
Abstract
Brettanomyces bruxellensis negatively impacts on the sensorial quality of wine by producing phenolic compounds associated with unpleasant odors. Thus, the control of this spoilage yeast is a critical factor during the winemaking process. A recent approach used to biocontrol undesired microorganisms is the use of yeast released antimicrobial peptides (AMPs), but this strategy has been poorly applied to wine-related microorganisms. The aim of this study was to evaluate the antifungal capacity of Candida intermedia LAMAP1790 against wine-spoilage strains of B. bruxellensis and fermentative strains of Saccharomyces cerevisiae, and also to determine the chemical nature of the compound. The exposure of strains to the supernatant of C. intermedia saturated cultures showed antifungal activity against B. bruxellensis, without affecting the growth of S. cerevisiae. By fractionation and concentration of C. intermedia supernatants, it was determined that the antifungal activity was related to the presence of heat-labile peptides with molecular masses under 5 kDa. To our knowledge, this is the first report of AMPs secreted by C. intermedia that control B. bruxellensis. This could lead to the development of new biocontrol strategies against this wine-spoilage yeast.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Brettanomyces bruxellensis is a major spoilage yeast in the winemaking industry, mainly due to its ability to synthesize non-desired aromatic phenolic compounds (Oelofse et al. 2008). Several methods have been used to eradicate this microorganism from the winemaking environment, including the use of pulsed electric fields or high hydrostatic pressure to inactive the yeast (González-Arenzana et al. 2016; Puértolas et al. 2009), and treatment of contaminated barrels with hot water (Fabrizio et al. 2015). However, the high cost associated with the use of these techniques makes their implementation unviable. The use of sulphur dioxide (SO2) as a chemical preservative is the most utilized method to control B. bruxellensis, but several strains are resistant to this compound. Moreover, it was reported that this chemical could potentially be harmful to the health of consumers (Devalia et al. 1994; Vigentini et al. 2013). In this context, some research groups have investigated the use of antimicrobial peptides (AMPs) as biocontrol agents for wine-spoilage yeasts (Branco et al. 2017; Enrique et al. 2007).
AMPs are molecules that are widely distributed in nature, spanning from prokaryotes to humans and are the first defense barrier against pathogens in pluricellular organisms. In general, AMPs are short peptides with a size less than 100 amino acids and high sequence variability. However, the majority of AMPs present charged and hydrophobic amino acids at physiological pH, defining their amphipathic nature (Zhang and Gallo 2016).
With the aim of analyzing Chilean yeast wine biodiversity, Ganga and Martínez (2004) isolated several yeasts present in spontaneous fermentations of grape harvests from 1999 to 2000. From this analysis, a Candida intermedia strain (LAMAP1790), showing a high endoglucanase activity in acidic pH (5–5.5) was isolated (Ganga and Martínez 2004). In addition, our research group demonstrated the antibacterial activity of this strain against Escherichia coli, Listeria monocytogenes and Salmonella typhimurium (Acuña-Fontecilla et al. 2017). Similar observations have been reported by Younis et al. (2017), where the authors obtained three isolates of C. intermedia from raw milk and fruit yoghurt with antimicrobial activity against E. coli, Staphylococcus aureus and Pseudomonas aeruginosa (Younis et al. 2017). Despite preliminary data showing that C. intermedia LAMAP1790 is active against B. bruxellensis strains, the chemical nature of the compound responsible for this activity is unknown. In this study, we demonstrated the nature and antifungal activity of the peptides released by C. intermedia LAMAP1790 against B. bruxellensis.
Materials and methods
Strains and culture media
B. bruxellensis LAMAP1359, LAMAP2480, LAMAP3276 (AWRI1499), LAMAP3294 (CBS2499), C. intermedia LAMAP1790, Saccharomyces cerevisiae BY4741 and EC1118 (Lallemand Inc.) were used in this study. All the strains were obtained from the Laboratory of Biotechnology and Applied Microbiology culture collection of University of Santiago de Chile. Strains were grown on GYEB media (yeast extract 5 g L−1 and glucose 20 g L−1) adjusted to pH 5.0 with 100 mmol L−1 phosphate-citrate buffer (Roostita et al. 2011).
