
Abstract
Isotopic enrichment offers structural insights that are not easily accessible with natural abundance isotopic composition. Deuterated cellulose has attracted considerable attention in the field of neutron scattering studies, providing information about the dynamics, structure of cellulose and its interactions with other plant cell wall components. The deuteration of cellulose also allows the analysis of cellulose hydrogen bonds by FTIR or NMR techniques. The chemical structure of cellulose contains both exchangeable hydroxyl and non-exchangeable alkyl hydrogens. Deuterium incorporation can be divided into two classifications: biological route which incorporates both alkyl and hydroxyl bound deuterium, and chemical route which typically replaces hydroxyl-bound exchangeable hydrogen. The biological route involves cultivating plants or microorganisms in a deuterium-enriched medium. The chemical route typically involves an exchange reaction between hydroxyl-bound hydrogen and D2O, often facilitating with an alkaline reagent. This review provides an overview of recent advances in deuteration methods and characterization as well as the application of deuterated cellulose.
Graphical Abstract

Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cellulose, one of the most abundant organic compound on earth, is a linear glucose polymer linked by β-1,4 glycosidic bonds (Fig. 1) (French 2017). The degree of polymerization depends on cellulose's origin and varies in the range of ~ 1000 to 10,000 in the herbaceous and woody plants. The linear cellulose chain forms hydrogen bonds through its hydroxyl groups with adjacent molecules, promoting association into a crystalline fibrous type of structure (Zhao et al. 2019). In addition to its traditional use in the manufacture of paper and textiles, and other products such as cellulose nitrate, acetate and mixed esters (Klemm et al. 2005), it has been extensively studied as a bioresource to replace petroleum-derived fuels, chemicals, and materials (Jiang et al. 2018).

Cellulose chemical structure
Cellulose has been subjected to a wide range of chemical modifications to functionalize its hydroxyl groups. Among all the cellulose modifications, cellulose deuteration, which refers to substitution of the 1H hydrogen atoms with the heavier isotope 2H (deuterium; D) atoms is an interesting technology. The obtained deuterium-incorporated cellulose has become one of the most insightful modifications for the structural study of cellulosic materials (Raghuwanshi et al. 2016). The zero-point-energy for a bond to deuterium is about 1.2–1.5 kcal/mol lower than that of a bond to hydrogen (Wiberg 1955). As a result, bonds to deuterium are shorter and stronger than corresponding bonds to hydrogen, slowing reaction rates for reactions that break hydrogen bonds (the kinetic isotope effect) but enabling resolution of deuterium-substituted positions by spectroscopic methods. For example, C-D and C-H bond dissociation energies are 341.4 and 338 kJ/mol, respectively (Rozenberg 1996). The dissociation constant for H2O, 1.00 × 10–14, is five-fold greater than that of D2O, 1.95 × 10–15 (Kirshenbaum 1951). Moreover, there are also isotope dilution effects for D bonds and H bonds for cellulose coupled vibrations (Nishiyama 2018).
Methods for the incorporation of deuterium into cellulose continue to be improved. Deuteration of cellulose has been produced mainly via two methodologies: (i) biological deuteration by cultivating plants (Evans and Shah 2015) or microorganisms in deuterated medium (Bali et al. 2013; O'Neill et al. 2015), which results in the substitution of hydrogens bonded to carbon, termed non-exchangeable hydrogens (i.e., alkyl hydrogens), as well as hydroxyl hydrogens that are not surface accessible, (ii) chemical deuteration by exchanging hydroxyl hydrogen atoms of cellulose with deuterium from solvent, typically D2O, often assisted by a chemical and/or thermal treatment (Bonhoeffer 1934; Lindh and Salmén 2017). Biological deuteration of cellulose is primarily used to examine the original cellulose structures and dynamics and to investigate biological and chemical reaction pathways by isotopic tracing and contrast. In general, the chemical exchange of hydroxyl hydrogen for deuterium is more commonly used for structural investigations than biological production. Exchange using D2O can be readily applied to many types of commercially relevant cellulose samples to probe hydrogen bonding patterns and crystallinity. In fact, both biological and chemical methods for deuteration of cellulose have had a very profound impact both on fundamental research on cellulosic structure and metabolism and on practical applications such as anti-counterfeiting materials (Song et al. 2021).
Deuterated cellulose is particularly useful for structural characterization in neutron scattering techniques because deuterium and hydrogen interact with the neutrons quite differently (Okuda et al. 2021). Scattering techniques using neutrons are non-destructive methods capable of providing critical insights into biomolecule structural properties over multiple length scales. Deuterium labeling is utilized to manipulate the scattering length densities of the biomolecules, enabling contrast variation and reduction in scattering background to facilitate the analysis of the molecular structure and dynamics of biological systems (O'Neill et al. 2015; Evans et al. 2019b; Bhagia et al. 2018).
Inclusion of the heavier deuterium atoms also enables studies of cellulose dynamics with vibrational spectroscopy (particularly low-frequency motions) and solid-state NMR spectroscopy (SSNMR). SSNMR has been used to characterize the structure and dynamics of the polysaccharides in the plant cell walls, and Gelenter et al. showed that the D (2H) and 13C labeled bacterial cellulose represented an excellent model system to study the polysaccharide motions and water accessibilities in the plant cell wall using the D-13C correlation NMR (Gelenter et al. 2017). Deuterium NMR spectroscopy offers local information about the environment around the nuclei via the nuclear quadrupolar interaction. It has been reported that the signal-to-noise ratio is improved upon deuteration by suppressing the spin diffusion and decreasing the relaxation rates of 13C or 15 N spins (Zhang 2020). Furthermore, deuterium labeling of otherwise chemically similar components (i.e., C–H vs. C–D) helped the assignment of specific spectroscopic peaks and investigation of phase separation due to the non-overlapping IR stretching vibrations (Russell et al. 2015). For example, both the C–H (2924–2954 cm−1) and O–H (3271–3480 cm−1) wavenumber on FTIR spectra are lowered in their corresponding C-D (2139–2251 cm−1) and O–D (2495 cm−1) wavenumber (Song et al. 2020). Moreover, after all of the cellulose hydroxyl groups are deuterated, the amorphous regions’ hydroxyl groups could be rehydrated under certain conditions and then analyzed by FTIR and NMR techniques; the cellulose hydroxyl groups accessibility and related reaction mechanism during various treatment were thus able to be studied (Fackler and Schwanninger 2011). For the purposes of this review paper, and due to its widespread use in earlier literature, the term “amorphous” will be used to denote the regions of cellulose that are accessible to water exchange.
Deuterated cellulose that is exchange-resistant to a particular environment could be developed as a novel cellulose-modified material since many properties such as thermostability, hydrolysis rate, and detectability were changed due to the isotope effect (Song et al. 2021). A previous review by Reishofer and Spirk summarizes achievements in the understanding of cellulose accessibility, structure, and function with a particular focus on its interactions with deuteration (Reishofer and Spirk 2016). In this review, we focus on recent advances in different chemical and biological routes of deuterium incorporation into cellulosic material and discuss the characterization of different deuterated cellulose preparations. In specific, the effects of deuterium incorporation on cellulose chemistry and physical structure are discussed. The scientific application of deuterated cellulose for investigation of its structure and dynamics as well as interaction with other polymers including cellulase proteins and polysaccharides (i.e., xylan and other hemicelluloses) using neutron scattering, FTIR, and NMR techniques from both fundamental and practical perspectives is also reviewed.
Biological routes of deuterium incorporation into cellulose
Biological routes of deuterium incorporation into cellulose include the growth of plants in D2O and the cultivation of cellulose-producing bacterial strains in deuterium-enriched medium. The deuterated cellulose or enriched cellulose is then obtained from deuterated plants or microorganisms.
Growth of plants in D 2 O
Studies of the effects of D2O on germination and growth of tobacco plants were carried out by Gilbert Lewis at the University of California soon after the discovery of deuterium by Harold Urey (Lewis 1933, 1934). These early studies demonstrated that some plants could germinate and grow in 50% D2O, while germination, rooting, and growth, were completely inhibited by 100% D2O. In contrast, some species of algae can adapt to growth in 90–100% D2O (Katz and Crespi 1966). The incorporation of deuterium into photosynthetic biomass was first demonstrated in 1934 by Reitz and Bonhoeffer, who cultivated Chlamydomonas algae in water with 12.3% D-content, finding that both exchangeable (hydroxyl) and non-exchangeable (alkyl) deuterium were present in the dried algal biomass (Reitz and Bonhoeffer 1934). Cultivation of Scenedesmus algae in 47% D2O was then shown to produce biomass with 31% deuterium incorporation (Reitz and Bonhoeffer 1935). Algae continue to be cultivated in 90–100% D2O as a source of deuterated biochemicals, including the deuterated glucose used in bacterial media (Crespi et al. 1960; Daboll et al. 1962; Crespi et al. 1970; Crespi & Katz 1972; Zachleder et al. 2018).
