
Abstract
Diversification of monoculture grass pastures may provide opportunity for increased productivity, efficiency, and sustainability of forage-livestock systems. Our objective was to compare herbage accumulation (HA), nutritive value, and animal performance on a grass monoculture control and two alternative systems, all based on ‘Marandu’ palisadegrass [Brachiaria (syn. Urochloa) brizantha (A. Rich.) Stapf] in the Amazon Biome. Alternatives were a crop-livestock system based on soybean [Glycine max (L.) Merr.] and palisadegrass, and a eucalyptus [Eucalyptus urograndis (hybrid of E. grandis W. Hill ex Maiden and E. urophylla S. T. Blake) clone H13]-palisadegrass silvopasture. From July 2017 to May 2018 all experimental units were stocked continuously with beef cattle using a variable stocking rate to maintain a palisadegrass canopy height of 30 cm. The grass monoculture and silvopasture systems had similar HA (13,200 kg dry matter [DM] ha−1) and herbage accumulation rate (HAR; 40 kg DM ha−1 day−1), whereas crop-livestock resulted in greater HA (127%) and HAR (158%) than the other systems. Animal weight gain ha−1 for crop-livestock was 860 kg, 15% greater than for grass monoculture and silvopasture systems (average of 750 kg). Greatest palisadegrass HA and animal performance following 2 years of soybean production support the crop-livestock as an alternative for sustainable intensification. The similar HA and gain ha−1 in the seventh year of the silvopasture and grass monoculture indicates tree-pastures systems need wider arrangements to warrant long-term productivity. The maintenance of soil fertility, appropriate grazing management, and animal nutritional supplementation can greatly improve forage-based systems in the Amazon Biome.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The demand for animal protein for human consumption is expected to increase in coming years, and Brazil is an important player in supplying global demand (OECD-FAO 2015). The Brazilian livestock industry, however, faces major challenges including degraded pastures due to inadequate management of forage-based systems. In addition, practices that maximize short-term resource utilization or economic return from grass monocultures have not been effective in sustaining ecosystem function (Sollenberger et al. 2019).
Efforts have been made to intensify and diversify these systems in order to improve productivity and efficiency of forage-based cattle operations and to better conserve natural resources (Pedreira et al. 2018). The rotation of crop and livestock in a system (crop-livestock) may be one way to improve livestock production while supporting societal aspirations toward sustainable intensification. To accomplish this goal, crop-livestock systems must capitalize on the synergy among soil, plant, and animal components (Magalhães et al. 2019). Besides, the crop-livestock systems can recover degraded pastures and improve soil fertility (Salton et al. 2014), nutrient cycling, and fertilizer use efficiency (Faccio Carvalho et al. 2010).
Silvopasture systems are another potential alternative to grass monocultures. The forestry component may improve animal welfare (Domiciano et al. 2018), and carbon sequestration (Adewopo et al. 2014). However, shading reduces the photosynthetically active radiation (PAR) to the pasture, often decreasing canopy photosynthesis (Nascimento et al. 2019) and HA, while potentially increasing nutritive value (Lima et al. 2019). Exceptions can occur to the typical response of reduced forage HA in the understory under drought stress conditions when moderately shaded forage plants maintain greater leaf water potential and photosynthetic rates than full-sun plants (Johnson et al. 1994).
Overall, both systems may improve forage-based livestock production through enhancing biodiversity, resulting in more agronomically, socially, and ecologically efficient systems (Costa et al. 2018), especially in the Brazilian Amazon (Carvalho et al. 2019; Domiciano et al. 2020). Based on these factors, we hypothesized that silvopasture and crop-livestock will affect pasture responses and animal productivity. In order to test this hypothesis, we developed two integrated systems containing Brachiaria brizantha ‘Marandu’ pastures and compared HA and nutritive value, and animal performance with that obtained in grass monoculture.
Material and methods
Site characteristics and fertilization management
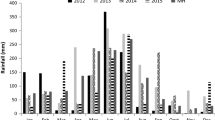
The study was carried out at Embrapa Agrossilvipastoril, in Sinop, Mato Grosso, Brazil, in the Amazon biome (11º51′ S, 55º35′ W, 370 m ASL) following the guidelines of the Committee on Ethics in Animal Use (Protocol No. 008/2015). The climate is classified according to the criteria of Köppen, as Am monsoon (Alvares et al. 2013). Annual average temperature is 25 °C and average annual relative air humidity is 70%, with 2250 mm of annual precipitation (Embrapa 2018). Weather data during the experimental period were obtained from a recording station located 500 m from the experiment site. The water balance was calculated assuming a soil water capacity of 40 cm (Fig. 1).

