni, and T. salvini (Table 3). Petenia splendida had very low dietary overlap with all of the other cichlid species except for M. uropthalmus.

Mean observed trophic niche overlap for Polo Stream (black boxes) and San Pedrito Lagoon (grey boxes) along with mean overlaps from the ECOSIM randomization (open circles) by nearest neighbor rank for eight common cichlid species in the Pantanos de Centla Biosphere Reserve
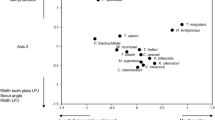
The first two axes from the PCA performed on morphological traits explained 59.9% and 16.0%, respectively, of variation among species (Fig. 3). The morphological gradient along the first axis was associated with differences in jaw protrusion, gape size, gill raker number, body depth, caudal peduncle width, anal fin length, pelvic fin length, dorsal fin height, and eye diameter. The second axis was most influenced by differences among species in head length, gut length, body size, eye position, and gill raker length. Petenia splendida had a high positive score on PC1 due to high jaw protrusion, large gape, relatively short fins, small eyes relative to body size, and few widely spaced gill rakers. Species with lowest scores on PC1 were characterized by having relatively long fins, deeper bodies, and more gill rakers. These included the Vieja and Thorichthys species. Thorichthys species, C. robertsoni, and P. splendida had high scores on PC 2, which was associated with a relatively longer head, high eye position, and short gut length. The two Vieja species, which had long guts and relatively large body sizes, had low PC2 scores.

PCA ordination of morphological traits defining the position of Lower Grijalva-Usumacinta cichlid species in two dimensions of trait space. Traits used to characterize the two axes had correlation coefficients ≥0.2. Abbreviations for morphological traits are given in Table 1
Canonical correspondence analysis revealed a significant relationship between diet and morphological traits (p < 0.01, 1000 permutations). The first two axes represented 51.1% and 29.7% of diet variation explained by morphology respectively (Fig. 4). The dominant axis (axis 1) separated the piscivorous P. splendida from all other species. Herbivore/detritivore species were separated from species with more invertebrates in their diets along the second axis. Gape width and extent of jaw protrusion were positively correlated with axis 1 and strongly associated with the proportion of fish in the diet. Gut length was associated with the consumption of algae, aquatic macrophytes, and detritus and was negatively correlated with axis 2. Head length, snout length, and eye position were positively correlated with axis 2 and with the frequency of mollusks, benthic meiofauna, and aquatic insect larvae in stomach contents. Based on the relative position of species in the CCA ordination, there were four main ecomorphological groups: piscivores with large gapes and highly protrusible jaws (P. splendida), benthic invertebrate feeders with relatively long heads and snouts and higher eye position (Thorichthys spp., C. robertsoni), herbivore/detritivores with long guts and shorter head lengths (Vieja spp.), and generalist feeders with intermediate-sized mouths, heads, and guts (M. urophthalmus and T. salvini).

Ordination of prey items and morphological traits of common Lower Grijalva-Usumacinta cichlid species on the first two axes of the CCA. Abbreviations for morphological traits are given in Table 1
Discussion
Twelve co-occurring native cichlid species were documented in sites within Pantanos de Centla Bioshpere Reserve, ranking these assemblages among the most species-rich in Mesoamerica (Myers 1966; Bussing 1985). High dietary niche overlap was observed among several cichlid species, particularly in Polo Stream, where vegetative detritus was a substantial component of the diet for all species. Relationships between feeding ecology and morphology identified by canonical correspondence were similar to those described for other Neotropical cichlids (e.g., Winemiller et al. 1995; Cochran-Biederman and Winemiller 2010; Montaña and Winemiller 2013). These strong, consistent ecomorphological relationships may be useful for predicting diet based on morphology for other cichlid assemblages that currently lack data.