Determination of antifungal activity of C. intermedia LAMAP1790
The screening of antifungal activity from C. intermedia LAMAP1790 against B. bruxellensis and S. cerevisiae was evaluated using a methodology proposed by Mehlomakulu et al. (2014). For this, 4 × 103 cells mL−1 were inoculated on 25 mL of warm agar MBA (5 g L−1 yeast extract, 5 g L−1 peptone, 20 g L−1 glucose and 15 g L−1 agar, adjusted to pH 5.0 with 100 mmol L−1 phosphate-citrate buffer and supplemented with 0.03 g L−1 of methylene blue). After vortexing, each inoculated media was plated into petri dishes. A surface inoculation was carried out using 10 µL of 1 × 108 cells mL−1 suspension from C. intermedia LAMAP1790. As negative control, 10 µL of 1 × 108 cells mL−1 suspension from S. cerevisiae BY4741 were inoculated. The plates were incubated for 7 days at 28 °C, and every assay was evaluated six times. The antimicrobial activity was evaluated measuring the diameter of the inhibition halo in the plates (Mehlomakulu et al. 2014). In addition, a viability assay was performed using the supernatant of C. intermedia LAMAP1790 in saturated culture media. For this, 50 mL of GYEB media were inoculated with C. intermedia LAMAP1719 and it was incubated for 48 h at 28 °C. The culture was centrifuged for 10 min at 5900×g, and the supernatant was sterilized using 0.22 µm filters (EDLAB®). Then, 10 mL of filtered supernatant was inoculated with 1 × 106 cells mL−1 of each B. bruxellensis and S. cerevisiae strains and incubated for 24 h at 28 °C with 120 rpm agitation. S. cerevisiae BY4741 supernatants were prepared following the same procedure and used as a control in this experiment. A cellular count of the cultures was carried out on YPD agar plates (5 g L−1 yeast extract, 5 g L−1 peptone, 20 g L−1 glucose and 15 g L−1 agar) incubated for 7 days at 28 °C.
Characterization of antifungal compound released from C. intermedia LAMAP1790
Two fractions of 10 mL from the supernatant of C. intermedia were used to characterize the antifungal compound. One was treated for 10 min at 100 °C and the other was maintained at 4 °C. Later, each fraction was inoculated with 1 × 107 cells of B. bruxellensis LAMAP2480 and incubated for 24 h at 28 °C and with 120 rpm agitation. Counts in plates from the cultures were evaluated on YPD agar for 7 days at 28 °C. Additionally, a proteolytic treatment of the antimicrobial supernatant was assessed (Comitini et al. 2004). Five hundred milliliters of a saturated culture from C. intermedia LAMAP1790 was centrifuged for 10 min at 5900×g and filtered using membranes with 0.45 µm pore size (Pall® Corp). The proteins in the media were concentrated 100X using ultrafiltration with a 10 kDa cutoff Amicon® Ultra-15 device (Merck-Millipore®). Using the same method, the eluted fraction (minor to 10 kDa) was concentrated 100X using lyophilization. Two hundred and fifty microliters of each concentrated fraction was treated with 20 mg mL−1 of Streptomyces griseus protease (Sigma®) and incubated for 4 h at 37 °C. As control, 250 µL of each concentrated fraction (non-treated with protease) and 250 µL of sterile deionized water treated with proteases were utilized. Once the incubation was finished, the tubes were supplemented with GYEB medium at 1:1 proportion and inoculated with 1 × 105 cells of B. bruxellensis LAMAP2480. The cultures were incubated for 72 h at 28 °C, and the cell counts were assessed on YPD agar plates.
Determination of the half inhibitory concentration (IC50) and minimum inhibitory concentration (MIC) against B. bruxellensis
Using the same method described previously, a 250X concentrated fraction of peptides with molecular weight inferior to 10 kDa was prepared. The protein concentration in the fraction 250X was measured using the method described by Bradford (1976). Using the concentrated fraction, the IC50 and MIC against B. bruxellensis were evaluated as described by Branco et al. (2017). In this work, IC50 was defined as the reduction of the final yeast viability by 50% compared to control growth, while the MIC corresponds to the reduction of the final yeast viability by 99.9% in relation to the control, both measured later after 120 h of incubation.