Daily photosynthetically active radiation (PAR, 7 a.m. to 5 p.m.), water balance and weather data from June 2017 to May 2018
The experimental area had a gently sloping topography, and the soil was a Rhodic Hapludox (Soil Survey Staff 2014), with soil texture characterized by 163 g kg–1 of coarse sand, 179 g kg–1 of fine sand, 620 g kg–1 of clay and 38 g kg–1 of silt. In June 2017, soil samples were collected at 0–20 cm for chemical analyses (Table 1). The pastures were fertilized on 18 Nov. 2017 with 29 kg P ha–1 in the form of single superphosphate. In addition, N and K were applied on 17 Jan. and 7 Mar. 2018. On each date, 50 kg N and 42 kg K were applied as urea and potassium chloride, respectively.
System description and experimental design
The systems were established in the spring of 2011 (Magalhães et al. 2019). The experimental period (17 July 2017 to 24 May 2018) was divided in two seasons: dry (17 July to 2 November 2017 and 7 April to 24 May 2018) and rainy seasons (3 November 2017 to 6 April 2018). The design was a randomized complete block with three production systems as treatments and four replications. Each experimental unit was 2 ha (200 × 100 m) in area, totaling 24 ha. The systems evaluated were Marandu palisadegrass pastures in grass monoculture, silvopasture, and crop-livestock.
For silvopasture, there were three alley cropping of trees per experimental units (Fig. 2). Each alley cropping consisted of three rows of trees, and the distance between alley cropping was 30 m. Trees were planted in Nov. 2011, in east–west rows with an intra-row spacing of 3 m and an inter-row spacing of 3.5 m, resulting in a density of 270 trees ha–1. This arrangement was selected based on previous research that demonstrated the importance of distance between alley cropping in determining the success of the silvopasture system (Gomes et al. 2019). However, in Sep. 2016, 50% of the trees were thinned to improve the understory light environment, resulting in a density of approximately 135 trees ha–1. The trees were 21 ± 2.2 m tall in July 2017. In the silvopasture, trees and pasture occupied 0.5 and 1.5 ha, respectively. Variables reported in the present study, as herbage mass, herbage accumulation, stocking rate and gain per hectare were calculated based on the effective pasture area, excluding the area occupied by trees (Magalhães et al. 2019).

Schematic representation and evaluation sites in the silvopasture (a) and grass monoculture or crop-livestock systems (b)
In the crop-livestock, the crop and livestock components rotate every 2 years. From Nov. 2011 to Jun. 2013, soybean was cultivated as the first crop (Oct.-Feb.) followed by maize (Zea mays L.) intercropped with palisadegrass as the second crop, and palisadegrass without grazing was grown from Jul. 2013 to Oct. 2015. The current evaluation was conducted in the first livestock period (Jul. 2017-May 2018) after two crop seasons (Oct. 2015-Feb. 2017). In these two previous crop seasons, soybean was grown as the first crop (Oct. 2015-Feb. 2016) followed by palisadegrass established in late Feb. In second crop season (Oct. 2016-Feb. 2017), pasture was deferred (Mar.-Apr. 2017) to ensure the establishment, followed by grazing.
The presence of spittlebug (Deois flavopicta Stall; Notozulia entreriana Berg, and Mahanarva spp.) was monitored and chemical pest control was used once in silvopasture (15 Nov. 2017) and twice in the grass monoculture (15 and 31 Nov. 2017), applying 300 mL ha−1 of Thiamethoxam (141 g L−1) + Lambda-cyhalothrin (106 g L−1). Biological pest control was applied on 13 Jan. and 7 Mar. 2018 using 0.5 kg ha−1 of Metarhizium anisopliae (Metschn.) Sorokin in Ssilvopasture and crop-livestock systems.
Pasture assessments
Pastures were continuously stocked using a variable stocking rate (Mott and Lucas 1952), and average pasture canopy height was targeted for 30 ± 5.0 cm (Da Silva et al. 2014). The canopy height was monitored weekly at 50 points per experimental units (Fig. 2) using ruler and transparency (Barthram 1985). In the grass monoculture and crop-livestock, measurements were taken at randomly selected points. In the silvopasture, a zigzag pattern was used to represent the range of possible distances from the alley cropping. Twenty-five points were sampled on each face (north and south) of the central alley cropping of trees. The canopy height was 30, 28 and 29 cm, on average, for grass monoculture, silvopasture and crop-livestock throughout the experimental period.