The effect of D2O on the growth properties of wheat (Pratt and Curry 1937), winter rye (Siegel et al. 1964; Waber and Sakai 1974), Arabidopsis (Xiaoyuan et al. 2010), barley (Zachleder et al. 2018) and duckweed (Cope et al. 1965) have been explored. Researchers found that seed germination, root growth, and flowering of these higher plants were severely impacted by D2O concentrations higher than 50%. Despite these issues, several species of higher plants have been cultivated in different D2O concentrations to obtain biomass with lower levels of deuterium incorporation that are sufficient for some purposes (Blake et al. 1967; Katz and Crespi 1966). For example, partially deuterium-labeled carrots, kale, and spinach were cultivated in 15–30% D2O for nutritional studies (Grusak 1997; Tang et al. 2005).
The highest deuterium incorporation was reported for the duckweed Lemna peruspilla grown photoheterotrophically, with 35% deuterium content of whole biomass obtained by growth with 0.5% glucose in 50% D2O and 59% for growth with 0.5% deuterated glucose in 50% D2O (Cope et al. 1965). Deuterium incorporation into stem biomass from kale (collards; Brassica oleracea L., subsp. acephala; var. viridis) cultivated in 30% D2O was determined by NMR and found to be ~ 31% deuterium incorporation in the cellulose (Foston et al. 2012). Production of cellulose with higher deuterium content was also reported from duckweed (Lemna minor and gibba) (Evans et al. 2019a), annual ryegrass (Lolium multiflorum) (Evans et al. 2014), and switchgrass (Panicum virgatum) (Evans et al. 2015) that were cultivated in 50% D2O solutions. The D2O concentration employed for plant cultivation and the resultant degree of deuterium incorporation into the plants available from recent literature are summarized in Table 1, in which the highest deuteration incorporation rate was 60%. Obtaining higher deuterium incorporation in plants is typically difficult as higher levels of D2O adversely affect the growth of plants. However, since plant cellulose is synthesized as a cell wall component in synchrony and in the presence of other cell wall components, growth of deuterated plants can be worth the effort to enable observation of structural features. Since seed germination and primary root extension are particularly impacted by high content D2O, cultivation of species that propagate by vegetative growth (i.e., switchgrass and duckweed), or establishment of plants in H2O before transferring to deuteration media generally produce better results (Evans and Shah 2015).
The cellulose obtained from the deuterated plants showed in molecular weight compared to control protiated plants that varied depending on the plants. For example, cellulose isolated from deuterated switchgrass had various changes in structural properties polydispersity index (DPI) and molecular weight profiles compared to control protio cellulose samples (Evans et al. 2015). The degree of polymerization (DPw) of switchgrass cellulose was increased 22% by deuteration, but the crystallinity was not statistically different with CrI of 42 ± 1.6% compared to 44 ± 1.6% for protiated (Evans et al. 2015). However, in a study of the duckweeds Lemna gibba and Lemna minor (Evans et al. 2019a), the partially deuterated cellulose DPw values were lower than those of the H2O grown controls, 63% of control for L. gibba and 85% of control for L. minor.
Bhagia et al. showed that the enzymatic hydrolysis of deuterated cellulose and holocellulose isolated from switchgrass was slower by 18 and 8%, respectively, than that of the corresponding protiated cellulosic substrates (Bhagia et al. 2018). A similar effect was observed for enzymatic hydrolysis of deuterated and protiated bacterial cellulose, which gave ~ 90 and 95% of the expected glucose yield, respectively (He et al. 2014). These lower glucose yields from deuterated cellulose versus protiated cellulose for the enzyme-catalyzed hydrolysis of their ß-1,4 glycosidic bonds fall within the range reported for kinetic isotope effects observed for enzymatic hydrolysis of aryl glucosides deuterated at C1 (Li et al. 2001).
Cultivation of microorganisms in deuterium-enriched medium
There have been methods reported for the production of deuterated biopolymers including polyhydroxyalkanoate, chitosan and cellulose, which provide not only multiple options for creating contrast in polymer blends and composites for structural studies but also insight into the biosynthetic pathways (Russell et al. 2015). Cell growth is inhibited in D2O for both prokaryotes and eukaryotes, although the tolerance to D2O is higher in prokaryotes compared to eukaryotes and especially higher order multicellular eukaryotes (Katz and Crespi 1966; O'Neill et al. 2015). Gradual adaptation with growth in increasing D2O concentrations has been successful with several prokaryotes and unicellular eukaryotes (Katz and Crespi 1966; O'Neill et al. 2015; Russell et al. 2015). The acetic acid bacteria, now assigned to the genus Komagataeibacter (synonym Gluconacetobacter,; basonym Acetobacter) are gram-negative bacteria that, in addition to acetic acid, produce large amounts of extracellular cellulose as shown in Fig. 2 (Brown 1886; Klemm et al. 2005). Microscopic observations revealed that the cellulose microfibrils are extruded through the cell membranes and cell walls directly into the growth media (Brown et al. 1976; Kondo et al. 2002). The deuterated medium and degree of deuterium incorporation for bacterial cellulose are summarized in Table 2.

Copyright Elsevier)
SEM image of protiated (hydrogenated) bacterial cellulose (HBC, left) and deuterated bacterial cellulose (DBC, right). Scale bars 1 μm. (Reprinted from Bali et al. (2013) The effect of deuteration on the structure of bacterial cellulose. Carbohydrate Research 374:82–88 with permission.
The scientific names of these bacteria have changed several times, so a brief explanation will be useful for understanding both current and previous literature. Originally named Bacillus xylinum (Brown 1886), they were later assigned to the genus Acetobacter, with two subgenera, Acetobacter and Gluconoacetobacter, in 1984 (Yamada and Yukphan 2008). The original species designation xylinum was later changed to xylinus to conform to Latin grammatical usage (Yamada et al. 1997). The change to Komagatabacter (Yamada et al. 2011) was later revised to Komagataeibacter (Yamada et al. 2012). The American Type Culture Collection (Manassas, Virginia, USA) currently uses Komagataeibacter xylinus as the scientific name for ATCC 53524 and 700178 (Table 2) (2021).
It is well known that bacterial cellulose differs in its physical properties from plant cellulose. For example, it has higher water retention capacity, tensile strength, and Young’s modulus than plant cellulose (Klemm et al. 2005; O'Neill et al. 2015). Incorporation of deuterium into bacterial cellulose is carried out by the cultivation of bacteria in deuteration media which can contain varying amounts of D2O, deuterated glycerol, or deuterated glucose, depending on the desired incorporation level in the cellulose (O'Neill et al. 2015). Bali et al. reported the production of deuterated bacterial cellulose with an 85% deuterium incorporation by cultivating the bacteria in 100% deuterated glycerol in the D2O medium (Bali et al. 2013). A bacterial cellulose-producing strain developed for efficient glucose utilization was studied in deuterated glucose-based media (Martínez-Sanz et al. 2016b; Martínez-Sanz et al. 2017c). Both D2O and deuterated carbon sources such as deuterated glucose and glycerol can provide a deuterium source for bacterial cellulose. Hence, deuterated bacterial cellulose's deuterium substitution rate could be higher than 90% (He et al. 2014). Studies have demonstrated that there were no significant differences in the molecular and morphological properties between the deuterated and protiated bacterial cellulose despite the relatively high level of deuterium incorporation. Figure 2 shows the scanning electron microscopy (SEM) images of purified, freeze-dried protiated (hydrogenated) and deuterated bacterial cellulose (Bali et al. 2013). On the other hand, the polydispersity index of molecular weight was higher for deuterated than protiated bacterial cellulose, which appeared to indicate the presence of higher molecular weight cellulose chains in the deuterated sample (O'Neill et al. 2015).