Tris-tricine-SDS PAGE
A sample from the 250X concentrated supernatant was used to perform a Tris-tricine-SDS PAGE. The concentrated fraction was treated with 125 µg mL−1 of deoxycholic acid and 6% v v−1 trichloroacetic acid (TCA-DOC precipitation). Later, the samples were analyzed using Tris-tricine-SDS buffer and silver stained (Schägger and von Jagow 1987; Schägger 2006).
Statistical analysis
All the data were analyzed using the Kruskal–Wallis test, with an initial analysis of the distribution goodness of fit using the Kolmogorov–Smirnov test. All analysis was carried out with Statgraphics Centurion XVI Software (Statpoint Technologies Inc.). The significant differences were validated with a probability < 0.05.
Results
A screening of antifungal activity from C. intermedia LAMAP1790 was performed against wine-spoilage strains of B. bruxellensis LAMAP1359, LAMAP2480, LAMAP3276, LAMAP3294, and fermentative strain of S. cerevisiae BY4741 and EC1118. The halos observed on B. bruxellensis assays were completely clear, demonstrating that the antifungal compound produced by C. intermedia can diffuse into the agar and affect the growth of this wine-spoilage yeast. In the case of S. cerevisiae, no halo formation was observed. Additionally, a viability assay of the B. bruxellensis strains exposed to the supernatant of C. intermedia saturated culture was assessed to confirm if the antimicrobial capacity observed in the screening was related with a secreted molecule. The results showed that the antifungal compound was released to the culture media and it did not affect the laboratory or industrial wine strains of S. cerevisiae (Table 1). For B. bruxellensis LAMAP1359, LAMAP2480 y LAMAP3276, no viable cells were detected after the incubation with the antifungal supernatant, but LAMAP3294 presented a minimum viability of 468 ± 233 CFU mL−1. However, the statistical analysis did not show significant differences between the B. bruxellensis strains tested (Table 1).
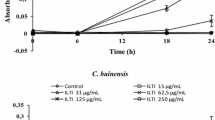
To elucidate the biochemical nature of the compound present in the C. intermedia, supernatant samples were incubated at 100 °C for 10 min, and then, the treated supernatant was used against B. bruxellensis LAMAP2480. The results showed that the supernatant exposed to temperature completely lost its antifungal activity (Fig. 1). This observation indicated that the released antifungal compound has proteinaceous nature. To confirm this hypothesis, two concentrated preparations of the supernatant were obtained by protein ultrafiltration; one larger and the other smaller than 10 kDa. Both fractions 100X were treated with protease and used against LAMAP2480. After the proteolytic treatment, it was demonstrated that the fraction 100X with peptides of molecular mass lower than 10 kDa completely lost the ability to inhibit the growth of B. bruxellensis LAMAP2480 (Fig. 2). This is indicative of the proteinic nature of the antifungal compound present in the C. intermedia fraction. Additionally, we analyzed the protein profile of the 250X fraction which contained the peptides with molecular masses under 10 kDa, using Tris-tricine-SDS PAGE. The results showed the presence of peptides with molecular masses under 4.6 kDa, and two zones with higher peptide concentration (Fig. 3, black arrows). The concentrated supernatant of S. cerevisiae BY4741 was used as a control in this experiment. The BY4741 supernatant did not present any appreciable band below 4.6 kDa, reaffirming the idea that low molecular peptides (≤ 5 kDa) are responsible for antifungal activity observed in the supernatant of C. intermedia LAMAP1790. All these results are classic characteristics of fungal AMPs, and to our knowledge, are the first report of Candida intermedia AMPs active again non-Saccharomyces yeasts.

