
Abstract
The Membrane Theory of Aging proposes that lifespan is inversely related to the level of unsaturation in membrane phospholipids. Calorie restriction (CR) without malnutrition extends lifespan in many model organisms, which may be related to alterations in membrane phospholipids fatty acids. During the last few years our research focused on studying how altering the predominant fat source affects the outcome of CR in mice. We have established four dietary groups: one control group fed 95 % of a pre-determined ad libitum intake (in order to prevent obesity), and three CR groups fed 40 % less than ad libitum intake. Lipid source for the control and one of the CR groups was soybean oil (high in n-6 PUFA) whereas the two remaining CR groups were fed diets containing fish oil (high in n-3 PUFA), or lard (high in saturated and monounsaturated fatty acids). Dietary intervention periods ranged from 1 to 18 months. We performed a longitudinal lifespan study and a cross-sectional study set up to evaluate several mitochondrial parameters which included fatty acid composition, H+ leak, activities of electron transport chain enzymes, ROS generation, lipid peroxidation, mitochondrial ultrastructure, and mitochondrial apoptotic signaling in liver and skeletal muscle. These approaches applied to different cohorts of mice have independently indicated that lard as a fat source often maximizes the effects of 40 % CR on mice. These effects could be due to significant increases of monounsaturated fatty acids levels, in accordance with the Membrane Theory of Aging.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Aging is the time-dependent progressive decline in physiological function with decreased fertility and increased susceptibility of the organism to endogenous and external threats, leading to a wide variety of related diseases such as degenerative and neoplastic disorders. Scientists have demonstrated a strong interest not only in unravelling the causes of aging, but also in discovering how we can manipulate potential causes of aging to decrease, stop, or even revert its rate of progression (Sousa-Victor et al. 2014; Li and Izpisua-Belmonte 2014). Denham Harman proposed that aging is the result of deleterious interactions between free radicals and cellular constituents (Harman 1956). Despite some current controversy, the so-called mitochondrial Free Radical Theory of Aging still stands today as one of the most widely cited theories to explain the causes of aging (Barja 2013, 2014). Closely related to this theory, the Membrane Theory of Aging proposes that lifespan is inversely related to the degree of unsaturation of membrane phospholipids (Pamplona et al. 1998, 2002; Hulbert et al. 2007; Hulbert 2008; Pamplona and Barja 2011). Alterations involved in aging are indeed multifactorial and involve diverse tissue-specific processes at the organismal level, such as genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intracellular communication (López-Otín et al. 2013). Some of these hallmarks point to the mitochondria as playing an essential role in the aging process.
Calorie restriction (CR) without malnutrition increases maximum lifespan and prevents or delays the onset of pathophysiological changes in multiple species (Sohal and Weindruch 1996). Despite intensive research to elucidate how CR improves longevity, the mechanisms responsible for the retardation of aging with CR are not yet entirely understood, although it is generally accepted that longevity-promoting effects of CR can be mediated, at least partially, by the optimization of mitochondrial function (López-Lluch et al. 2006).
Mitochondrial fatty acids, calorie restriction and longevity
Fatty acids are components that could influence lifespan in CR animals. As stated above, the Membrane Theory of Aging proposes that lifespan is inversely related to the level of unsaturation, and in particular the level of n-3 polyunsaturated fatty acids (PUFA) in membrane phospholipids (Pamplona et al. 1998, 2002; Hulbert et al. 2007; Hulbert 2008; Pamplona and Barja 2011). Of note, only two traits currently correlate inversely with maximal longevity among animal species: both the rate of reactive oxygen species (ROS) generation in mitochondria and the degree of fatty acid unsaturation of tissue membranes (Pamplona and Barja 2011; Barja 2014). In accordance, a decrease in long-chain PUFA in mitochondrial membrane phospholipids may be a mechanism contributing to the anti-aging effects of CR (Laganiere and Yu 1989, 1993; Yu et al. 2002). Interestingly, the drug atenolol, a β1-blocker that decreases fatty acid unsaturation in heart and skeletal muscle mitochondria and changes the lipid profile towards that found in long-lived mammals, reverted several age-associated detrimental alterations in mice but did not extend longevity, possibly due to unwanted side effects of the drug (Sánchez-Román et al. 2010, 2014; Gómez et al. 2014). Furthermore, a recent lipidomic approach has demonstrated that CR significantly altered the hepatic lipidome in male C57BL/6 mice and caused a change in the relative abundance of specific triglycerides and phosphatidylethanolamines and reduced hepatic 1-palmitoyl-2-glutaryl-sn-glycero-3-phosphatidylcholine content, a specific product of phospholipid peroxidation (Jové et al. 2014). Less susceptibility of membranes to peroxidation was explained on the basis of a redistribution in the type of unsaturation: CR increased monounsaturated fatty acids (MUFA) in liver, whereas the levels of PUFA were decreased without any observed changes in saturated fatty acids (SFA). These specific changes may be the result of a metabolic reprogramming leading to lower levels of oxidative damage which could contribute to the increased lifespan of CR mice (Jové et al. 2014).
These recent observations are in agreement with the theories that link fatty acid composition of mitochondrial phospholipids to aging, with lipid peroxidation as the mechanism through which fatty acids influence lifespan. In addition, mitochondrial phospholipid fatty acids could also influence aging by altering the activity of membrane proteins (Innis and Clandinin 1981; Daum 1985; Dowhan et al. 2004; Lee 2004; Marsh 2008), membrane permeability (Brand et al. 1994; Brookes et al. 1998), ROS production (Ramsey et al. 2005; Hagopian et al. 2010) or other membrane-linked processes. Membrane n-6 and n-3 fatty acids of 20 carbons in length may also serve as precursors for the formation of eicosanoids, which modulate inflammatory responses (Calder 2004, 2007; Schmitz and Ecker 2008; Deckelbaum and Torrejon 2012). It has been recently demonstrated that chronic, progressive low-grade inflammation induced by knockout of the nfkb1 subunit of the transcription factor NF-kB induces premature aging in mice (Jurk et al. 2014). Thus, membrane phospholipid fatty acids may influence aging by promoting oxidative damage, influencing mitochondrial function and modulating inflammation.
Dietary lipids and lifespan
There is considerable interest in the role that specific dietary lipids play in human health and longevity (Lands 2014). In particular, several epidemiological studies have focused on the potential adverse effects of saturated fats (Staessen et al. 1997; Kromhout et al. 2000; Leosdottir et al. 2005; Tucker et al. 2005; Chen et al. 2011) and the positive effects of fish oil (König et al. 2005; León et al. 2008; Yamagishi al. 2008; Gopinath et al. 2011). It has been reported that saturated fat consumption is positively associated with coronary heart disease mortality (Tucker et al. 2005; Chen et al. 2011) and all-cause mortality in men (Staessen et al. 1997; Kromhout et al. 2000). In contrast, several studies have reported that fish oil consumption is negatively associated with coronary heart disease mortality (König et al. 2005; León et al. 2008; Yamagishi al. 2008). However, these retrospective studies do not contain the dietary controls needed to truly determine if a specific fat source alone is capable of altering longevity. To address this question, a few rodent longevity studies have been completed where the animals were fed diets that differed only in their lipid component. Most of these studies have largely been completed in short-lived mouse or rat models and have produced mixed results. For example, it has been reported that lifespan is increased in autoimmune lupus-prone mice fed a diet containing fish oil versus corn oil (Jolly et al. 2001; Halade et al. 2010). Other studies, however, have indicated that lifespan is decreased in both diabetic rats fed fish versus corn oil (Berdanier 1992) and senescence-accelerated mice fed fish oil (Tsuduki et al. 2011) or perilla oil (source of 18:3 n-3) (Umezawa et al. 2000) versus safflower oil. Moreover, a recent longevity study carried out with long-lived, male, B6C3F1 mice fed diets supplemented with krill oil and Lovaza, a pharmaceutical grade fish oil, beginning at 12 months of age, has demonstrated a 6.6 % lifespan shortening relative to controls (Spindler et al. 2014). Thus, there is little information about the influence of dietary fatty acids on lifespan in long-lived strains of mice (or rats). Also, previous studies in humans and rodents have been almost exclusively completed in individuals allowed ad libitum access to food, and it is not known if specific lipids have the same effect on health and longevity under either CR or ad libitum feeding conditions.
Optimization of pro-longevity actions of CR diets
It is generally assumed that the reduction in caloric intake itself is the major dietary factor responsible for the extension of life with feeding restriction in the rat. In addition, it is also clear that an extreme reduction of methionine intake can extend the life of rats to a similar extent as can CR (Masoro 2006). However, little is known about whether there is an optimum diet composition for promoting lifespan extension with CR. A number of diets, with a range of ingredients, have been used for CR and aging studies (Pugh et al. 1999), but so far very few studies have attempted to compare diets to ascertain if dietary composition influences the response to CR. It has been reported that mean lifespan is increased in CR rats consuming a diet with sucrose versus cornstarch as the primary carbohydrate source (Murtagh-Mark et al. 1995). It has also been shown that the upper 10th percent survival is slightly increased in CR rats consuming a high versus moderate protein diet (Masoro et al. 1989). The potential importance of dietary composition to the retardation of aging with CR is reinforced by the recent idea that differences in diet composition could have contributed to the different outcomes in the two studies investigating the influence of CR on lifespan in rhesus macaques (Cava and Fontana 2013; Colman et al. 2014). In particular, the study at the Wisconsin National Primate Research Center (WNPRC) demonstrated a longevity increase in the CR group (Colman et al. 2009) while no differences were observed between diet groups in the National Institute of Aging (NIA, NIH) study (Mattison et al. 2012). Interestingly, if we compare the diets used in the WNPRC and NIA rhesus macaques studies, it is unveiled that all of the NIA monkeys received a whole-food diet rich in phytochemicals whereas the WNPRC monkeys received semipurified diets with sucrose and corn oil. Although experimental validation is needed, the possibility exists that the beneficial effects on lifespan of the combination of phytochemical-rich pescovegetarian diets and mild CR in the NIA control monkeys are already maximized (Cava and Fontana, 2013). Additional information is clearly needed to determine the extent to which diet composition influences the response to CR.