Photosynthetically active radiation (PAR) was monitored monthly throughout the day (7 a.m. to 5 p.m.) using quantum linear sensors (PQS1, Kipp & Zonen BV, Netherlands) connected to an automated data acquisition system (CR3000, Campbell Scientific, UK) in one experimental unit of each system. In silvopasture, four recording stations were placed at 7.5 and 15 m from the tree trunks of the edge row, on the north and south face of the central alley cropping of trees. In the grass monoculture and crop-livestock, one recording station was placed at the center of the experimental units and PAR was determined based on the average of both. In the grass monoculture and crop-livestock, the daily PAR was 802 μmol photons m–2 s–1 and in the silvopasture was 652 μmol photons m–2 s–1.
Herbage accumulation (HA) and herbage mass (HM) were quantified every 28-days and total herbage accumulation (THA) was quantified by summing the HA at each cycle 28-day. During winter, the frequency of measurement was 56 days because the herbage accumulation rate (HAR) was slowed. The HA was measured using the paired cage technique (Klingman et al. 1943), and HAR was obtained by dividing the HA by number of days between sampling events.
In the grass monoculture and crop-livestock, exclosure cages (0.64 m2 in area and 1.2 m tall) were placed at four sites per experimental units. In silvopasture, four cages were placed on the north face and four on the south face of the central alley cropping of trees, with two cages each at 7.5 and 15 m from the tree trunks of the edge row (Fig. 2).
At each cage site, paired sites were chosen with similar canopy height. A cage was placed at one of the paired sites and at the other the HM was measured by clipping all herbage to soil level inside a circular quadrat (0.64 m2). On the next sampling date, HM inside the cage was quantified by clipping the herbage inside the quadrat to soil level and drying it at 55 °C in a forced-air dryer until constant weight. New paired sampling sites were chosen on that same day to start the next accumulation period. The HA was calculated as the difference between HM in the cage and HM measured at the paired site on the day that cages were allocated earlier. Herbage bulk density was calculated as the quotient of HM and the canopy height at the time of measurement.
In order to evaluate plant-part composition, subsamples of HM were taken in four cycles, two during the dry season (Aug. and Oct. 2017) and two cycles in the rainy season (Jan. and Mar. 2018). In each of these four cycles, HM sub-samples (described previously) were separated into leaf, stem plus sheath, and dead material. The samples were dried at 55 °C in a forced-air dryer until constant weight and plant-part proportion calculated.
In each experimental units, in the same four cycles, 50 sites that represented mean canopy condition were selected and hand-plucked samples taken (Vendramini et al. 2007), separated (leaf, stem plus sheath, and dead material), and dried in an air-forced oven at 55 °C to constant weight. Subsequently, plant parts were combined and total hand-plucked herbage samples were ground to pass a 1-mm screen to determine concentrations of dry matter (DM), crude protein (AOAC 1990), neutral detergent fiber (NDF) and acid detergent fiber (ADF; Van Soest et al. 1991).
Animal performance
Animal performance was evaluated with Nellore (Bos taurus indicus L.) steers with average initial body weight (BW) of 338 ± 10 kg. Animals were classified as either testers or grazers. Three tester animals were allocated to each experimental unit based on weight and age. The grazer animals were moved on and off pastures as needed to adjust the stocking rate (SR) so that canopy height target was kept constant. Throughout the evaluation period, supplement (2 g kg–1 BW) was offered to the animals. The supplement (corn, soybean meal and probiotics) had 300 g kg–1 crude protein (135 g non-protein nitrogen kg–1, urea) during the dry season and 200 g kg–1 crude protein during the rainy season. The animals were weighed every 28 days after a 16-h feed and water fast.
The SR was calculated as the quotient of the average weight of the animals (testers and grazers) and the number of days in the experimental units (Petersen and Lucas 1968). The minimum (SRmin) and maximum (SRmax) stocking rates were reported by season. The herbage allowance (HAL) was calculated for each system as the average HM in each 28-d period divided by the total average steer BW during that period (Sollenberger et al. 2005). Only testers were used to calculate the average daily gain (ADG). The average body weight gain per area (gain ha−1) was obtained by multiplying the ADG of the tester animals by the total number of animals days supported in that experimental units (Petersen and Lucas 1968) and total gain ha–1 was quantified by summing the gain ha–1 in each cycle.
Statistical analysis
The data were analyzed using mixed models methods with parametric structure in the covariance matrix, through the MIXED procedure (SAS Studio, v. 9.4) (Littell et al. 2006) using the maximum likelihood restricted method (REML). The ‘seasonal’ response variables were analyzed as repeated measurements in the seasons (rainy and dry). Block and the block × system interaction were considered as random effects, whereas system, season and season × system interaction were fixed effects. Linear predictor and quantil-quantile plots of the residues were used to verify homogeneity of variance and error normality. The command “TYPE” specifies the covariance matrix structure, which was selected considering the smallest Akaike information criterion (AIC) (Wolfinger 1993). The means were estimated by least squares mean (LSMEANS) and comparisons were performed using the Tukey–Kramer test (P < 0.05).