Temperature effect over the antifungal activity from C. intermedia LAMAP1790 supernatant against B. bruxellensis LAMAP2480. The antifungal activity was observed measuring the viability of B. bruxellensis exposed to supernatants (Y-axis). GYEB medium was used as a control. The statistically significant differences are indicated with asterisk (p < 0.05)

Antifungal activity of 100X supernatant from C. intermedia LAMAP1790 against B. bruxellensis LAMAP2480, after the proteolytic treatment of the proteins fractions under and above 10 kDa. The antifungal activity of each fraction was quantified measuring the viability of B. bruxellensis after its expositions (Y-axis). Deionized water was used as a control. −P: no protease; +P: 20 mg mL−1 of Streptomyces griseus protease (Sigma®). Different letters indicate statistically significant differences (p < 0.05)

Tris-tricine-SDS PAGE of 250X protein fractions under 10 kDa obtained from S. cerevisiae BY4741 and C. intermedia LAMAP1790. The proteins bands were stained with silver nitrate. Lane ST: spectra low-range protein ladder (Thermo-Scientific®); Lane BY4741: fraction ≤ 10 kDa obtained from S. cerevisiae BY4741 supernatant; Lane LAMAP1790: fraction ≤ 10 kDa obtained from C. intermedia LAMAP1790 supernatant. Black arrows indicate low molecular weight zones with more concentration of peptides, not present in S. cerevisiae concentrated supernatant
Finally, the IC50 and MIC of the 250X protein fraction under 10 kDa obtained from C. intermedia LAMAP1790 supernatant were determined. The results show that the exposure of B. bruxellensis to 0.05 µg µL−1 of protein results in 39.8% viability, lower than the IC50 value. However, exposure to 0.1 µg µL−1 of protein results in 4.1% viability. Exposures greater than 0.15 µg µL−1 showed a complete removal of B. bruxellensis (nul viability). Using this data, we determined that the IC50 is ≤ 0.05 µg µL−1 and the MIC is 0.15 µg µL−1.
Discussion
One of the main challenges in the wine industry is reducing the losses produced by contamination with undesired yeasts. The focus of this investigation was evaluating a new method for controlling this spoilage yeast. In this context, some authors have reported alternative strategies of biocontrol (e.g., use of killer toxin). Candida pyralidae produces two killer toxins able to biocontrol several B. bruxellensis strains, with effectiveness under conditions similar to the winemaking process (Mehlomakulu et al. 2014, 2017). In this study, a similar effect of C. intermedia against B. bruxellensis was observed. Moreover, considering that the molecular mass of the C. pyralidae killer toxins is greater than 50 kDa (Mehlomakulu et al. 2017), we conclude that the antifungal peptides released by C. intermedia LAMAP1790 are not killer toxins.
In this study, to evaluate the antifungal action of C. intermedia, we selected two strains of S. cerevisiae: BY4741 derived from the most studied type strain (S288c) and EC1118 is a common commercial strain used to perform wine fermentation. Additionally, both strains present important genotypic and phenotypic variability (Liti et al. 2009; Warringer et al. 2011). In the case of B. bruxellensis, we use four strains which have been characterized by their physiological and molecular features, including genomic sequencing (Curtin et al. 2012; Piškur et al. 2012; Valdes et al. 2014). In addition, these strains represent a wide range of behaviours, such as differences in their growth rates and different conversion ratios of hydroxycinnamic acids into vinyl and ethyl derivates (Coronado et al. 2015; Godoy et al. 2016, 2017).
In all B. bruxellensis strains exposed to C. intermedia LAMAP1790, an inhibition halo was formed, suggesting the production of diffusible antifungal peptides (Table 1). This was not observed with either the laboratory strain (BY4741) or the industrial strain (EC1118) of S. cerevisiae, indicating that these species are resistant to the antifungal peptides released by C. intermedia. Some authors have described an initial screening of antimicrobial activities using the diameter of inhibition halos, because these measures are an indicator of the radial diffusion capacity of the studied compounds. However, this information corresponds to a qualitative observation (Roostita et al. 2011; Mehlomakulu et al. 2014; Acuña-Fontecilla et al. 2017; Younis et al. 2017). On the other hand, the viability evaluation was carried out exposing the four B. bruxellensis strains to the sterile supernatant. In this case, C. intermedia saturated medium did not allow the growth of three out of four B. bruxellensis strains studied. B. bruxellensis LAMAP3294 presented a minimal number of viable colonies after the treatment, whose counts do not show statistically significant differences (Table 1). These phenomenon observed between strains of B. bruxellensis could be related to the high genotypic and phenotypic variability, which would impact on the sensitivity to different antifungal peptides (Hellborg and Piškur 2009; Avramova et al. 2018). Similar effects were reported by Enrique et al. (2007) using a set of synthetic peptides derived from PAF protein of Penicillium chrysogenum and bovine lactoferricin. In that work was demonstrated the effectiveness of these peptides against B. bruxellensis, Cryptococcus albidus, Pichia membranifaciens, Zygosaccharomyces bailli and Zygosaccharomyces bosporus in culture medium. However, the antifungal activity was also detected against the fermentative yeast S. cerevisiae. In this study, the antifungal peptides do not affect the growth of the laboratory strain (BY4741) and industrial strain (EC1118) of S. cerevisiae (Table 1).