Calorie restriction diets differing in the major fat source
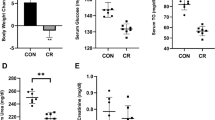
Since fatty acids profile may play a prominent role in determining the positive effects of CR on longevity (Jové et al. 2014), manipulation of membrane fatty acids by feeding CR animals with diets containing different lipid compositions is a valuable strategy to determine their role in CR intervention. We have focused our research on how altering the predominant fat source affects the outcome of CR in mice. For these studies, C57BL/6 J mice were randomly assigned into 4 dietary groups fed semi-purified diets based on the AIN-93 formulas. The control group was fed 95 % of a pre-determined ad libitum intake (12.5 kcal) in order to prevent obesity, whereas the CR dietary groups were fed 40 % less than ad libitum intake. All diets were identical except for dietary lipid source, which was soybean oil (high in n-6 PUFA) for both control and one of the CR groups. The two remaining CR groups were fed diets containing either fish oil (high in n-3 PUFA) or lard (high in saturated and monounsaturated fatty acids) as main sources of dietary fat respectively. Soybean oil (14 % of total fat content) was added to the fish oil (AIN93G and AIN93M) and lard (AIN93M only) diets to insure adequate intake of linoleic acid. Three independent cohorts of mice were subjected to dietary intervention. Two of these cohorts were maintained at the University of California, Davis and used in a longitudinal lifespan study and a cross-sectional study set up to evaluate several mitochondrial parameters which included fatty acid composition, H+ leak, activities of electron transport chain (ETC) enzymes, ROS generation and lipid peroxidation in liver and skeletal muscle. The third cohort was maintained at the University Pablo de Olavide (Sevilla, Spain) and used in a cross-sectional study set up to evaluate mitochondrial ultrastructure and mitochondrial apoptotic signaling. The aim of this paper is to provide a comprehensive review of these previous investigations, and the main results of our studies are summarized in the following sections.
The effect of dietary fat on mitochondrial fatty acid composition in CR mice
In ad libitum fed animals, it has been shown that dietary lipids can alter the fatty acid profile of mitochondrial membranes in multiple tissues, including liver, heart, brain, and skeletal muscle (Yamaoka et al. 1988; Quiles et al. 2002; Ramsey et al. 2005; Tahin et al. 1981). Do these alterations also occur in CR animals? Our first major goal was to determine if alterations in dietary lipids could lead to changes in mitochondrial phospholipids fatty acid composition in CR mice, since a previous study has reported that CR dampened dietary fat-induced changes in liver plasma membrane phospholipid composition (Cha and Jones 2000). Membrane n-3 fatty acid levels were increased in all phospholipids classes in both skeletal muscle (Chen et al. 2012) and liver mitochondria (Chen et al. 2013) in 1 month CR mice consuming fish oil compared to all other diet groups. Similarly, membrane linoleic acid (18:2 n-6) levels were increased in liver and skeletal muscle mitochondria from CR mice consuming soybean oil (high in 18:2 n-6) compared to all other CR groups. In liver, this increase in 18:2 n-6 took place in all phospholipids, while in skeletal muscle the increase of 18:2 n-6 was due primarily to phosphatidylcholine. Mice consuming lard had increased levels of MUFA in liver and skeletal muscle phospholipids, but the MUFA increase in skeletal muscle was limited to phosphatidylcholine (Chen et al. 2012, 2013). In the case of mice fed experimental diets for 8 mo, it was found that fish oil markedly increased n-3 fatty acids whereas soybean oil increased 18:2 n-6 levels in skeletal muscle mitochondrial phospholipids (Chen et al. 2014). Taken together, our results demonstrate that skeletal muscle and liver mitochondrial phospholipid fatty acids readily change to reflect the dietary fat source in CR mice, indicating that dietary lipid manipulations can be used to test the role of mitochondrial membrane phospholipid fatty acid composition on CR action.
Mitochondrial proton leak
Mitochondrial H+ leak, whereby protons bypass the ATP synthase and passively cross the mitochondrial inner membrane, is a major energy expending process responsible for approximately 20 % of resting energy expenditure (Ramsey et al. 2000). Overall H+ leak is thought to consist of basal leak that is unregulated and inducible leak that is regulated by either the uncoupling proteins or the adenosine monophosphate/adenine nucleotide translocase (Brookes et al. 2005). It is well established that basal H+ leak increases with age (Hagen et al. 1997; Harper et al. 1998; Lal et al. 2001) although the CR effects on H+ leak have provided conflicting results, with CR inducing a decrease (Hagopian et al. 2005), increase (Lambert and Merry 2004), or no change (Ramsey et al. 2004; Lambert and Merry 2005) in liver mitochondrial H+ leak. In the case of skeletal muscle, proton leak is either not altered or decreased with CR depending on duration of CR and/or animal age (Asami et al. 2008).
Thus, we were interested in determining how CR and dietary fat affect H+ leak in both liver and skeletal muscle of mice fed our experimental diets. In liver, 1 month of CR did not markedly alter H+ leak in comparison with the control group. However, when comparing the three CR groups that differed in fat source, we found that the lowest H+ leak occurred in CR-Lard group whereas the CR-Fish animals had increased H+ leak (Chen et al. 2013). Similarly, in skeletal muscle, mitochondrial H+ leak was also lower in CR mice consuming lard compared to all other groups (Chen et al. 2012), although these differences disappeared at 8 months CR (Chen et al. 2014). It has previously been shown that age-related changes in H+ leak kinetics of skeletal muscle mitochondria primarily take place in control rather than in CR mice (Asami et al. 2008). It is thus possible that time-related differences in H+ leak kinetics between CR and control mice reflect the fact that CR mitigates age-related changes in H+ leak. The mechanism through which dietary fatty acids influence mitochondrial H+ leak is not entirely clear. Comparative studies have reported that membrane unsaturation index (UI) and n-3 PUFA are positively correlated with mitochondrial H+ leak (Porter et al. 1996; Brookes et al. 1998). However, studies using liposomes have found that proton leak through phospholipid bilayers (lipid–lipid interactions) account for only a small amount of mitochondrial H+ leak (Brookes et al. 1997) and studies using a range of mammalian species indicate that mitochondrial membrane area has a much greater influence on proton leak than membrane fatty acid composition (Porter et al. 1996). In our studies, the fact that differences in H+ leak among the diet groups disappeared in skeletal muscle at 8 months of CR, despite the fact that mitochondrial fatty acid composition continued to differ dramatically between groups, supports the notion that membrane fatty acid composition is not the primary determinant of mitochondrial H+ leak. Additional research is needed to determine if dietary lipids influence mitochondrial H+ leak by altering mitochondrial morphology, changing interactions between membrane proteins and fatty acids or other processes. In sum, our results suggest that lard may help to induce mitochondrial changes which conserve energy in CR mice. Such an adaptation may be beneficial when animals are faced with maintaining energetically costly post-mitotic tissues while decreasing energy intake.
Activities of the mitochondrial electron transport complexes
In liver mitochondria, we found that 1 month of CR did not significantly alter ETC enzyme activities (Chen et al. 2013). This is in contrast with previous results obtained with skeletal muscle mitochondria, which showed that the activities of ETC Complexes I, III and IV were lower in mitochondria from CR compared with control mice at 10 months of age (or ∼26 weeks of CR). However, at older age (20 months), CR mice did not experience the age-related decrease in ETC enzyme activity observed in control animals (Desai et al. 1996). Young adult rats (8–10 months old) on CR for 4.5–6.5 months were also reported to have lower Complex IV activity in muscle mitochondria compared with their age-matched ad libitum controls (Hepple et al. 2006). When we studied the effect of dietary fat in CR mice, it was found that the CR-Fish group had lower Complex I activity than the CR-Soy group and lower Complex II activity than both the CR-Soy and CR-Lard groups. A decrease in Complex I activity in the CR-Lard group was the only difference between the CR-Lard and CR-Soy groups. Our results point out the importance of the tissue under study (liver vs. muscle) and the duration of CR regime on the effects of this intervention in ETC enzyme activities. Furthermore, dietary lipids can differentially influence the activities of ETC enzymes in mitochondria from CR mice, which agrees with the previously reported role of cardiolipin fatty acid composition on ETC activities (Chen et al. 2013).