Bali and coworkers also found that almost all hydrogens were replaced by deuterium (including C–D and R–O–D signals) except for partial replacement at carbon C6 in the deuterated bacterial cellulose (Bali et al. 2013). Figure 3 shows the deuteration arrangement on each anhydroglucose unit for different levels of bacterial cellulose biodeuteration (Su et al. 2016). It can be seen that the hydroxyl groups were deuterated preferentially than alkyl hydrogen once they are accessible to the solvent (e.g., D2O). The hydrogens bound to oxygen atom may exchange with the deuterium (e.g., C6H7D3O5 vs. C6H4D6O5 in Fig. 3), while those linked to a carbon atom cannot exchange. The location of deuterium in the glucose subunits and the degree of deuteration of bacterial cellulose can be controlled by varying the deuterated component composition of the medium (O'Neill et al. 2015). For example, glucose with specific deuterium labeling and no other source of deuterium enables labeling of specific positions within the glucose unit. The selectively deuterated bacterial cellulose could thus be used to understand the cellulose structure mechanism, such as the nature of phase transitions within the cellulose crystallite (Russell et al. 2015). However, bacterial metabolic pathways can cause positional-specific replacement of deuterium from deuterated glucose or glycerol by hydrogen from H2O in the growth media. Analysis by NMR of cellulose produced by growth on deuterated glucose-d7 in H2O and in D2O found that bacterial metabolic pathways resulted in replacement by hydrogen from water of 40% of the deuterium from C2, C3, C4, and C5 of deuterated glucose, while only 10% was lost from C1 and C6 (Barnoud et al. 1971). Use of the mutant strain ATCC 53,524 which was selected for increased cellulose yield through knockout of competing gluconate and keto-gluconate production (Johnson and Neogi 1989) minimized this effect according to characterization of deuterated cellulose produced with this strain (Russell et al. 2015; Martínez-Sanz et al. 2015a, 2016b, 2017a). Deuterated bacterial cellulose has also been used to reveal water-cellulose dynamics (e.g., water diffusion on cellulose) at the atomic scale by neutron scattering assisted by molecular dynamics simulation (O'Neill et al. 2017).

Copyright Springer Nature 2016)
Molecular structure of cellulose at different deuteration levels. (Reprinted with permission from Su et al. (2016) Smooth deuterated cellulose films for the visualisation of adsorbed bio-macromolecules Scientific reports 6:36,119.
Chemical routes of deuterium incorporation into cellulose
The chemical isotope exchange can be readily utilized to produce cellulose containing exchangable deuterium for applications that solely rely on deuterated hydroxyl groups (Tarmian et al. 2017). The isotope exchange of hydroxyl hydrogen for deuterium by soaking cellulose in D2O and alkaline reagents or exposure to D2O vapor is commonly used for structural investigations such as hydrogen bonding, accessibility and crystallinity (Hishikawa et al. 2017). Without the use of alkaline reagents, simply soaking cellulose in D2O without heating leaves crystalline regions relatively inaccessible to solvent, as shown in a study of the molecular structure of intact cotton fibers using SANS and SAXS (Martínez-Sanz et al. 2017b). Depending on the conditions used to treat the cellulose and to carry out the exchange reactions, the chemical exchange treatment could be classified into crystalline and amorphous deuteration categories by the extent and penetration of deuterium into the cellulose microfibrils, which enables tailoring the deuterium-substitution for specific investigations of surface and crystalline cellulose properties.
Crystalline OH group substitution by chemical exchange
All the RO-H groups from cellulose crystallite were reported to be able to exchange with RO-D groups through treatment in 0.1 N NaOD in D2O at 210 ℃ for 30 min to 1 h (Nishiyama et al. 2003), or 260 ℃ for 30 min to 1 h (Nishiyama et al. 2010). Under these severe conditions, the fully deuterated cellulose crystallite could be obtained without any loss of crystallinity, which provides an opportunity to locate the exact position of the H atom in cellulose based on the different scattering length of H and D for neutron study (Langan et al. 1999). Most notably, Nishiyama et al. successfully obtained different fully OH-deuterated cellulose crystallites including cellulose Iα (Nishiyama et al. 2003), Iβ (Nishiyama et al. 2002) and cellulose II (Nishiyama et al. 1999a, 2002), respectively. The crystal structure data as well as hydrogen-bonding system data were thus revealed by neutron studies (Nishiyama et al. 1999b). Neutron scattering analysis of the fully hydroxyl-deuterated cellulose was used to compare the different cellulose crystallites as well, such as the differences in diffraction pattern between cellulose I and cellulose II, and in the hydrogen-bonding system of cellulose I and cellulose III (Nishiyama et al. 2010). Langan and co-workers also developed a protocol to prepare deuterated ammonia-cellulose I and cellulose IIII (decomplexation of ammonia-cellulose I) sample using deuterated ammonia (ND3) and used these deuterated cellulose samples along with neutron and molecular dynamic simulation studies to reveal the crystal structure of cellulose IIII (Wada et al. 2004) and the rearrangement of hydrogen bonding in the complex of cellulose with ammonia (Wada et al. 2011). Figure 4 illustrates the neutron fiber diffraction patterns for different cellulose crystalline structures.

Copyright Elsevier)
Series of neutron fiber diffraction patterns for cellulose I and cellulose II with vertical fibre axes and displayed at the same scale in reciprocal space; a from a reconstituted sample of tunicin (cellulose Iβ) microcrystals; b as in (a), but after substituting all OHs by ODs; c from a reconstituted sample of Cladophora cellulose (cellulose Iα + Iβ); d as in (c) but after substituting all OHs by ODs; e from mercerized flax in standard NaOH/H2O; f as in (e), but mercerized in NaOD/D2O. (Printed with permission from Nishiyama et al. (1999b). High resolution neutron fibre diffraction data on hydrogenated and deuterated cellulose International journal of biological macromolecules 26:279–283.
Partial substitution of OH groups by chemical exchange
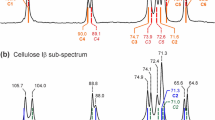
There are significant differences existing in the deuteration rates of the crystalline and amorphous regions of cellulose. Amorphous cellulose deuteration occurs quickly, followed by the slow exchange in crystalline region of cellulose. Meanwhile, the deuterated amorphous regions could be rehydrogenated once exposed to H2O (Reishofer and Spirk 2016). The partial deuterated crystalline area would retain the OD groups known as "resistant OD groups". Hishikawa et al. concluded that 10.6% of the total hydroxyl groups in nematically ordered cellulose II, which was produced by stretching cellulose swollen in lithium chloride-dimethylacetamide to align the fibrils before mercerization, were non-replaceable by D2O vapor-phase exchange, corresponding to the cellulose crystalline area (Hishikawa et al. 2010). With resistant hydroxyl groups in the crystalline area generated by alkaline reagents that did not exchange with vapor-phase D2O, data on specific hydrogen bonds in crystalline cellulose materials were thus readily generated (Hishikawa et al. 2017). Alkaline reagents like NaOH not only lowered the activation energy of R-O–H breaking but also swelled the internal cellulose structure. We have reported previously that prolonged soaking time, higher temperature and alkaline reagents could facilitate the deuteration of inaccessible cellulose crystalline area (Song et al. 2020). Our studies also suggested that a higher crystalline index contributed to a higher resistant deuterium incorporation. These findings are in agreement with previous reports that adjusting the treatment parameter could promote deuterium incorporation into cellulose (Frilette et al. 1948). The FTIR spectral data after deuterium incorporation are shown in Fig. 5. The hydrogen bonds around 3400 cm−1 are shifted to around 2500 cm−1, corresponding to deuterium bonds, which would help the calculation of resistant deuterium incorporation through the OD and OH characteristic intensity (Song et al. 2020, 2021). In addition, differences of the resistant deuterium incorporation position between cotton (natural cellulose, strongest peak at 2480 cm−1) and Tencel (regenerated cellulose, strongest peak at 2550 cm−1 and 2475 cm−1) have been observed.

Copyright Springer)
FTIR of the original and deuterated cellulose fibers (C: cotton; T: Tencel; Black solid line: original protonic fibers; Red dotted line: deuterated fibers). (Reprinted with permission from Song et al. (2020) The production of hydrogen–deuterium exchanged cellulose fibers with exchange-resistant deuterium incorporation Cellulose 27:6163–6174.
Characterization of deuterated cellulose
Characterization of biological route deuterated cellulose from plants
As mentioned aboved, He et al. also reported that enzymatic hydrolysis of deuterated bacterial cellulose and protiated cellulose yield ~ 90 and 95% of the expected glucose yield, respectively (He et al. 2014). The lower glucose yield from deuterated cellulose versus protiated cellulose is attributed to the secondary kinetic isotope effects on the enzyme-catalyzed hydrolysis of cellulose to cellobiose and finally glucose. These kinetic isotope effects for 35–85% deuterated cellulose fall within the range reported for kinetic isotope effects observed for enzymatic hydrolysis of aryl glucosides deuterated at C1 (Li et al. 2001).