Results
Pasture productivity
The THA was affected by systems, where crop-livestock was 127% greater than grass monoculture and silvopasture. There was a system × season interaction for HM, HA, HAR, herbage bulk density and for leaf, stem and dead material proportions (Table 2). Across seasons, crop-livestock had the greatest HM, HA, HAR and herbage bulk density. The HM was 40% and 77% greater in crop-livestock than in grass monoculture and silvopasture, respectively, in the dry season. In addition, during the rainy season, HM was 131% greater in crop-livestock than in grass monoculture and silvopasture. The crop-livestock had the same HM in both seasons.
The HA in crop-livestock was 74% and 228% greater than the average of grass monoculture and silvopasture in the dry and rainy seasons, respectively (Table 2). The silvopasture maintained similar HA across the seasons, while grass monoculture and crop-livestock had 60% and 110% greater HA, respectively, in rainy than dry seasons. The HAR in crop-livestock was 146 and 174% greater than the average of grass monoculture and silvopasture during dry and rainy seasons, respectively. The HAR was, on average, 55% less in dry than rainy seasons, across systems. The crop-livestock had greater herbage bulk density than grass monoculture and silvopasture, with an increase of 60% and 116% in the dry and rainy seasons, respectively.
Leaf and stem proportions in HM were greatest in the crop-livestock, which did not differ among seasons (Table 2). However, in grass monoculture and silvopasture, the leaf and stem proportions in HM were greatest and the dead material proportion least in the rainy season.
Hand-plucked plant-part composition and nutritive value
Leaf, stem and dead material proportions were affected by a system × season interaction (Table 3). The greatest leaf proportion and the least dead material proportion were measured during the rainy season. The silvopasture had the least leaf proportion regardless of the season. In addition, during the dry season, crop-livestock had a greater leaf proportion and lesser dead material proportion than grass monoculture and silvopasture.
The NDF and crude protein concentrations were affected by a system × season interaction (Table 3). In the rainy season, NDF concentration did not differ among systems, however, grass monoculture and silvopasture were the greatest during the dry season. Crude protein was greater in the rainy than the dry season for all systems. The greatest dry season crude protein concentration was measured in crop-livestock. In the rainy season, crude protein was also greater for crop-livestock than grass monoculture but crop-livestock and silvopasture did not differ during the rainy season. Season also affected the ADF concentration, with greater values during the dry (330 g kg−1 DM) than rainy (315 g kg−1 DM) season.
Animal responses
The HAL and SR were affected by a system × season interaction (Table 4). The greatest HAL was recorded for all systems during the dry season. Within seasons, HAL was least for silvopasture in the dry season and for grass monoculture and silvopasture during the rainy season. However, the greatest average, minimum and maximum SR occurred in the rainy season for all systems. In the dry season, the average SR was 30% greater for crop-livestock than grass monoculture, while in the rainy season average SR in crop-livestock was 24% greater than grass monoculture and silvopasture.
The SRmin was affected by system and season (Table 4), and it was 43% greater for crop-livestock than grass monoculture and silvopasture. In addition, SRmin was 84% greater during the rainy than dry season. The SRmax, ADG and gain ha−1 were affected by season (Table 4), where these responses were 47, 21 and 104% greater during the rainy than dry season, respectively. In addition, the total gain ha–1 was affected by systems, with the crop-livestock presenting 15% greater gain than the grass monoculture and silvopasture.
Discussion
Measures of herbage quantity and nutritive value were generally superior in the rainy vs. the dry season (Table 2) and were associated with the greater rainfall (Fig. 1) and fertilizer application during that season, both of which support enhanced plant growth and often impact nutritive value (Bourscheidt et al. 2019). Although HA in the dry season was expected to be only 20–30% that observed in the rainy season (Castro Santos et al. 2016), the grass monoculture and silvopasture contributed about 40% of the THA during the dry season.
The greatest responses of HM, HA, HAL, herbage bulk density and animal responses were observed in the crop-livestock both across and within seasons illustrates the impact of 2 years of crop production management, in particular fertilization, on subsequent forage production (Costa et al. 2015). They may increase soil organic matter (Costa et al. 2015), base saturation and cation exchange capacity (Salton et al. 2014), further benefiting forage growth potential after a cropping period. In contrast, the grass monoculture and silvopasture generally underperformed crop-livestock likely due to lesser nutrient input in the previous years and absence of crop residue addition. In our study, the major difference in soil chemical characteristics among systems was soil K which was four times greater for crop-livestock than for grass monoculture and silvopasture (Table 1). However, soil P was two times greater in crop-livestock than grass monoculture, and the sum of bases, cation exchange capacity, and base saturation were 8–38% greater in crop-livestock than grass monoculture.