Most cichlid species consumed a variety of prey items, including insect larvae, other benthic invertebrates, detritus, and plant material. Despite the high overlap, four basic feeding groups could be identified. Petenia splendida, a specialized piscivore, had a very narrow trophic niche distinct from all other species. Thorichthys helleri, T. pasionis, and C. robertsoni fed primarily on benthic invertebrates. Vieja bifasciata and V. melanura consumed larger proportions of detritus and aquatic macrophytes. Finally, T. salvini and M. urophthalmus had generalist diets, consuming many kinds of invertebrates as well as detritus, algae and plant material. This is consistent with findings for P. splendida and C. robertsoni in cichlid assemblages in Quintana Roo, México, (Valtierra-Vega and Schmitter-Soto 2000) and in the Bladen River, Belize (Cochran-Biederman and Winemiller 2010). Trichromis salvini appears to be more omnivorous in the lowland habitats in southern Mexico (this study; Hinojosa-Garro et al. 2013) compared to upland streams in Belize (Cochran-Biederman and Winemiller 2010). Similar to our findings for Thorichthys helleri and T. pasionis, other species in the genus Thorichthys have been found to feed predominantly on benthic invertebrates (Valtierra-Vega and Schmitter-Soto 2000; Cochran-Biederman and Winemiller 2010). Thorichthys species have been described as substrate winnowers or sifters that ingest sand or other loose sediments; ingested material is then manipulated within the orobranchial chamber in order to separate invertebrate prey from debris, the latter being expelled from the mouth or opercular opening (López-Fernández et al. 2014).
Similar spectra of trophic niches were described for a less diverse Mesoamerican cichlid assemblage (six species) in Belize (Cochran-Biederman and Winemiller 2010). In addition to piscivores, benthic invertebrate feeders, herbivore-detritivores, and generalists, Winemiller et al. (1995) found that a Costa Rican cichlid assemblage with high species richness (14 species) contained two additional, unique trophic specialists: algae scraper and frugivore. Some cichlid assemblages in South America contain more co-occurring species, but herbivorous and detritivorous cichlids are rarer in these systems (Winemiller et al. 1995). Myers (1966) suggested that Mesoamerican cichlid species have filled an exceptionally diverse set of ecological niches owing to the low diversity of ostariophysan fishes compared to other tropical regions. Compared to cichlids in South America, Mesoamerican cichlids, as a group, consume more detritus, algae and vegetation, perhaps due to less competition for those resources from characiform and siluriform fishes, both historically as well as presently (Winemiller et al. 1995).
Dietary niche overlap was higher than would be predicted based on random expectations in the Polo Stream cichlid assemblage, and did not differ from random expectations in the San Pedrito Lagoon assemblage. In both sites, dietary overlap was particularly high among benthic invertebrate feeders (Thorichthys species and C. robertsoni) and the herbivore-detritivore trophic group (Vieja species). High niche overlap among congeneric species (Thorichthys and Vieja species) suggests that phylogenetic niche conservatism maintains similar feeding ecology within clades, and our analyses revealed that consumption of some prey items had a strong phylogenetic signal. Niche overlaps higher than or similar to randomized simulations in these assemblages did not support expectations for trophic niche partitioning among co-occurring cichlids. This was somewhat surprising because sampling took place at the height of the dry season, the period during which low water levels are expected to constrain resources, resulting in more pronounced resource partitioning among fishes in tropical lotic systems (Lowe-McConnell 1987; Winemiller 1991b; Jepsen et al. 1997). A previous study in San Pedrito Lagoon showed that based on stable isotope signatures, fishes and other aquatic consumers used a narrower set of food resources during the dry season compared to the wet season when flooding provided access to a broader array of resources (Sepúlveda-Lozada et al. 2017). Our results suggest that although the diversity of available food resources may be lower in the dry season in this system, low-quality resources (detritus in Polo Stream and submerged vegetation in San Pedrito Lagoon) might remain plentiful enough during low-water conditions to preclude distinct niche partitioning within cichlid assemblages overall. Because many Mesoamerican cichlids are capable of foraging on detritus and aquatic vegetation, they likely overlap in the use of these resources when others are less available. To better understand the influence of resource availability in this system, the effects of seasonal environmental variation in the Grijalva-Usumacinta delta (Arévalo-Frías and Mendoza-Carranza 2015) on food resources and fish foraging need to be investigated.