Mitochondrial ROS generation
CR has been reported to decrease mitochondrial production of ROS in a variety of tissues (Gredilla and Barja 2005; Sohal and Weindruch 1996), but the influence of short-term CR on liver ROS production is not entirely clear since studies in rats have reported that ROS production is not decreased in liver mitochondria following 1 month 40 % CR (Ramsey et al. 2004). Furthermore, 6–7 weeks 8.5 or 25 % CR did not change ROS production in liver mitochondria although different markers of protein oxidation, glycoxidation and lipoxidation were significantly decreased (Gomez et al. 2007). In contrast, it has been reported that ROS production is decreased at 6 weeks of 40 % CR in rat liver mitochondria respiring on pyruvate/malate (Gredilla et al. 2001). In our model, we found that 1 month of CR did not decrease ROS production in liver mitochondria respiring on either substrate alone or substrate plus rotenone (an inhibitor of Complex I). In addition, a decrease in ROS production was only observed with CR when liver mitochondria were respiring on pyruvate/malate plus the Complex III antimycin A (Chen et al. 2013).
When we tested the effect of dietary fat, it was found that mitochondrial H2O2 production was significantly lower in the CR-Fish compared with the other CR groups under all substrate (succinate, or pyruvate/malate, or pyruvate/malate/succinate) and substrate plus inhibitor conditions (Chen et al. 2013). These results are in accordance with our previous results obtained with fat-1 mice, which express the C. elegans fat-1 gene encoding a desaturase that uses n-6 fatty acids as a substrate for the formation of n-3 fatty acids. Liver mitochondria obtained from these mice were enriched in n-3 PUFA and had lower H2O2 production when respiring on either succinate or succinate/glutamate/malate compared with control mice (Hagopian et al. 2010).
Long-term (1 year or longer) CR decreases ROS production in skeletal muscle mitochondria (Bevilacqua et al. 2005; Drew et al. 2003), whereas ROS production was either decreased (Bevilacqua et al. 2004) or not changed (Gredilla et al. 2004) with short-term CR. Skeletal muscle ROS production was not altered in mitochondria respiring on substrates in the absence of ETC inhibitors from short-term CR mice (Faulks et al. 2006). In accordance with these previous studies, we found that H2O2 production was not altered at 1 month of CR in mitochondria respiring on substrate (succinate, pyruvate/malate, or succinate/pyruvate/malate) without ETC inhibitors, but was altered in the presence of ETC inhibitors with a diminished ROS producing capacity in both Complexes I and III in all three CR groups regardless of dietary fat, without significant differences among CR groups (Chen et al. 2012). However, after 8 months of CR there were no differences between control and CR groups in H2O2 production regardless of substrates and/or inhibitors used. The CR-Lard group had decreased ROS producing capacity from complex III although dietary lipids had little influence on ROS produced from backflow into complex I (Chen et al. 2014).
In sum, short-term CR may decrease maximal capacity for ROS production from Complex III in liver mitochondria (Chen et al. 2013). In addition, our studies with mice fed CR diets with different fat sources support the idea that phospholipid fatty acid composition may have a major influence on liver mitochondrial ROS production. Fish oil consumption changes mitochondrial phospholipid fatty acid composition and decreases ROS production in both Complexes I and III (Chen et al. 2013). In skeletal muscle CR produces relatively rapid (within 1 month) changes in the mitochondrial ETC which could influence ROS production under conditions which increase the reduction state of the ETC components (i.e. in the presence of ETC inhibitors). Changes elicited by short-term CR may be due to protein modifications and/or ETC complex assembly, and are unlikely to be related to fatty acid modifications (Chen et al. 2012). Although these alterations were attenuated by age, a protective effect of CR-Lard diet against mitochondrial ROS production under certain assay conditions becomes increasingly apparent in skeletal muscle from mice fed on a long-term CR regime. Thus, the length of CR and/or age may influence changes between dietary lipid groups in ROS production by skeletal muscle mitochondria (Chen et al. 2014).
Lipid peroxidation
A cornerstone of the Membrane Theory of Aging is that increased lipid peroxidation and oxidative damage to membranes with increased levels of highly unsaturated fatty acids (HUFA) lead to a decrease in lifespan (Pamplona et al. 1998, 2002; Hulbert et al. 2007; Hulbert 2008; Pamplona and Barja 2011). In the case of liver mitochondria there were no differences in lipid peroxidation (thiobarbituric acid–reacting substance (TBARS) assay) between the CR-Soy and control groups, and among the three CR groups, at 1 month of CR, which indicates that liver mitochondrial lipid peroxidation is not altered by dietary lipid source in short-term CR mice (Chen et al. 2013). We observed that skeletal muscle mitochondria from the CR-Fish group had increased lipid peroxidation compared with both control and CR-Lard groups, despite consuming a diet containing twice the amount of the antioxidant t-butylhydroquinone as the other groups (Chen et al. 2012). These results are in agreement with other studies showing that fatty acid UI is positively associated with the level of mitochondrial oxidative damage (Herrero et al. 2001; Pamplona et al. 2004). Long-term ingestion of fish oil increased oxidative stress and decreased lifespan in senescence-accelerated mice (Tsuduki et al. 2011), but we did not observe any significant differences between any of the groups of mice in mitochondrial lipid peroxidation at 8 months of CR (Chen et al. 2014). However, it has to be taken into account that these results were obtained by using TBARS method, which has proven to work reasonably well when applied to defined systems, such as liposomes and microsomes, but also to be prone to interferences when applied to body fluids and tissue samples (Meagher and Fitzgerald 2000). Thus, these observations should be taken with caution until confirmed by additional methodologies.
Markers of mitochondrial ultrastructure and dynamics
Liver was studied in mice fed experimental diets for 6 and 18 months starting at an age of 3 months. Mitochondria from CR-Lard fed young adult mice showed increased size compared with all other diets, but this parameter remained unaltered in CR-Fish and CR-Soy compared with the control group (Khraiwesh et al. 2013). CR also induced changes in mitochondrial shape, with a decrease of circularity coefficients in all CR groups, and particularly in CR-Fish, compared with the control group. In addition, the number of cristae per mitochondrion was significantly higher in all CR groups compared with the control group, which could represent a metabolic adaptation to the low-energy state under CR conditions (Khraiwesh et al. 2013). The percentage of cell volume occupied by mitochondria and the number of mitochondria per cell volume unit were significantly increased in all CR groups compared with the control group, which agrees with a previous quantitative study (López-Lluch et al. 2006). On the other hand, no differences in these parameters were observed among the three CR groups (Khraiwesh et al. 2013). Interestingly, similar changes were also observed in mice fed a diet supplemented with resveratrol, a mimetic of CR (Agarwal and Baur 2011; Villalba and Alcaín 2012).
In the case of 21 months old mice, CR also resulted in increased mitochondrial number and volume, although these results also depended on the fat source since CR-Lard and CR-Soy groups exhibited larger mitochondria than the CR-Fish group. Mitochondrial shape also varied with CR in old mice, with more spherical mitochondria being found in all the CR groups regardless of dietary fat. Mean number of cristae per mitochondria and mean crista length were also increased when comparing CR animals with controls and this change was also affected by dietary fat, since a sequential increase in cristal length was found with CR-Soy > CR-Lard > CR-Fish > control (Khraiwesh et al. 2014). These results contrast with those reported in young mice subjected to 6 months of CR where mitochondria of increased size were found only in the CR-Lard group (Khraiwesh et al. 2013).
We found increased levels of proteins related to mitochondrial fission (Fis1 and Drp1) by CR in young adults, but no changes were detected in proteins involved in mitochondrial fusion (Mfn1, Mfn2 and OPA1). Our results on Drp1 and Fis1 are in accordance with previous investigations (Nisoli et al. 2005; López-Lluch et al. 2006) showing an increased number of mitochondria together with an increase in parameters related to mitochondrial biogenesis in animals subjected to CR. A similar result has been reported in animals fed a resveratrol-supplemented diet (Baur et al. 2006). Interestingly, the effect of CR on fission proteins was exacerbated by lard and diminished by fish oil, even while liver mitochondria were the largest in CR-Lard compared with the other dietary groups (Khraiwesh et al. 2013). However, these dietary fat-induced differences in markers of mitochondrial dynamics were attenuated by aging (Khraiwesh et al. 2014).
In sum, modification of mitochondrial ultrastructure by CR and dietary lipid composition is influenced by age and/or duration of dietary intervention. Long-term CR is associated with increases of mitochondrial abundance and cristal number and length in mouse liver. The increase of mitochondrial abundance by this intervention is independent of level of fatty acid unsaturation (Khraiwesh et al. 2013) and is also produced by the CR mimetic resveratrol. Interestingly, although some of the changes are attenuated by aging, the effect of dietary fat under CR conditions is more pronounced in aged mice.