Lesser HM in silvopasture than crop-livestock could be a result of the 23% less PAR incidence (Nascimento et al. 2019) and the effect of light quality (red: far-red) that affects tillering (Peri et al. 2007). The light absorption by the forest canopy component reduces the incident PAR on the pasture canopy understory and can negatively affect HA (Gomes et al. 2020), although in the current experiment there was no difference in THA among grass monoculture and silvopasture. Moderately shaded pasture plants can maintain greater leaf water potential and photosynthetic rates under drought conditions than full-sun plants (Johnson et al. 1994). Marandu palisadegrass can also show physiological adaptations (e.g., maintain leaf photosynthesis, increase specific leaf area and chlorophyll indexes) to support HA in a silvopasture (Nascimento et al. 2019). Under moderate shade (reduced PAR), plants were found to acclimate, maximizing radiation use efficiency and meeting the demand for maintenance and production of new tissues (Valladares and Niinemets 2008).
The increase in the concentration of fiber components is associated with increasing plant maturity due to accumulation of cellulose, hemicellulose and lignin (Minson 1990). Grazing management guided by a steady state canopy height, however, could reduce variation in canopy structure and composition, regardless of the season, and contribute to maintaining a vegetative state (Pedreira et al. 2017) and more constant nutritive value (Nave et al. 2011). Lack of seasonal differences in NDF for crop-livestock occurred because leaf proportion was sustained during the dry season to a much greater degree than the other systems.
In tropical grass pastures, there is a concern about crude protein concentration, which could limit activity of ruminal microorganisms and reduce fibrous substrate digestibility and rate of passage (Lazzarini et al. 2009). As a general guide, the forage crude protein concentration for grazing beef cattle is recommended to be above 70 g kg−1 DM (Minson 1990), although all systems in the current study provided levels greater than that required by many categories of livestock. In all systems, animals were supplemented throughout the year to improve animal performance and the production potential (Vendramini et al. 2007).
From a systems perspective, is also important to have an adequate HAL, which is a determinant factor of forage intake and performance of grazing animals (Sollenberger and Vanzant 2011). In this experiment, HAL was a response to the canopy target height imposed and thus it varied seasonally and among treatments due to differences in HA and herbage bulk density (Table 4). Greater HA during the rainy season required an increase in SR to maintain the target canopy height and resulted in lesser HAL during the rainy than dry season. Greater herbage bulk density and HM for crop-livestock at the same canopy height than the other systems contributed to generally greater HAL in the crop-livestock. Levels of approximately 3.5 kg DM kg−1 BW have been suggested for palisadegrass (Herling et al. 2011) as contributing to optimal animal intake in pasture-based systems. Studies with other C4 grasses have shown increasing ADG with increasing HAL up to 3.3 kg DM kg–1 BW for pearlmillet (Pennisetum glaucum L.[R. Br.]) (McCartor and Rouquette 1977) and up to ~ 4.0 for stargrass (Cynodon nlemfuensis Vanderyst) (Hernández Garay et al. 2004). Thus, HAL may not have been sufficiently great to maximize ADG in all treatments and during both seasons in the current study, but the addition of supplement likely reduced any negative impacts of HAL on ADG.
The ADG did not differ among systems, but ADG was greater in the rainy than in the dry season for all systems (Table 4). This is likely associated with greater leaf proportion and lesser dead material proportion, lesser NDF and ADF during the rainy season. It is important to note, however, the ADG in the dry season was 0.61 kg day−1, which was only 21% less than the rainy season ADG. Dry season ADG in the current study was greater than those reported (~ 0.300 kg day−1) in several studies (Paciullo et al. 2011; Euclides et al. 2016; Domiciano et al. 2018), likely due to the shorter dry season and the supplementation in our study.
Greater HA in the crop-livestock was associated with the greatest SR among systems. This increased HA and SR are due to the synergistic effects among crops and livestock, resulting in better soil characteristics for crop-livestock. Considering the similar ADG among systems, gain ha−1 was mainly driven by differences in SR. The SRmax and SRmin values illustrate the relative herbage production difference between the rainy and dry seasons, and the need to develop strategic feeding to account for differences among systems. For example, if a system allows greater SR during the rainy season, a greater supplemental fodder source (e.g. stockpiled forage, silage, hay) will be required during the dry season to accommodate the needs of this larger herd. In addition, varying feed supplementation strategies (e.g. feedlots or high concentrate supplement) are likely needed seasonally to maintain high levels of animal performance throughout the year and reduce time to animal harvest (Pedreira et al. 2018).