Incorporation of deuterium into plant cellulose can extend the information obtained by analytical techniques. Small-angle neutron scattering (SANS) has been shown to be a useful technique for examination of the molecular structure of cellulose and lignocellulosic biomass (Pingali et al. 2010a; Martínez-Sanz et al. 2015b, 2017b), and the effects of thermochemical pretreatment on them (Pingali et al. 2010b, 2017; Pingali et al. 2020). Substitution of deuterium for hydrogen in polymers increases the scattering length density (SLD) observed with SANS as demonstrated for cellulose and other polymers (Martínez-Sanz et al. 2016a). The ability to differentiate the neutron scattering pattern of deuterated cellulose from those of other component polymers with lower deuterium content provides new capabilities to understand both structure and deconstruction mechanisms of plant biomass. This property can be leveraged for experimental investigation without the necessity to isolate the deuterated cellulose. SANS was used to compare the nanoscale structural features of biomass in switchgrass biomass with different levels of deuterium incorporation (Evans et al. 2019b). An example of a plot used to calculate SLD for partially deuterated and control switchgrass samples is provided in Fig. 6 (Evans et al. 2019b). This includes labile exchange that occurs between solvent and sample during the solvent contrast variation experiments. The plot shows the variation in scattering intensity of the SANS data (square-root of the scattering intensity) versus deuterium concentration in solvent for a fixed value of Q (= 0.0045 Å−1) for contrast series of four biomass samples from switchgrass grown in different D2O concentrations. The SLD of the D2O concentration at the intersection of the straight line fit with the y = 0 line is the SLD of that system (y = 0 represents the parabola vertex of a I(Q) vs D2O% profile). The results indicate that both field-grown and hydroponic plants grown with H2O have a lower SLD than the partially deuterated switchgrass.

Copyright 2019 American Chemical Society)
Plots of the square root of scattering intensity at Q = 0.0045 Å−1 as a function of D2O: H2O solvent mixture are used to determine the contrast match points of control and partially deuterated switchgrass biomass. The line is a straight-line fit of the parabolic I(Q0) vs. D2O% relation, therefore data points in the plot undergo a flip in their sign ‘ + ’ to ‘–’ (or vice versa) as the parabola vertex is crossed. The line at y = 0 represents the vertex of the parabola from the I vs. Q plot. The deuterated and hydrogenated samples were assigned opposite slopes and flipped as the vertex was crossed. The D2O: H2O solvent intercept (x-value) between y = 0 axis and the fit is the contrast-matching D2O: H2O solvent mixture. Three of the samples are hydroponically grown switchgrass in 100% H2O (red dots; red line), 50% D2O (blue filled up-triangle; blue line), 40% D2O at 30 °C (orange filled down-triangle; orange line), and the fourth is field-grown switchgrass in 100% H2O (green filled squares; green line). (Reprinted with permission from Evans et al. “Structural Studies of Deuterium-Labeled Switchgrass Biomass”, In Understanding Lignocellulose: Synergistic Computational and Analytic Methods, ACS Symposium Series 1338, Ed. M. D. Smith, Chapter 2, 17–32, ACS Publications, Washington D.C. (2019).
Figure 7 is an example of how the SANS data helped to reveal the internal biomass structure (Evans et al. 2019b). Based on our current understanding of the plant cell wall's structural characteristics, the three distinct Q-regions were related to the cellulose microfibril, disordered or amorphous plant polymers, and the plant cell wall surface, respectively. As indicated in Fig. 7, the switchgrass grown in 50% (blue filled up-triangle) and 40% D2O (orange filled down-triangle) both had a significantly higher contrast-matching D2O percentage (~ 75–85%) in the high-Q region known to correspond to cellulose microfibrils than H2O-grown hydroponic and field grown controls (~ 40%), suggesting that the cellulose had a significant D incorporation. A smaller clear increase was observed in the low-Q range corresponding to scattering by amorphous polymers even though the change is relatively small compared to the high-Q (Evans et al. 2019b). Therefore, it can be concluded that the substitution of hydrogen by deuterium in the deuterated biomass improves the ability to deconvolute the structural features of the individual biopolymer components in SANS experiments. Also evident from the plot is that the effect of this partial deuteration on SLD is the most pronounced for cellulose microfibrils, enabling deconvolution of cellulose scattering from that of amorphous polymers hemicellulose and lignin in whole biomass. This result is consistent with NMR studies of stem biomass of kale grown in 31% D2O indicating higher deuterium incorporation in cellulose than in lignin (Foston et al. 2012).

Copyright 2019 American Chemical Society)
The variation in contrast-matching D2O solvent mixture as a function of scattering-vector, Q. Three samples are switchgrass grown hydroponically in 100% H2O (red dots; red line), 50% D2O (blue filled up-triangle; blue line), 40% D2O at 30 °C (orange filled down-triangle; orange line), and the fourth is field-grown switchgrass grown with 100% H2O (green filled squares; green line). (Reprinted with permission from Evans et al. “Structural Studies of Deuterium-Labeled Switchgrass Biomass”, In Understanding Lignocellulose: Synergistic Computational and Analytic Methods, ACS Symposium Series 1338, Ed. M. D. Smith, Chapter 2, 17–32, ACS Publications, Washington D.C. (2019).
Characterization of biologically deuterated cellulose from bacteria
Deuterium labeling of bacterial cellulose was used to reveal cellulose's biosynthetic pathways (Barnoud et al. 1971; O'Neill et al. 2015). Raghuwanshi and coworkers used deuterated bacterial cellulose to investigate the mechanism of cellulose dissolution in ionic liquid (1-ethyl-3-methylimidazolium acetate) by applying small-angle neutron scattering (SANS) with contrast variation, and the results showed that besides disrupting the intermolecular hydrogen bonding, the ionic liquid also imparts an effective charge to the cellulose chains hindering their agglomeration in the solution (Raghuwanshi et al. 2018). Deuterated bacterial cellulose has also been used to form composites with hemicelluloses isolated from plants that mimic the plant cell wall structure (Martínez-Sanz et al. 2015a, 2017a; Shah et al. 2019). The partially deuterated bacterial cellulose hydrogels were synthesized in the presence of different plant cell wall polysaccharides (xylan, galactoglucomannan, glucomannan, or xyloglucan) to identify the different structural roles of these plant cell wall polysaccharides and their distinct interaction mechanisms with cellulose (Martínez-Sanz et al. 2016b, 2017a; Penttilä et al. 2018; Shah et al. 2019). The results suggest that cellulose microfibrils are composed of an impermeable crystalline core surrounded by a partially hydrated paracrystalline shell in their native state and cellulose ribbons consist of a network of cellulose microfibrils and tightly bound solvent. With deuterium labeling, the structural role of water in cellulose hydrogels can be explored and identified (O'Neill et al. 2017). Deuterated bacterial cellulose can be dissolved in appropriate solvents and reconstituted into thin films to examine the adsorption mechanisms of cellulose-binding materials with SANS and other techniques. Deuterated bacterial cellulose films were used for enhanced contrast in neutron reflectometry to enable the visualization of adsorbed proteins such as IgG antibodies (Raghuwanshi et al. 2017) and human carbonic anhydrase (Koruza et al. 2018).
Characterization of chemically deuterated cellulose
With the complete chemical deuteration of all cellulose OH groups, the cellulose crystallite structural details were revealed through scattering studies (Nishiyama et al. 2002, 2003). Furthermore, resistant deuterated cellulose has the potential to be applied as anti-counterfeiting materials such as specialty paper. Moreover, after all of the OH groups in cellulose crystallites were deuterated, deuteration and rehydrogenation could be combined to characterize cellulose accessibility since the amorphous regions would be easily re-exchanged with hydrogen under certain conditions. Horikawa et al. showed that deuterated cellulosic substrates could be rehydrogenated by simply soaking it in water at room temperature, in which case only the OD groups on the surface could be rehydrogenated. The complete deuterated cellulose treated by a rehydrogenation process was termed as "intra-crystalline" deuterated cellulose (Horikawa and Sugiyama 2008). Consequently, the ratio of OH/OD absorbance of "intra-crystalline" deuterated cellulose could be determined by FTIR. It could also be used to demonstrate the cellulose microfibril size and accessibility. Similarly, the localization of Iα and Iβ domains within a cellulose microfibril could be obtained via "intra-crystalline" deuteration and rehydrogenation of cellulose samples; and it was suggested that the simple “skin–core” distribution model of Iα and Iβ domains is not realistic, at least, for these native celluloses of Iα-rich algae (Horikawa and Sugiyama 2009). Moreover, the partially deuterated amorphous cellulose made through deuteration/rehydration treatment was utilized to investigate the reaction mechanism of a microwave-assisted low-temperature decomposition process to produce high-quality fuels from biomass based on the difference of the calorific value of char obtained from cellulose processed conventionally and in the presence of microwave as well as deuterated cellulose (Budarin et al. 2010).