Apoptotic signaling
Apoptosis regulates cellular turnover in mitotic tissues (such as liver) and is involved in the onset of sarcopenia in aging skeletal muscle (Evan and Littlewood 1998; Kanzler and Galle 2000; Dirks and Leeuwenburgh 2002, 2004; Phillips and Leeuwenburgh 2005; Chung and Ng 2006; Adams and Cory 2007; Marzetti et al. 2008a, b, 2009; Seo et al. 2008; Wohlgemuth et al. 2010; Hanahan and Weinberg 2011). Our studies of apoptotic signaling were carried out on mice fed experimental diets for 6 or 18 months starting at an age of 3 months and analyses were performed both in liver and in skeletal muscle. An age-linked increase in the mitochondrial apoptotic pathway was detected with CR in liver, including a decrease in Bcl-2/Bax ratio, an enhanced release of cytochrome c to the cytosol and higher caspase-9 activity. However, these changes were not fully transmitted to the effectors AIF and caspase-3. In addition, CR (which completely abated aging-related inflammatory alterations in liver) and dietary fat altered the activities of caspases-8, -9 and -3. Of note, two well-recognized makers of aging liver, as DNA fragmentation and nuclear mean area, were dramatically increased in all aged animals with the remarkable exception of the CR-Lard group (López-Domínguez et al. 2014a).
In skeletal muscle, 6 months of CR downregulated several proapoptotic mediators, such as lipid hydroperoxides, plasma membrane neutral sphingomyelinase, Bax levels, and release/accumulation of cytochrome c and AIF into the cytosol. CR also improved structural features of gastrocnemius fibers by increasing cross-sectional area and decreasing circularity of fibers in cross sections. Fish oil augmented the protective effect of CR in young animals, decreasing plasma membrane neutral sphingomyelinase, Bax levels, caspase-8 and -9 activities, while increasing levels of the antioxidant coenzyme Q at the plasma membrane, and potentiating the increase of cross-sectional area and the decrease of fiber circularity in cross sections. Many of these changes were not observed in the CR-Lard group (López-Domínguez et al. 2013). On the other hand, the most prominent change observed in aged mice was found for caspase-9 activity, a marker of mitochondrial apoptosis, which exhibited a dramatic increase with aging in the CR-Fish group but not with CR-Lard, while CR-Soy showed an intermediate effect (López-Domínguez et al. manuscript in preparation).
Our observations support the idea that the influence of CR and dietary fat on apoptotic signaling in liver and skeletal muscle is age dependent. Lard elicits protective changes in hepatic homeostasis with aging in mice fed under CR (López-Domínguez et al. 2014a). In addition, although fish oil attenuates skeletal muscle apoptotic signaling in young CR mice, most of these changes were abolished or even reverted in aged mice, with a significant decrease of caspase-9 activity, a marker of mitochondrial apoptosis, in the CR-Lard group.
Effect of dietary fat on lifespan in CR mice
Lifespan was increased in CR mice consuming lard > soybean oil > fish oil-containing diets (López-Domínguez et al. 2014b). There were no differences in prevalence of neoplasms or other major measures of end-of-life pathology between the three CR diet groups. Thus, differences in lifespan between the CR-Lard mice and the other CR groups were likely due to delay in onset of disease rather than preventing the occurrence of specific disease conditions. Longevity improvement in the CR-Lard group is consistent with the Membrane Theory of Aging, and questions the efficacy of feeding diets high in PUFA to CR animals. Furthermore, these results suggest that lipid composition of the diet should be considered when designing diets to maximize lifespan extension with CR.
A possible explanation for dietary fat source effects on lifespan in CR mice
There has been considerable interest in the possible health benefits of dietary n-3 PUFA for humans and other animals, mainly by the role of these fatty acids in inflammation (Lands 2014). In particular, n-3 PUFA form eicosanoids, resolvins and docosanoids, which are anti-inflammatory, in contrast with n-6 PUFA, which form pro-inflammatory eicosanoids (Calder 2004, 2007, 2012). Also, some studies have reported that fat mass is decreased in rodents (Jones 1989; Hill et al. 1993; Su and Jones 1993; Baillie et al. 1999; Tsuboyama-Kasaoka et al. 2008) and humans (Couet et al. 1997; Noreen et al. 2010) consuming diets containing fish oil (n-3 PUFA). Fatty acid-induced changes in mitochondria could be a major contributor to this decrease in adiposity (Nakamura et al. 2014). There is evidence that consumption of fish oil increases mitochondrial biogenesis (Flachs et al. 2005), at least in some tissues, and increases capacity for mitochondrial fatty acid oxidation (Halminski et al. 1991; Ide et al. 2000). Dietary fish oil has been shown to increase the activities of enzymes involved in fatty acid oxidation (Ide et al. 2000; Hong et al. 2003) and, in particular, to increase the activity of the membrane-bound enzyme carnitine palmitoyltransferase I (Power et al. 1994; Power and Newsholme 1997; Hong et al. 2003), which plays a central role in regulating the rate of mitochondrial β-oxidation. There is also some evidence that diet-induced increases in mitochondrial n-3 PUFA alter the activities of ETC enzymes (Yamaoka et al. 1988; McMillin et al. 1992; Barzanti et al. 1994; Infante et al. 2001), although additional work is needed to determine the extent of these changes. We have reported that mitochondrial ROS production may be decreased in response to increased mitochondrial phospholipid n-3 PUFA, and this may contribute to protect the mitochondria from oxidative damage (Ramsey et al. 2005; Hagopian et al. 2010). Thus, fish oil induces a number of beneficial mitochondrial changes which may lead to increased lifespan in ad libitum fed animals.
The possibility exists, however, that dietary n-3 PUFA may not be beneficial to CR animals. CR induces many of the same physiological changes as dietary n-3 PUFA, including decreased ROS production (Sohal and Weindruch 1996; Gredilla and Barja 2005), adiposity (Ramsey and Hagopian 2006; Speakman and Mitchell 2011) and inflammation (Chung et al. 2001; Fontana 2009), and increased mitochondrial biogenesis (López-Lluch et al. 2006; Nisoli et al. 2005; Civitarese et al. 2007) and fatty acid oxidation (Bruss et al. 2009). Thus, n-3 PUFA may not be able to induce additional changes in CR animals. On the other hand, long chain n-3 PUFA are very susceptible to peroxidation (Crockett 2008). Increased peroxidizability of membranes from CR animals consuming diets enriched in n-3 fatty acids could cause impaired mitochondrial function and decreased lifespan, as previously indicated (Pamplona et al. 1998; Hulbert et al. 2007; Hulbert 2008; Pamplona and Barja 2011), and these alterations could be particularly important if n-3 PUFA induce no benefits beyond those already obtained by CR per se. There is evidence that consumption of diets high in saturated and monounsaturated fatty acids may increase ETC activity in old rats (Bronnikov et al. 2010). It has also been reported that mitochondrial ROS production and oxidative damage are decreased (Huertas et al. 1999; Mataix et al. 2006; Mujahid et al. 2009) and mitochondrial function is improved (Mataix et al. 2006) in animals consuming diets high in monounsaturated fats versus diets high in PUFA. Thus, the possibility exists that diets high in saturated and/or monounsaturated fatty acids may show clear benefits in CR animals that are not overweight and have low levels of inflammation.
Concluding remarks
Very different approaches (biochemical, ultrastructural, lifespan analysis) applied to three different cohorts of mice have independently indicated that lard as a fat source often maximizes the effects of 40 % CR on mice, in accordance with the Membrane Theory of Aging (see Table 1). A limitation of our studies, and of all studies using complex dietary lipid sources, is that various dietary lipids differ in multiple fatty acids. Thus, studies using diets with purified fatty acids will be likely required to identify the specific fatty acids which influence lifespan in CR mice. The fact that the CR-Lard diet significantly increased MUFA levels in liver and muscle phospholipids and the recent demonstration that CR produces a redistribution in the type of unsaturation with a significant increase of MUFA in liver (Jové et al. 2014), make it very likely that MUFA increases may be a causal factor in the observed effects of CR-lard diet. Additional studies will also be needed to determine if the increased lifespan in 40 % CR mice consuming the lard diet would also be observed at other levels of energy intake. Although the focus of our studies was on dietary lipid composition, it is possible that composition of protein, carbohydrates, or other dietary components may also influence lifespan. Studies comparing various diets under identical conditions in animals maintained on specific levels of energy intake are needed to truly determine the extent to which various diets influence lifespan in animals maintained on CR.