Overall, compared with grass monoculture, the crop-livestock offers greater animal and pasture productivity, increased production diversification, reduced market risks and the ability to minimize economic instability (Reis et al. 2018). However, the grass monoculture and silvopasture supported levels of production that far exceed averages for the current Brazilian livestock scenario (750 vs. 167 kg of liveweight total gain ha−1; Zen et al. 2018). It is important to note that these results occurred in the seventh year of imposing these systems and when considering the total area (pasture more forestry) in silvopasture, the system productivity is increased by 25%, providing evidence of system sustainability. Going forward, more research is needed to address the impacts of establishment patterns and densities on shading of the forage component and its relationship to herbage and animal production, and the effect of shading on thermal comfort indices of animals in pasture-livestock systems. As the systems continue to be imposed and trees are harvested, the forestry component adds a new income source, which can be commercialized or used on-farm (fences, corral, wooden gates, etc.). Finally, our study highlights the potential of traditional (grass monoculture) and alternative (silvopasture and crop-livestock) forage-based production systems in the Brazilian Amazon Biome when recognized technologies (maintenance of soil fertility and appropriate grazing management associated with nutritional supplementation) are properly applied.
Conclusions
-
1.
The increase in herbage accumulation of palisadegrass and animal performance after 2 years of soybean production support the crop-livestock system as an option for sustainable intensification.
-
2.
The similar pasture and animal productivity in the seventh year of the silvopasture and grass monoculture systems suggests that adequate tree arrangement ensures long-term pasture productivity, besides several trees benefits.
-
3.
The maintenance of soil fertility, appropriate grazing management, and nutritional supplementation can greatly improve forage-based systems in the Amazon Biome.
References
Adewopo JB, Silveira ML, Xu S et al (2014) Management intensification impacts on soil and ecosystem carbon stocks in subtropical grasslands. Soil Sci Soc Am J 78:977–986. https://doi.org/10.2136/sssaj2013.12.0523
Alvares CA, Stape JL, Sentelhas PC et al (2013) Köppen’s climate classification map for Brazil. Meteorol Zeitschrift 22:711–728. https://doi.org/10.1127/0941-2948/2013/0507
AOAC (1990) Official methods of analysis, 15th edn. Association of Official Analytical Chemists, Virginia
Barthram GT (1985) Experimental techniques: the HFRO sward stick. Hill Farming Res Organ Rep 1984/1985, pp 29–30
Bourscheidt MLB, Pedreira BC, Pereira DH et al (2019) Nitrogen input strategies in pastures: mineral fertilizer, bacterial inoculant and consortium with forage peanuts. Sci Electron Arch 12:137–147. https://doi.org/10.36560/1232019784
Carvalho PCF, Anghinoni I, de Moraes A et al (2010) Managing grazing animals to achieve nutrient cycling and soil improvement in no-till integrated systems. Nutr Cycl Agroecosystems 88:259–273. https://doi.org/10.1007/s10705-010-9360-x
Carvalho P, Domiciano LF, Mombach MA et al (2019) Forage and animal production on palisadegrass pastures growing in monoculture or as a component of integrated crop–livestock–forestry systems. Grass Forage Sci 74:650–660. https://doi.org/10.1111/gfs.12448
Costa MP, Schoeneboom JC, Oliveira SA et al (2018) A socio-eco-efficiency analysis of integrated and non-integrated crop-livestock-forestry systems in the Brazilian Cerrado based on LCA. J Clean Prod 171:1460–1471. https://doi.org/10.1016/j.jclepro.2017.10.063
Costa NR, Andreotti M, Mascarenhas Lopes KS et al (2015) Atributos do solo e acúmulo de carbono na integração lavoura-pecuária em sistema plantio direto. Rev Bras Cienc do Solo 39:852–863. https://doi.org/10.1590/01000683rbcs20140269
Da Silva SC, Pereira LET, Sbrissia AF, Hernandez-Garay A (2014) Carbon and nitrogen reserves in marandu palisade grass subjected to intensities of continuous stocking management. J Agric Sci 153:1449–1463. https://doi.org/10.1017/S0021859614001130