It is also possible that food resource partitioning occurs at a scale of resolution finer than the one analyzed in this study. Subdividing prey items, such as insect larvae, into finer taxonomic categories might reveal stronger evidence of niche segregation. For example, Reinthal (1990) found that many co-occurring mbuna cichlids in Lake Malawi mostly consumed algae, and resource partitioning was detected only when algae were subdivided into different taxa. In our study, all cichlids were collected from similar macrohabitat types in both sites (Vallisneria beds in San Pedrito Lagoon and slow-moving, vegetated shorelines of Polo Stream), but potential differences in microhabitat use among species were not investigated.
Morphological differences among cichlid species in these assemblages were strongly associated with feeding (e.g., jaw protrusion, gape size, gill raker length and number, gut length) as well as locomotion and habitat use (body depth, fin lengths, caudal peduncle width). Our results further confirm strong associations between functional morphology and diet in cichlid assemblages (Liem 1973; Winemiller et al. 1995; Barlow 2000). In agreement with studies of cichlids in other regions, species with larger mouth gape and greater jaw protrusibility, such as P. splendida and M. urophthalmus, consumed more fish (Cochran-Biederman and Winemiller 2010; Montaña and Winemiller 2013). Functional morphology studies of jaw protrusion in P. splendida and other Neotropical cichlids suggest that increased protrusion is linked to improved performance in capturing relatively small and evasive prey, such as decapod crustaceans and small pelagic fish (Wainwright et al. 2001; Waltzek and Wainwright 2003; Hulsey and García de Leon 2005). Cichlids that were more herbivorous and detritivorous, including Vieja species, had longer guts and relatively short heads and snouts. Winemiller et al. (1995) also found an association between these traits and high proportions of detritus in diets of riverine cichlids across continents. Greater intestine length allows for longer processing times for relatively refractory material of lower nutritional quality, such as vegetative detritus (Gatz 1979; Bowen 1983). Our analyses also revealed that longer snout and head lengths were associated with diets consisting of benthic invertebrates. Winemiller et al. (1995) found an association between cichlid head length and consumption of mayfly nymphs, and snout length has been shown to correlate with benthic invertebrate feeding by South American cichlids (López-Fernández et al. 2012; Montaña and Winemiller 2013). Longer snouts may enable cichlids to sift more efficiently for benthic invertebrates in soft substrate (López-Fernández et al. 2014). Ecomorphological similarity between the Thorichthys species and C. robertsoni (sediment-sifting benthic invertebrate feeders with long heads and snouts) is considered a result of evolutionary convergence (Winemiller et al. 1995; Roe et al. 1997).
Because phylogenetic constraints may have a confounding effect in comparative studies, relationships between form and feeding performance are expected to be strongest when comparing species within the same family (Douglas and Matthews 1992). Elucidation of relationships is enhanced when assemblages contain species capable of using a broad array of resources (Hugueny and Pouilly 1999). In this study, we examined morphological traits using traditional morphometric methods rather than geometric morphometric methods (e.g., Costa and Cataudella 2007; Lombarte et al. 2012). Franssen et al. (2015) found that while both approaches identified relationships between morphology and habitat use for cyprinids, traditional morphometric traits were more strongly related to diet. Some of the strongest relationships between diet and morphology in this study involved traits such as intestine length, gape size, jaw protrusion, and gill raker length that are not captured using geometric morphometric methods. Traditional morphometrics often reveal relationships between morphology and feeding ecology in a directly interpretable manner. For example, traits such as gape size and jaw protrusion are important biomechanical components of feeding performance in cichlids (e.g. Hulsey and García de Leon 2005; Arbour and Lopez-Fernandez 2014). In general, the links between cichlid morphology and feeding ecology in these assemblages confirm relationships found in other regions (e.g.; Winemiller et al. 1995; López-Fernández et al. 2012; Montaña and Winemiller 2013). Discovery of consistent ecomorphological relationships provides a basis for inferring aspects of the ecology of poorly-studied taxa lacking data on resource use (Hugueny and Pouilly 1999; Winemiller et al. 2015).