The heat treatment performed on the supernatant of C. intermedia affected its antifungal effect (Fig. 1), which led us to consider that the compound may have a protein nature. Moreover, Narayanan and Rao (1976) have described the secretion of secondary metabolites with antifungal characteristics, such as β-Indoleethanol (β-IEA) and β-Indolelacetic acid (β-ILA) in species of the genus Candida (including C. intermedia). These metabolites are synthesized from l-tryptophan and, given their physicochemical characteristics, can also be susceptible to heat treatment, mainly by evaporation (Narayanan and Rao 1976). Thus, to determine the nature of the compound, the separation of the proteins present in the supernatant by their molecular masses and the subsequent treatments of the fractions with proteases were performed. After the proteolytic treatment, it was demonstrated that the fraction 100X of peptides with molecular masses lower than 10 kDa completely lost its ability to inhibit the growth of B. bruxellensis (Fig. 2). This is indicative of the proteinic nature of the antifungal activity present in the C. intermedia LAMAP1790 supernatant. Additionally, the protein pattern of the fraction 250X lower than 10 kDa showed peptides whose molecular masses are lower than 4.6 kDa (Fig. 3). These molecular masses are consistent with reported masses for AMPs secreted by various organisms (Branco et al. 2015; Zhang and Gallo 2016). However, we are working on the sequence characterization of the peptides under 10 kDa contained in the culture supernatant of C. intermedia LAMAP1790 to confirm which of the highlighted regions in Fig. 3 are responsible for the antifungal activity.
Only a limited number of articles have reported AMPs secreted by yeast with activity against non-Saccharomyces wine-related strains. The most studied are AMPs from S. cerevisiae with activity against a variety of wine-related yeasts (Albergaria et al. 2010; Branco et al. 2014). The reported S. cerevisiae peptides are C-terminal fragments of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) isoenzymes with a molecular mass close to 1.6 kDa (Branco et al. 2014, 2017). Antifungal peptides secreted by S. cerevisiae have shown good results when controlling other yeast species such as Hanseniaspora guilliermondii, Kluyveromyces marxianus, Lachancea thermotolerans and Torulaspora delbrueckii, whose MICs vary between 250 and 500 μg mL−1 of purified peptide fraction. In the case of B. bruxellensis, the MIC obtained increased to 1000 μg mL−1, whereas the IC50 was 260 μg mL−1 (Branco et al. 2017). These values are substantially higher than MIC and IC50 obtained for the unpurified peptide fraction obtained from C. intermedia LAMAP1790. These results also demonstrate a higher antifungal effect of the C. intermedia fraction against B. bruxellensis strains. Matsuzaki (2009) reported that the MICs of different AMPs are dependent on the net charge that they present. Thus, peptides with an anionic nature have weaker antimicrobial activity (MIC > 600 μM) than peptides with cationic nature (MIC 10–100 μM). Considering that the peptides identified in S. cerevisiae are anionic, this could be an indicative of the high MIC and IC50 values necessary to affect the growth of B. bruxellensis. As mentioned previously, we are currently performing the identification of the sequences obtained in the peptides observed in Fig. 3, to carry out a parallel between their relationship with other AMPs, their three-dimensional structure and their possible degree of conservation, compared to other AMPs secreted by species of the Fungi kingdom.
One of the important factors to be defined is the effectiveness of the antifungal peptides secreted by C. intermedia LAMAP1790 as biocontrol agents of contaminating microorganisms in winemaking conditions. In this context, Branco et al. (2017) have determined that saccharomycin exhibit considerable antifungal activity on non-Saccharomyces yeasts in experiments using synthetic must. However, for the specific case of B. bruxellensis, the log growth of these experiments was reduced in 0.4 log cycles using the MIC (1000 μg mL−1) according to the authors. Therefore, this suggests that the effectiveness of saccharomycin under fermentation conditions is low and is not suitable for the control of B. bruxellensis from an industrial point of view. In this context, parallel results have shown that when a synthetic grape medium is supplemented with 1 µg of total proteins obtained from C. intermedia LAMAP1790 supernatant, the viability of B. bruxellensis and Pichia guilliermondii decrease in one log cycle (unpublished data).