Domiciano LF, Mombach MA, Carvalho P et al (2018) Performance and behaviour of Nellore steers on integrated systems. Anim Prod Sci 58:920. https://doi.org/10.1071/AN16351
Domiciano LF, Pedreira BC, Silva NMF da, et al (2020) Agroforestry systems: an alternative to intensify forage-based livestock in the Brazilian Amazon. Agrofor Syst X:1–11. doi: 10.1007/s10457–020–00499–1
Embrapa (2018) Weather Station (Estação Meteorológica). In: Embrapa Agrossilvipastoril
Euclides VPB, Montagner DB, Barbosa RA et al (2016) Animal performance and sward characteristics of two cultivars of Brachiaria brizantha (BRS Paiaguás and BRS Piatã). Rev Bras Zootec 45:85–92. https://doi.org/10.1590/S1806-92902016000300001
Gomes FJ, Pedreira BC, Santos PM et al (2020) Microclimate effects on canopy characteristics of shaded palisadegrass pastures in a silvopastoral system in the Amazon biome of central Brazil. Eur J Agron 115:126029. https://doi.org/10.1016/j.eja.2020.126029
Gomes FJ, Pedreira CGS, Bosi C et al (2019) Shading effects on Marandu palisadegrass in a silvopastoral system: plant morphological and physiological responses. Agron J 111:1–9. https://doi.org/10.2134/agronj2019.01.0052
Herling VR, Pedreira CGS, De PH et al (2011) Performance and productivity of Nellore steers on rotationally stocked palisadegrass (Brachiaria brizantha) pastures in response to herbage allowance. J Agric Sci 149:761–768. https://doi.org/10.1017/S0021859611000116
Hernández Garay A, Sollenberger LE, McDonald DC et al (2004) Nitrogen fertilization and stocking rate affect stargrass pasture and cattle performance. Crop Sci 44:1348–1354. https://doi.org/10.2135/cropsci2004.1348
Johnson SE, Sollenberger LE, Bennett JM (1994) Yield and reserve status of rhizoma peanut growing under shade. Crop Sci 34:757–761. https://doi.org/10.2135/cropsci1994.0011183X003400030028x
Klingman DL, Miles SR, Mott GO (1943) The cage method for determining consumption and yield of pasture herbage. J Am Soc Agron 35:739–746
Lazzarini I, Detmann E, Sampaio CB et al (2009) Transit and degradation dynamics of neutral detergent fiber in cattle fed low-quality tropical forage and nitrogenous compounds (Portuguese). Arq Bras Med Veterinária e Zootec 61:635–647. https://doi.org/10.1590/S0102-09352009000300017
Lima MA, Paciullo DSC, Morenz MJF et al (2019) Productivity and nutritive value of Brachiaria decumbens and performance of dairy heifers in a long-term silvopastoral system. Grass Forage Sci 74:160–170. https://doi.org/10.1111/gfs.12395
Littell RC, Milliken GA, Stroup WW et al (2006) SAS for mixed model, 2nd edn. SAS Publishing, Cary
Magalhães CAS, Pedreira BC, Tonini H, Farias Neto AL (2019) Crop, livestock and forestry performance assessment under different production systems in the north of Mato Grosso, Brazil. Agrofor Syst 93:2085–2096. https://doi.org/10.1007/s10457-018-0311-x
McCartor MM, Rouquette FM (1977) Grazing pressures and animal performance from pearl millet. Agron J 69:983. https://doi.org/10.2134/agronj1977.00021962006900060020x
Minson DJ (1990) Forage in ruminant nutrition. Academic Press, San Diego
Mott GO, Lucas HL (1952) The design, conduct and interpretation of grazing trials on cultivated and improved pastures. In: IGC, Proceedings of the 6th International Grassland Congress. State College Press, Pennsylvania, pp 1380–1395
Nascimento HLB, Pedreira BC, Sollenberger LE et al (2019) Physiological characteristics and forage accumulation of grazed Marandu palisade grass (Brachiaria brizantha) growing in monoculture and in silvopasture with Eucalyptus urograndis. Crop Pasture Sci 70:384–394. https://doi.org/10.1071/CP18403
Nave RLG, Pedreira CGS, Pedreira BC (2011) Nutritive value and physical characteristics of Xaraes palisadegrass as affected by grazing strategy. S Afr J Anim Sci 40:285–293. https://doi.org/10.4314/sajas.v40i4.65236
OECD-FAO (2015) OCDE-FAO Perspectivas Agrícolas 2015–2024 (OECD-FAO Agricultural Outlook 2015–2024). OECD Publishing, Paris
Paciullo DSC, Castro CRT, Gomide CAM et al (2011) Performance of dairy heifers in a silvopastoral system. Livest Sci 141:166–172. https://doi.org/10.1016/j.livsci.2011.05.012