Cichlids are major components of fisheries of the Grijalva-Usumacinta delta (Mendoza-Carranza et al. 2013), a region currently vulnerable to environmental impacts from the petroleum industry, agricultural practices, and non-native invasive species. Non-native cichlids, including Oreochromis species and Parachromis managuensis, are established in the lower Grijalva-Usumacinta (Amador-del Ángel and Wakida-Kusunoki 2014), and they may compete with native cichlids. Invasive loricariid catfish, Pterygoplichthys sp., now abundant in the region, have affected ecosystem processes (Capps and Flecker 2013) and already have been implicated in fisheries declines (Wakida-Kusunoki et al. 2007). Documenting the ecology of these diverse native cichlid assemblages provides important information for management and conservation in the Lower Grijalva-Usumacinta. Better understanding of how resources that support cichlid stocks and species coexistence, for example, is critical for constructing models to estimate the impacts of environmental change in the region and to guide effective ecosystem-based management of fisheries resources.
References
Abouheif E (1999) A method for testing the assumption of phylogenetic independence in comparative data. Evol Ecol Res 1:895–909
Amador-del Ángel LE, Wakida-Kusunoki AT (2014) Peces invasores en el sureste de México. In: Mendoza R, Koleff P (eds) Especies acuáticas invasoras en México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, pp 425–433
Arbour JH, Lopez-Fernandez H (2014) Adaptive landscape and functional diversity of Neotropical cichlids: implications for the ecology and evolution of Cichlinae (Cichlidae; Cichliformes). J Evol Biol 27:2431–2442
Arévalo Frías WDC, Carranza M (2012) Larvas y juveniles de peces en ambientes estuarinos de la Reserva de la Biosfera Pantanos de Centla y su zona costera adyacente. In: Sánchez A, Chiappa-Carrara X, Pérez B (eds) Recursos Acuáticos Costeros del Sureste: Tendencias actuales en investigación y estado del arte, vol 2. RECORECOS, CONCYTEY, UNAC, AR, UJAT, ECOSUR, UNAM, México, pp 242–269
Arévalo-Frías W, Mendoza-Carranza M (2015) Influence of temporal and spatial factors on abundance and richness of fish early stages in shallow tropical estuaries. Environ Biol Fish 98:891–904
Barlow GW (2000) The Cichlid Fishes. Perseus Publishing, Cambridge
Bowen SH (1983) Detritivory in neotropical fish communities. Environ Biol Fish 9:137–144
Bussing WA (1985) Patterns of distribution of the Central American ichthyofauna. In: Stehli FG, Webb SD (eds) The great American biotic interchange. Plenum Publishing, New York, pp 453–473
Capps KA, Flecker AS (2013) Invasive aquarium fish transform ecosystem nutrient dynamics. Proc R Soc B 280:20131520
Cochran-Biederman JL, Winemiller KO (2010) Relationships among habitat, ecomorphology and diets of cichlids in the Bladen River, Belize. Environ Biol Fish 88:143–152
Costa C, Cataudella S (2007) Relationships between shape and trophic ecology of selected species of Sparids of the Caprolace coastal lagoon (Central Tyrrhenian Sea). Environ Biol Fish 78:115–123
Douglas ME, Matthews WJ (1992) Does morphology predict ecology? Hypothesis testing within a freshwater stream fish assemblage. Oikos 65:213–224