As mentioned above, the amorphous cellulose regions’ deuteration is exchangeable and reversible because rehydrogenation occurs readily in the surface and noncrystalline region. The substitution of deuterium in amorphous regions demonstrated that the cellulosic materials' amorphous area consists of at least three types of domains at the IR level, supported by different deuteration rates along with treatment time (Hishikawa et al. 2005). The crystallinity and moisture content of the cellulose materials could also be calculated based on the deuterium accessibility results (Lee and Bismarck 2011). During the hydrogen–deuterium exchange process in the cellulose amorphous area, the hydration behavior of cellulosic materials as well as the reaction mechanism were studied by combined deuteration and FTIR techniques (Driemeier et al. 2015). The deuteration of the amorphous area was also utilized to determine which hydroxyl groups are involved in the hydrogen bond of this amorphous cellulose (Hattori and Arai 2016). Furthermore, the characteristic of structure details and hydrogen-bonding pattern of the cellulose interior chains in woody cell walls were able to be observed at the single-cell level by cellulose deuteration. For example, crystalline lattice spacing of cellulose glucan chains and dimensions of cellulose microfibrils have been characterized by wide-angle neutron scattering (WANS) over a range of equatorial and azimuthal angles, using exchange with D2O to improve contrast. This technique was used to examine the native structure of several cellulosic materials including bamboo wood (Thomas et al. 2015) and celery fibers (Thomas et al. 2013). The microfibril dimensions of cherry, birch and sunflower microfibrils perpendicular to the [200] crystal plane were estimated as 3.0, 3.4 and 3.3 nm, respectively (Thomas et al. 2014). These techniques were recently applied to examine the properties of spruce wood and its structural changes under tension (Thomas et al. 2021). In addition, Guo and Altaner monitored the orientation of hydroxyl groups in deuterated eucalypt wood, suggesting that the inaccessible fraction of cellulose was the primary load-carrying structure (Guo and Altaner 2019).
The most common use of hydrogen–deuterium exchange treatment of cellulose is the determination of cellulose hydroxyl group accessibility (Leboucher et al. 2020). Table 3 summarizes studies that examined the accessibility of cellulose hydroxyl groups to deuteration with FTIR, NIR, DVS (dynamic vapor sorption), NMR and SANS analysis methods. Cellulose internal structural details could also be revealed during these investigations. For example, Hofstetter et al. studied the roles of different hydrogen bonds in cellulose moisture uptake with cellulose deuteration and indicated that there was a shift of the load transfer towards the backbone of the cellulose structure during both H2O and D2O moisture conditioning (Hofstetter et al. 2006). Utilization of 2H MAS NMR provided evidence for two absorbed D2O phases of different mobility in addition to the deuterated hydroxyls (“deuteroxyls”) following vapor phase D2O exchange (Lindh et al. 2017). Comparison of time-resolved SANS and FTIR analyses enabled tracking of both interfibrillar D2O penetration and D exchange of hydroxyl groups (Penttillä et al. 2021). Therefore, the chemical deuteration of water-accessible cellulose helped to uncover the alteration mechanisms of cellulose structure during certain reactions. The native structure of spruce wood was probed by introduction of exchange-resistant OD groups on cellulose surfaces underlying associated xylan chains through by mild treatment with 0.1 M potassium hydroxide in D2O, enabling SANS observation of wider cellulose microfibrillar spacing consistent with the presence of xylan between cellulose microfibrils (Thomas et al. 2020).
5. Summary and outlook
Cellulose deuteration is a promising technology that can be applied to examine cellulose structure, hydrogen bonding, adsorption progress and other cellulose-related reaction mechanisms. Deuterated cellulose can be produced by the biological cultivation of plants or microorganisms in deuterated medium or chemical hydrogen–deuterium exchange treatment, depending on the type of exchangeable or non-exchangeable hydrogen substitution desired.
Deuterium incorporation into cellulose via biological routes is insightful as it could provide deuterium-labeled cellulose without significantly altering cellulose structure, in which deuterated bacteria could possess higher deuterium incorporation rate than the deuterated plant. In general, biological deuteration takes more effort and constrained by species response, while chemical deuteration is easier to accomplish and could be applied to a variety of cellulosic materials. Biological deuteration of cellulose is primarily used when replacement of non-exchangeable (i.e., alkyl) hydrogens is desired. It also results in the deuterium substitution of hydroxyl hydrogens that are not surface accessible without changing the nature of lignocellulosic biomass. Thus, it would help to reveal the original native cellulose internal structure. Chemical routes, which typically substitute solely hydroxyl groups, are more commonly applied for structural investigations of properties such as hydrogen bonding and crystallinity. Characterization and analysis of deuterated cellulose by methods including SANS, FTIR and NMR, has helped to reveal many of the structural and dynamic details of cellulose. Particularly for neutron diffraction, reflectometry, and scattering techniques, deuterium incorporation plays a significant role in fundamental research.
However, there is a lack of material application research for deuterated cellulose. Since the C–D and O–D bond strength is stronger than C–H and O–H bond strength, respectively, it has an impact on chemical reaction rates and other isotope effects (such as biological isotope effect, spectroscopy isotope effect, kinetic isotope effect and thermomechanical isotope effect) related to the properties of deuterated cellulose. These isotope effects are suggestive of future material applications which substantial potential benefit could be gained from use of deuterated cellulose.
References
Bali G, Foston MB, O’Neill HM, Evans BR, He J, Ragauskas AJ (2013) The effect of deuteration on the structure of bacterial cellulose. Carbohydr Res 374:82–88
Barnoud F, Gagnaire D, Odier L, Vincendon M (1971) Biosynthèse de la cellulose bacterienne deutériée: étude par R. M. N. du taux d’incorporation et de la localisation du deutérium. Polymers 10:2269–2273
Bhagia S et al (2018) Ultrastructure and enzymatic hydrolysis of deuterated switchgrass. Sci Rep 8:13226
Blake M, Crane F, Uphaus R, Katz J (1967) Effect of heavy water on the germination of a number of species of seeds. Planta 78:35–38. https://doi.org/10.1007/BF00384855
Bonhoeffer KF (1934) Zeitschrift für Elektrochemie 40 (7b): 469–474, Reaktionen mit schweren Wasserstoff
Brown AJ (1886) On an acetic ferment which forms cellulose. J Chem Soc (London) 49:432–439
Brown RM Jr, Willison JHM, Richardson CL (1976) Cellulose biosynthesis in Acetobacter xylinum: visualization of the site of synthesis and direct measurement of the in vivo process Proc. Natl Acad Sci USA 73:4565–4569
Budarin V, Clark JH, Lanigan B, Shuttleworth PS, Macquarrie DJ (2010) Microwave assisted decomposition of cellulose: a new thermochemical route for biomass exploitation. Biores Technol 101:3776–3779
Cope BT, Bose S, Katz C (1965) Growth of Lemna in H2O–D2O Mixtures: enhancement by Kinetin. Botanical Gazette 126:214–221
Crespi HL, Conrad SM, Uphaus RA, Katz JJ (1960) Cultivation of microorganisms in heavy water. Ann New York Acad Sci 84:648–666
Crespi HL, Daboll HF, Katz JJ (1970) Production of deuterated algae. Biochim Biophys Acta 200(1):26–33