References
Adams JM, Cory S (2007) The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene 26:1324–1337. doi:10.1038/sj.onc.1210220
Agarwal B, Baur JA (2011) Resveratrol and life extension. Ann N Y Acad Sci 1215:138–143. doi:10.1111/j.1749-6632.2010.05850.x
Asami DK, McDonald RB, Hagopian K, Horwitz BA, Warman D, Hsiao A, Warden C (2008) Ramsey JJ (2008) Effect of aging, caloric restriction, and uncoupling protein 3 (UCP3) on mitochondrial proton leak in mice. Exp Gerontol 43(12):1069–1076. doi:10.1016/j.exger.2008.09.010
Baillie RA, Takada R, Nakamura M, Clarke SD (1999) Coordinate induction of peroxisomal acyl-CoA oxidase and UCP-3 by dietary fish oil: a mechanism for decreased body fat deposition. Prostaglandins Leukot Essent Fatty Acids 60:351–356. doi:10.1016/S0952-3278(99)80011-8
Barja G (2013) Updating the mitochondrial free radical theory of aging: an integrated view, key aspects, and confounding concepts. Antioxid Redox Signal 20:1420–1445. doi:10.1089/ars.2012.5148
Barja G (2014) The mitochondrial free radical theory of aging. Prog Mol Biol Transl Sci 127:1–27. doi:10.1016/B978-0-12-394625-6.00001-5
Barzanti V, Battino M, Baracca A, Cavazzoni M, Cocchi M, Noble R, Maranesi M, Turchetto E, Lenaz G (1994) The effect of dietary lipid changes on the fatty acid composition and function of liver, heart and brain mitochondria in the rat at different ages. Br J Nutr 71:193–202. doi:http://dx.doi.org/10.1079/BJN19940126
Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, Boss O, Gwinn D, Wang M, Ramaswamy S, Fishbein KW, Spencer RG, Lakatta EG, Le Couteur D, Shaw RJ, Navas P, Puigserver P, Ingram DK, de Cabo R, Sinclair DA (2006) Resveratrol improves health and survival of mice on a high-calorie diet. Nature 444:337–342. doi:10.1038/nature05354
Berdanier CD, Johnson B, Hartle DK, Crowell W (1992) Life span is shortened in BHE/cdb rats fed a diet containing 9% menhaden oil and 1% corn oil. J Nutr 122:1309–1317
Bevilacqua L, Ramsey JJ, Hagopian K, Weindruch R, Harper ME (2004) Effects of short- and medium-term calorie restriction on muscle mitochondrial proton leak and reactive oxygen species production. Am J Physiol Endocrinol Metab 286:E852–E861. doi:10.1152/ajpendo.00367.2003
Bevilacqua L, Ramsey JJ, Hagopian K, Weindruch R, Harper ME (2005) Long-term caloric restriction increases UCP3 content but decreases proton leak and reactive oxygen species production in rat skeletal muscle mitochondria. Am J Physiol Endocrinol Metab 289:E429–E438. doi:10.1152/ajpendo.00435.2004
Brand MD, Couture P, Hulbert AJ (1994) Liposomes from mammalian liver mitochondria are more polyunsaturated and leakier to protons than those from reptiles. Comp Biochem Physiol Biochem Mol Biol 108:181–188. doi:10.1016/0305-0491(94)90064-7
Bronnikov GE, Kulagina TP, Aripovsky AV (2010) Dietary supplementation of old rats with hydrogenated peanut oil restores activities of mitochondrial respiratory complexes in skeletal muscles. Biochemistry (Mosc) 75:1491–1497. doi:10.1134/S0006297910120102
Brookes PS (2005) Mitochondrial H(+) leak and ROS generation: an odd couple. Free Radic Biol Med 38:12–23. doi:10.1016/j.freeradbiomed.2004.10.016
Brookes PS, Rolfe DF, Brand MD (1997) The proton permeability of liposomes made from mitochondrial inner membrane phospholipids: comparison with isolated mitochondria. J Membre Biol 155:167–174. doi:10.1007/s002329900168
Brookes PS, Buckingham JA, Tenreiro AM, Hulbert AJ, Brand MD (1998) The proton permeability of the inner membrane of liver mitochondria from ectothermic and endothermic vertebrates and from obese rats: correlations with standard metabolic rate and phospholipid fatty acid composition. Comp Biochem Physiol B: Biochem Mol Biol 119:325–334. doi:10.1016/S0305-0491(97)00357-X
Bruss MD, Khambatta CF, Ruby MA, Aggarwal I, Hellerstein MK (2009) Calorie restriction increases fatty acid synthesis and whole body fat oxidation rates. Am J Physiol Endocrinol Metab 298:E108–E116. doi:10.1152/ajpendo.00524.2009
Calder PC (2004) N-3 polyunsaturated fatty acids and inflammation: from molecular biology to the clinic. Lipids 38:343–352. doi:10.1007/s11745-003-1068-y
Calder PC (2007) Immunomodulation by omega-3 fatty acids. Prostaglandins Leukot Essent Fatty Acids 77:327–335. doi:10.1016/j.plefa.2007.10.015
Calder PC (2012) Mechanisms of action of (n-3) fatty acids. J Nutr 142:592S–599S. doi:10.3945/jn.111.155259
Cava E, Fontana L (2013) Will calorie restriction work in humans? Aging (Albany NY) 5:507–514
Cha MC, Jones PJ (2000) Energy restriction dilutes the changes related to dietary fat type in membrane phospholipid fatty acid composition in rats. Metabolism 49:977–983. doi:10.1053/meta.2000.7725
Chen BK, Seligman B, Farquhar JW, Goldhaber-Fiebert JD (2011) Multi-Country analysis of palm oil consumption and cardiovascular disease mortality for countries at different stages of economic development: 1980–1997. Global Health 7:45. doi:10.1186/1744-8603-7-45
Chen Y, Hagopian K, McDonald RB, Bibus D, López-Lluch G, Villalba JM, Navas P, Ramsey JJ (2012) The influence of dietary lipid composition on skeletal muscle mitochondria from mice following 1 month of calorie restriction. J Gerontol A Biol Sci Med Sci 67:1121–1131. doi:10.1093/gerona/gls113
Chen Y, Hagopian K, Bibus D, Villalba JM, López-Lluch G, Navas P, Kim K, McDonald RB, Ramsey JJ (2013) The influence of dietary lipid composition on liver mitochondria from mice following 1 month of calorie restriction. Biosci Rep 33:83–95. doi:10.1042/BSR20120060
Chen Y, Hagopian K, Bibus D, Villalba JM, López-Lluch G, Navas P, Kim K, Ramsey JJ (2014) The influence of dietary lipid composition on skeletal muscle mitochondria from mice following eight months of calorie restriction. Physiol Res 63:57–71
Chung L, Ng Y-C (2006) Age-related alterations in expression of apoptosis regulatory proteins and heat shock proteins in rat skeletal muscle. Biochim Biophys Acta 1762:103–109. doi:10.1016/j.bbadis.2005.08.003
Chung HY, Kim HJ, Kim JW, Yu BP (2001) The inflammation hypothesis of aging: molecular modulation by calorie restriction. Ann N Y Acad Sci 928:327–335. doi:10.1111/j.1749-6632.2001.tb05662.x
Civitarese AE, Carling S, Heilbronn LK, Hulver MH, Ukropcova B, Deutsch WA, Smith SR, Ravussin E, Pennington Team CALERIE (2007) Calorie restriction increases muscle mitochondrial biogenesis in healthy humans. PLoS Med 4(3):e76. doi:10.1371/journal.pmed.0040076
Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kosmatka KJ, Beasley TM, Allison DB, Cruzen C, Simmons HA, Kemnitz JW, Weindruch R (2009) Caloric restriction delays disease onset and mortality in rhesus monkeys. Science 325:201–204. doi:10.1126/science.1173635
Colman RJ, Beasley TM, Kemnitz JW, Johnson SC, Weindruch R, Anderson RM (2014) Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys. Nat Commun 5:3557. doi:10.1038/ncomms4557
Couet C, Delarue J, Ritz P, Antoine JM, Lamisse F (1997) Effect of dietary fish oil on body fat mass and basal fat oxidation in healthy adults. Int J Obes Relat Metab Disord 21:637–643. doi:10.1038/sj.ijo.0800451
Crockett EL (2008) The cold but not hard fats in ectotherms: consequences of lipid restructuring on susceptibility of biological membranes to peroxidation, a review. J Comp Physiol B 178:795–809. doi:10.1007/s00360-008-0275-7
Daum G (1985) Lipids of mitochondria. Biochim Biophys Acta 822:1–42
Deckelbaum RJ, Torrejon C (2012) The omega-3 fatty acid nutritional landscape: health benefits and sources. J Nutr 142:587S–5591S. doi:10.3945/jn.111.148080
Desai VG, Weindruch R, Hart RW, Feuers RJ (1996) Influences of age and dietary restriction on gastrocnemius electron transport system activities in mice. Arch Biochem Biophys 1333:145–151. doi:10.1006/abbi.1996.0375
Dirks A, Leeuwenburgh C (2002) Apoptosis in skeletal muscle with aging. Am J Physiol Regul Integr Comp Physiol 282:R519–R527. doi:10.1152/ajpregu.00458.2001
Dirks AJ, Leeuwenburgh C (2004) Aging and lifelong calorie restriction result in adaptations of skeletal muscle apoptosis repressors, apoptosis-inducing factor, X-linked inhibitor of apoptosis, caspase-3, and caspase-12. Free Radic Biol Med 36:27–39. doi:10.1016/j.freeradbiomed.2003.10.003