Entsminger, GL (2014) EcoSim Professional: Null modeling software for ecologists, Version 1. Acquired Intelligence Inc., Kesey-Bear, & Pinyon Publishing. Montrose, CO 81403. http://www.garyentsminger.com/ecosim/index.htm
Espinal JC, Salles P, Morán DK (2007) Storm surge and sediment process owing to hurricane Isidore in Terminos Lagoon, Campeche. In: Kraus NC, Rosati JD (eds) Coastal sediments’07: proceedings of the Sixth International Symposium on Coastal Engineering and Science of Coastal Sediment Processes, May 13–17, 2007. ASCE Publications, New Orleans, pp 996–1007
Franssen NR, Goodchild CG, Shepard DB (2015) Morphology predicting ecology: incorporating new methodological and analytical approaches. Environ Biol Fish 98:713–724
Gatz AJ (1979) Ecological morphology of freshwater stream fishes. Tulane Stud Zool Bot 21:91–124
Genner MJ, Turner GF, Barker S, Hawkins SJ (1999a) Niche segregation among Lake Malawi cichlid fishes? Evidence from stable isotope signatures. Ecol Lett 2:185–190
Genner MJ, Turner GF, Hawkins SJ (1999b) Foraging of rocky habitat cichlid fishes in Lake Malawi: coexistence through niche partitioning? Oecologia 121:283–292
Grossman GD, Ratajczak RE, Crawford M, Freeman MC (1998) Assemblage organization in stream fishes: Effects of environmental variation and interspecific interactions. Ecol Monogr 68:395–420
Herder F, Freyhof J (2006) Resource partitioning in a tropical stream fish assemblage. J Fish Biol 69:571–589
Hinojosa-Garro D, Arceo-Gomez J, Zambrano L, Escalera-Vazquez LH (2013) Fish diet composition in permanent and semi-permanent pools in tropical wetlands of the Yucatan Peninsula. Neotrop Ichthyol 11:881–890
Hugueny B, Pouilly M (1999) Morphological correlates of diet in an assemblage of West African freshwater fishes. J Fish Biol 54:1310–1325
Hulsey CD, García de Leon FJ (2005) Cichlid jaw mechanics: linking morphology to feeding specialization. Funct Ecol 19:487–494
Hulsey CD, García de Leon FJ, Johnson YS, Hendrickson DA, Near TJ (2004) Temporal diversification of Mesoamerican cichlid fishes across a major biogeographic boundary. Mol Phylogenet Evol 31:754–764
Jepsen DB, Winemiller KO, Taphorn DC (1997) Temporal patterns or resource partitioning among Cichla species in a Venezuelan blackwater river. J Fish Biol 51:1085–1108
Levins R (1968) Evolution in changing environments. Princeton University Press, Princeton
Liem KF (1973) Evolutionary strategies and morphological innovations: cichlid pharyngeal jaws. Syst Zool 22:425–441
Lombarte A, Gordoa A, Whitfield AK, James NC, Tuset VM (2012) Ecomorphological analysis as a complementary tool to detect changes in fish communities following major perturbations in two South African estuarine systems. Environ Biol Fish 94:601–614
López-Fernández H, Arbour JH, Willis S, Watkins C, Honeycutt RL, Winemiller KO (2014) Morphology and efficiency of a specialized foraging behavior, sediment sifting, in Neotropical cichlid fishes. PLoS One 9(3):e89832. https://doi.org/10.1371/journal.pone.0089832
López-Fernández H, Arbour JH, Winemiller KO, Honeycutt RL (2013) Testing for ancient adaptive radiations in neotropical cichlid fishes. Evolution 67:1321–1337
López-Fernández H, Winemiller KO, Montaña C, Honeycutt RL (2012) Diet-morphology correlations in the radiation of South American Geophagine Cichlids (Perciformes: Cichlidae: Cichlinae). PLoS One 7(4):e33997. https://doi.org/10.1371/journal.pone.0033997