In summary, it has been shown that C. intermedia LAMAP1790 produces an antifungal compound with protein nature that affects the growth of several strains of B. bruxellensis, without affecting S. cerevisiae fermentative yeast. Along with this, it was observed that the released peptides have a low molecular mass, which is in accordance with the molecular masses described for AMPs secreted by different organisms from the Fungi kingdom. Thus, this would constitute the first report of the production of AMPs secreted by C. intermedia with specific antifungal activity against B. bruxellensis.
References
Acuña-Fontecilla A, Silva-Moreno E, Ganga MA, Godoy L (2017) Evaluation of antimicrobial activity from native wine yeast against food industry pathogenic microorganisms. CYTA J Food 15(3):457–465
Albergaria H, Francisco D, Gori K, Arneborg N, Gírio F (2010) Saccharomyces cerevisiae CCMI 885 secretes peptides that inhibit the growth of some non-Saccharomyces wine-related strains. Appl Microbiol Biotechnol 86(3):965–972
Avramova M, Cibrario A, Peltier E, Coton M, Coton E et al (2018) Brettanomyces bruxellensis population survey reveals a diploid-triploid complex structured according to substrate of isolation and geographical distribution. Sci Rep 8(1):1–13
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1–2):248–254
Branco P, Francisco D, Chambon C, Hébraud M, Arneborg N et al (2014) Identification of novel GAPDH-derived antimicrobial peptides secreted by Saccharomyces cerevisiae and involved in wine microbial interactions. Appl Microbiol Biotechnol 98(2):843–853
Branco P, Viana T, Albergaria H, Arneborg N (2015) Antimicrobial peptides (AMPs) produced by Saccharomyces cerevisiae induce alterations in the intracellular pH, membrane permeability and culturability of Hanseniaspora guilliermondii cells. Int J Food Microbiol 205:112–118
Branco P, Francisco D, Monteiro M, Almeida MG, Caldeira J et al (2017) Antimicrobial properties and death-inducing mechanisms of saccharomycin, a biocide secreted by Saccharomyces cerevisiae. Appl Microbiol Biotechnol 101(1):159–171
Comitini F, De Ingeniis J, Pepe L, Mannazzu I, Ciani M (2004) Pichia anomala and Kluyveromyces wickerhamii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microbiol Lett 238(1):235–240
Coronado P, Aguilera S, Carmona L, Godoy L, Martínez C et al (2015) Comparison of the behaviour of Brettanomyces bruxellensis strain LAMAP L2480 growing in authentic and synthetic wines. Antonie Leeuwenhoek 107(5):1217–1223
Curtin CD, Borneman AR, Chambers PJ, Pretorius IS (2012) De-novo assembly and analysis of the heterozygous triploid genome of the wine spoilage yeast Dekkera bruxellensis AWRI1499. PLoS ONE 7(3):1–10
Devalia JL, Rusznak C, Herdman MJ, Trigg CJ, Davies RJ et al (1994) Effect of nitrogen dioxide and sulphur dioxide on airway response of mild asthmatic patients to allergen inhalation. Lancet 344(8938):1668–1671
Enrique M, Marcos JF, Yuste M, Martínez M, Vallés S et al (2007) Antimicrobial action of synthetic peptides towards wine spoilage yeasts. Int J Food Microbiol 118(3):318–325
Fabrizio V, Vigentini I, Parisi N, Picozzi C, Compagno C et al (2015) Heat inactivation of wine spoilage yeast Dekkera bruxellensis by hot water treatment. Lett Appl Microbiol 61(2):186–191
Ganga MA, Martínez C (2004) Effect of wine yeast monoculture practice on the biodiversity of non-Saccharomyces yeasts. J Appl Microbiol 96(1):76–83
Godoy L, Vera-Wolf P, Martinez C, Ugalde JA, Ganga MA (2016) Comparative transcriptome assembly and genome-guided profiling for Brettanomyces bruxellensis LAMAP2480 during p-coumaric acid stress. Sci Rep 6:1–13
Godoy L, Silva-Moreno E, Mardones W, Guzman D, Cubillos FA et al (2017) Genomics perspectives on metabolism, survival strategies, and biotechnological applications of Brettanomyces bruxellensis LAMAP2480. J Mol Microbiol Biotechnol 27(3):147–158