Dowhan W, Mileykovskaya E, Bogdanov M (2004) Diversity and versatility of lipid-protein interactions revealed by molecular genetic approaches. Biochim Biophys Acta 1666:19–39. doi:10.1016/j.bbamem.2004.04.010
Drew B, Phaneuf S, Dirks A, Selman C, Gredilla R, Lezza A, Barja G, Leeuwenburgh C (2003) Effects of aging and caloric restriction on mitochondrial energy production in gastrocnemius muscle and heart. Am J Physiol Regul Integr Comp Physiol 284:R474–R480. doi:10.1152/ajpregu.00455.2002
Evan G, Littlewood T (1998) A matter of life and cell death. Science 281:1317–1322. doi:10.1126/science.281.5381.1317
Faulks SC, Turner N, Else PL, Hulbert AJ (2006) Calorie restriction in mice: effects on body composition, daily activity, metabolic rate, mitochondrial reactive oxygen species production, and membrane fatty acid composition. J Gerontol A Biol Sci Med Sci 61:781–794
Flachs P, Horakova O, Brauner P, Rossmeisl M, Pecina P, Franssen-van Hal N, Ruzickova J, Sponarova J, Drahota Z, Vlcek C, Keijer J, Houstek J, Kopecky J (2005) Polyunsaturated fatty acids of marine origin upregulate mitochondrial biogenesis and induce beta-oxidation in white fat. Diabetologia 48:2365–2375. doi:10.1007/s00125-005-1944-7
Fontana L (2009) Neuroendocrine factors in the regulation of inflammation: excessive adiposity and calorie restriction. Exp Gerontol 44:41–45. doi:10.1016/j.exger.2008.04.005
Gómez J, Caro P, Naudí A, Portero-Otin M, Pamplona R, Barja G (2007) Effect of 8.5% and 25% caloric restriction on mitochondrial free radical production and oxidative stress in rat liver. Biogerontology 8:555–566. doi:10.1007/s10522-007-9099-1
Gómez A, Sánchez-Roman I, Gomez J, Cruces J, Mate I, Lopez-Torres M, Naudi A, Portero-Otin M, Pamplona R, De la Fuente M, Barja G (2014) Lifelong treatment with atenolol decreases membrane fatty acid unsaturation and oxidative stress in heart and skeletal muscle mitochondria and improves immunity and behavior, without changing mice longevity. Aging Cell 13:551–560. doi:10.1111/acel.12205
Gopinath B, Buyken AE, Flood VM, Empson M, Rochtchina E, Mitchell P (2011) Consumption of polyunsaturated fatty acids, fish, and nuts and risk of inflammatory disease mortality. Am J Clin Nutr 93:1073–1079. doi:10.3945/ajcn.110.009977
Gredilla R, Barja G (2005) Minireview: the role of oxidative stress in relation to caloric restriction and longevity. Endocrinology 146:3713–3717. doi:http://dx.doi.org/10.1210/en.2005-0378
Gredilla R, Barja G, López-Torres M (2001) Effect of short-term caloric restriction on H2O2 production and oxidative DNA damage in rat liver mitochondria and location of the free radical source. J Bioenerg Biomembr 33:279–287. doi:10.1023/A:1010603206190
Gredilla R, Phaneuf S, Selman C, Kendaiah S, Leeuwenburgh C, Barja G (2004) Short-term caloric restriction and sites of oxygen radical generation in kidney and skeletal muscle mitochondria. Ann N Y Acad Sci 1019:333–432. doi:10.1196/annals.1297.057
Hagen TM, Yowe DL, Bartholomew JC, Wehr CM, Do KL, Park JY, Ames BN (1997) Mitochondrial decay in hepatocytes from old rats: membrane potential declines, heterogeneity and oxidants increase. Proc Natl Acad Sci U S A 94:3064–3069
Hagopian K, Harper ME, Ram JJ, Humble SJ, Weindruch R, Ramsey JJ (2005) Long-term calorie restriction reduces proton leak and hydrogen peroxide production in liver mitochondria. Am J Physiol Endocrinol Metab 288:E674–E684. doi:10.1152/ajpendo.00382.2004
Hagopian K, Weber KL, Hwee DT, Van Eenennaam AL, López-Lluch G, Villalba JM, Burón I, Navas P, German JB, Watkins SM, Chen Y, Wei A, McDonald RB, Ramsey JJ (2010) Complex I-associated hydrogen peroxide production is decreased and electron transport chain enzyme activities are altered in n-3 enriched fat-1 mice. PLoS ONE 5(9):e12696. doi:10.1371/journal.pone.0012696
Halade GV, Rahman MM, Bhattacharya A, Barnes JL, Chandrasekar B, Fernandes G (2010) Docosahexaenoic acid-enriched fish oil attenuates kidney disease and prolongs median and maximal life span of autoimmune lupus-prone mice. J Immunol 184:5280–5286. doi:10.4049/jimmunol.0903282
Halminski MA, Marsh JB, Harrison EH (1991) Differential effects of fish oil, safflower oil and palm oil on fatty acid oxidation and glycerolipid synthesis in rat liver. J Nutr 121:1554–1561
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144:646–674. doi:10.1016/j.cell.2011.02.013
Harman DJ (1956) Aging: a theory based on free radical and radiation chemistry. J Gerontol 11:298–300
Harper ME, Monemdjou S, Ramsey JJ, Weindruch R (1998) Age-related increase in mitochondrial proton leak and decrease in ATP turnover reactions in mouse hepatocytes. Am J Physiol 275:E197–E206
Hepple RT, Baker DJ, McConkey M, Murynka T, Norris R (2006) Caloric restriction protects mitochondrial function with aging in skeletal and cardiac muscles. Rejuvenation Res 9:219–222. doi:10.1089/rej.2006.9.219
Herrero A, Portero-Otín M, Bellmunt MJ, Pamplona R, Barja G (2001) Effect of the degree of fatty acid unsaturation of rat heart mitochondria on their rates of H2O2 production and lipid and protein oxidative damage. Mech Ageing Dev 122:427–443. doi:10.1016/S0047-6374(01)00214-7
Hill JO, Peters JC, Lin D, Yakubu F, Greene H, Swift L (1993) Lipid accumulation and body fat distribution is influenced by type of dietary fat fed to rats. Int J Obes Relat Metab Disord 17:223–236
Hong DD, Takahashi Y, Kushiro M, Ide T (2003) Divergent effects of eicosapentaenoic and docosahexaenoic acid ethyl esters, and fish oil on hepatic fatty acid oxidation in the rat. Biochim Biophys Acta 1635:29–36. doi:10.1016/j.bbalip.2003.10.003
Huertas JR, Martinez-Velasco E, Ibáñez S, López-Frias M, Ochoa JJ, Quiles J, Parenti Castelli G, Mataix J, Lenaz G (1999) Virgin olive oil and coenzyme Q10 protect heart mitochondria from peroxidative damage during aging. BioFactors 9:337–343
Hulbert AJ (2008) The links between membrane composition, metabolic rate and lifespan. Comp Biochem Physiol A: Mol Integr Physiol 150:196–203. doi:10.1016/j.cbpa.2006.05.014
Hulbert AJ, Pamplona R, Buffenstein R, Buttemer WA (2007) Life and death: metabolic rate, membrane composition, and life span of animals. Physiol Rev 87:1175–1213. doi:10.1152/physrev.00047.2006
Ide T, Kobayashi H, Ashakumary L, Rouyer IA, Takahashi Y, Aoyama T, Hashimoto T, Mizugaki M (2000) Comparative effects of perilla and fish oils on the activity and gene expression of fatty acid oxidation enzymes in rat liver. Biochim Biophys Acta 1485:23–35. doi:10.1016/S1388-1981(00)00026-3
Infante JP, Kirwan RC, Brenna JT (2001) High levels of docosahexaenoic acid (22:6n-3)-containing phospholipids in high-frequency contraction muscles of hummingbirds and rattlesnakes. Comp Biochem Physiol B: Biochem Mol Biol 130:291–298. doi:10.1016/S1096-4959(01)00443-2
Innis SM, Clandinin MT (1981) Dynamic modulation of mitochondrial membrane physical properties and ATPase activity by diet lipid. Biochem J 198:167–175
Jolly CA, Muthukumar A, Avula CP, Troyer D, Fernandes G (2001) Life span is prolonged in food-restricted autoimmune-prone (NZB x NZW)F(1) mice fed a diet enriched with (n-3) fatty acids. J Nutr 131:2753–2760
Jones PJ (1989) Effect of fatty acid composition of dietary fat on energy balance and expenditure in hamsters. Can J Physiol Pharmacol 67:994–998
Jové M, Naudí A, Ramírez-Núñez O, Portero-Otín M, Selman C, Withers DJ, Pamplona R (2014) Caloric restriction reveals a metabolomic and lipidomic signature in liver of male mice. Aging Cell 13:828–837. doi:10.1111/acel.12241
Jurk D, Wilson C, Passos JF, Oakley F, Correia-Melo C, Greaves L, Saretzki G, Fox C, Lawless C, Anderson R, Hewitt G, Pender SL, Fullard N, Nelson G, Mann J, van de Sluis B, Mann DA, von Zglinicki T (2014) Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nat Commun 2:4172. doi:10.1038/ncomms5172
Kanzler S, Galle PR (2000) Apoptosis and the liver. Semin Cancer Biol 10:173–184. doi:10.1006/scbi.2000.0318