Lowe-McConnell RH (1987) Ecological studies in tropical fish communities. Cambridge University Press, Cambridge
Lowe-McConnell RH (1991) Ecology of cichlids in South American and African waters, excluding the African great lakes. In: MHA K (ed) Cichlid fishes: behavior, ecology and evolution. Chapman and Hall, New York, pp 129–150
Martin AP, Bermingham E (1998) Systematics and evolution of lower Central American cichlids inferred from analysis of cytochrome b gene sequences. Mol Phylogenet Evol 9:192–203
McNyset KM (2009) Ecological niche conservatism in North American freshwater fishes. Biol J Linn Soc 96:282–295
Mendoza-Carranza M, Arévalo-Frías W, Inda-Díaz E (2013) Common pool resources dilemmas in tropical inland small-scale fisheries. Ocean Coast Manag 82:119–126
Mendoza-Carranza M, Hoeinghaus DJ, Garcia AM, Romero-Rodriguez A (2010) Aquatic food webs in mangrove and seagrass habitats of Centla Wetland, a Biosphere Reserve in Southeastern Mexico. Neotrop Ichthyol 8:171–178
Miller RR (2005) Freshwater Fishes of Mexico. University of Chicago Press, Chicago
Montaña CG, Winemiller KO (2013) Evolutionary convergence in Neotropical cichlids and Nearctic centrarchids: evidence from morphology, diet, and stable isotope analysis. Biol J Linn Soc 109:146–164
Myers GS (1966) Derivation of the freshwater fish fauna of Central America. Copeia 1966:766–773
Pavoine S, Ollier S, Pontier D, Chessel D (2008) Testing for phylogenetic signal in phenotypic traits: New matrices of phylogenetic proximities. Theor Popul Biol 73:79–91
Pease AA, González-Díaz AA, Rodiles-Hernandez R, Winemiller KO (2012) Functional diversity and trait-environment relationships of stream fish assemblages in a large tropical catchment. Freshw Biol 57:1060–1075
Pianka ER (1973) The structure of lizard communities. Annu Rev Ecol Syst 4:53–74
R Foundation for Statistical Computing (2010) R version 2.11.1. R Foundation for Statistical Computing, Vienna
Reinthal PN (1990) The feeding habits of a group of herbivorous rock-dwelling cichlid fishes (Cichlidae: Perciformes) from Lake Malawi, Africa. Environ Biol Fish 27:215–233
Rican O, Pialek L, Dragova K, Novak J (2016) Diversity and evolution of the Middle American cichlid fishes (Teleostei: Cichlidae) with revised classification. Vertebr Zool 66:3–102
Rodiles-Hernández R, Diaz-Pardo E, Lyons J (1999) Patterns in the species diversity and composition of the fish community of the Lacanja River, Chiapas, Mexico. J Freshw Ecol 14:455–468
Roe KJ, Conkel D, Lydeard C (1997) Molecular systematics of middle American cichlid fishes and the evolution of trophic-types in 'Cichlasoma (Amphilophus)' and 'C-(Thorichthys)'. Mol Phylogenet Evol 7:366–376
Ross ST (1986) Resource partitioning in fish assemblages: a review of field studies. Copeia 1986:352–388
Schluter D (2000) The ecology of adaptive radiation. Oxford Univ. Press, New York
Sepúlveda-Lozada A, Mendoza-Carranza M, Wolff M, Saint-Paul U, Ponce-Mendoza A (2015) Differences in food web structure of mangroves and freshwater marshes: evidence from stable isotope studies in the Southern Gulf of Mexico. Wetl Ecol Manag 23:293–314
Sepúlveda-Lozada A, Saint-Paul U, Mendoza-Carranza M, Wolff M, Yáñez-Arancibia A (2017) Flood pulse induced changes in isotopic niche and resource utilization of consumers in a Mexican floodplain system. Aquat Sci 79:597–616