Crespi HL, Katz JJ (1972) Chapter 27, Preparation of Deuterated Proteins and Enzymes. In Methods in Enzymology Vol 26, Enzyme Structure Part C, Ed. CHW Hirs and SN Timasheff, pp. 627–637, Academic Press, New York and London
Daboll H, Crespi H, Katz J (1962) Mass cultivation of algae in pure heavy water. Biotechnol Bioeng 4:281–297. https://doi.org/10.1002/bit.260040304
Driemeier C, Mendes FM, Ling LY (2015) Hydrated fractions of cellulosics probed by infrared spectroscopy coupled with dynamics of deuterium exchange. Carbohydr Polym 127:152–159
Evans BR, Bali G, Reeves DT, O’Neill HM, Sun Q, Shah R, Ragauskas AJ (2014) Effect of D2O on growth properties and chemical structure of annual ryegrass (Lolium multiflorum). J Agric Food Chem 62:2595–2604. https://doi.org/10.1021/jf4055566
Evans BR et al (2015) Production of deuterated switchgrass by hydroponic cultivation. Planta 242:215–222. https://doi.org/10.1007/s00425-015-2298-0
Evans BR, Shah R (2015) Chapter Ten—Development of approaches for deuterium incorporation in plants. In: Methods in Enzymology, vol 565. Academic Press, pp. 213–243. https://doi.org/10.1016/bs.mie.2015.07.014
Evans BR, Pingali SV, Bhagia S, O'Neill HM, Ragauskas AJ (2019b) Structural studies of deuterium-labeled switchgrass biomass. In: Understanding Lignocellulose: Synergistic Computational and Analytic Methods, vol 1338. ACS Symposium Series, vol 1338. American Chemical Society, pp. 17–32. https://doi.org/10.1021/bk-2019-1338.ch002
Evans BR et al. (2019a) Production of deuterated biomass by cultivation of Lemna minor (duckweed) in D2O Planta, 249:1465–1475
Fackler K, Schwanninger M (2011) Accessibility of hydroxyl groups of brown-rot degraded spruce wood to heavy water. J Near Infrared Spectrosc 19:359–368. https://doi.org/10.1255/jnirs.943
Foston MB, McGaughey J, O’Neill H, Evans BR, Ragauskas A (2012) Deuterium incorporation in biomass cell wall components by NMR analysis. Analyst 137:1090–1093. https://doi.org/10.1039/c2an16025k
French AD (2017) Glucose, not cellobiose, is the repeating unit of cellulose and why that is important. Cellulose 24:4605–4609. https://doi.org/10.1007/s10570-017-1450-3
Frilette VJ, Hanle J, Mark H (1948) Rate of exchange of cellulose with heavy water. J Am Chem Soc 70:1107–1113. https://doi.org/10.1021/ja01183a071
Gelenter M, Wang T, Liao S-Y, O’Neill H, Hong M (2017) 2H–13C correlation solid-state NMR for investigating dynamics and water accessibilities of proteins and carbohydrates. J Biomolecular NMR 68:257–270. https://doi.org/10.1007/s10858-017-0124-7
Grusak M (1997) Intrinsic stable isotope labeling of plants for nutritional investigations in humans. J Nutr Biochem 8:164–171. https://doi.org/10.1016/S0955-2863(97)00017-X
Guo F, Altaner CM (2019) Effects of mechanical stretching, desorption and isotope exchange on deuterated eucalypt wood studied by near infrared spectroscopy. Spectrochim Acta Part A Mol Biomol Spectrosc 211:254–259. https://doi.org/10.1016/j.saa.2018.12.012
Han L, Guo J, Wang K, Grönquist P, Li R, Tian X, Yin Y (2020) Hygroscopicity of waterlogged archaeological wood from Xiaobaijiao No.1 Shipwreck related to its deterioration state. Polymers 12:834. https://doi.org/10.3390/polym12040834
Hattori K, Arai A (2016) Preparation and hydrolysis of water-stable amorphous cellulose. ACS Sustain Chem Eng. https://doi.org/10.1021/acssuschemeng.5b01247
He J et al (2014) Controlled incorporation of deuterium into bacterial cellulose. Cellulose 21:927–936. https://doi.org/10.1007/s10570-013-0067-4
Hishikawa Y, Inoue S-I, Magoshi J, Kondo T (2005) Novel tool for characterization of Noncrystalline regions in cellulose: a FTIR deuteration monitoring and generalized two-dimensional correlation spectroscopy. Biomacromolecules 6:2468–2473. https://doi.org/10.1021/bm050032k
Hishikawa Y, Togawa E, Kondo T (2010) Molecular orientation in the Nematic Ordered Cellulose film using polarized FTIR accompanied with a vapor-phase deuteration method. Cellulose 17:539–545. https://doi.org/10.1007/s10570-010-9410-1
Hishikawa Y, Togawa E, Kondo T (2017) Characterization of individual hydrogen bonds in crystalline regenerated cellulose using resolved polarized FTIR spectra. ACS Omega 2:1469–1476. https://doi.org/10.1021/acsomega.6b00364
Hofstetter K, Hinterstoisser B, Salmén L (2006) Moisture uptake in native cellulose—the roles of different hydrogen bonds: a dynamic FT-IR study using Deuterium exchange. Cellulose 13:131–145
Horikawa Y, Sugiyama J (2008) Accessibility and size of Valonia cellulose microfibril studied by combined deuteration/rehydrogenation and FTIR technique. Cellulose 15:419–424
Horikawa Y, Sugiyama J (2009) Localization of crystalline allomorphs in cellulose microfibril. Biomacromol 10:2235–2239
Jiang W et al (2018) A green degumming process of ramie. Ind Crops Prod 120:131–134. https://doi.org/10.1016/j.indcrop.2018.04.045
Johnson DC, Neogi AN (1989) Sheeted products formed from reticulated microbial cellulose. US Patent 4,863,565 dated Sep 5, 1989
Katz JJ, Crespi HL (1966) Deuterated organisms: cultivation and uses. Science 151:1187–1194. https://doi.org/10.1126/science.151.3715.1187
Kirshenbaum I (1951) Physical properties and analysis of heavy water. In H. C. Urey and G. M. Murphy (Series Eds.), National Nuclear Energy Series, Vol. 4 A. 1st edition. New York: McGraw-Hill Book Company, Inc
Klemm D, Heublein B, Fink H-P, Bohn A (2005) Cellulose: fascinating biopolymer and sustainable raw material. Angew Chem Intl Ed 44:3358–3393. https://doi.org/10.1002/anie.200460587
Kondo T, Nojiri M, Hishikawa Y, Togawa E, Romanovicz D, Brown R (2002) Biodirected epitaxial nanodeposition of polymers on oriented macromolecular templates. Proc Natl Acad Sci USA 99:14008–14013. https://doi.org/10.1073/pnas.212238399
Koruza K, Lafumat B, Végvári Á, Knecht W, Fisher SZ (2018) Deuteration of human carbonic anhydrase for neutron crystallography: Cell culture media, protein thermostability, and crystallization behavior. Archives Biochem Biophys 645:26–33. https://doi.org/10.1016/j.abb.2018.03.008
Langan P, Nishiyama Y, Chanzy H (1999) A revised structure and hydrogen-bonding system in cellulose II from a neutron fiber diffraction analysis. J Am Chem Soc 121:9940–9946
Leboucher J et al (2020) High-yield cellulose hydrolysis by HCl vapor: co-crystallization, deuterium accessibility and high-temperature thermal stability. Cellulose 27:3085–3105. https://doi.org/10.1007/s10570-020-03002-2
Lee KY, Bismarck A (2011) Chapter Ten-Assessing the moisture uptake behavior of natural fibres. In: Zafeiropoulos NE (ed.) Interface Engineering of Natural Fibre Composites for Maximum Performance. Woodhead Publishing, pp. 275–288. https://doi.org/10.1533/9780857092281.2.275
Lewis GN (1933) The biochemistry of water containing hydrogen isotope. J Agric Food Chem 55:3503–3504
Lewis GN (1934) The biology of heavy water. Science 79:151–153
Li Y-K, Chir J, Chen F-Y (2001) Catalytic mechanism of a family 3 β-glucosidase and mutagenesis study on residue Asp-247. Biochem J 355:835–840. https://doi.org/10.1042/bj3550835
Lindh EL, Salmén L (2017) Surface accessibility of cellulose fibrils studied by hydrogen–deuterium exchange with water. Cellulose 24:21–33
Lindh EL, Terenzi C, Salmén L, Furó I (2017) Water in cellulose: evidence and identification of immobile and mobile adsorbed phases by 2H MAS NMR. Phys Chem Chem Phys 19:4360–4369
Martínez-Sanz M, Gidley MJ, Gilbert EP (2015a) Application of X-ray and neutron small angle scattering techniques to study the hierarchical structure of plant cell walls: a review. Carbohyd Polym 125:120–134. https://doi.org/10.1016/j.carbpol.2015.02.010
Martínez-Sanz M, Lopez-Sanchez P, Gidley MJ, Gilbert EP (2015b) Evidence for differential interaction mechanism of plant cell wall matrix polysaccharides in hierarchically-structured bacterial cellulose. Cellulose 22:1541–1563. https://doi.org/10.1007/s10570-015-0614-2