Khraiwesh H, López-Domínguez JA, López-Lluch G, Navas P, de Cabo R, Ramsey JJ, Villalba JM, González-Reyes JA (2013) Alterations of ultrastructural and fission/fusion markers in hepatocyte mitochondria from mice following calorie restriction with different dietary fats. J Gerontol A Biol Sci Med Sci 68:1023–1034. doi:10.1093/gerona/glt006
Khraiwesh H, López-Domínguez JA, Fernández del Río L, Gutierrez-Casado E, López-Lluch G, Navas P, de Cabo R, Ramsey JJ, Burón MI, Villalba JM, González-Reyes JA (2014) Mitochondrial ultrastructure and markers of dynamics in hepatocytes from aged, calorie restricted mice fed with different dietary fats. Exp Gerontol 56:77–88. doi:10.1016/j.exger.2014.03.023
König A, Bouzan C, Cohen JT, Connor WE, Kris-Etherton PM, Gray GM, Lawrence RS, Savitz DA, Teutsch SM (2005) A quantitative analysis of fish consumption and coronary heart disease mortality. Am J Prev Med 29:335–346. doi:10.1016/j.amepre.2005.07.001
Kromhout D, Bloemberg B, Feskens E, Menotti A, Nissinen A (2000) Saturated fat, vitamin C and smoking predict long-term population all-cause mortality rates in the Seven Countries Study. Int J Epidemiol 29:260–265. doi:10.1093/ije/29.2.260
Laganiere S, Yu BP (1989) Effect of chronic food restriction in aging rats. II. Liver cytosolic antioxidants and related enzymes. Mech Ageing Dev 48:221–230. doi:10.1016/0047-6374(89)90084-5
Laganiere S, Yu BP (1993) Modulation of membrane phospholipid fatty acid composition by age and food restriction. Gerontology 39:7–18. doi:10.1159/000213509
Lal SB, Ramsey JJ, Monemdjou S, Weindruch R, Harper ME (2001) Effects of caloric restriction on skeletal muscle mitochondrial proton leak in aging rats. J Gerontol A Biol Sci Med Sci 56:B116–B122. doi:10.1093/gerona/56.3.B116
Lambert AJ, Merry BJ (2004) Effect of caloric restriction on mitochondrial reactive oxygen species production and bioenergetics: reversal by insulin. Am J Physiol Regul Integr Comp Physiol 286:R71–R79. doi:10.1152/ajpregu.00341.2003
Lambert AJ, Merry BJ (2005) Lack of effect of caloric restriction on bioenergetics and reactive oxygen species production in intact rat hepatocytes. J Gerontol A Biol Sci Med Sci 60:175–180. doi:10.1093/gerona/60.2.175
Lands B (2014) Historical perspectives on the impact of n-3 and n-6 nutrients on health. Prog Lipid Res 55:17–29. doi:10.1016/j.plipres.2014.04.002
Lee AG (2004) How lipids affect the activities of integral membrane proteins. Biochim Biophys Acta 1666:62–87. doi:10.1016/j.bbamem.2004.05.012
León H, Shibata MC, Sivakumaran S, Dorgan M, Chatterley T, Tsuyuki RT (2008) Effect of fish oil on arrhythmias and mortality: systematic review. BMJ 23(337):a2931. doi:10.1136/bmj.a2931
Leosdottir M, Nilsson PM, Nilsson JA, Månsson H, Berglund G (2005) Dietary fat intake and early mortality patterns–data from The Malmö Diet and Cancer Study. J Intern Med 258:153–165. doi:10.1111/j.1365-2796.2005.01520.x
Li M, Izpisua-Belmonte JC (2014) Ageing: genetic rejuvenation of old muscle Nature 506: 304–305. doi: 10.1038/nature13058
López-Domínguez JA, Khraiwesh H, González-Reyes JA, López-Lluch G, Navas P, Ramsey JJ, de Cabo R, Burón MI, Villalba JM (2013) Dietary fat modifies mitochondrial and plasma membrane apoptotic signaling in skeletal muscle of calorie-restricted mice. Age (Dordr) 35:2027–2044. doi:10.1007/s11357-012-9492-9
López-Domínguez JA, Khraiwesh H, González-Reyes JA, López-Lluch G, Navas P, Ramsey JJ, de Cabo R, Burón MI, Villalba JM (2014a) Dietary fat and aging modulate apoptotic signaling in liver of calorie-restricted mice. J Gerontol A Biol Sci Med Sci doi:10.1093/gerona/glu045
López-Domínguez JA, Ramsey JJ, Tran D, Imai DM, Koehne A, Laing ST, Griffey SM, Kim K, Taylor SL, Hagopian K, Villalba JM, López-Lluch G, Navas P, McDonald RB (2014b) The influence of dietary fat source on life span in calorie restricted mice. J Gerontol A Biol Sci Med Sci doi:10.1093/gerona/glu177
López-Lluch G, Hunt N, Jones B, Zhu M, Jamieson H, Hilmer S, Cascajo MV, Allard J, Ingram DK, Navas P, de Cabo R (2006) Calorie restriction induces mitochondrial biogenesis and bioenergetic efficiency. Proc Natl Acad Sci U S A. 103:1768–1773. doi:10.1073/pnas.0510452103
López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G (2013) The hallmarks of aging. Cell 153:1194–1217. doi:10.1016/j.cell.2013.05.039
Marsh D (2008) Protein modulation of lipids, and vice versa, in membranes. Biochim Biophys Acta 1778:1545–1575. doi:10.1016/j.bbamem.2008.01.015
Marzetti E, Lawler JM, Hiona A, Manini T, Seo AY, Leeuwenburgh C (2008a) Modulation of age-induced apoptotic signaling and cellular remodeling by exercise and calorie restriction in skeletal muscle. Free Radic Biol Med 44:160–168. doi:10.1016/j.freeradbiomed.2007.05.028
Marzetti E, Wohlgemuth SE, Lees HA, Chung H-Y, Giovannini S, Leeuwenburgh C (2008b) Age-related activation of mitochondrial caspase-independent apoptotic signaling in rat gastrocnemius muscle. Mech Ageing Dev 129:542–549. doi:10.1016/j.mad.2008.05.005
Marzetti E, Carter CS, Wohlgemuth SE, Lees HA, Giovannini S, Anderson B, Quinn LS, Leeuwenburgh C (2009) Changes in IL-15 expression and death-receptor apoptotic signaling in rat gastrocnemius muscle with aging and lifelong calorie restriction. Mech Ageing Dev 130:272–280. doi:10.1016/j.mad.2008.12.008
Masoro EJ (2006) Caloric restriction and aging: controversial issues. J Gerontol A Biol Sci Med Sci 61:14–19
Masoro EJ, Iwasaki K, Gleiser CA, McMahan CA, Seo EJ, Yu BP (1989) Dietary modulation of the progression of nephropathy in aging rats: an evaluation of the importance of protein. Am J Clin Nutr 49:1217–1227
Mataix J, Ochoa JJ, Quiles JL (2006) Olive oil and mitochondrial oxidative stress. Int J Vitam Nutr Res 76:178–183
Mattison JA, Roth GS, Beasley TM, Tilmont EM, Handy AM, Herbert RL, Longo DL, Allison DB, Young JE, Bryant M, Barnard D, Ward WF, Qi W, Ingram DK, de Cabo R (2012) Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nature 489:318–321. doi:10.1038/nature11432
McMillin JB, Bick RJ, Benedict CR (1992) Influence of dietary fish oil on mitochondrial function and response to ischemia. Am J Physiol 263:H1479–H1485
Meagher EA, Fitzgerald GA (2000) Indices of lipid peroxidation in vivo: strengths and limitations. Free Radic Biol Med 28:1745–1750. doi:10.1016/S0891-5849(00)00232-X
Mujahid A, Akiba Y, Toyomizu M (2009) Olive oil-supplemented diet alleviates acute heat stress-induced mitochondrial ROS production in chicken skeletal muscle. Am J Physiol Regul Integr Comp Physiol 297:R690–R698. doi:10.1152/ajpregu.90974.2008
Murtagh-Mark CM, Reiser KM, Harris R Jr, McDonald RB (1995) Source of dietary carbohydrate affects life span of Fischer 344 rats independent of caloric restriction. J Gerontol A Biol Sci Med Sci 50:B148–B154. doi:10.1093/gerona/50A.3.B148
Nakamura MT, Yudell BE, Loor JJ (2014) Regulation of energy metabolism by long-chain fatty acids. Prog Lipid Res 53:124–144. doi:10.1016/j.plipres.2013.12.001
Nisoli E, Tonello C, Cardile A, Cozzi V, Bracale R, Tedesco L, Falcone S, Valerio A, Cantoni O, Clementi E, Moncada S, Carruba MO (2005) Calorie restriction promotes mitochondrial biogenesis by inducing the expression of eNOS. Science 310:314–317. doi:10.1126/science.1117728
Noreen EE, Sass MJ, Crowe ML, Pabon VA, Brandauer J, Averill LK (2010) Effects of supplemental fish oil on resting metabolic rate, body composition, and salivary cortisol in healthy adults. J Int Soc Sports Nutr 7:31. doi:10.1186/1550-2783-7-31
Pamplona R, Barja G (2011) An evolutionary comparative scan for longevity-related oxidative stress resistance mechanisms in homeotherms. Biogerontology 12:409–435. doi:10.1007/s10522-011-9348-1
Pamplona R, Portero-Otín M, Riba D, Ruiz C, Prat J, Bellmunt MJ, Barja G (1998) Mitochondrial membrane peroxidizability index is inversely related to maximum life span in mammals. J Lipid Res 39:1989–1994
Pamplona R, Barja G, Portero-Otín M (2002) Membrane fatty acid unsaturation, protection against oxidative stress, and maximum life span: a homeoviscous-longevity adaptation? Ann N Y Acad Sci 959:475–490. doi:10.1111/j.1749-6632.2002.tb02118.x