Servicio Meteorológico Nacional (2017) Resumenes mensuales de temperature y lluvia. Available at: http://smn.cna.gob.mx/es/climatologia/temperaturas-y-lluvias/resumenes-mensuales-de-temperaturas-y-lluvias. Accessed Aug 2017
Soria-Barreto M, Rodiles-Hernández R (2008) Spatial distribution of cichlids in Tzendales River, Biosphere Reserve Montes Azules, Chiapas, Mexico. Environ Biol Fish 83:459–469
Valtierra-Vega MT, Schmitter-Soto JJ (2000) Hábitos alimentarios de las mojarras (Perciformes: Cichlidae) de la laguna Caobas, Quintana Roo, México. Rev Biol Trop 48:503–508
Wainwright PC, Ferry-Graham LA, Waltzek TB, Carroll AM, Hulsey CD, Grubich JR (2001) Evaluating the use of ram and suction during prey capture by cichlid fishes. J Exp Biol 204:3039–3051
Wakida-Kusunoki AT, Ruis-Carus R, Amador del Angel E (2007) Amazon sailfin catfish, Pterygoplichthys pardalis (Loricariidae), another exotic species established in southeastern Mexico. Southwest Nat 52:141–144
Waltzek TB, Wainwright PC (2003) Functional morphology of extreme jaw protrusion in Neotropical cichlids. J Morphol 257:96–106
Winemiller KO (1990) Spatial and temporal variation in tropical fish trophic networks. Ecol Monogr 60:331–367
Winemiller KO (1991a) Ecomorphological diversification in lowland freshwater fish assemblages from five biotic regions. Ecol Monogr 61:343–365
Winemiller KO (1991b) Comparative ecology of Serranochromis species (Teleostei: Cichlidae) in the Upper Zambezi River floodplain. J Fish Biol 39:617–639
Winemiller KO, Fitzgerald DR, Bower LM, Pianka ER (2015) Functional traits, convergent evolution, and periodic tables of niches. Ecol Lett 18:737–751
Winemiller KO, Kelso-Winemiller LC (2003) Food habits of tilapiine cichlids of the Upper Zambezi River and floodplain during the descending phase of the hydrologic cycle. J Fish Biol 63:120–128
Winemiller KO, Kelso-Winemiller LC, Brenkert AL (1995) Ecomorphological diversification and convergence in fluvial cichlid fishes. Environ Biol Fish 44:235–261
Winemiller KO, Pianka ER (1990) Organization in natural assemblages of desert lizards and tropical fishes. Ecol Monogr 60:27–55
Yáñez-Arancibia A, Day JW, Currie-Alder B (2009) Functioning of the Grijalva–Usumacinta river delta, Mexico: challenges for coastal management. Ocean Yearbook. The University of Chicago Press, Chicago, pp 479–507
Acknowledgements
Funding was provided to MMC by Consejo Nacional de Ciencia y Tecnología and to KOW by the National Science Foundation (DEB 0516831). Hernán López-Fernández, Timothy Grabowski, and Matthew Barnes provided helpful comments on an earlier version of this manuscript. We thank Benjamin “Mincho” Martinez, Gustavo Salvador, Alfredo Hernández, and Candelario Jimenez for help with field fish collection and the directors of Centla Wetland Biosphere Reserve for access to facilities during this research. Fish collections were carried out following a protocol approved by the Texas A&M University Institutional Animal Care and Use Committee (AUP 2005-117).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Pease, A.A., Mendoza-Carranza, M. & Winemiller, K.O. Feeding ecology and ecomorphology of cichlid assemblages in a large Mesoamerican river delta. Environ Biol Fish 101, 867–879 (2018). https://doi.org/10.1007/s10641-018-0743-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-018-0743-1