Martínez-Sanz M, Gidley MJ, Gilbert EP (2016a) Hierarchical architecture of bacterial cellulose and composite plant cell wall polysaccharide hydrogels using small angle neutron scattering. Soft Matter 12:1534–1549
Martínez-Sanz M, Mikkelsen D, Flanagan B, Gidley MJ, Gilbert EP (2016b) Multi-scale model for the hierarchical architecture of native cellulose hydrogels. Carbohydr Polym 147:542–555
Martínez-Sanz M, Mikkelsen D, Flanagan BM, Rehm C, de Campo L, Gidley MJ, Gilbert EP (2016c) Investigation of the micro- and nano-scale architecture of cellulose hydrogels with plant cell wall polysaccharides: a combined USANS/SANS study. Polymer 105:449–460. https://doi.org/10.1016/j.polymer.2016.07.015
Martínez-Sanz M, Mikkelsen D, Flanagan BM, Gidley MJ, Gilbert EP (2017) Multi-scale characterisation of deuterated cellulose composite hydrogels reveals evidence for different interaction mechanisms with arabinoxylan, mixed-linkage glucan and xyloglucan. Polymer 124:1–11. https://doi.org/10.1016/j.polymer.2017a.07.036
Martínez-Sanz M, Pettolino F, Flanagan B, Gidley MJ, Gilbert EP (2017b) Structure of cellulose microfibrils in mature cotton fibres. Carbohyd Polym 175:450–463. https://doi.org/10.1016/j.carbpol.2017.07.090
Nishiyama Y (2018) Molecular interactions in nanocellulose assembly. Philosophical Trans Royal Soc Math Phys Eng Sci 376:20170047. https://doi.org/10.1098/rsta.2017.0047
Nishiyama Y, Isogai A, Okano T, Müller M, Chanzy H (1999a) Intracrystalline deuteration of native cellulose. Macromolecules 32:2078–2081. https://doi.org/10.1021/ma981563m
Nishiyama Y, Okano T, Langan P, Chanzy H (1999b) High resolution neutron fibre diffraction data on hydrogenated and deuterated cellulose. Int J Biol Macromol 26:279–283
Nishiyama Y, Langan P, Chanzy H (2002) Crystal structure and hydrogen-bonding system in cellulose Iβ from synchrotron X-ray and neutron fiber diffraction. J Am Chem Soc 124:9074–9082
Nishiyama Y, Sugiyama J, Chanzy H, Langan P (2003) Crystal structure and hydrogen bonding system in cellulose Iα from synchrotron X-ray and neutron fiber diffraction. J Am Chem Soc 125:14300–14306
Nishiyama Y, Langan P, Wada M, Forsyth VT (2010) Looking at hydrogen bonds in cellulose. Acta Crystallogr D Biol Crystallogr 66:1172–1177
Okuda A et al (2021) Deuteration aiming for neutron scattering. Biophys Physicobiol. https://doi.org/10.2142/biophysico.bppb-v18.003
O’Neill H et al (2017) Dynamics of water bound to crystalline cellulose. Sci Rep 7:11840. https://doi.org/10.1038/s41598-017-12035-w
O'Neill H et al. (2015) Chapter Six—Production of Bacterial Cellulose with Controlled Deuterium–Hydrogen Substitution for Neutron Scattering Studies. In: Methods in enzymology, vol. 565. Academic Press, pp. 123–146
Penttilä PA, Imai T, Sugiyama J, Schweins R (2018) Biomimetic composites of deuterated bacterial cellulose and hemicelluloses studied with small-angle neutron scattering. Eur Polymer J 104:177–183
Penttilä PA, Zitting A, Lourençon T, Altgen M, Schwerin R, Rautkari L (2021) Water-accessibility of interfibrillar spaces in spruce wood cell walls. Cellulose 28:11231–11245. https://doi.org/10.1007/s10570-021-04253-3
Pingali SV et al (2010a) Sans study of cellulose extracted from switchgrass. Acta Crystallographica Sect D Biol Crystallography D66:1189–1193. https://doi.org/10.1107/S0907444910020408
Pingali SV, Urban VS, Heller WT, McGaughey J, O’Neill HM, Foston M, Myles DA, Ragauskas A, Evans BR (2010b) Breakdown of cell wall nanostructure in dilute acid pretreated biomass. Biomacromol 11:2329–2335
Pingali SV, Urban VS, Heller WT, McGaughey J, O’Neill H, Foston MB, Li H, Wyman CE, Myles DA, Langan P, Ragauskas A, Davison B, Evans BR (2017) Understanding multiscale structural changes during dilute acid pretreatment of switchgrass and poplar. ACS Sustain Chem Eng 5(1):426–435. https://doi.org/10.1021/acssuschemeng.6b01803
Pingali SV, Smith MD, Liu SH, Rawal TB, Pu Y, Shah R, Evans BR, Urban VS, Davison BH, Ca CM, Ragauskas AJ, O’Neill HM, Smith JC, Petridis L (2020) Deconstruction of biomass enabled by local demixing of cosolvents at cellulose and lignin surfaces. Proc Natl Acad Sci U SA 117:16776–16781. https://doi.org/10.1073/pnas.1922883117
Pönni R, Rautkari L, Hill CAS, Vuorinen T (2014) Accessibility of hydroxyl groups in birch kraft pulps quantified by deuterium exchange in D2O vapor. Cellulose 21:1217–1226. https://doi.org/10.1007/s10570-014-0166-x
Ponni R, Galvis L, Vuorinen T (2014) Changes in accessibility of cellulose during kraft pulping of wood in deuterium oxide. Carbohyd Polym 101:792–797. https://doi.org/10.1016/j.carbpol.2013.10.001
Pratt R, Curry J (1937) Growth of Roots in Deuterium Oxide. Am J Botany. https://doi.org/10.2307/2436423
Raghuwanshi VS et al (2016) Bio-deuterated cellulose thin films for enhanced contrast in neutron reflectometry. Cellulose 24:11–20. https://doi.org/10.1007/s10570-016-1108-6
Raghuwanshi VS, Su J, Garvey CJ, Holt SA, Holden PJ, Batchelor WJ, Garnier G (2017) Visualization and quantification of IgG antibody adsorbed at the cellulose-liquid interface. Biomacromol 18:2439–2445. https://doi.org/10.1021/acs.biomac.7b00593
Raghuwanshi VS, Cohen Y, Garnier G, Garvey CJ, Russell RA, Darwish T, Garnier G (2018) Cellulose dissolution in ionic liquid: ion binding revealed by neutron scattering. Macromolecules 51:7649–7655. https://doi.org/10.1021/acs.macromol.8b01425
Reishofer D, Spirk S (2016) Deuterium and Cellulose: A Comprehensive Review. In: Rojas OJ (ed) Cellulose Chemistry and Properties: Fibers, Nanocelluloses and Advanced Materials. Springer International Publishing, Cham, pp. 93–114. https://doi.org/10.1007/12_2015_321
Reitz O, Bonhoeffer K (1934) Über den Einbau von schwerem Wasserstoff in wachsende Organismen. Naturwissenschaften 22:744
Reitz O, Bonhoeffer K (1935) Über den Einbau von schwerem Wasserstoff in wachsende Organismen. Z Phys Chem 172:369–388
Rozenberg M (1996) IR spectra and hydrogen bond energies of crystalline acid salts of carboxylic acids Spectrochimica Acta Part A-molecular and Biomolecular. Spectroscopy 52:1559–1563. https://doi.org/10.1016/0584-8539(96)01703-5
Russell RA, Garvey CJ, Darwish TA, Foster LJR, Holden PJ (2015) Chapter five—biopolymer deuteration for neutron scattering and other isotope-sensitive techniques. Methods Enzymol 565:97–121. https://doi.org/10.1016/bs.mie.2015.06.015
Shah R, Huan S, Pingali SV, Sawada D, Kim S, Evans BR, Davison BH, O’Neill H (2019) Hemicellulose-cellulose composites reveal differences in cellulose organization after dilute acid pretreatment. Biomacromolecules 20(2):893–903
Siegel S, Halpern L, Giumarro C (1964) Germination and seedling growth of winter rye in deuterium oxide. Nature 201:1244–1245. https://doi.org/10.1038/2011244a0
Song Y, Jiang W, Ben H, Meng X, Zhang Y, Han G, Ragauskas AJ (2020) The production of hydrogen–deuterium exchanged cellulose fibers with exchange-resistant deuterium incorporation. Cellulose 27:6163–6174. https://doi.org/10.1007/s10570-020-03230-6
Song Y, Jiang W, Ben H, Zhang Y, Han G, Ragauskas AJ (2021) The preparation and characterization of chemically deuterium incorporated cotton fibers. Cellulose 28:5351–5361. https://doi.org/10.1007/s10570-021-03869-9
Su J et al (2016) Smooth deuterated cellulose films for the visualisation of adsorbed bio-macromolecules. Sci Rep 6:36119. https://doi.org/10.1038/srep36119
Tang G, Qin J, Dolnikowski G, Russell R, Grusak M (2005) Spinach or carrots can supply significant amounts of vitamin A as assessed by feeding with intrinsically deuterated vegetables. Am J Clin Nutr 82:821–828. https://doi.org/10.1093/ajcn/82.4.821
Tarmian A, Burgert I, Thybring EE (2017) Hydroxyl accessibility in wood by deuterium exchange and ATR-FTIR spectroscopy: methodological uncertainties. Wood Sci Technol 51:845–853. https://doi.org/10.1007/s00226-017-0922-9