Pamplona R, Portero-Otín M, Sanz A, Requena J, Barja G (2004) Modification of the longevity-related degree of fatty acid unsaturation modulates oxidative damage to proteins and mitochondrial DNA in liver and brain. Exp Gerontol 39:725–733. doi:10.1016/j.exger.2004.01.006
Phillips T, Leeuwenburgh C (2005) Muscle fiber specific apoptosis and TNF-alpha signaling in sarcopenia are attenuated by life-long calorie restriction. FASEB J 19:668–670. doi:10.1096/fj.04-2870fje
Porter RK, Hulbert AJ, Brand MD (1996) Allometry of mitochondrial proton leak: influence of membrane surface area and fatty acid composition. Am J Physiol 271:R1550–R1560
Power GW, Newsholme EA (1997) Dietary fatty acids influence the activity and metabolic control of mitochondrial carnitine palmitoyltransferase I in rat heart and skeletal muscle. J Nutr 127:2142–2150
Power GW, Yaqoob P, Harvey DJ, Newsholme EA, Calder PC (1994) The effect of dietary lipid manipulation on hepatic mitochondrial phospholipid fatty acid composition and carnitine palmitoyltransferase I activity. Biochem Mol Biol Int 34:671–684
Pugh TD, Klopp RG, Weindruch R (1999) Controlling caloric consumption: protocols for rodents and rhesus monkeys. Neurobiol Aging 20:157–165. doi:10.1016/S0197-4580(99)00043-3
Quiles JL, Martínez E, Ibáñez S, Ochoa JJ, Martín Y, López-Frías M, Huertas JR, Mataix J (2002) Ageing-related tissue-specific alterations in mitochondrial composition and function are modulated by dietary fat type in the rat. J Bioenerg Biomembr 34:517–524. doi:10.1023/A:1022530512096
Ramsey JJ, Hagopian K (2006) Energy expenditure and restriction of energy intake: could energy restriction alter energy expenditure in companion animals? J Nutr 136:1958S–1966S
Ramsey JJ, Harper ME, Weindruch R (2000) Restriction of energy intake, energy expenditure, and aging. Free Radic Biol Med 29:946–968. doi:10.1016/S0891-5849(00)00417-2
Ramsey JJ, Hagopian K, Kenny TM, Koomson EK, Bevilacqua L, Weindruch R, Harper ME (2004) Proton leak and hydrogen peroxide production in liver mitochondria from energy-restricted rats. Am J Physiol Endocrinol Metab 286:E31–E40. doi:10.1152/ajpendo.00283.2003
Ramsey JJ, Harper ME, Humble SJ, Koomson EK, Ram JJ, Bevilacqua L, Hagopian K (2005) Influence of mitochondrial membrane fatty acid composition on proton leak and H2O2 production in liver. Comp Biochem Physiol B: Biochem Mol Biol 140:99–108. doi:10.1016/j.cbpc.2004.09.016
Sanchez-Roman I, Gomez J, Naudi A, Ayala V, Portero-Otín M, Lopez-Torres M, Pamplona R, Barja G (2010) The β-blocker atenolol lowers the longevity-related degree of fatty acid unsaturation, decreases protein oxidative damage, and increases extracellular signal-regulated kinase signaling in the heart of C57BL/6 mice. Rejuvenation Res 13:683–693. doi:10.1089/rej.2010.1062
Sanchez-Roman I, Gomez A, Naudí A, Jove M, Gómez J, Lopez-Torres M, Pamplona R, Barja G (2014) Independent and additive effects of atenolol and methionine restriction on lowering rat heart mitochondria oxidative stress. J Bioenerg Biomembr 46:159–172. doi:10.1007/s10863-013-9535-7
Schmitz G, Ecker J (2008) The opposing effects of n-3 and n-6 fatty acids. Prog Lipid Res 47:147–155. doi:10.1016/j.plipres.2007.12.004
Seo AY, Xu J, Servais S, Hofer T, Marzetti E, Wohlgemuth SE, Knutson MD, Chung HY, Leeuwenburgh C (2008) Mitochondrial iron accumulation with age and functional consequences. Aging Cell 7:706–716. doi:10.1111/j.1474-9726.2008.00418.x
Sohal RS, Weindruch R (1996) Oxidative stress, caloric restriction, and aging. Science 273:59–63. doi:10.1126/science.273.5271.59
Sousa-Victor P, Gutarra S, García-Prat L, Rodriguez-Ubreva J, Ortet L, Ruiz-Bonilla V, Jardí M, Ballestar E, González S, Serrano AL, Perdiguero E, Muñoz-Cánoves P (2014) Geriatric muscle stem cells switch reversible quiescence into senescence. Nature 506:316–321. doi:10.1038/nature13013
Speakman JR, Mitchell SE (2011) Caloric restriction. Mol Aspects Med 32:159–221. doi:10.1016/j.mam.2011.07.001
Spindler SR, Mote PL, Flegal JM (2014) Dietary supplementation with Lovaza and krill oil shortens the life span of long-lived F1 mice. Age 36:1345–1352. doi:10.1007/s11357-014-9659-7
Staessen L, De Bacquer D, De Henauw S, De Backer G, Van Peteghem C (1997) Relation between fat intake and mortality: an ecological analysis in Belgium. Eur J Cancer Prev 6:374–381
Su W, Jones PJ (1993) Dietary fatty acid composition influences energy accretion in rats. J Nutr 123:2109–2114
Tahin QS, Blum M, Carafoli E (1981) The fatty acid composition of subcellular membranes of rat liver, heart, and brain: diet-induced modifications. Eur J Biochem 121:5–13. doi:10.1111/j.1432-1033.1981.tb06421.x
Tsuboyama-Kasaoka N, Sano K, Shozawa C, Osaka T, Ezaki O (2008) Studies of UCP2 transgenic and knockout mice reveal that liver UCP2 is not essential for the antiobesity effects of fish oil. Am J Physiol Endocrinol Metab 294:E600–E606. doi:10.1152/ajpendo.00551.2007
Tsuduki T, Honma T, Nakagawa K, Ikeda I, Miyazawa T (2011) Long-term intake of fish oil increases oxidative stress and decreases lifespan in senescence-accelerated mice. Nutrition 27:334–337. doi:10.1016/j.nut.2010.05.017
Tucker KL, Hallfrisch J, Qiao N, Muller D, Andres R, Fleg JL, Baltimore Longitudinal Study of Aging (2005) The combination of high fruit and vegetable and low saturated fat intakes is more protective against mortality in aging men than is either alone: the Baltimore Longitudinal Study of aging. J Nutr 135:556–561
Umezawa M, Takeda T, Kogishi K, Higuchi K, Matushita T, Wang J, Chiba T, Hosokawa M (2000) Serum lipid concentrations and mean life span are modulated by dietary polyunsaturated fatty acids in the senescence-accelerated mouse. J Nutr 130:221–227
Villalba JM, Alcaín FJ (2012) Sirtuin activators and inhibitors. BioFactors 38:349–359. doi:10.1002/biof.1032
Wohlgemuth SE, Seo AY, Marzetti E, Lees HA, Leeuwenburgh C (2010) Skeletal muscle autophagy and apoptosis during aging: effects of calorie restriction and life-long exercise. Exp Gerontol 45:138–148. doi:10.1016/j.exger.2009.11.002
Yamagishi K, Iso H, Date C, Fukui M, Wakai K, Kikuchi S, Inaba Y, Tanabe N, Tamakoshi A, Japan Collaborative Cohort Study for Evaluation of Cancer Risk Study Group (2008) Fish, omega-3 polyunsaturated fatty acids, and mortality from cardiovascular diseases in a nationwide community-based cohort of Japanese men and women the JACC (Japan Collaborative Cohort Study for Evaluation of Cancer Risk) Study. J Am Coll Cardiol 52:988–996. doi:10.1016/j.jacc.2008.06.018
Yamaoka S, Urade R, Kito M (1988) Mitochondrial function in rats is affected by modification of membrane phospholipids with dietary sardine oil. J Nutr 118:290–296
Yu BP, Lim BO, Sugano M (2002) Dietary restriction downregulates free radical and lipid peroxide production: plausible mechanism for elongation of life span. J Nutr Sci Vitaminol (Tokyo) 48:257–264
Acknowledgments
Supported by NIH grant 1R01AG028125 (to JJR, PN and JMV), Ministerio de Economía y Competitividad BFU2011-23578 (to JMV), Junta de Andalucía Proyectos de Excelencia grant P09-CVI-4887 (to JMV), Junta de Andalucía Proyectos Internacionales grant (to JMV), and BIO-276 (Junta de Andalucía and the University of Córdoba, to JMV). RdC is funded by the Intramural Research program of the NIA/NIH. JALD, JA, LFdR and EGC were funded by a predoctoral fellowship of the Spanish Ministerio de Educación and BIO-276. HK was funded by a predoctoral fellowship of the Agencia Española de Cooperación Internacional al Desarrollo and BIO-276. MCR and MdR were supported by BIO-276.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Villalba, J.M., López-Domínguez, J.A., Chen, Y. et al. The influence of dietary fat source on liver and skeletal muscle mitochondrial modifications and lifespan changes in calorie-restricted mice. Biogerontology 16, 655–670 (2015). https://doi.org/10.1007/s10522-015-9572-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10522-015-9572-1