Pedreira BC, Domiciano LF, Vilela L et al (2018) O estado da arte e estudos de caso em sistemas integrados de produção agropecuária no Centro-Oeste do Brasil. In: de Souza ED, da Silva FD, Assmann TS, et al. (eds) Sistemas Integrados de Produção Agropecuária no Brasil, 1st edn. Copiart, Tubarão, pp 277–300
Pedreira CGS, Silva VJ, Pedreira BC, Sollenberger LE (2017) Herbage accumulation and organic reserves of palisadegrass in response to grazing management based on canopy targets. Crop Sci 57:2283. https://doi.org/10.2135/cropsci2016.11.0957
Peri PL, Lucas RJ, Moot DJ (2007) Dry matter production, morphology and nutritive value of Dactylis glomerata growing under different light regimes. Agrofor Syst 70:63–79. https://doi.org/10.1007/s10457-007-9029-x
Petersen RG, Lucas HL (1968) Computing methods for the evaluation of pastures by means of animal response. Agron J 60:682–687. https://doi.org/10.2134/agronj1968.00021962006000060031x
Reis JC dos, Kamoi MYT, Michetti M, Wruck FJ (2018) Análise dos benefícios econômicos da diversificação da produção em sistemas de integração lavoura-pecuáriafloresta. In: 56° Congresso da Sociedade Brasileira de Economia, Administração e Sociologia Rural - SOBER. Congresso da Sociedade Brasileira de Economia, Administração e Sociologia Rural, Campinas, SP
Salton JC, Mercante FM, Tomazi M et al (2014) Integrated crop-livestock system in tropical Brazil: toward a sustainable production system. Agric Ecosyst Environ 190:70–79. https://doi.org/10.1016/j.agee.2013.09.023
Santos DC, Guimarães Júnior R, Vilela L et al (2016) Forage dry mass accumulation and structural characteristics of Piatã grass in silvopastoral systems in the Brazilian savannah. Agric Ecosyst Environ 233:16–24. https://doi.org/10.1016/j.agee.2016.08.026
Soil Survey Staff (2014) Keys to soil taxonomy by soil, 12th edn. USDA - Natural Resources Conservation Service, Washington
Sollenberger LE, Kohmann MM, Dubeux JCB, Silveira ML (2019) Grassland management affects delivery of regulating and supporting ecosystem services. Crop Sci 59:1–19. https://doi.org/10.2135/cropsci2018.09.0594
Sollenberger LE, Moore JE, Allen VG, Pedreira CGS (2005) Reporting forage allowance in grazing experiments. Crop Sci 45:896–900. https://doi.org/10.2135/cropsci2004.0216
Sollenberger LE, Vanzant ES (2011) Interrelationships among forage nutritive value and quantity and individual animal performance. Crop Sci 51:420–432. https://doi.org/10.2135/cropsci2010.07.0408
Valladares F, Niinemets Ü (2008) Shade tolerance, a key plant feature of complex nature and consequences. Annu Rev Ecol Evol Syst 39:237–257. https://doi.org/10.1146/annurev.ecolsys.39.110707.173506
Van Soest PJ, Robertson JB, Lewis BA (1991) Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 74:3583–3597. https://doi.org/10.3168/jds.s0022-0302(91)78551-2
Vendramini JMB, Sollenberger LE, Dubeux JCB et al (2007) Concentrate supplementation effects on the performance of early weaned calves grazing Tifton 85 bermudagrass. Agron J 99:399–404. https://doi.org/10.2134/agronj2005.0355
Wolfinger R (1993) Covariance structure selection in general mixed models. Commun Stat Simul Comput 22:1079–1106. https://doi.org/10.1080/03610919308813143
Zen S de, Moreira R, Gomes M, Penazzi G (2018) Em 10 anos, produtividade média da pecuária nacional cresce mais de 22%. Bol Ativos da Pecuária Corte 10:1–5. https://www.cnabrasil.org.br/boletins
Acknowledgements
This work was supported by CNPq [479409/2013-7 and 303438/2015-0]; Embrapa [Grant Numbers 02.16.05.007.00.00]; Acrimat and ICLF Network. The authors acknowledge CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil) [Finance Code 001] and FAPEMAT for granting a scholarship to the first author; and Acrinorte for the partnership with the beef cattle used [44.771/14 and 50.168/2017].
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
da Silva, F.S., Domiciano, L.F., Gomes, F.J. et al. Herbage accumulation, nutritive value and beef cattle production on marandu palisadegrass pastures in integrated systems. Agroforest Syst 94, 1891–1902 (2020). https://doi.org/10.1007/s10457-020-00508-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-020-00508-3