Thomas LH, Forsyth VT, Sturcova A, Kennedy CJ, May RP, Altaner CM, Apperley DC, Wess TJ, Jarvis MC (2013) Structure of cellulose microfibris in primary cell walls from collenchyma. Plant Physiol 161:465–476
Thomas LH, Forsyth VT, Martel A, Grillo I, Altaner CM, Jarvis MC (2014) Structure and spacing of cellulose microfibrils in woody cell walls of dicots. Cellulose 21:3887–3895. https://doi.org/10.1007/s10570-014-0431-z
Thomas LH, Forsyth VT, Martel A, Grillo I, Altaner CM, Jarvis MC (2015) Diffraction evidence for the structure of cellulose microfibrils in bamboo, a model for grass and cereal celluloses. BMC Plant Biol 15:153
Thomas LH, Martel A, Grillo I, Jarvis MC (2020) Hemicellulose binding and the spacing of cellulose microfibrils in spruce wood. Cellulose 27:4249–4254. https://doi.org/10.1007/s10570-020-03091-z
Thomas LH, Altaner CM, Forsyth VT, Mossou E, Kennedy CJ, Martel A, Jarvis MC (2021) Nanostructural deformation of high-stiffness spruce wood under tension. Sci Rep 11:453. https://doi.org/10.1038/s41598-020-79676-2h
Thybring E, Thygesen L, Burgert I (2017) Hydroxyl accessibility in wood cell walls as affected by drying and re-wetting procedures. Cellulose. https://doi.org/10.1007/s10570-017-1278-x
Traber MG, Leonard SW, Gerd B, Xueyan F, Edward S, Grusak MA, Booth SL (2015) α-Tocopherol disappearance rates from plasma depend on lipid concentrations: studies using deuterium-labeled collard greens in younger and older adults. Am J Clin Nutrition 101:752–759
Tsuchikawa S, Siesler H (2003) Near-infrared spectroscopic monitoring of the diffusion process of deuterium-labeled molecules in wood. Part II: Hardwood Appl Spectroscopy 57:675–681
Väisänen S, Pönni R, Hämäläinen A, Vuorinen T (2018) Quantification of accessible hydroxyl groups in cellulosic pulps by dynamic vapor sorption with deuterium exchange. Cellulose 25:6923–6934. https://doi.org/10.1007/s10570-018-2064-0
Waber J, Sakai W (1974) The effect of growth in 99.8 deuterium oxide on the ultrastructure of winter rye. Plant Physiol 53:128–130. https://doi.org/10.1104/pp.53.1.128
Wada M, Chanzy H, Nishiyama Y, Langan P (2004) Cellulose IIIICrystal structure and hydrogen bonding by synchrotron X-ray and neutron fiber diffraction. Macromolecules 37:8548–8555. https://doi.org/10.1021/ma0485585
Wada M, Nishiyama Y, Bellesia G, Forsyth T, Gnanakaran S, Langan P (2011) Neutron crystallographic and molecular dynamics studies of the structure of ammonia-cellulose I: rearrangement of hydrogen bonding during the treatment of cellulose with ammonia. Cellulose 18:191–206. https://doi.org/10.1007/s10570-010-9488-5
Wiberg KB (1955) The deuterium isotope effect. Chem Rev 55:713–743. https://doi.org/10.1021/cr50004a004
Xiaoyuan Y, Chen W-P, Rendahl A, Hegeman A, Gray W, Cohen J (2010) Measuring the turnover rates of Arabidopsis proteins using deuterium oxide: an auxin signaling case study. Plant J Cell Molecular Biol 63:680–695. https://doi.org/10.1111/j.1365-313X.2010.04266.x
Yamada Y, Yukphan P (2008) Genera and species in acetic acid bacteria. Int J Food Microbiol 125:15–24. https://doi.org/10.1016/j.ijfoodmicro.2007.11.077
Yamada Y, Hoshino KI, Ishikawa T (1997) The phylogeny of acetic acid bacteria based on the partial sequences of 16S Ribosomal RNA: the elevation of the subgenus gluconoacetobacter to the generic level. Biosci Biotechnol. Biochem. 61:1244–1251
Yamada Y, Yukphan P, Vu H, Muramatsu Y, Ochaikul D, Tanasupawat S, Nakagawa Y (2012) Description of Komagataeibacter gen. nov., with proposals of new combinations (Acetobacteraceae). J General Appl Microbiol 58:397–404. https://doi.org/10.2323/jgam.58.397
Yamada Y, Yukphan P, Vu H, Muramatsu Y, Ochaikul D, Nakagawa Y (2011) Subdivision of the genus Gluconacetobacter Yamada, Hoshino and Ishikawa 1998: the proposal of Komagatabacter gen. nov., for strains accommodated to the Gluconacetobacter xylinus group in the α-Proteobacteria Annals of Microbiology 62. https://doi.org/10.1007/s13213-011-0288-4
Yonenobu H, Tsuchikawa S, Sato K (2013) Near-infrared spectroscopic analysis of aging degradation in antique washi paper using a deuterium exchange method. Vib Spectrosc 51:100–104. https://doi.org/10.1016/j.vibspec.2008.11.001
Zachleder V et al (2018) Stable isotope compounds—production, detection, and application. Biotechnol Adv. https://doi.org/10.1016/j.biotechadv.2018.01.010
Zhang M (2020) Recent developments of methyl-labeling strategies in Pichia pastoris for NMR spectroscopy. Protein Expr Purif 166:105521
Zhao D et al (2019) Exploring structural variations of hydrogen-bonding patterns in cellulose during mechanical pulp refining of tobacco stems. Carbohyd Polym 204:247–254. https://doi.org/10.1016/j.carbpol.2018.10.024
Acknowledgments
The authors (YS, WJ, HB, YZ, SZ, GH) acknowledge the support by Science Research Foundation of Dezhou University (2021xjrc207 and 2019xgrc04), National Natural Science Foundation of China (51903131), the National Key Research and Development Program of China (2017YFB0309702), Natural Science Foundation of Shandong Province (ZR2019QEM007 and ZR2020ME076), Key Research and Development Program of Shandong Province (2020CXGC011101), State Key Laboratory of Bio-Fibers and Eco-Textiles (Qingdao University) and Special Foundation of “Taishan Scholar” Construction Program. This study was also supported, in part, by the U. S. Department of Energy Office of Science through the Genomic Science Program, Office of Biological and Environmental Research, under contract FWP ERKP752. Use of transmission electron microscopy and ultramicrotomy was provided by the Center for Nanophase Materials Sciences (CNMS), a U. S. Department of Energy Office of Science User Facility at Oak Ridge National Laboratory (ORNL), Oak Ridge, Tennessee, under proposal no. CNMS2016-R83. Oak Ridge National Laboratory is managed by UT Battelle, LLC, for the U. S. Department of Energy under Contract DE-AC05-00OR22725. The views and opinions of the authors expressed herein do not necessarily state or reflect those of the United States Government or any agency thereof. Neither the United States Government nor any agency thereof, nor any of their employees, makes any warranty, expressed or implied, or assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, apparatus, product, or process disclosed, or represents that its use would not infringe privately owned rights.
Funding
This research was supported by the Science Research Foundation of Dezhou University; the National Natural Science Foundation of China; the National Key Research and Development Program of Shangdong Province; State Key Laboratory of Bio-Fibers and Eco-Textiles; Special Foundation of Taishan Scholar Construction Program; and by the U. S. Department of Energy Office of Science.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This manuscript has been authored by UT-Battelle, LLC, under contract DE-AC05-00OR22725 with the US Department of Energy (DOE). The US government retains and the publisher, by accepting the article for publication, acknowledges that the US government retains a nonexclusive, paid-up, irrevocable, worldwide license to publish or reproduce the published form of this manuscript, or allow others to do so, for US government purposes. DOE will provide public access to these results of federally sponsored research in accordance with the DOE Public Access Plan (http://energy.gov/downloads/doe-public-access-plan)
Rights and permissions
About this article

Cite this article
Song, Y., Meng, X., Jiang, W. et al. Deuterium incorporation into cellulose: a mini-review of biological and chemical methods. Cellulose 29, 4269–4286 (2022). https://doi.org/10.1007/s10570-022-04551-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10570-022-04551-4