González-Arenzana L, Sevenich R, Rauh C, López R, Knorr D et al (2016) Inactivation of Brettanomyces bruxellensis by high hydrostatic pressure technology. Food Control 59:188–195
Hellborg L, Piškur J (2009) Complex nature of the genome in a wine spoilage yeast, Dekkera bruxellensis. Eukaryot Cell 8(11):1739–1749
Liti G, Carter DM, Moses AM, Warringer J, Parts L et al (2009) Population genomics of domestic and wild yeasts. Nature 458(7236):337–341
Matsuzaki K (2009) Control of cell selectivity of antimicrobial peptides. Biochim Biophys Acta 1788(8):1687–1692
Mehlomakulu NN, Setati ME, Divol B (2014) Characterization of novel killer toxins secreted by wine-related non-Saccharomyces yeasts and their action on Brettanomyces spp. Int J Food Microbiol 188:83–91
Mehlomakulu NN, Prior KJ, Setati ME, Divol B (2017) Candida pyralidae killer toxin disrupts the cell wall of Brettanomyces bruxellensis in red grape juice. J Appl Microbiol 122(3):747–758
Narayanan TK, Rao GR (1976) Beta-indoleethanol and beta-indolelactic acid production by Candida species: their antibacterial and autoantibiotic action. Antimicrob Agents Chemother 9(3):375–380
Oelofse A, Pretorius IS, du Toit M (2008) Significance of Brettanomyces and Dekkera during winemaking: a synoptic review. S Afr J Enol Vitic 29(2):128–144
Piškur J, Ling Z, Marcet-Houben M, Ishchuk OP, Aerts A et al (2012) The genome of wine yeast Dekkera bruxellensis provides a tool to explore its food-related properties. Int J Food Microbiol 157(2):202–209
Puértolas E, López N, Condón S, Raso J, Álvarez I (2009) Pulsed electric fields inactivation of wine spoilage yeast and bacteria. Int J Food Microbiol 130(1):49–55
Roostita LB, Fleet GH, Wendry SP, Apon ZM, Gemilang LU (2011) Determination of yeasts antimicrobial activity in milk and meat products. Adv J Food Sci Technol 3(6):442–445
Schägger H (2006) Tricine-SDS-PAGE. Nat Protocols 1(1):16–22
Schägger H, von Jagow G (1987) Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal Biochem 166(2):368–379
Valdes J, Tapia P, Cepeda V, Varela J, Godoy L et al (2014) Draft genome sequence and transcriptome analysis of the wine spoilage yeast Dekkera bruxellensis LAMAP2480 provides insights into genetic diversity, metabolism and survival. FEMS Microbiol Lett 361(2):104–106
Vigentini I, Lucy Joseph CM, Picozzi C, Foschino R, Bisson LF (2013) Assessment of the Brettanomyces bruxellensis metabolome during sulphur dioxide exposure. FEMS Yeast Res 13(7):597–608
Warringer J, Zörgö E, Cubillos FA, Zia A, Gjuvsland A et al (2011) Trait variation in yeast is defined by population history. PLoS Genet 7(6):e1002111. https://doi.org/10.1371/journal.pgen.1002111
Younis G, Awad A, Dawod RE, Yousef NE (2017) Antimicrobial activity of yeasts against some pathogenic bacteria. Vet World 10(8):979–983
Zhang L, Gallo RL (2016) Antimicrobial peptides. Curr Biol 26(1):R14–R19
Acknowledgements
This work has been supported by Proyecto Fortalecimiento USACH USA 1398_GM181622 Grant. Rubén Peña is funded by the Comisión Nacional de Investigación Científica y Tecnológica CONICYT-PCHA/Doctorado Nacional/2013-21130439 Doctoral Fellowship.
Author information
Authors and Affiliations
Contributions
MAG conceived and designed the study. RP performed research and analyzed data. MAG and RP wrote the paper.
Corresponding author
Ethics declarations
Conflict of interest
The authors hereby declare they do not have any conflict of interest associated to this work.
Rights and permissions
About this article

Cite this article
Peña, R., Ganga, M.A. Novel antimicrobial peptides produced by Candida intermedia LAMAP1790 active against the wine-spoilage yeast Brettanomyces bruxellensis. Antonie van Leeuwenhoek 112, 297–304 (2019). https://doi.org/10.1007/s10482-018-1159-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10482-018-1159-9